
Testis-Specific Isoform of Na+-K+ ATPase and Regulation of Bull Fertility

{kind=link}
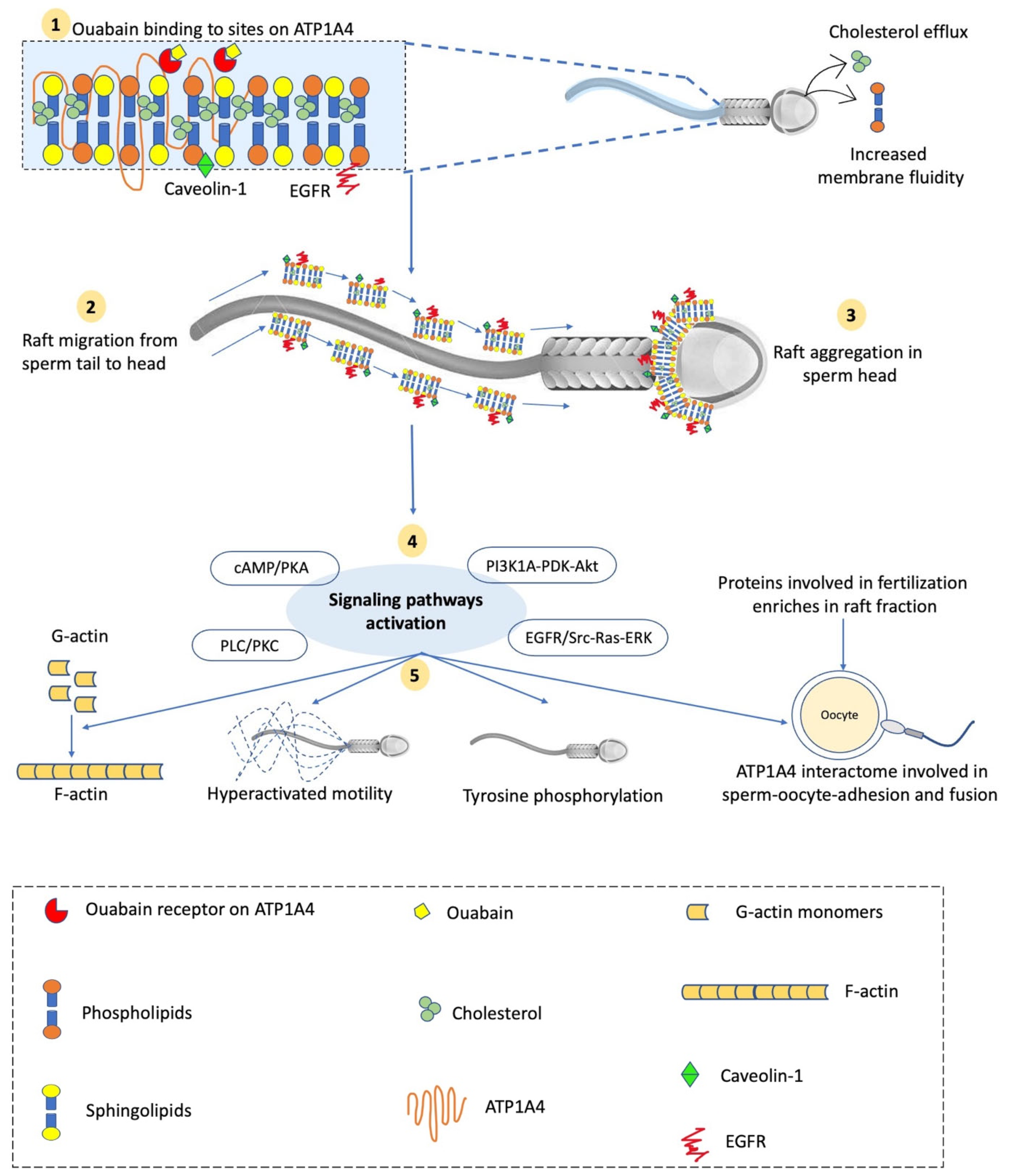
Abstract
1. Introduction
2. Na+-K+ ATPase (NKA) Ion Channel
3. NKA in Somatic Cells
4. Distribution of ATP1A4 in the Testis and Sperm
5. Role of NKA α4 Isoform in Sperm Physiology
5.1. ATP1A4-Associated Signaling Pathways Involved in Bull Sperm Capacitation
5.2. ATP1A4 Interactome in Sperm Raft and Non-Raft Fractions during Capacitation
5.3. ATP1A4 Function in Sperm Motility and Capacitation-Associated Hyperactivation
5.4. AT1A4 in Sperm–Oocyte Interaction and Activation
6. De Novo ATP1A4 Translation during Capacitation
7. ATP1A4 as a Potential Candidate Biomarker in Male Fertility Prediction
8. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ATP1A4 | Na+-K+ ATPase α4 isoform |
| ADAM32 | ADAM metallopeptidase domain 32 |
| BSE | breeding soundness examinations |
| cAMP | cyclic adenosine monophosphate |
| cBiMPS | 5,6-dichloro-1-β-D-ribofuranosylbenzimidazole-3′,5′-monophosphorothioate |
| [Ca2+]i | cytosolic/free intracellular calcium |
| DAG | diacylglycerol |
| EGFR | epidermal growth factor receptor |
| ERK | extracellular signal-regulated kinases |
| IP3 | inositol 1,4,5-trisphosphate |
| ICSI | intracytoplasmic sperm injection |
| MAPK | microtubule-associated protein kinases |
| NCX | Na+/Ca2+ ion channel |
| NKA | Na+-K+ ATPase |
| NHE | Na+/H+ exchanger |
| PIP2 | phosphatidylinositol 4,5-bisphosphate |
| PKA | protein kinase A |
| PLC/PKC | phospholipase C/protein kinase C |
| PI3K | phosphoinositide 3-kinase |
| PDK | 3-phosphoinositide-dependent kinase |
References
- FAO. Transforming Food and Agriculture to Acheive the SDGs; FAO: Rome, Italy, 2018; p. 71. Available online: http://www.fao.org/fao-stories/article/en/c/1184363/ (accessed on 13 May 2022).
- Kastelic, J.P. Male Involvement in Fertility and Factors Affecting Semen Quality in Bulls. Anim. Front. 2013, 3, 20–25. [Google Scholar] [CrossRef]
- Thundathil, J.C.; Rajamanickam, G.D.; Kastelic, J.P. Na/K-ATPase and Regulation of Sperm Function. Anim. Reprod. 2018, 15, 711–720. [Google Scholar] [CrossRef]
- Lewis, S.E.M. Is Sperm Evaluation Useful in Predicting Human Fertility? Reproduction 2007, 134, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Kastelic, J.; Thundathil, J. Breeding Soundness Evaluation and Semen Analysis for Predicting Bull Fertility. Reprod. Domest. Anim. 2008, 43, 368–373. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, H. Laboratory Semen Assessment and Prediction of Fertility: Still Utopia? Reprod. Domest. Anim. 2003, 38, 312–318. [Google Scholar] [CrossRef]
- Gadea, J.; Sellés, E.; Marco, M. The Predictive Value of Porcine Seminal Parameters on Fertility Outcome under Commercial Conditions: Porcine Seminal Parameters and Fertility Outcome. Reprod. Domest. Anim. 2004, 39, 303–308. [Google Scholar] [CrossRef]
- DeJarnette, J.M.; Marshall, C.E.; Lenz, R.W.; Monke, D.R.; Ayars, W.H.; Sattler, C.G. Sustaining the Fertility of Artificially Inseminated Dairy Cattle: The Role of the Artificial Insemination Industry. J. Dairy Sci. 2004, 87, E93–E104. [Google Scholar] [CrossRef]
- David, A.; Vilensky, A.; Nathan, H. Temperature Changes in the Different Parts of the Rabbit’s Oviduct. Int. J. Gynecol. Obstet. 1972, 10, 52–56. [Google Scholar] [CrossRef]
- Bahat, A.; Eisenbach, M.; Tur-Kaspa, I. Periovulatory Increase in Temperature Difference within the Rabbit Oviduct. Hum. Reprod. 2005, 20, 2118–2121. [Google Scholar] [CrossRef]
- Hunter, R.H.F.; Nichol, R. A Preovulatory Temperature Gradient between the Isthmus and Ampulla of Pig Oviducts during the Phase of Sperm Storage. Reproduction 1986, 77, 599–606. [Google Scholar] [CrossRef]
- El-Sheikh Ali, H.; Kitahara, G.; Tamura, Y.; Kobayashi, I.; Hemmi, K.; Torisu, S.; Sameshima, H.; Horii, Y.; Zaabel, S.; Kamimura, S. Presence of a Temperature Gradient among Genital Tract Portions and the Thermal Changes within These Portions over the Estrous Cycle in Beef Cows. J. Reprod. Dev. 2012, 59, 59–65. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Emmens, C.W. The Motility and Viability of Rabbit Spermatozoa at Different Hydrogen-Ion Concentrations. J. Physiol. 1947, 106, 471. [Google Scholar] [CrossRef]
- Pholpramool, C.; Chaturapanich, G. Effect of Sodium and Potassium Concentrations and PH on the Maintenance of Motility of Rabbit and Rat Epididymal Spermatozoa. Reproduction 1979, 57, 245–251. [Google Scholar] [CrossRef]
- Dale, B.; Menezo, Y.; Cohen, J.; DiMatteo, L.; Wilding, M. Intracellular PH Regulation in the Human Oocyte. Hum. Reprod. 1998, 13, 964–970. [Google Scholar] [CrossRef] [PubMed]
- Lishko, P.V.; Kirichok, Y.; Ren, D.; Navarro, B.; Chung, J.-J.; Clapham, D.E. The Control of Male Fertility by Spermatozoan Ion Channels. Annu. Rev. Physiol. 2012, 74, 453–475. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R.; Usui, N. Calcium Dependence of the Acrosome Reaction and Activation of Guinea Pig Spermatozoa. Exp. Cell Res. 1974, 89, 161–174. [Google Scholar] [CrossRef]
- Babcock, D.F.; Rufo, G.A.; Lardy, H.A. Potassium-Dependent Increases in Cytosolic PH Stimulate Metabolism and Motility of Mammalian Sperm. Proc. Natl. Acad. Sci. USA 1983, 80, 1327–1331. [Google Scholar] [CrossRef]
- Newton, L.D.; Kastelic, J.P.; Wong, B.; van der Hoorn, F.; Thundathil, J. Elevated Testicular Temperature Modulates Expression Patterns of Sperm Proteins in Holstein Bulls. Mol. Reprod. Dev. 2009, 76, 109–118. [Google Scholar] [CrossRef]
- Rajamanickam, G.D.; Kastelic, J.P.; Thundathil, J.C. Na/K-ATPase Regulates Bovine Sperm Capacitation through Raft- and Non-Raft-Mediated Signaling Mechanisms. Mol. Reprod. Dev. 2017, 84, 1168–1182. [Google Scholar] [CrossRef]
- Jimenez, T.; Sánchez, G.; Wertheimer, E.; Blanco, G. Activity of the Na, K-ATPase A4 Isoform Is Important for Membrane Potential, Intracellular Ca2+, and PH to Maintain Motility in Rat Spermatozoa. Reproduction 2010, 139, 835–845. [Google Scholar] [CrossRef]
- Wagoner, K.; Sanchez, G.; Nguyen, A.-N.; Enders, G.C.; Blanco, G. Different Expression and Activity of the A1 and A4 Isoforms of the Na, K-ATPase during Rat Male Germ Cell Ontogeny. Reproduction 2005, 130, 627–641. [Google Scholar] [CrossRef] [PubMed]
- McDermott, J.P.; Sánchez, G.; Mitra, A.; Numata, S.; Liu, L.C.; Blanco, G. Na, K-ATPase A4, and Not Na, K-ATPase A1, Is the Main Contributor to Sperm Motility, but its High Ouabain Binding Affinity Site Is Not Required for Male Fertility in Mice. J. Membr. Biol. 2021, 254, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Kocak-toker, N.; Aktan, G.; Aykac-toker, G. The Role of Na, K-ATPase in Human Sperm Motility. Int. J. Androl. 2002, 25, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Lestari, S.W.; Hinting, A.; Lunardi, H.; Aditya, D.; Miati, D.N.; Rizki, M.D. Sperm Na+, K+-ATPase and Dynein ATPase Activity: A Study of Embryo Development in In Vitro Fertilization (IVF); AIP Publishing LLC: Kurdistan, Iraq, 2019; p. 040019. [Google Scholar]
- Takei, G.L.; Fujinoki, M. Regulation of Hamster Sperm Hyperactivation by Extracellular Na. Reprod 2016, 151, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Skou, J.C. The Influence of Some Cations on an Adenosine Triphosphatase from Peripheral Nerves. Biochim. Biophys. Acta 1957, 23, 394–401. [Google Scholar] [CrossRef]
- Blanco, G.; Mercer, R.W. Isozymes of the Na-K-ATPase: Heterogeneity in Structure, Diversity in Function. Am. J. Physiol. Ren. Physiol. 1998, 275, F633–F650. [Google Scholar] [CrossRef]
- Blanco, G.; Sánchez, G.; Melton, R.J.; Tourtellotte, W.G.; Mercer, R.W. The A4 Isoform of the Na, K-ATPase is Expressed in the Germ Cells of the Testes. J. Histochem. Cytochem. 2000, 48, 1023–1032. [Google Scholar] [CrossRef]
- Woo, A.L.; James, P.F.; Lingrel, J.B. Sperm Motility Is Dependent on a Unique Isoform of the Na, K-ATPase. J. Biol. Chem. 2000, 275, 20693–20699. [Google Scholar] [CrossRef]
- Lingrel, J.; Moseley, A.; Dostanic, I.; Cougnon, M.; He, S.; James, P.; Woo, A.; O’Connor, K.; Neumann, J. Functional Roles of the α Isoforms of the Na, K-ATPase. Ann. N. Y. Acad. Sci. 2003, 986, 354–359. [Google Scholar] [CrossRef]
- Crambert, G.; Hasler, U.; Beggah, A.T.; Yu, C.; Modyanov, N.N.; Horisberger, J.-D.; Lelièvre, L.; Geering, K. Transport and Pharmacological Properties of Nine Different Human Na, K-ATPase Isozymes. J. Biol. Chem. 2000, 275, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Geering, K. Functional Roles of Na, K-ATPase Subunits. Curr. Opin. Nephrol. Hypertens. 2008, 17, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Schmalzing, G. Na+/K+-Pump Beta Subunits: Structure and Functions. Cell. Physiol. Biochem. 1994, 4, 96–114. [Google Scholar] [CrossRef]
- Kaplan, J.H. Biochemistry of Na, K-ATPase. Annu. Rev. Biochem. 2002, 71, 511–535. [Google Scholar] [CrossRef] [PubMed]
- McDonough, A.A.; Geering, K.; Farley, R.A. The Sodium Pump Needs Its β Subunit. FASEB J. 1990, 4, 1598–1605. [Google Scholar] [CrossRef]
- Liang, M.; Tian, J.; Liu, L.; Pierre, S.; Liu, J.; Shapiro, J.; Xie, Z.-J. Identification of a Pool of Non-Pumping Na/K-ATPase. J. Biol. Chem. 2007, 282, 10585–10593. [Google Scholar] [CrossRef]
- Clausen, T. Potassium and Sodium Transport and pH Regulation. Can. J. Physiol. Pharmacol. 1992, 70, S219–S222. [Google Scholar] [CrossRef]
- Gao, J.; Wymore, R.S.; Wang, Y.; Gaudette, G.R.; Krukenkamp, I.B.; Cohen, I.S.; Mathias, R.T. Isoform-Specific Stimulation of Cardiac Na/K Pumps by Nanomolar Concentrations of Glycosides. J. Gen. Physiol. 2002, 119, 297–312. [Google Scholar] [CrossRef]
- Haas, M.; Askari, A.; Xie, Z. Involvement of Src and Epidermal Growth Factor Receptor in the Signal-Transducing Function of Na+/K+-ATPase. J. Biol. Chem. 2000, 275, 27832–27837. [Google Scholar] [CrossRef]
- Mohammadi, K.; Kometiani, P.; Xie, Z.; Askari, A. Role of Protein Kinase C in the Signal Pathways That Link Na+/K+-ATPase to ERK1/2. J. Biol. Chem. 2001, 276, 42050–42056. [Google Scholar] [CrossRef]
- Tian, J.; Gong, X.; Xie, Z. Signal-Transducing Function of Na+-K+-ATPase Is Essential for Ouabain’s Effect on [Ca2+]i in Rat Cardiac Myocytes. Am. J. Physiol. Heart Circ. Physiol. 2001, 281, H1899–H1907. [Google Scholar] [CrossRef] [PubMed]
- Sweadner, K.J. Enzymatic Properties of Separated Isozymes of the Na, K-ATPase. Substrate Affinities, Kinetic Cooperativity, and Ion Transport Stoichiometry. J. Biol. Chem. 1985, 260, 11508–11513. [Google Scholar] [CrossRef]
- Erdmann, E.; Werdan, K.; Brown, L. Multiplicity of Cardiac Glycoside Receptors in the Heart. Trends Pharmacol. Sci. 1985, 6, 293–295. [Google Scholar] [CrossRef]
- Inoue, N.; Matsui, H.; Tsukui, H.; Hatanaka, H. The Appearance of a Highly Digitalis-Sensitive Isoform of Na+, K+-ATPase during Maturation In Vitro of Primary Cultured Rat Cerebral Neurons. J. Biochem. 1988, 104, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Obrien, W.J.; Lingrel, J.B.; Wallick, E.T. Ouabain Binding Kinetics of the Rat Alpha Two and Alpha Three Isoforms of the Sodium-Potassium Adenosine Triphosphate. Arch. Biochem. Biophys. 1994, 310, 32–39. [Google Scholar] [CrossRef]
- Liu, J.; Tian, J.; Haas, M.; Shapiro, J.I.; Askari, A.; Xie, Z. Ouabain Interaction with Cardiac Na+/K+-ATPase Initiates Signal Cascades Independent of Changes in Intracellular Na+ and Ca2+ Concentrations. J. Biol. Chem. 2000, 275, 27838–27844. [Google Scholar] [CrossRef]
- Cai, T.; Wang, H.; Chen, Y.; Liu, L.; Gunning, W.T.; Quintas, L.E.M.; Xie, Z.-J. Regulation of Caveolin-1 Membrane Trafficking by the Na/K-ATPase. J. Cell Biol. 2008, 182, 1153–1169. [Google Scholar] [CrossRef]
- Staubach, S.; Hanisch, F.-G. Lipid Rafts: Signaling and Sorting Platforms of Cells and Their Roles in Cancer. Expert Rev. Proteom. 2011, 8, 263–277. [Google Scholar] [CrossRef]
- Kometiani, P.; Li, J.; Gnudi, L.; Kahn, B.B.; Askari, A.; Xie, Z. Multiple Signal Transduction Pathways Link Na+/K+-ATPase to Growth-Related Genes in Cardiac Myocytes. J. Biol. Chem. 1998, 273, 15249–15256. [Google Scholar] [CrossRef]
- Aizman, O.; Uhlén, P.; Lal, M.; Brismar, H.; Aperia, A. Ouabain, a Steroid Hormone That Signals with Slow Calcium Oscillations. Proc. Natl. Acad. Sci. USA 2001, 98, 13420–13424. [Google Scholar] [CrossRef]
- Aydemir-Koksoy, A.; Abramowitz, J.; Allen, J.C. Ouabain-Induced Signaling and Vascular Smooth Muscle Cell Proliferation. J. Biol. Chem. 2001, 276, 46605–46611. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Cai, T.; Yuan, Z.; Wang, H.; Liu, L.; Haas, M.; Maksimova, E.; Huang, X.-Y.; Xie, Z.-J. Binding of Src to Na+/K+-ATPase Forms a Functional Signaling Complex. Mol. Biol. Cell 2006, 17, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Askari, A. Na+/K+-ATPase as a Signal Transducer. Eur. J. Biochem. 2002, 269, 2434–2439. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhao, X.; Pierre, S.V.; Askari, A. Association of PI3K-Akt Signaling Pathway with Digitalis-Induced Hypertrophy of Cardiac Myocytes. Am. J. Physiol. Cell Physiol. 2007, 293, C1489–C1497. [Google Scholar] [CrossRef] [PubMed]
- Shamraj, O.I.; Lingrel, J.B. A Putative Fourth Na+, K+-ATPase Alpha-Subunit Gene Is Expressed in Testis. Proc. Natl. Acad. Sci. USA 1994, 91, 12952–12956. [Google Scholar] [CrossRef]
- Sanchez, G.; Nguyen, A.-N.T.; Timmerberg, B.; Tash, J.S.; Blanco, G. The Na, K-ATPase A4 Isoform from Humans Has Distinct Enzymatic Properties and Is Important for Sperm Motility. MHR Basic Sci. Reprod. Med. 2006, 12, 565–576. [Google Scholar] [CrossRef]
- Hickey, K.D.; Buhr, M.M. Characterization of Na+K+-ATPase in Bovine Sperm. Theriogenology 2012, 77, 1369–1380. [Google Scholar] [CrossRef]
- Thundathil, J.C.; Anzar, M.; Buhr, M.M. Na+/K+ATPase as a Signaling Molecule during Bovine Sperm Capacitation. Biol. Reprod. 2006, 75, 308–317. [Google Scholar] [CrossRef]
- Unnikrishnan, V.; Kastelic, J.P.; Thundathil, J.C. Ouabain-Induced Activation of Phospholipase C Zeta and Its Contributions to Bovine Sperm Capacitation. Cell Tissue Res. 2021, 385, 785–801. [Google Scholar] [CrossRef]
- Clausen, M.V.; Nissen, P.; Poulsen, H. The A4 Isoform of the Na+, K+-ATPase Is Tuned for Changing Extracellular Environments. FEBS J. 2016, 283, 282–293. [Google Scholar] [CrossRef]
- Rajamanickam, G.D.; Kroetsch, T.; Kastelic, J.P.; Thundathil, J.C. Testis-Specific Isoform of Na/K-ATPase (ATP1A4) Regulates Sperm Function and Fertility in Dairy Bulls through Potential Mechanisms Involving Reactive Oxygen Species, Calcium and Actin Polymerization. Andrology 2017, 5, 814–823. [Google Scholar] [CrossRef]
- Yanagimachi, R. The Movement of Golden Hamster Spermatozoa before and after Capacitation. Reproduction 1970, 23, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Gadella, B.M.; Tsai, P.; Boerke, A.; Brewis, I.A. Sperm Head Membrane Reorganisation during Capacitation. Int. J. Dev. Biol. 2008, 52, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Salicioni, A.M.; Platt, M.D.; Wertheimer, E.V.; Arcelay, E.; Allaire, A.; Sosnik, J.; Visconti, P.E. Signalling Pathways Involved in Sperm Capacitation. Soc. Reprod. Fertil. Suppl. 2007, 65, 245–259. [Google Scholar] [PubMed]
- Leese, H.J. The Formation and Function of Oviduct Fluid. Reproduction 1988, 82, 843–856. [Google Scholar] [CrossRef]
- Daniel, L.; Etkovitz, N.; Weiss, S.R.; Rubinstein, S.; Ickowicz, D.; Breitbart, H. Regulation of the Sperm EGF Receptor by Ouabain Leads to Initiation of the Acrosome Reaction. Dev. Biol. 2010, 344, 650–657. [Google Scholar] [CrossRef]
- Hobkirk, R. Steroid Sulfotransferases and Steroid Sulfate Sulfatases: Characteristics and Biological Roles. Can. J. Biochem. Cell Biol. 1985, 63, 1127–1144. [Google Scholar] [CrossRef]
- Libersky, E.A.; Boatman, D.E. Effects of Progesterone on In Vitro Sperm Capacitation and Egg Penetration in the Golden Hamster1. Biol. Reprod. 1995, 53, 483–487. [Google Scholar] [CrossRef]
- Hernández-Silva, G.; Chirinos, M. Proteins from Male and Female Reproductive Tracts Involved in Sperm Function Regulation. Zygote 2019, 27, 5–16. [Google Scholar] [CrossRef]
- Sandtner, W.; Egwolf, B.; Khalili-Araghi, F.; Sánchez-Rodríguez, J.E.; Roux, B.; Bezanilla, F.; Holmgren, M. Ouabain Binding Site in a Functioning Na+/K+ ATPase. J. Biol. Chem. 2011, 286, 38177–38183. [Google Scholar] [CrossRef]
- Sajeevadathan, M.; Pettitt, M.J.; Buhr, M.M. Are Isoforms of Capacitating Na+-K+-ATPase Localized to Sperm Head Rafts? Mol. Reprod. Dev. 2021, 88, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Rajamanickam, G.D.; Kastelic, J.P.; Thundathil, J.C. Testis-Specific Isoform of Na/K-ATPase (ATP1A4) Interactome in Raft and Non-Raft Membrane Fractions from Capacitated Bovine Sperm. Int. J. Mol. Sci. 2019, 20, 3159. [Google Scholar] [CrossRef] [PubMed]
- Ickowicz, D.; Finkelstein, M.; Breitbart, H. Mechanism of Sperm Capacitation and the Acrosome Reaction: Role of Protein Kinases. Asian J. Androl. 2012, 14, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Rotfeld, H.; Hillman, P.; Ickowicz, D.; Breitbart, H. PKA and CaMKII Mediate PI3K Activation in Bovine Sperm by Inhibition of the PKC/PP1 Cascade. Reproduction 2014, 147, 347–356. [Google Scholar] [CrossRef][Green Version]
- Aquila, S.; Sisci, D.; Gentile, M.; Middea, E.; Catalano, S.; Carpino, A.; Rago, V.; Andò, S. Estrogen Receptor (ER)α and ERβ Are Both Expressed in Human Ejaculated Spermatozoa: Evidence of Their Direct Interaction with Phosphatidylinositol-3-OH Kinase/Akt Pathway. J. Clin. Endocrinol. Metab. 2004, 89, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Awda, B.J.; Buhr, M.M. Extracellular Signal-Regulated Kinases (ERKs) Pathway and Reactive Oxygen Species Regulate Tyrosine Phosphorylation in Capacitating Boar Spermatozoa1. Biol. Reprod. 2010, 83, 750–758. [Google Scholar] [CrossRef]
- Anpalakan, K.; Buhr, M.M.; Hickey, K.D. Src Is a Signaling Molecule in Ouabain-Na+K+-ATPase-Stimulated Signaling Pathway during Bull Sperm Capacitation. Biol. Reprod. 2010, 83, 497. [Google Scholar] [CrossRef]
- Sajeevadathan, M.; Pettitt, M.J.; Buhr, M. Interaction of Ouabain and Progesterone on Induction of Bull Sperm Capacitation. Theriogenology 2019, 126, 191–198. [Google Scholar] [CrossRef]
- Morrill, G.A.; Erlichman, J.; Gutierrez-Juarez, R.; Kostellow, A.B. The Steroid-Binding Subunit of the Na/K-ATPase as a Progesterone Receptor on the Amphibian Oocyte Plasma Membrane. Steroids 2005, 70, 933–945. [Google Scholar] [CrossRef]
- Morrill, G.A.; Kostellow, A.B.; Askari, A. Progesterone Modulation of Transmembrane Helix-Helix Interactions between the α-Subunit of Na/K-ATPase and Phospholipid N-Methyltransferase in the Oocyte Plasma Membrane. BMC Struct. Biol. 2010, 10, 12. [Google Scholar] [CrossRef]
- Sajeevadathan, M. Role of Na+/K+ ATPase in Bull Sperm Capacitation. Ph.D. Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 2018. [Google Scholar]
- Saha, S.R.; Sakase, M.; Fukushima, M.; Harayama, H. Effects of Digoxin on Full-Type Hyperactivation in Bovine Ejaculated Spermatozoa with Relatively Lower Survivability for Incubation with Stimulators of cAMP Signaling Cascades. Theriogenology 2020, 154, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.M.; Rubenstein, J.L.R.; Parce, J.W.; McConnell, H.M. Lateral Diffusion of M-13 Coat Protein in Mixtures of Phosphatidylcholine and Cholesterol. Biochemistry 1980, 19, 5907–5911. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Vaz, W.L.C. Model Systems, Lipid Rafts, and Cell Membranes. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 269–295. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.E. Lipid-Mediated Localization of Signaling Proteins. In Handbook of Cell Signaling; Elsevier: Amsterdam, The Netherlands, 2010; pp. 365–371. ISBN 978-0-12-374145-5. [Google Scholar]
- Munro, S. Lipid Rafts. Cell 2003, 115, 377–388. [Google Scholar] [CrossRef]
- Watanabe, H.; Takeda, R.; Hirota, K.; Kondoh, G. Lipid Raft Dynamics Linked to Sperm Competency for Fertilization in Mice. Genes Cells 2017, 22, 493–500. [Google Scholar] [CrossRef]
- Shadan, S.; James, P.S.; Howes, E.A.; Jones, R. Cholesterol Efflux Alters Lipid Raft Stability and Distribution during Capacitation of Boar Spermatozoa. Biol. Reprod. 2004, 71, 253–265. [Google Scholar] [CrossRef]
- Ushiyama, A.; Tajima, A.; Ishikawa, N.; Asano, A. Characterization of the Functions and Proteomes Associated with Membrane Rafts in Chicken Sperm. PLoS ONE 2017, 12, e0186482. [Google Scholar] [CrossRef]
- Cross, N.L. Reorganization of Lipid Rafts during Capacitation of Human Sperm. Biol. Reprod. 2004, 71, 1367–1373. [Google Scholar] [CrossRef]
- van Gestel, R.A.; Brewis, I.A.; Ashton, P.R.; Brouwers, J.F.; Gadella, B.M. Multiple Proteins Present in Purified Porcine Sperm Apical Plasma Membranes Interact with the Zona Pellucida of the Oocyte. MHR Basic Sci. Reprod. Med. 2007, 13, 445–454. [Google Scholar] [CrossRef]
- Boarelli, P.V.; Monclus, M.D.L.A.; Lancelotti, T.S.; Cabrillana, M.E.; Simón, L.Y.; Funes, A.K.; Fornes, M.W. Simultaneous Study of Cholesterol and GM1 Ganglioside by Specific Probes: Lipid Distribution during Maturation, Capacitation and the Acrosome Reaction. J. Cytol. Histol. 2016, 7, 1000412. [Google Scholar] [CrossRef]
- Jones, R.; James, P.S.; Howes, L.; Bruckbauer, A.; Klenerman, D. Supramolecular Organization of the Sperm Plasma Membrane during Maturation and Capacitation. Asian J. Androl. 2007, 9, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Mukherjee, S.; Maxfield, F.R. Cholesterol Depletion Induces Large Scale Domain Segregation in Living Cell Membranes. Proc. Natl. Acad. Sci. USA 2001, 98, 13072–13077. [Google Scholar] [CrossRef] [PubMed]
- Osheroff, J.E. Regulation of Human Sperm Capacitation by a Cholesterol Efflux-Stimulated Signal Transduction Pathway Leading to Protein Kinase A-Mediated up-Regulation of Protein Tyrosine Phosphorylation. Mol. Hum. Reprod. 1999, 5, 1017–1026. [Google Scholar] [CrossRef]
- Tanphaichitr, N. New Insights into Sperm-Zona Pellucida Interaction: Involvement of Sperm Lipid Ra. Front. Biosci. 2007, 12, 1748–1766. [Google Scholar] [CrossRef] [PubMed]
- Nixon, B.; Bielanowicz, A.; Mclaughlin, E.A.; Tanphaichitr, N.; Ensslin, M.A.; Aitken, R.J. Composition and Significance of Detergent Resistant Membranes in Mouse Spermatozoa. J. Cell. Physiol. 2009, 218, 122–134. [Google Scholar] [CrossRef]
- Nixon, B.; Mitchell, L.A.; Anderson, A.L.; Mclaughlin, E.A.; O’bryan, M.K.; Aitken, R.J. Proteomic and Functional Analysis of Human Sperm Detergent Resistant Membranes. J. Cell. Physiol. 2011, 226, 2651–2665. [Google Scholar] [CrossRef]
- Maehashi, E.; Sato, C.; Ohta, K.; Harada, Y.; Matsuda, T.; Hirohashi, N.; Lennarz, W.J.; Kitajima, K. Identification of the Sea Urchin 350-KDa Sperm-Binding Protein as a New Sialic Acid-Binding Lectin That Belongs to the Heat Shock Protein 110 Family. J. Biol. Chem. 2003, 278, 42050–42057. [Google Scholar] [CrossRef]
- Sleight, S.B.; Miranda, P.V.; Plaskett, N.-W.; Maier, B.; Lysiak, J.; Scrable, H.; Herr, J.C.; Visconti, P.E. Isolation and Proteomic Analysis of Mouse Sperm Detergent-Resistant Membrane Fractions: Evidence for Dissociation of Lipid Rafts during Capacitation. Biol. Reprod. 2005, 73, 721–729. [Google Scholar] [CrossRef]
- Asano, A.; Selvaraj, V.; Buttke, D.E.; Nelson, J.L.; Green, K.M.; Evans, J.E.; Travis, A.J. Biochemical Characterization of Membrane Fractions in Murine Sperm: Identification of Three Distinct Sub-Types of Membrane Rafts. J. Cell. Physiol. 2009, 218, 537–548. [Google Scholar] [CrossRef]
- Asano, A.; Nelson, J.L.; Zhang, S.; Travis, A.J. Characterization of the Proteomes Associating with Three Distinct Membrane Raft Sub-Types in Murine Sperm. Proteomics 2010, 10, 3494–3505. [Google Scholar] [CrossRef]
- Kelsey, K.M.; Zigo, M.; Thompson, W.E.; Kerns, K.; Manandhar, G.; Sutovsky, M.; Sutovsky, P. Reciprocal Surface Expression of Arylsulfatase A and Ubiquitin in Normal and Defective Mammalian Spermatozoa. Cell Tissue Res. 2020, 379, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Ojaghi, M.; Kastelic, J.; Thundathil, J.C. Testis-Specific Isoform of Angiotensin-Converting Enzyme (tACE) as a Candidate Marker for Bull Fertility. Reprod. Fertil. Dev. 2018, 30, 1584. [Google Scholar] [CrossRef] [PubMed]
- Coonrod, S.A.; Herr, J.C.; Westhusin, M.E. Inhibition of Bovine Fertilization in Vitro by Antibodies to SP-10. Reproduction 1996, 107, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Tardif, S.; Wilson, M.D.; Wagner, R.; Hunt, P.; Gertsenstein, M.; Nagy, A.; Lobe, C.; Koop, B.F.; Hardy, D.M. Zonadhesin Is Essential for Species Specificity of Sperm Adhesion to the Egg Zona Pellucida. J. Biol. Chem. 2010, 285, 24863–24870. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Xu, W.; Yi, Y.-J.; Sutovsky, P.; Oko, R. The Extracellular Protein Coat of the Inner Acrosomal Membrane Is Involved in Zona Pellucida Binding and Penetration during Fertilization: Characterization of Its Most Prominent Polypeptide (IAM38). Dev. Biol. 2006, 290, 32–43. [Google Scholar] [CrossRef]
- Post, H.; Schwarz, A.; Brandenburger, T.; Aumüller, G.; Wilhelm, B. Arrangement of PMCA4 in Bovine Sperm Membrane Fractions: PMCA in Sperm Membranes. Int. J. Androl. 2010, 33, 775–783. [Google Scholar] [CrossRef]
- Wennemuth, G.; Babcock, D.F.; Hille, B. Calcium Clearance Mechanisms of Mouse Sperm. J. Gen. Physiol. 2003, 122, 115–128. [Google Scholar] [CrossRef]
- Chakraborti, S.; Dhalla, N.S. Regulation of Membrane Na+-K+ ATPase; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 978-3-319-79672-7. [Google Scholar]
- Visconti, P.E. Understanding the Molecular Basis of Sperm Capacitation through Kinase Design. Proc. Natl. Acad. Sci. USA 2009, 106, 667–668. [Google Scholar] [CrossRef]
- Takei, G.L.; Hayashi, K. Na+/K+-ATPase A4 Regulates Sperm Hyperactivation While Na+/K+-ATPase A1 Regulates Basal Motility in Hamster Spermatozoa. Theriogenology 2020, 157, 48–60. [Google Scholar] [CrossRef]
- Paoli, D.; Gallo, M.; Rizzo, F.; Baldi, E.; Francavilla, S.; Lenzi, A.; Lombardo, F.; Gandini, L. Mitochondrial Membrane Potential Profile and Its Correlation with Increasing Sperm Motility. Fertil. Steril. 2011, 95, 2315–2319. [Google Scholar] [CrossRef]
- Counillon, L.; Pouysségur, J. The Expanding Family of Eukaryotic Na+/H+Exchangers. J. Biol. Chem. 2000, 275, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, C.B.; Goltz, J.S.; Kanous, K.S.; Gardner, T.K.; Olds-Clarke, P. Evidence for an Increased Sensitivity to Ca2+ in the Flagella of Sperm from Tw32/+ Mice. Mol. Reprod. Dev. 1990, 26, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Bedu-Addo, K.; Costello, S.; Harper, C.; Machado-Oliveira, G.; Lefievre, L.; Ford, C.; Barratt, C.; Publicover, S. Mobilisation of Stored Calcium in the Neck Region of Human Sperm a Mechanism for Regulation of Flagellar Activity. Int. J. Dev. Biol. 2008, 52, 615–626. [Google Scholar] [CrossRef]
- Zhang, S.; Malmersjö, S.; Li, J.; Ando, H.; Aizman, O.; Uhlén, P.; Mikoshiba, K.; Aperia, A. Distinct Role of the N-Terminal Tail of the Na, K-ATPase Catalytic Subunit as a Signal Transducer. J. Biol. Chem. 2006, 281, 21954–21962. [Google Scholar] [CrossRef]
- Tian, J.; Xie, Z. The Na-K-ATPase and Calcium-Signaling Microdomains. Physiology 2008, 23, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Lestari, S.W.; Miati, D.N.; Seoharso, P.; Sugiyanto, R.; Pujianto, D.A. Sperm Na+, K+-ATPase A4 and Plasma Membrane Ca2+-ATPase (PMCA) 4 Regulation in Asthenozoospermia. Syst. Biol. Reprod. Med. 2017, 63, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Takezawa, Y.; Yoshida, K.; Miyado, K.; Sato, M.; Nakamura, A.; Kawano, N.; Sakakibara, K.; Kondo, T.; Harada, Y.; Ohnami, N.; et al. β-Catenin Is a Molecular Switch That Regulates Transition of Cell-Cell Adhesion to Fusion. Sci. Rep. 2011, 1, 68. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, S.; Yoshida, N.; Fukui, T.; Amanai, M.; Isobe, T.; Itagaki, C.; Izumi, T.; Perry, A.C.F. Mammalian Phospholipase Cζ Induces Oocyte Activation from the Sperm Perinuclear Matrix. Dev. Biol. 2004, 274, 370–383. [Google Scholar] [CrossRef]
- Escoffier, J.; Yassine, S.; Lee, H.C.; Martinez, G.; Delaroche, J.; Coutton, C.; Karaouzène, T.; Zouari, R.; Metzler-Guillemain, C.; Pernet-Gallay, K.; et al. Subcellular Localization of Phospholipase Cζ in Human Sperm and Its Absence in DPY19L2-Deficient Sperm Are Consistent with Its Role in Oocyte Activation. MHR Basic Sci. Reprod. Med. 2015, 21, 157–168. [Google Scholar] [CrossRef]
- Kurokawa, M.; Sato, K.; Fissore, R.A. Mammalian Fertilization: From Sperm Factor to Phospholipase Cζ. Biol. Cell 2004, 96, 37–45. [Google Scholar] [CrossRef]
- Ducibella, T.; Huneau, D.; Angelichio, E.; Xu, Z.; Schultz, R.M.; Kopf, G.S.; Fissore, R.; Madoux, S.; Ozil, J.-P. Egg-to-Embryo Transition Is Driven by Differential Responses to Ca2+ Oscillation Number. Dev. Biol. 2002, 250, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.; Agarwal, A.; Pushparaj, P.; Ahmad, G.; Sharma, R. Reduced Semen Quality in Patients with Testicular Cancer Seminoma Is Associated with Alterations in the Expression of Sperm Proteins. Asian J. Androl. 2020, 22, 88. [Google Scholar] [CrossRef] [PubMed]
- Gur, Y.; Breitbart, H. Protein Synthesis in Sperm: Dialog between Mitochondria and Cytoplasm. Mol. Cell. Endocrinol. 2008, 282, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Gur, Y.; Breitbart, H. Mammalian Sperm Translate Nuclear-Encoded Proteins by Mitochondrial-Type Ribosomes. Genes Dev. 2006, 20, 411–416. [Google Scholar] [CrossRef]
- Castillo, J.; Bogle, O.A.; Jodar, M.; Torabi, F.; Delgado-Dueñas, D.; Estanyol, J.M.; Ballescà, J.L.; Miller, D.; Oliva, R. Proteomic Changes in Human Sperm During Sequential In Vitro Capacitation and Acrosome Reaction. Front. Cell Dev. Biol. 2019, 7, 295. [Google Scholar] [CrossRef]
- Li, Y.; Li, R.-H.; Ran, M.-X.; Zhang, Y.; Liang, K.; Ren, Y.-N.; He, W.-C.; Zhang, M.; Zhou, G.-B.; Qazi, I.H.; et al. High Throughput Small RNA and Transcriptome Sequencing Reveal Capacitation-Related MicroRNAs and MRNA in Boar Sperm. BMC Genom. 2018, 19, 736. [Google Scholar] [CrossRef]
- Cortadellas, N.; Durfort, M. Fate and Composition of Cytopiasmic Droplet of Hamster Epididymal Spermatozoa. J. Morphol. 1994, 221, 199–210. [Google Scholar] [CrossRef]
- Premkumar, E.; Bhargava, P.M. Transcription and Translation in Bovine Spermatozoa. Nat. New Biol. 1972, 240, 139–143. [Google Scholar] [CrossRef]
- Abaigar, T.; Holt, W.V.; Harrison, R.A.P.; del Barrio, G. Sperm Subpopulations in Boar (Sus Scrofa) and Gazelle (Gazella Dama Mhorr) Semen as Revealed by Pattern Analysis of Computer-Assisted Motility Assessments. Biol. Reprod. 1999, 60, 32–41. [Google Scholar] [CrossRef]
- Muiño, R.; Tamargo, C.; Hidalgo, C.O.; Peña, A.I. Identification of Sperm Subpopulations with Defined Motility Characteristics in Ejaculates from Holstein Bulls: Effects of Cryopreservation and between-Bull Variation. Anim. Reprod. Sci. 2008, 109, 27–39. [Google Scholar] [CrossRef]
- Ibanescu, I.; Siuda, M.; Bollwein, H. Motile Sperm Subpopulations in Bull Semen Using Different Clustering Approaches–Associations with Flow Cytometric Sperm Characteristics and Fertility. Anim. Reprod. Sci. 2020, 215, 106329. [Google Scholar] [CrossRef] [PubMed]
- Thurston, L.M.; Watson, P.F.; Holt, W.V. Sources of Variation in the Morphological Characteristics of Sperm Subpopulations Assessed Objectively by a Novel Automated Sperm Morphology Analysis System. Reproduction 1999, 117, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Valverde, A.; Arenán, H.; Sancho, M.; Contell, J.; Yániz, J.; Fernández, A.; Soler, C. Morphometry and Subpopulation Structure of Holstein Bull Spermatozoa: Variations in Ejaculates and Cryopreservation Straws. Asian J. Androl. 2016, 18, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Nesci, S.; Spinaci, M.; Galeati, G.; Nerozzi, C.; Pagliarani, A.; Algieri, C.; Tamanini, C.; Bucci, D. Sperm Function and Mitochondrial Activity: An Insight on Boar Sperm Metabolism. Theriogenology 2020, 144, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Boerke, A.; Dieleman, S.J.; Gadella, B.M. A Possible Role for Sperm RNA in Early Embryo Development. Theriogenology 2007, 68, S147–S155. [Google Scholar] [CrossRef]
- Hosken, D.J.; Hodgson, D.J. Why Do Sperm Carry RNA? Relatedness, Conflict, and Control. Trends Ecol. Evol. 2014, 29, 451–455. [Google Scholar] [CrossRef]
- Wang, J.; Fan, H.C.; Behr, B.; Quake, S.R. Genome-Wide Single-Cell Analysis of Recombination Activity and de Novo Mutation Rates in Human Sperm. Cell 2012, 150, 402–412. [Google Scholar] [CrossRef]
- Pausch, H.; Kölle, S.; Wurmser, C.; Schwarzenbacher, H.; Emmerling, R.; Jansen, S.; Trottmann, M.; Fuerst, C.; Götz, K.-U.; Fries, R. A Nonsense Mutation in TMEM95 Encoding a Nondescript Transmembrane Protein Causes Idiopathic Male Subfertility in Cattle. PLoS Genet. 2014, 10, e1004044. [Google Scholar] [CrossRef]
- Sutovsky, P.; Neuber, E.; Schatten, G. Ubiquitin-Dependent Sperm Quality Control Mechanism Recognizes Spermatozoa with DNA Defects as Revealed by Dual Ubiquitin-TUNEL Assay. Mol. Reprod. Dev. 2002, 61, 406–413. [Google Scholar] [CrossRef]
- Sutovsky, P.; Aarabi, M.; Miranda-Vizuete, A.; Oko, R. Negative Biomarker Based Male Fertility Evaluation: Sperm Phenotypes Associated with Molecular-Level Anomalies. Asian J. Androl. 2015, 17, 554. [Google Scholar] [CrossRef]
- Kennedy, C.E.; Krieger, K.B.; Sutovsky, M.; Xu, W.; Vargovič, P.; Didion, B.A.; Ellersieck, M.R.; Hennessy, M.E.; Verstegen, J.; Oko, R.; et al. Protein Expression Pattern of PAWP in Bull Spermatozoa Is Associated with Sperm Quality and Fertility Following Artificial Insemination: Protein expression pattern of PAWP. Mol. Reprod. Dev. 2014, 81, 436–449. [Google Scholar] [CrossRef] [PubMed]
- Kaya, A.; Dogan, S.; Vargovic, P.; Kutchy, N.A.; Ross, P.; Topper, E.; Oko, R.; van der Hoorn, F.; Sutovsky, P.; Memili, E. Sperm Proteins ODF2 and PAWP as Markers of Fertility in Breeding Bulls. Cell Tissue Res. 2022, 387, 159–171. [Google Scholar] [CrossRef]
- Somashekar, L.; Selvaraju, S.; Parthipan, S.; Patil, S.K.; Binsila, B.K.; Venkataswamy, M.M.; Karthik Bhat, S.; Ravindra, J.P. Comparative Sperm Protein Profiling in Bulls Differing in Fertility and Identification of Phosphatidylethanolamine-Binding Protein 4, a Potential Fertility Marker. Andrology 2017, 5, 1032–1051. [Google Scholar] [CrossRef] [PubMed]
- Ojaghi, M.; Varghese, J.; Kastelic, J.P.; Thundathil, J.C. Characterization of the Testis-Specific Angiotensin Converting Enzyme (tACE)-Interactome during Bovine Sperm Capacitation. Curr. Issues Mol. Biol. 2022, 44, 449–469. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiwari, S.; Rajamanickam, G.; Unnikrishnan, V.; Ojaghi, M.; Kastelic, J.P.; Thundathil, J.C. Testis-Specific Isoform of Na+-K+ ATPase and Regulation of Bull Fertility. Int. J. Mol. Sci. 2022, 23, 7936. https://doi.org/10.3390/ijms23147936
Tiwari S, Rajamanickam G, Unnikrishnan V, Ojaghi M, Kastelic JP, Thundathil JC. Testis-Specific Isoform of Na+-K+ ATPase and Regulation of Bull Fertility. International Journal of Molecular Sciences. 2022; 23(14):7936. https://doi.org/10.3390/ijms23147936
Chicago/Turabian StyleTiwari, Saurabh, Gayathri Rajamanickam, Veena Unnikrishnan, Mina Ojaghi, John P. Kastelic, and Jacob C. Thundathil. 2022. "Testis-Specific Isoform of Na+-K+ ATPase and Regulation of Bull Fertility" International Journal of Molecular Sciences 23, no. 14: 7936. https://doi.org/10.3390/ijms23147936
APA StyleTiwari, S., Rajamanickam, G., Unnikrishnan, V., Ojaghi, M., Kastelic, J. P., & Thundathil, J. C. (2022). Testis-Specific Isoform of Na+-K+ ATPase and Regulation of Bull Fertility. International Journal of Molecular Sciences, 23(14), 7936. https://doi.org/10.3390/ijms23147936