
Metabolomic Evidence for Peroxisomal Dysfunction in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome

 , , , ,
, , , ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
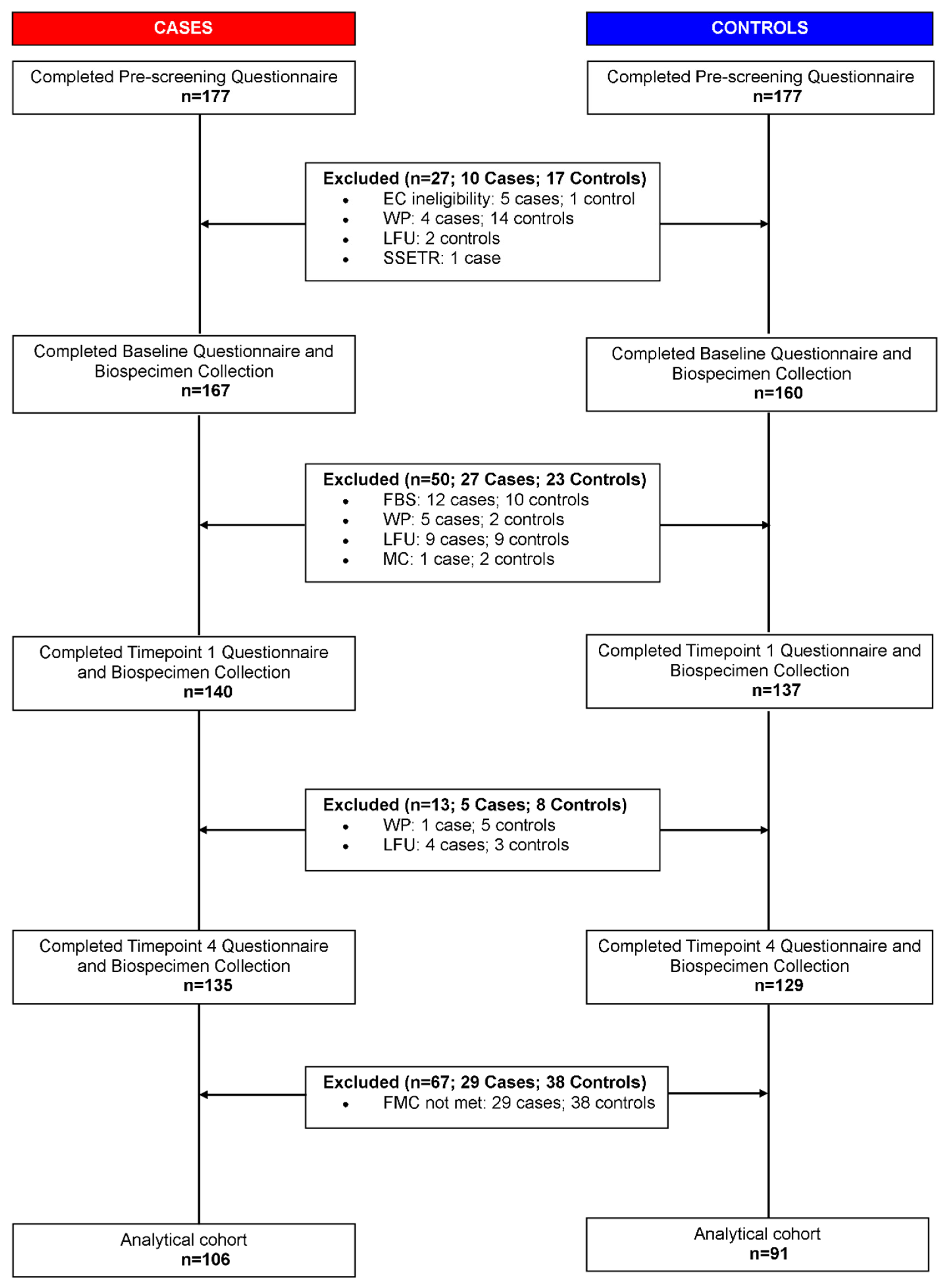
2.1. Study Population Characteristics
2.2. Metabolomic Dataset
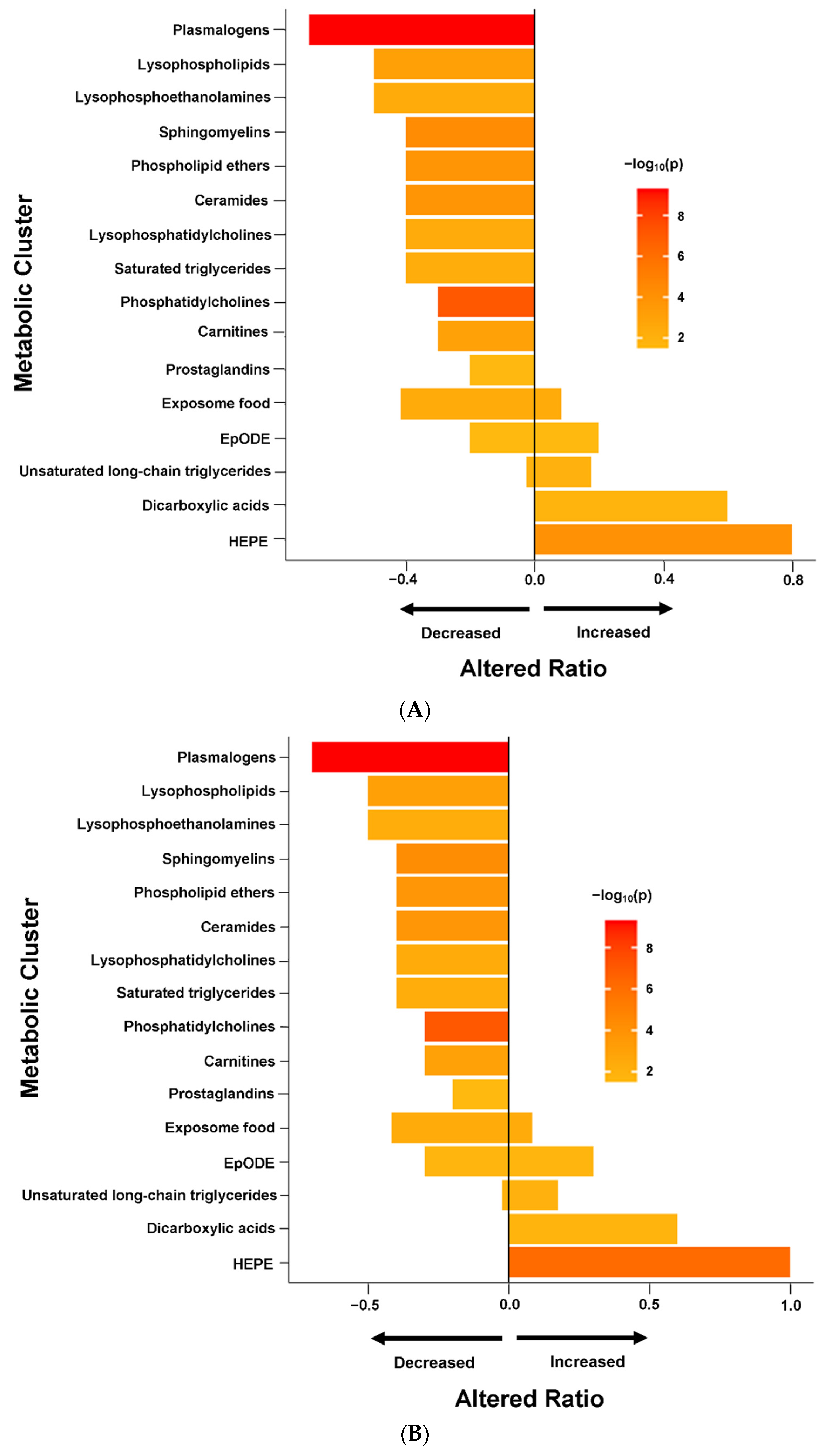
2.3. ME/CFS Associated with Altered Metabolomic Profile
2.4. Altered Metabolomic Profiles in Female and Male ME/CFS Patients
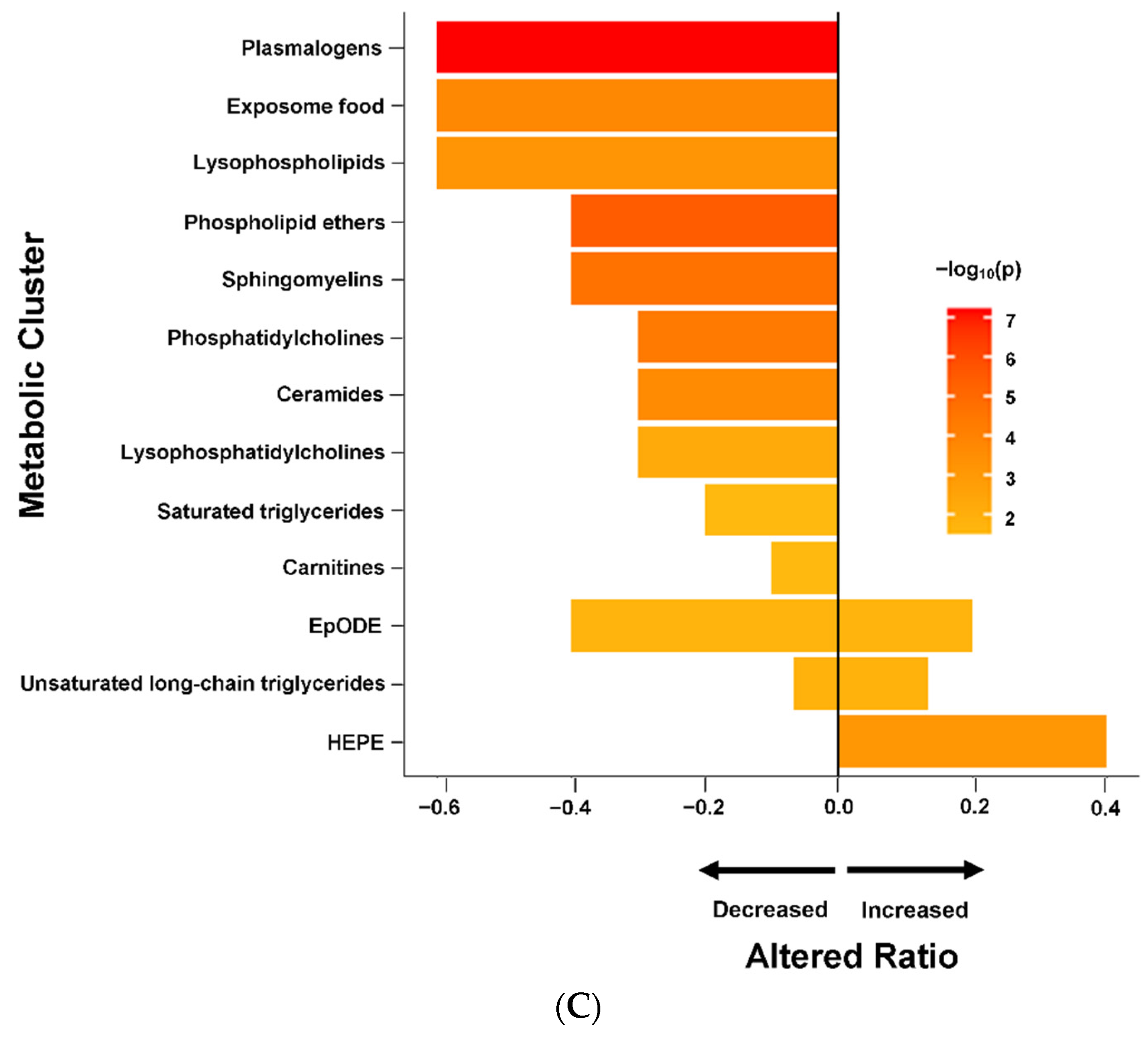
2.5. Altered Metabolomics Profile in ME/CFS Patients without sr-IBS
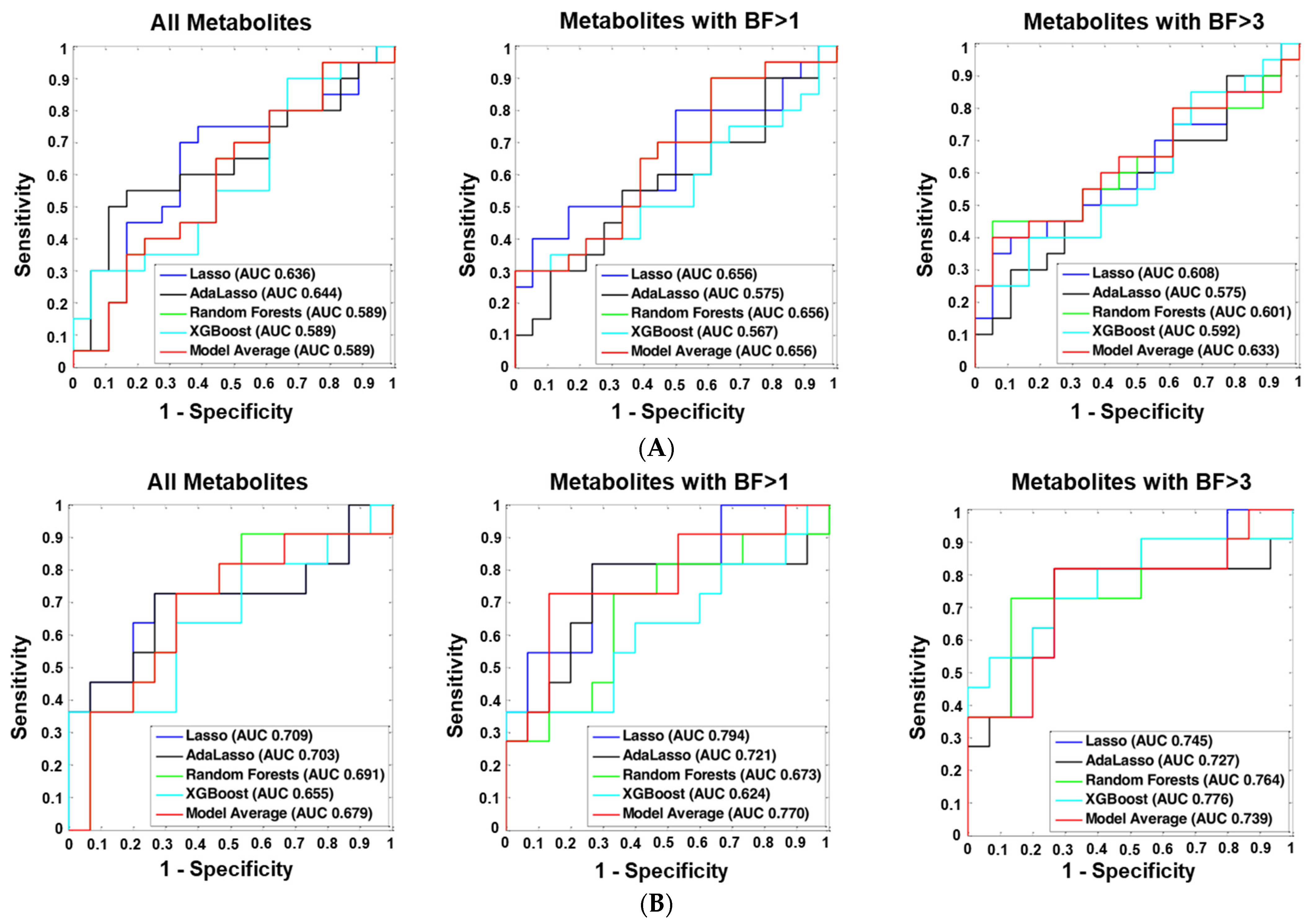
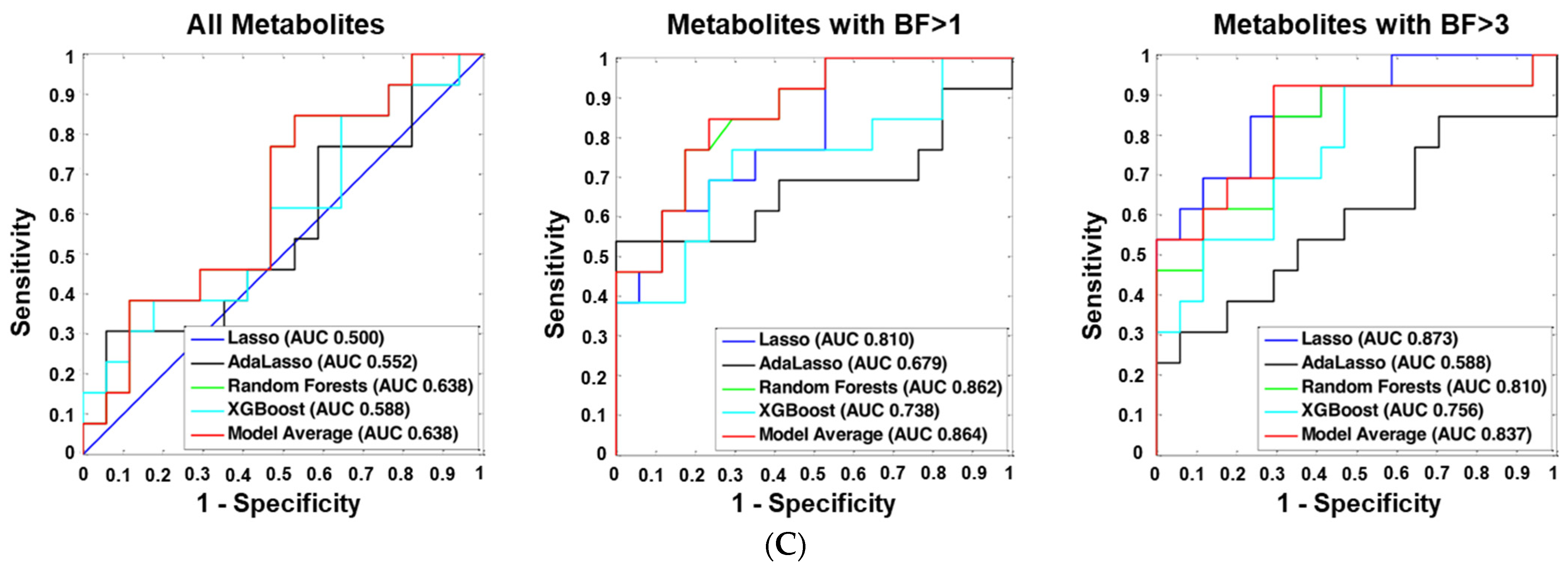
2.6. Machine Learning Analyses
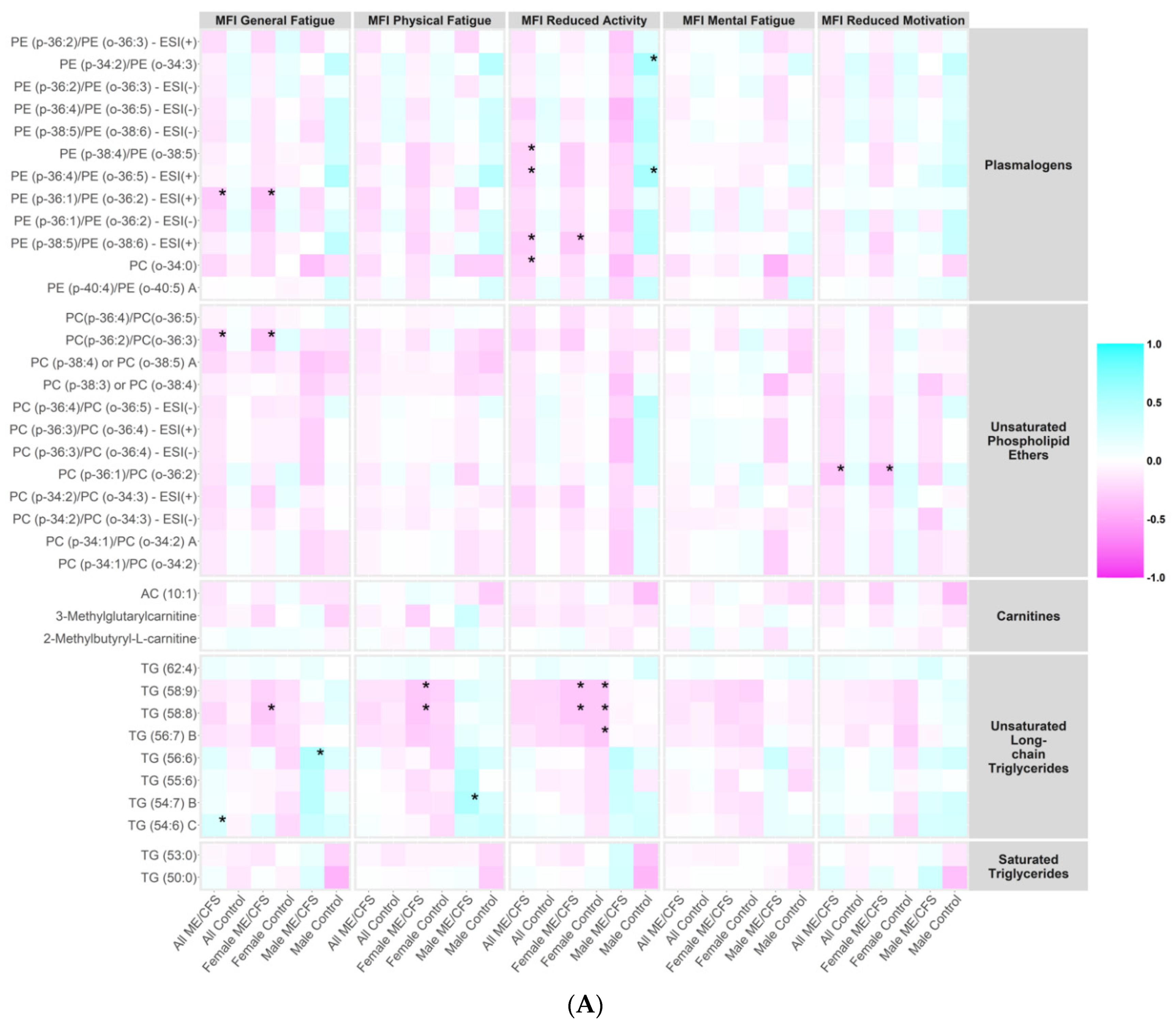
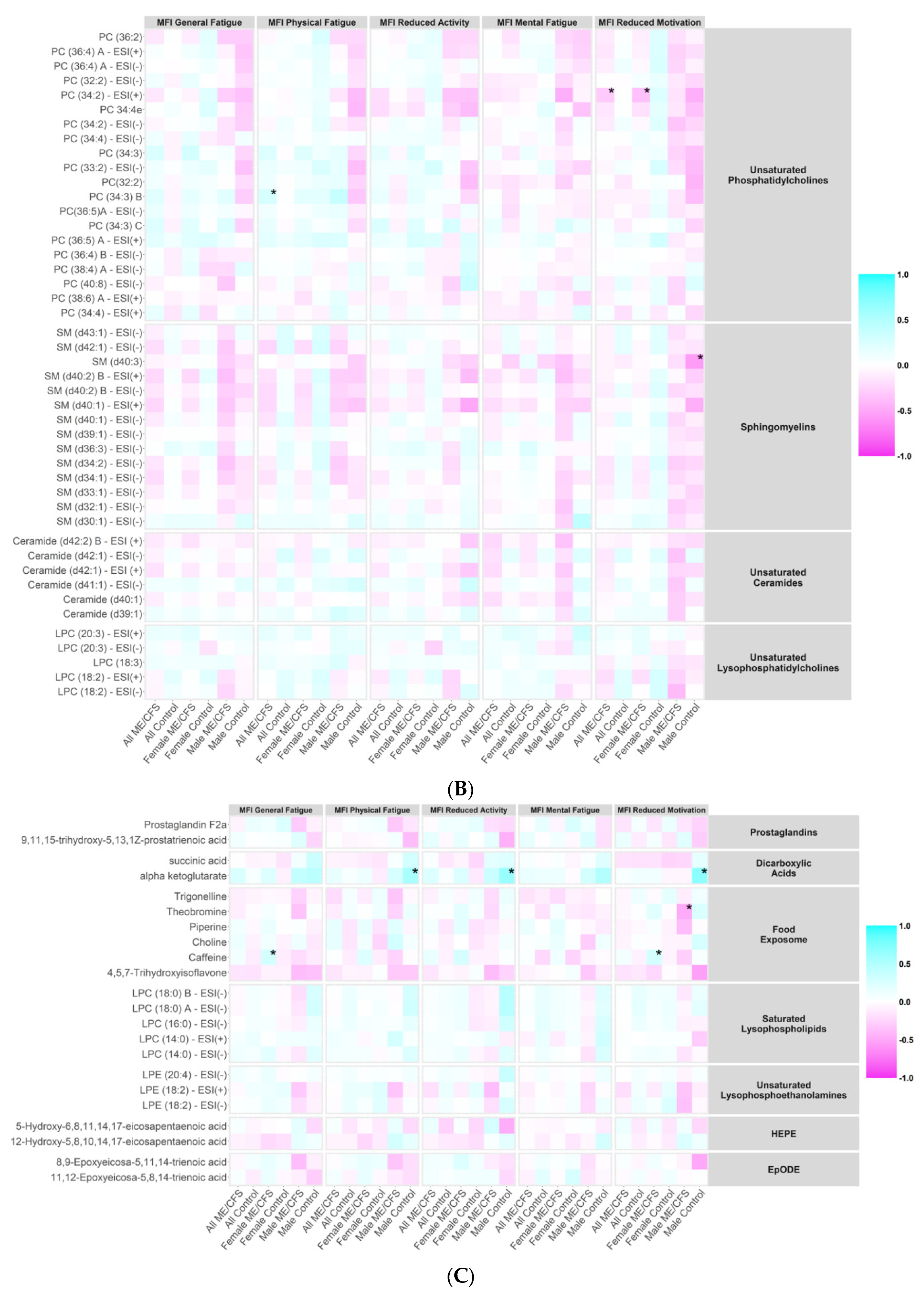
2.7. Correlations between Metabolites and ME/CFS Symptom Severity Scores
3. Discussion
3.1. Lipid Metabolism Abnormalities
3.2. TCA Cycle and Other Abnormalities
3.3. Metabolomic Findings as Biomarkers of Disease and of Disease Severity
3.4. Strengths and Limitations
4. Materials and Methods
4.1. Study Population
4.2. Plasma Collection
4.3. Clinical Assessment
4.4. Metabolomics Analysis
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Committee on the Diagnostic Criteria for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome; Board on the Health of Select Populations; Institute of Medicine. Beyond Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: Redefining an Illness. In The National Academies Collection: Reports Funded by National Institutes of Health; National Academies Press: Washington, DC, USA, 2015. [Google Scholar]
- Carruthers, B.M.; van de Sande, M.I.; De Meirleir, K.L.; Klimas, N.G.; Broderick, G.; Mitchell, T.; Staines, D.; Powles, A.C.; Speight, N.; Vallings, R.; et al. Myalgic encephalomyelitis: International Consensus Criteria. J. Intern. Med. 2011, 270, 327–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jason, L.A.; Mirin, A.A. Updating the National Academy of Medicine ME/CFS prevalence and economic impact figures to account for population growth and inflation. Fatigue Biomed. Health Behav. 2021, 9, 9–13. [Google Scholar] [CrossRef]
- Haney, E.; Smith, M.E.; McDonagh, M.; Pappas, M.; Daeges, M.; Wasson, N.; Nelson, H.D. Diagnostic Methods for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: A Systematic Review for a National Institutes of Health Pathways to Prevention Workshop. Ann. Intern. Med. 2015, 162, 834–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheibenbogen, C.; Freitag, H.; Blanco, J.; Capelli, E.; Lacerda, E.; Authier, J.; Meeus, M.; Castro Marrero, J.; Nora-Krukle, Z.; Oltra, E.; et al. The European ME/CFS Biomarker Landscape project: An initiative of the European network EUROMENE. J. Transl. Med. 2017, 15, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, C.W.; McGregor, N.R.; Lewis, D.P.; Butt, H.L.; Gooley, P.R. Metabolic profiling reveals anomalous energy metabolism and oxidative stress pathways in chronic fatigue syndrome patients. Metabolomics 2015, 11, 1626–1639. [Google Scholar] [CrossRef]
- Armstrong, C.W.; McGregor, N.R.; Lewis, D.P.; Butt, H.L.; Gooley, P.R. The association of fecal microbiota and fecal, blood serum and urine metabolites in myalgic encephalomyelitis/chronic fatigue syndrome. Metabolomics 2017, 13, 8. [Google Scholar] [CrossRef]
- Armstrong, C.W.; McGregor, N.R.; Sheedy, J.R.; Buttfield, I.; Butt, H.L.; Gooley, P.R. NMR metabolic profiling of serum identifies amino acid disturbances in chronic fatigue syndrome. Clin. Chim. Acta. 2012, 413, 1525–1531. [Google Scholar] [CrossRef]
- Fluge, O.; Mella, O.; Bruland, O.; Risa, K.; Dyrstad, S.E.; Alme, K.; Rekeland, I.G.; Sapkota, D.; Rosland, G.V.; Fossa, A.; et al. Metabolic profiling indicates impaired pyruvate dehydrogenase function in myalgic encephalopathy/chronic fatigue syndrome. JCI Insight 2016, 1, e89376. [Google Scholar] [CrossRef] [Green Version]
- Germain, A.; Barupal, D.K.; Levine, S.M.; Hanson, M.R. Comprehensive Circulatory Metabolomics in ME/CFS Reveals Disrupted Metabolism of Acyl Lipids and Steroids. Metabolites 2020, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Germain, A.; Ruppert, D.; Levine, S.M.; Hanson, M.R. Metabolic profiling of a myalgic encephalomyelitis/chronic fatigue syndrome discovery cohort reveals disturbances in fatty acid and lipid metabolism. Mol. Biosyst. 2017, 13, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Germain, A.; Ruppert, D.; Levine, S.M.; Hanson, M.R. Prospective Biomarkers from Plasma Metabolomics of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Implicate Redox Imbalance in Disease Symptomatology. Metabolites 2018, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGregor, N.R.; Armstrong, C.W.; Lewis, D.P.; Gooley, P.R. Post-Exertional Malaise Is Associated with Hypermetabolism, Hypoacetylation and Purine Metabolism Deregulation in ME/CFS Cases. Diagnostics 2019, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy-Szakal, D.; Barupal, D.K.; Lee, B.; Che, X.; Williams, B.L.; Kahn, E.J.R.; Ukaigwe, J.E.; Bateman, L.; Klimas, N.G.; Komaroff, A.L.; et al. Insights into myalgic encephalomyelitis/chronic fatigue syndrome phenotypes through comprehensive metabolomics. Sci. Rep. 2018, 8, 10056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naviaux, R.K.; Naviaux, J.C.; Li, K.; Bright, A.T.; Alaynick, W.A.; Wang, L.; Baxter, A.; Nathan, N.; Anderson, W.; Gordon, E. Metabolic features of chronic fatigue syndrome. Proc. Natl. Acad. Sci. USA 2016, 113, E5472–E5480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamano, E.; Sugimoto, M.; Hirayama, A.; Kume, S.; Yamato, M.; Jin, G.; Tajima, S.; Goda, N.; Iwai, K.; Fukuda, S.; et al. Index markers of chronic fatigue syndrome with dysfunction of TCA and urea cycles. Sci. Rep. 2016, 6, 34990. [Google Scholar] [CrossRef]
- Valdez, A.R.; Hancock, E.E.; Adebayo, S.; Kiernicki, D.J.; Proskauer, D.; Attewell, J.R.; Bateman, L.; DeMaria, A., Jr.; Lapp, C.W.; Rowe, P.C.; et al. Estimating Prevalence, Demographics, and Costs of ME/CFS Using Large Scale Medical Claims Data and Machine Learning. Front. Pediatr. 2018, 6, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milivojevic, M.; Che, X.; Bateman, L.; Cheng, A.; Garcia, B.A.; Hornig, M.; Huber, M.; Klimas, N.G.; Lee, B.; Lee, H.; et al. Plasma proteomic profiling suggests an association between antigen driven clonal B cell expansion and ME/CFS. PLoS ONE 2020, 15, e0236148. [Google Scholar] [CrossRef]
- Tomic, S.; Brkic, S.; Maric, D.; Mikic, A.N. Lipid and protein oxidation in female patients with chronic fatigue syndrome. Arch. Med. Sci. 2012, 8, 886–891. [Google Scholar] [CrossRef]
- Aaron, L.A.; Herrell, R.; Ashton, S.; Belcourt, M.; Schmaling, K.; Goldberg, J.; Buchwald, D. Comorbid clinical conditions in chronic fatigue: A co-twin control study. J. Gen. Intern. Med. 2001, 16, 24–31. [Google Scholar] [CrossRef]
- Giloteaux, L.; Goodrich, J.K.; Walters, W.A.; Levine, S.M.; Ley, R.E.; Hanson, M.R. Reduced diversity and altered composition of the gut microbiome in individuals with myalgic encephalomyelitis/chronic fatigue syndrome. Microbiome 2016, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Maes, M.; Bosmans, E.; Kubera, M. Increased expression of activation antigens on CD8+ T lymphocytes in Myalgic Encephalomyelitis/chronic fatigue syndrome: Inverse associations with lowered CD19+ expression and CD4+/CD8+ ratio, but no associations with (auto)immune, leaky gut, oxidative and nitrosative stress biomarkers. Neuro. Endocrinol. Lett. 2015, 36, 439–446. [Google Scholar] [PubMed]
- Nagy-Szakal, D.; Williams, B.L.; Mishra, N.; Che, X.; Lee, B.; Bateman, L.; Klimas, N.G.; Komaroff, A.L.; Levine, S.; Montoya, J.G.; et al. Fecal metagenomic profiles in subgroups of patients with myalgic encephalomyelitis/chronic fatigue syndrome. Microbiome 2017, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Makowski, D.; Ben-Shachar, M.S.; Chen, S.H.A.; Ludecke, D. Indices of Effect Existence and Significance in the Bayesian Framework. Front. Psychol. 2019, 10, 2767. [Google Scholar] [CrossRef] [PubMed]
- Jeffreys, H. Theory of Probability, 3rd ed.; Clarendon Press: Oxford, UK, 1961. [Google Scholar]
- Tibshirani, R. Regression Shrinkage and Selection Via the Lasso. J. R. Stat. Soc. Series B Stat. Methodol. 1996, 58, 267–288. [Google Scholar] [CrossRef]
- Zou, H. The Adaptive Lasso and Its Oracle Properties. J. Am. Stat. Assoc. 2012, 101, 1418–1429. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17August 2016; pp. 785–794. [Google Scholar]
- Hoel, F.; Hoel, A.; Pettersen, I.K.; Rekeland, I.G.; Risa, K.; Alme, K.; Sorland, K.; Fossa, A.; Lien, K.; Herder, I.; et al. A map of metabolic phenotypes in patients with myalgic encephalomyelitis/chronic fatigue syndrome. JCI Insight 2021, 6, e149217. [Google Scholar] [CrossRef]
- Messias, M.C.F.; Mecatti, G.C.; Priolli, D.G.; de Oliveira Carvalho, P. Plasmalogen lipids: Functional mechanism and their involvement in gastrointestinal cancer. Lipids Health Dis. 2018, 17, 41. [Google Scholar] [CrossRef] [Green Version]
- Wanders, R.J.; Poll-The, B.T. Role of peroxisomes in human lipid metabolism and its importance for neurological development. Neurosci. Lett. 2017, 637, 11–17. [Google Scholar] [CrossRef]
- Honsho, M.; Fujiki, Y. Plasmalogen homeostasis-regulation of plasmalogen biosynthesis and its physiological consequence in mammals. FEBS Lett. 2017, 591, 2720–2729. [Google Scholar] [CrossRef] [Green Version]
- Wanders, R.J.; Waterham, H.R.; Ferdinandusse, S. Metabolic Interplay between Peroxisomes and Other Subcellular Organelles Including Mitochondria and the Endoplasmic Reticulum. Front. Cell Dev. Biol. 2016, 3, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Missailidis, D.; Sanislav, O.; Allan, C.Y.; Smith, P.K.; Annesley, S.J.; Fisher, P.R. Dysregulated Provision of Oxidisable Substrates to the Mitochondria in ME/CFS Lymphoblasts. Int. J. Mol. Sci. 2021, 22, 2046. [Google Scholar] [CrossRef] [PubMed]
- Kujawski, S.; Cossington, J.; Slomko, J.; Zawadka-Kunikowska, M.; Tafil-Klawe, M.; Klawe, J.J.; Buszko, K.; Jakovljevic, D.G.; Kozakiewicz, M.; Morten, K.J.; et al. Relationship between Cardiopulmonary, Mitochondrial and Autonomic Nervous System Function Improvement after an Individualised Activity Programme upon Chronic Fatigue Syndrome Patients. J. Clin. Med. 2021, 10, 1542. [Google Scholar] [CrossRef]
- Flanagan, J.L.; Simmons, P.A.; Vehige, J.; Willcox, M.D.; Garrett, Q. Role of carnitine in disease. Nutr. Metab. 2010, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.L.; Wang, Q.Y.; Luan, H.Y.; Kang, Z.C.; Wang, C.B. Effects of L-carnitine against oxidative stress in human hepatocytes: Involvement of peroxisome proliferator-activated receptor alpha. J. Biomed. Sci. 2012, 19, 32. [Google Scholar] [CrossRef] [Green Version]
- Vacha, G.M.; Giorcelli, G.; Siliprandi, N.; Corsi, M. Favorable effects of L-carnitine treatment on hypertriglyceridemia in hemodialysis patients: Decisive role of low levels of high-density lipoprotein-cholesterol. Am. J. Clin. Nutr. 1983, 38, 532–540. [Google Scholar] [CrossRef] [Green Version]
- Violante, S.; Ijlst, L.; Te Brinke, H.; Koster, J.; Tavares de Almeida, I.; Wanders, R.J.; Ventura, F.V.; Houten, S.M. Peroxisomes contribute to the acylcarnitine production when the carnitine shuttle is deficient. Biochim. Biophys. Acta. 2013, 1831, 1467–1474. [Google Scholar] [CrossRef]
- Demarquoy, J.; Le Borgne, F. Crosstalk between mitochondria and peroxisomes. World J. Biol. Chem. 2015, 6, 301–309. [Google Scholar] [CrossRef]
- Paul, B.D.; Lemle, M.D.; Komaroff, A.L.; Snyder, S.H. Redox imbalance links COVID-19 and myalgic encephalomyelitis/chronic fatigue syndrome. Proc. Natl. Acad. Sci. USA. 2021, 118, e2024358118. [Google Scholar] [CrossRef]
- Sperka-Gottlieb, C.D.; Hermetter, A.; Paltauf, F.; Daum, G. Lipid topology and physical properties of the outer mitochondrial membrane of the yeast, Saccharomyces cerevisiae. Biochim. Biophys. Acta. 1988, 946, 227–234. [Google Scholar] [CrossRef]
- Zinser, E.; Sperka-Gottlieb, C.D.; Fasch, E.V.; Kohlwein, S.D.; Paltauf, F.; Daum, G. Phospholipid synthesis and lipid composition of subcellular membranes in the unicellular eukaryote Saccharomyces cerevisiae. J. Bacteriol. 1991, 173, 2026–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibellini, F.; Smith, T.K. The Kennedy pathway--De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB Life. 2010, 62, 414–428. [Google Scholar] [CrossRef] [PubMed]
- Birner, R.; Burgermeister, M.; Schneiter, R.; Daum, G. Roles of phosphatidylethanolamine and of its several biosynthetic pathways in Saccharomyces cerevisiae. Mol. Biol. Cell. 2001, 12, 997–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowhan, W.; Bogdanov, M. Lipid-dependent membrane protein topogenesis. Annu. Rev. Biochem. 2009, 78, 515–540. [Google Scholar] [CrossRef] [Green Version]
- Furt, F.; Moreau, P. Importance of lipid metabolism for intracellular and mitochondrial membrane fusion/fission processes. Int. J. Biochem. Cell Biol. 2009, 41, 1828–1836. [Google Scholar] [CrossRef]
- Schuler, M.H.; Di Bartolomeo, F.; Martensson, C.U.; Daum, G.; Becker, T. Phosphatidylcholine Affects Inner Membrane Protein Translocases of Mitochondria. J. Biol. Chem. 2016, 291, 18718–18729. [Google Scholar] [CrossRef] [Green Version]
- Schuler, M.H.; Di Bartolomeo, F.; Bottinger, L.; Horvath, S.E.; Wenz, L.S.; Daum, G.; Becker, T. Phosphatidylcholine affects the role of the sorting and assembly machinery in the biogenesis of mitochondrial beta-barrel proteins. J. Biol. Chem. 2015, 290, 26523–26532. [Google Scholar] [CrossRef] [Green Version]
- Yoda, E.; Hachisu, K.; Taketomi, Y.; Yoshida, K.; Nakamura, M.; Ikeda, K.; Taguchi, R.; Nakatani, Y.; Kuwata, H.; Murakami, M.; et al. Mitochondrial dysfunction and reduced prostaglandin synthesis in skeletal muscle of Group VIB Ca2+-independent phospholipase A2gamma-deficient mice. J. Lipid. Res. 2010, 51, 3003–3015. [Google Scholar] [CrossRef] [Green Version]
- Dawaliby, R.; Trubbia, C.; Delporte, C.; Masureel, M.; Van Antwerpen, P.; Kobilka, B.K.; Govaerts, C. Allosteric regulation of G protein-coupled receptor activity by phospholipids. Nat. Chem. Biol. 2016, 12, 35–39. [Google Scholar] [CrossRef]
- Li, Z.; Vance, D.E. Phosphatidylcholine and choline homeostasis. J. Lipid Res. 2008, 49, 1187–1194. [Google Scholar] [CrossRef] [Green Version]
- Momchilova, A.; Markovska, T. Phosphatidylethanolamine and phosphatidylcholine are sources of diacylglycerol in ras-transformed NIH 3T3 fibroblasts. Int. J. Biochem. Cell Biol. 1999, 31, 311–318. [Google Scholar] [CrossRef]
- Okamoto, Y.; Morishita, J.; Tsuboi, K.; Tonai, T.; Ueda, N. Molecular characterization of a phospholipase D generating anandamide and its congeners. J. Biol. Chem. 2004, 279, 5298–5305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathias, S.; Kolesnick, R. Ceramide: A novel second messenger. Adv. Lipid Res. 1993, 25, 65–90. [Google Scholar] [PubMed]
- Cabral-Marques, O.; Marques, A.; Giil, L.M.; De Vito, R.; Rademacher, J.; Gunther, J.; Lange, T.; Humrich, J.Y.; Klapa, S.; Schinke, S.; et al. GPCR-specific autoantibody signatures are associated with physiological and pathological immune homeostasis. Nat. Commun. 2018, 9, 5224. [Google Scholar] [CrossRef]
- Loebel, M.; Grabowski, P.; Heidecke, H.; Bauer, S.; Hanitsch, L.G.; Wittke, K.; Meisel, C.; Reinke, P.; Volk, H.D.; Fluge, O.; et al. Antibodies to beta adrenergic and muscarinic cholinergic receptors in patients with Chronic Fatigue Syndrome. Brain Behav. Immun. 2016, 52, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Wirth, K.; Scheibenbogen, C. A Unifying Hypothesis of the Pathophysiology of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): Recognitions from the finding of autoantibodies against ss2-adrenergic receptors. Autoimmun. Rev. 2020, 19, 102527. [Google Scholar] [CrossRef]
- Martinez-Reyes, I.; Chandel, N.S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, M.; Zhu, W.; Yu, J.; Wang, Q.; Zhang, J.; Cui, Y.; Pan, X.; Gao, X.; Sun, H. Succinate accumulation induces mitochondrial reactive oxygen species generation and promotes status epilepticus in the kainic acid rat model. Redox Biol. 2020, 28, 101365. [Google Scholar] [CrossRef]
- Palmieri, E.M.; Gonzalez-Cotto, M.; Baseler, W.A.; Davies, L.C.; Ghesquiere, B.; Maio, N.; Rice, C.M.; Rouault, T.A.; Cassel, T.; Higashi, R.M.; et al. Nitric oxide orchestrates metabolic rewiring in M1 macrophages by targeting aconitase 2 and pyruvate dehydrogenase. Nat. Commun. 2020, 11, 698. [Google Scholar] [CrossRef]
- Morris, G.; Berk, M.; Klein, H.; Walder, K.; Galecki, P.; Maes, M. Nitrosative Stress, Hypernitrosylation, and Autoimmune Responses to Nitrosylated Proteins: New Pathways in Neuroprogressive Disorders Including Depression and Chronic Fatigue Syndrome. Mol. Neurobiol. 2017, 54, 4271–4291. [Google Scholar] [CrossRef]
- Morris, G.; Maes, M. Mitochondrial dysfunctions in myalgic encephalomyelitis/chronic fatigue syndrome explained by activated immuno-inflammatory, oxidative and nitrosative stress pathways. Metab. Brain Dis. 2014, 29, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, B.D.; Marinello, M.; Heinz, J.; Rymut, N.; Sansbury, B.E.; Riley, C.O.; Sadhu, S.; Hosseini, Z.; Kojima, Y.; Tang, D.D.; et al. Resolvin D1 promotes the targeting and clearance of necroptotic cells. Cell Death Differ. 2020, 27, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Komaroff, A.L. Inflammation correlates with symptoms in chronic fatigue syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, 8914–8916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatomi, Y.; Kuratsune, H.; Watanabe, Y. Neuroinflammation in the Brain of Patients with Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Brain Nerve 2018, 70, 19–25. [Google Scholar] [PubMed]
- Fukuda, K.; Straus, S.E.; Hickie, I.; Sharpe, M.C.; Dobbins, J.G.; Komaroff, A. The chronic fatigue syndrome: A comprehensive approach to its definition and study. International Chronic Fatigue Syndrome Study Group. Ann. Intern. Med. 1994, 121, 953–959. [Google Scholar] [CrossRef]
- Carruthers, B.M.; Jain, A.K.; De Meirleir, K.L.; Peterson, D.L.; Klimas, N.G.; Lerner, A.M.; Bested, A.C.; Flor-Henry, P.; Joshi, P.; Powles, A.C.P.; et al. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. J. Chronic Fatigue Syndr. 2003, 11, 7–115. [Google Scholar] [CrossRef]
- Jason, L.A.; Evans, M.; Porter, N.; Brown, M.; Brown, A.; Hunnell, J.; Anderson, V.; Lerch, A.; De Meirleir, K.; Friedberg, F. The Development of a Revised Canadian Myalgic Encephalomyelitis Chronic Fatigue Syndrome Case Definition. Am. J. Biochem. Biotechnol. 2010, 6, 120–135. [Google Scholar] [CrossRef] [Green Version]
- Buysse, D.J.; Reynolds, C.F., 3rd; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh Sleep Quality Index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Ware, J.E., Jr.; Sherbourne, C.D. The MOS 36-item short-form health survey (SF-36). I. Conceptual framework and item selection. Med. Care. 1992, 30, 473–483. [Google Scholar] [CrossRef]
- Smets, E.M.; Garssen, B.; Bonke, B.; De Haes, J.C. The Multidimensional Fatigue Inventory (MFI) psychometric qualities of an instrument to assess fatigue. J. Psychosom. Res. 1995, 39, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spectrometry: Combined Targeted and Untargeted Profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30.4.1–30.4.32. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Wohlgemuth, G.; Lee, D.Y.; Lu, Y.; Palazoglu, M.; Shahbaz, S.; Fiehn, O. FiehnLib: Mass spectral and retention index libraries for metabolomics based on quadrupole and time-of-flight gas chromatography/mass spectrometry. Anal. Chem. 2009, 81, 10038–10048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cajka, T.; Smilowitz, J.T.; Fiehn, O. Validating Quantitative Untargeted Lipidomics Across Nine Liquid Chromatography-High-Resolution Mass Spectrometry Platforms. Anal. Chem. 2017, 89, 12360–12368. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Kind, T.; Liu, K.H.; Lee, D.Y.; DeFelice, B.; Meissen, J.K.; Fiehn, O. LipidBlast in silico tandem mass spectrometry database for lipid identification. Nat. Methods 2013, 10, 755–758. [Google Scholar] [CrossRef] [Green Version]
- Bakovic, M.; Fullerton, M.D.; Michel, V. Metabolic and molecular aspects of ethanolamine phospholipid biosynthesis: The role of CTP:phosphoethanolamine cytidylyltransferase (Pcyt2). Biochem. Cell Biol. 2007, 85, 283–300. [Google Scholar] [CrossRef]
- DeFelice, B.C.; Mehta, S.S.; Samra, S.; Cajka, T.; Wancewicz, B.; Fahrmann, J.F.; Fiehn, O. Mass Spectral Feature List Optimizer (MS-FLO): A Tool To Minimize False Positive Peak Reports in Untargeted Liquid Chromatography-Mass Spectroscopy (LC-MS) Data Processing. Anal. Chem. 2017, 89, 3250–3255. [Google Scholar] [CrossRef]
- Fan, S.; Kind, T.; Cajka, T.; Hazen, S.L.; Tang, W.H.W.; Kaddurah-Daouk, R.; Irvin, M.R.; Arnett, D.K.; Barupal, D.K.; Fiehn, O. Systematic Error Removal Using Random Forest for Normalizing Large-Scale Untargeted Lipidomics Data. Anal. Chem. 2019, 91, 3590–3596. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate-A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Series B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Barupal, D.K.; Fiehn, O. Chemical Similarity Enrichment Analysis (ChemRICH) as alternative to biochemical pathway mapping for metabolomic datasets. Sci. Rep. 2017, 7, 14567. [Google Scholar] [CrossRef]
- Goodrich, B.; Gabry, J.; Ali, I.; Brilleman, S. Rstanarm: Bayesian Applied Regression Modeling via Stan. Available online: mc-stan.org/rstanarm/ (accessed on 13 April 2021).
- Makowski, D.; Ben-Shachar, M.S.; Lüdecke, D. bayestestR: Describing Effects and their Uncertainty, Existence and Significance within the Bayseian Framework. J. Open Source Softw. 2019, 4, 1541. [Google Scholar] [CrossRef]
- Gelman, A.; Jakulin, A.; Pittau, M.G.; Su, Y.-S. A weakly informative default prior distribution for logistic and other regression models. Ann. Appl. Stat. 2008, 2, 1360–1383. [Google Scholar] [CrossRef]
- Muth, C.; Oravecz, Z.; Gabry, J. User-friendly Bayesian regression modeling: A tutorial with rstanarm and shinystan. Quant. Meth. Psych. 2018, 14, 99–119. [Google Scholar] [CrossRef]
- Fan, J.; Li, R. Variable Selection via Nonconcave Penalized Likelihood and its Oracle Properties. J. Am. Stat. Assoc. 2001, 96, 1348–1360. [Google Scholar] [CrossRef]
- Hoeting, J.A.; Madigan, D.; Raftery, A.E.; Volinsky, C.T. Bayesian Model Averaging: A Tutorial. Stat. Sci. 1999, 14, 382–401. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject Characteristics | ME/CFS (n = 106) | Controls (n = 91) | p-Value * | |
|---|---|---|---|---|
| Sex | Female | 75 | 69 | 0.42 |
| Male | 31 | 22 | ||
| Age | Mean ± SD | 47.8 ± 13.7 | 47.0 ± 14.1 | 0.78 |
| Median (Range) | 51.0 (21.6–70.0) | 50.6 (21.2–68.2) | ||
| Race and Ethnicity | White and not Hispanic | 93 | 85 | 0.40 |
| Hispanic | 6 | 3 | ||
| Not White and not Hispanic | 7 | 3 | ||
| Site of Collection | Incline Village, NV | 23 | 17 | 0.30 |
| Miami, FL | 15 | 5 | ||
| New York, NY | 17 | 19 | ||
| Salt Lake City, UT | 32 | 32 | ||
| Palo Alto, CA | 19 | 19 | ||
| Season of Collection | Winter | 33 | 14 | 0.004 |
| Spring | 72 | 70 | ||
| Summer | 1 | 7 | ||
| sr-IBS Comorbidity † | Yes | 35 | 3 | <0.001 |
| No | 71 | 88 | ||
| BMI | Mean ± SD | 26.1 ± 5.2 | 25.2 ± 4.7 | 0.31 |
| Median (Range) | 25.0 (18.1–41.2) | 25.1 (16.9–38.7) | ||
| Duration of ME/CFS ‡ | Mean ± SD | 15.0 ± 9.8 | n/a | n/a |
| Median (Range) | 14.4 (1.2–44.2) | n/a | ||
| ≥3 years | 92 | n/a | ||
| <3 years | 8 | n/a | ||
| SF-36 Scales § Median (IQR) | Emotional Limitations | 83.3 (0.0–100.0) | 100.0 (100.0–100.0) | <0.001 |
| Emotional Well-being | 72.0 (56.0–84.0) | 88.0 (80.0–92.0) | <0.001 | |
| General Health | 20.0 (15.0–30.0) | 90.0 (75.0–95.0) | <0.001 | |
| Pain | 45.0 (22.5–66.25) | 90.0 (90.0–100.0) | <0.001 | |
| Physical Functioning | 35.0 (20.0–55.0) | 100.0 (95.0–100.0) | <0.001 | |
| Physical Limitations | 0.0 (0.0–0.0) | 100.0 (100.0–100.0) | <0.001 | |
| Social Functioning | 22.5 (10.0–45.0) | 100.0 (90.0–100.0) | <0.001 | |
| Vitality | 5.0 (0.0–20.0) | 80.0 (70.0–85.0) | <0.001 | |
| MFI Scales || Median (IQR) | General Fatigue | 91.7 (71.9–100.0) | 16.7 (4.2–33.3) | <0.001 |
| Mental Fatigue | 62.5 (45.8–75.0) | 12.5 (0.0–35.4) | <0.001 | |
| Physical Fatigue | 87.5 (66.7–100.0) | 12.5 (4.2–20.8) | <0.001 | |
| Reduced Activity | 79.2 (58.3–95.8) | 4.2 (2.1–29.2) | <0.001 | |
| Reduced Motivation | 45.8 (29.2–66.7) | 12.5 (4.2–25.0) | <0.001 | |
| Metabolite | Enrichment Cluster | Regression Model | ME/CFS vs. Control | ||||
|---|---|---|---|---|---|---|---|
| Estimated Coefficient | 95% CI | p-Value | FDR | Bayes Factor | |||
| Biogenic Amines (BA) | |||||||
| Acetominophen | drugs | Lognormal | 0.068 | (0.028, 0.108) | 0.001 | 0.103 | 3.035 |
| Complex Lipids (CL) | |||||||
| PE (p-36:2)/PE (o-36:3)—ESI(+) | plasmalogens | Lognormal | −0.028 | (−0.043, −0.013) | 0.000 | 0.074 | 20.935 |
| PE (p-34:2)/PE (o-34:3) | plasmalogens | Lognormal | −0.037 | (−0.060, −0.014) | 0.002 | 0.126 | 5.662 |
| LPC (18:2)—ESI(−) | unsaturated LPC | Lognormal | −0.019 | (−0.032, −0.007) | 0.003 | 0.139 | 4.102 |
| PC (36:2) | unsaturated PC | Lognormal | −0.007 | (−0.011, −0.003) | 0.000 | 0.074 | 11.241 |
| PC (36:4) A—ESI(+) | unsaturated PC | Lognormal | −0.018 | (−0.028, −0.008) | 0.000 | 0.074 | 8.134 |
| PC (36:4) A—ESI(−) | unsaturated PC | Lognormal | −0.019 | (−0.031, −0.008) | 0.001 | 0.103 | 4.032 |
| PC (32:2)—ESI(−) | unsaturated PC | Lognormal | −0.027 | (−0.043, −0.010) | 0.002 | 0.135 | 7.389 |
| PC 34:4e | unsaturated PC | Lognormal | −0.022 | (−0.036, −0.008) | 0.003 | 0.139 | 4.327 |
| PC (p-34:2)/PC (o-34:3)—ESI(+) | unsaturated PLE | Lognormal | −0.018 | (−0.027, −0.009) | 0.000 | 0.062 | 44.620 |
| PC (p-34:1)/PC (o-34:2) | unsaturated PLE | Lognormal | −0.021 | (−0.032, −0.010) | 0.000 | 0.062 | 178.678 |
| PC (p-36:1)/PC (o-36:2) | unsaturated PLE | Lognormal | −0.055 | (−0.086, −0.024) | 0.001 | 0.074 | 11.555 |
| PC (p-34:2)/PC (o-34:3)—ESI(−) | unsaturated PLE | Lognormal | −0.020 | (−0.032, −0.009) | 0.001 | 0.074 | 12.281 |
| PC (p-36:4)/PC (o-36:5)—ESI(−) | unsaturated PLE | Lognormal | −0.021 | (−0.034, −0.009) | 0.001 | 0.103 | 7.046 |
| PC (p-34:1)/PC (o-34:2) A | unsaturated PLE | Lognormal | −0.027 | (−0.044, −0.011) | 0.002 | 0.125 | 5.655 |
| Oxylipins (OL) | |||||||
| Resolvin D1 | OH-FA_22_6_1 | Gamma | −0.528 | (−0.846, −0.210) | 0.002 | 0.134 | 6.635 |
| Metabolite | Enrichment Cluster | Regression Model | Female ME/CFS vs. Female Control | ||||
| Estimated Coefficient | 95% CI | p-Value | FDR | Bayes Factor | |||
| Biogenic Amines (BA) | |||||||
| Alprazolam | drugs | Lognormal | 0.081 | (0.030, 0.132) | 0.002 | 0.121 | 3.486 |
| Acyclovir | drugs | Lognormal | 0.152 | (0.057, 0.247) | 0.002 | 0.121 | 3.179 |
| Complex Lipids (CL) | |||||||
| PE (p-36:2)/PE (o-36:3)—ESI(+) | plasmalogens | Lognormal | −0.033 | (−0.049, −0.017) | 0.000 | 0.048 | 24.602 |
| PE (p-34:2)/PE (o-34:3) | plasmalogens | Lognormal | −0.042 | (−0.066, −0.018) | 0.001 | 0.064 | 6.155 |
| SM (d40:3) | sphingomyelins | Lognormal | −0.035 | (−0.055, −0.014) | 0.001 | 0.064 | 6.392 |
| PC (36:2) | unsaturated PC | Lognormal | −0.009 | (−0.013, −0.004) | 0.000 | 0.054 | 14.972 |
| PC (36:4) A—ESI(+) | unsaturated PC | Lognormal | −0.022 | (−0.033, −0.010) | 0.000 | 0.054 | 8.061 |
| PC (36:4) A—ESI(−) | unsaturated PC | Lognormal | −0.025 | (−0.038, −0.011) | 0.000 | 0.054 | 11.432 |
| PC 34:4e | unsaturated PC | Lognormal | −0.029 | (−0.045, −0.014) | 0.000 | 0.054 | 13.135 |
| PC (34:2)—ESI(+) | unsaturated PC | Lognormal | −0.006 | (−0.010, −0.003) | 0.000 | 0.054 | 6.913 |
| PC (p-34:2)/PC (o-34:3)—ESI(+) | unsaturated PLE | Lognormal | −0.022 | (−0.032, −0.011) | 0.000 | 0.048 | 36.107 |
| PC (p-34:1)/PC (o-34:2) | unsaturated PLE | Lognormal | −0.025 | (−0.038, −0.012) | 0.000 | 0.054 | 26.013 |
| PC (p-36:1)/PC (o-36:2) | unsaturated PLE | Lognormal | −0.067 | (−0.106, −0.029) | 0.001 | 0.064 | 5.458 |
| PC (p-34:2)/PC (o-34:3)—ESI(−) | unsaturated PLE | Lognormal | −0.025 | (−0.038, −0.011) | 0.001 | 0.054 | 7.542 |
| PC (p-36:4)/PC (o-36:5)—ESI(−) | unsaturated PLE | Lognormal | −0.025 | (−0.040, −0.011) | 0.001 | 0.064 | 7.044 |
| PC (p-34:1)/PC (o-34:2) A | unsaturated PLE | Lognormal | −0.035 | (−0.055, −0.015) | 0.001 | 0.064 | 6.285 |
| Metabolite | Enrichment Cluster | Regression Model | ME/CFS without sr-IBS vs. Control without sr-IBS | ||||
| Estimated Coefficient | 95% CI | p-Value | FDR | Bayes Factor | |||
| Complex Lipids (CL) | |||||||
| PE (p-36:2)/PE (o-36:3)—ESI(+) | plasmalogens | Lognormal | −0.029 | (−0.045, −0.013) | 0.000 | 0.081 | 6.915 |
| PC (36:2) | unsaturated PC | Lognormal | −0.008 | (−0.012, −0.004) | 0.000 | 0.076 | 8.626 |
| PC (p-34:2)/PC (o-34:3)—ESI(+) | unsaturated PLE | Lognormal | −0.019 | (−0.028, −0.010) | 0.000 | 0.076 | 16.961 |
| PC (p-34:1)/PC (o-34:2) | unsaturated PLE | Lognormal | −0.022 | (−0.033, −0.010) | 0.000 | 0.076 | 30.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Che, X.; Brydges, C.R.; Yu, Y.; Price, A.; Joshi, S.; Roy, A.; Lee, B.; Barupal, D.K.; Cheng, A.; Palmer, D.M.; et al. Metabolomic Evidence for Peroxisomal Dysfunction in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Int. J. Mol. Sci. 2022, 23, 7906. https://doi.org/10.3390/ijms23147906
Che X, Brydges CR, Yu Y, Price A, Joshi S, Roy A, Lee B, Barupal DK, Cheng A, Palmer DM, et al. Metabolomic Evidence for Peroxisomal Dysfunction in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. International Journal of Molecular Sciences. 2022; 23(14):7906. https://doi.org/10.3390/ijms23147906
Chicago/Turabian StyleChe, Xiaoyu, Christopher R. Brydges, Yuanzhi Yu, Adam Price, Shreyas Joshi, Ayan Roy, Bohyun Lee, Dinesh K. Barupal, Aaron Cheng, Dana March Palmer, and et al. 2022. "Metabolomic Evidence for Peroxisomal Dysfunction in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome" International Journal of Molecular Sciences 23, no. 14: 7906. https://doi.org/10.3390/ijms23147906
APA StyleChe, X., Brydges, C. R., Yu, Y., Price, A., Joshi, S., Roy, A., Lee, B., Barupal, D. K., Cheng, A., Palmer, D. M., Levine, S., Peterson, D. L., Vernon, S. D., Bateman, L., Hornig, M., Montoya, J. G., Komaroff, A. L., Fiehn, O., & Lipkin, W. I. (2022). Metabolomic Evidence for Peroxisomal Dysfunction in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. International Journal of Molecular Sciences, 23(14), 7906. https://doi.org/10.3390/ijms23147906