
Phytohormones and Transcriptome Analyses Revealed the Dynamics Involved in Spikelet Abortion and Inflorescence Development in Rice

 ,
,  ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
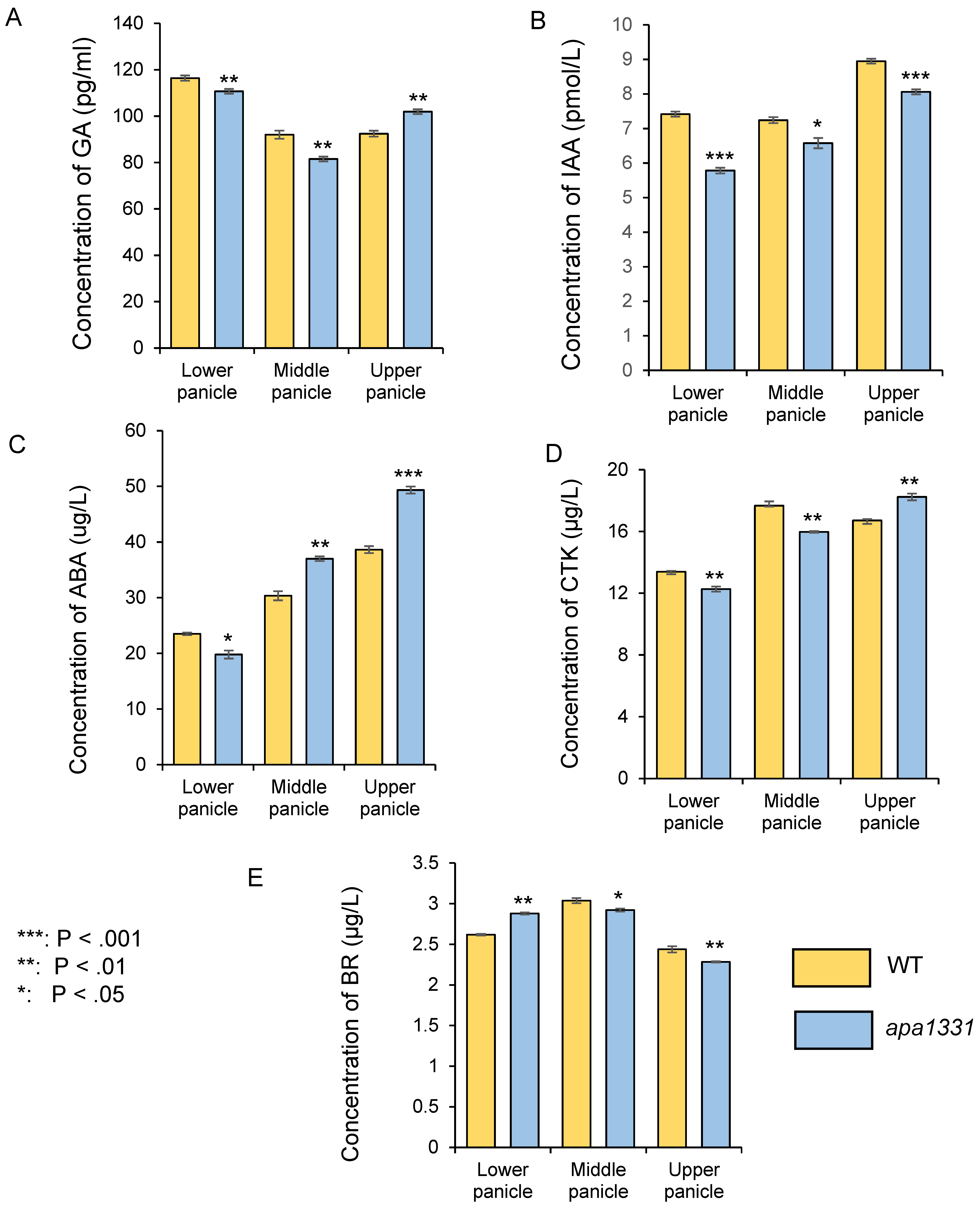
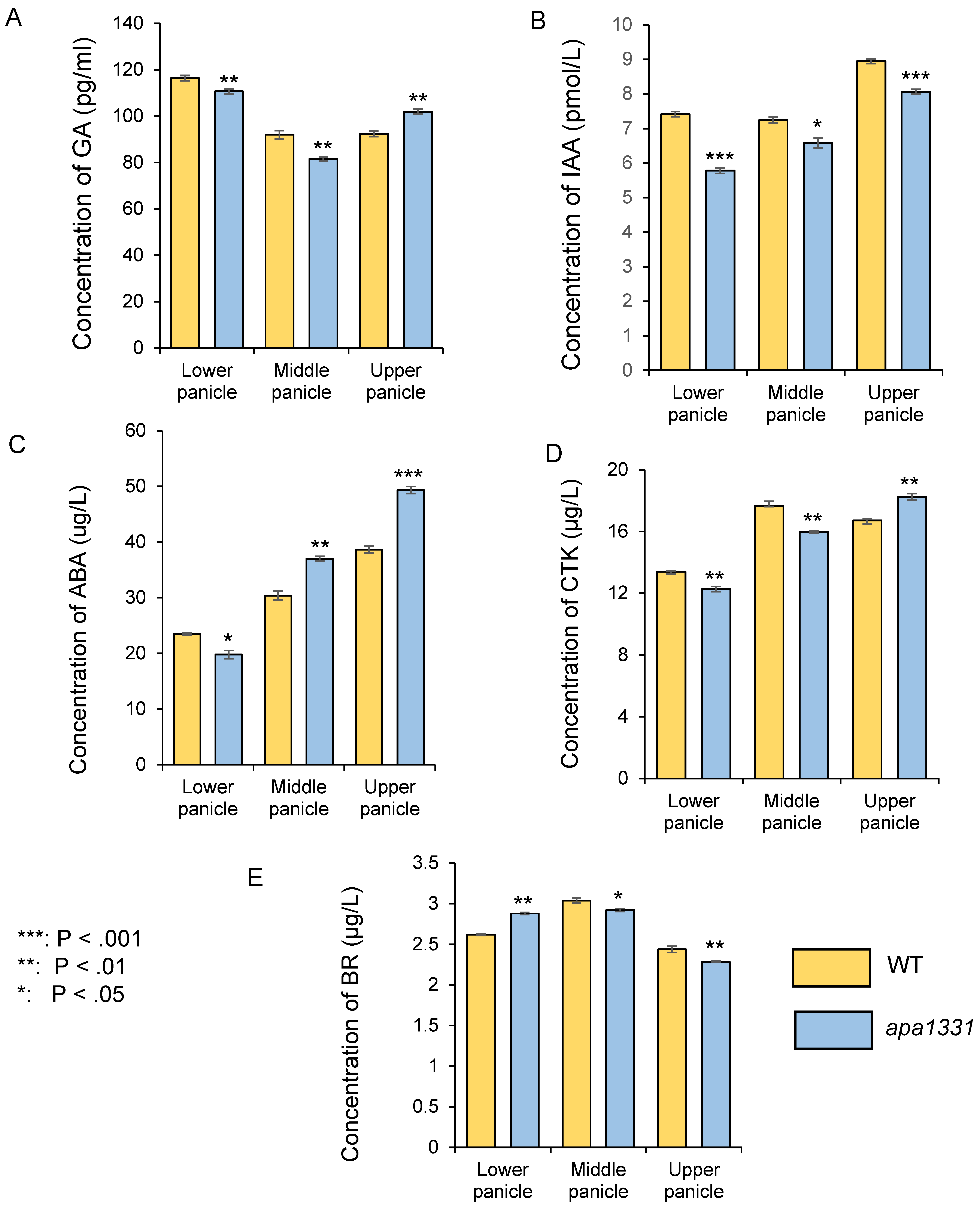
2.1. Apical Spikelets of apa1331 Showed a Differential Load of Phytohormones, Especially IAA and ABA
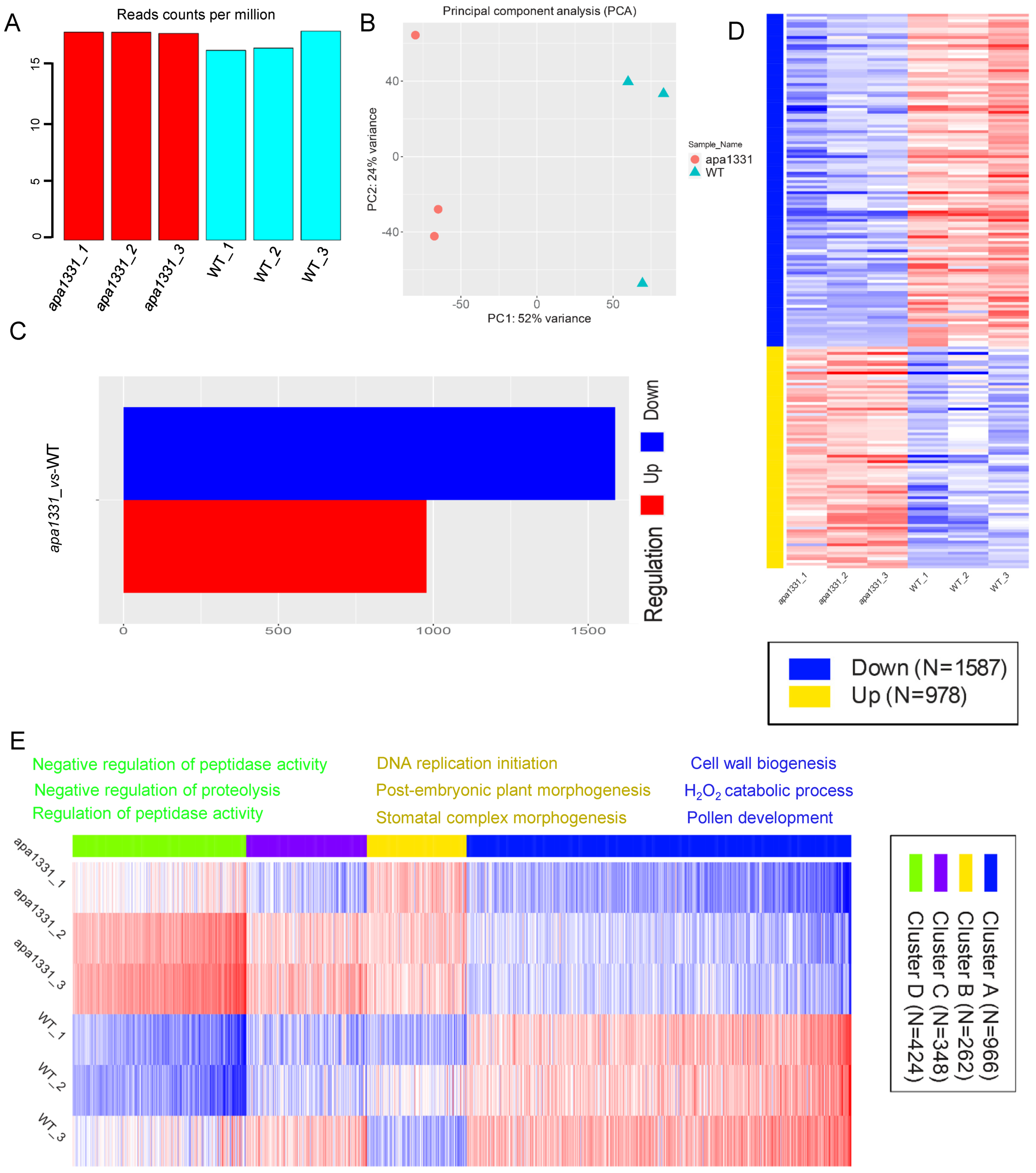
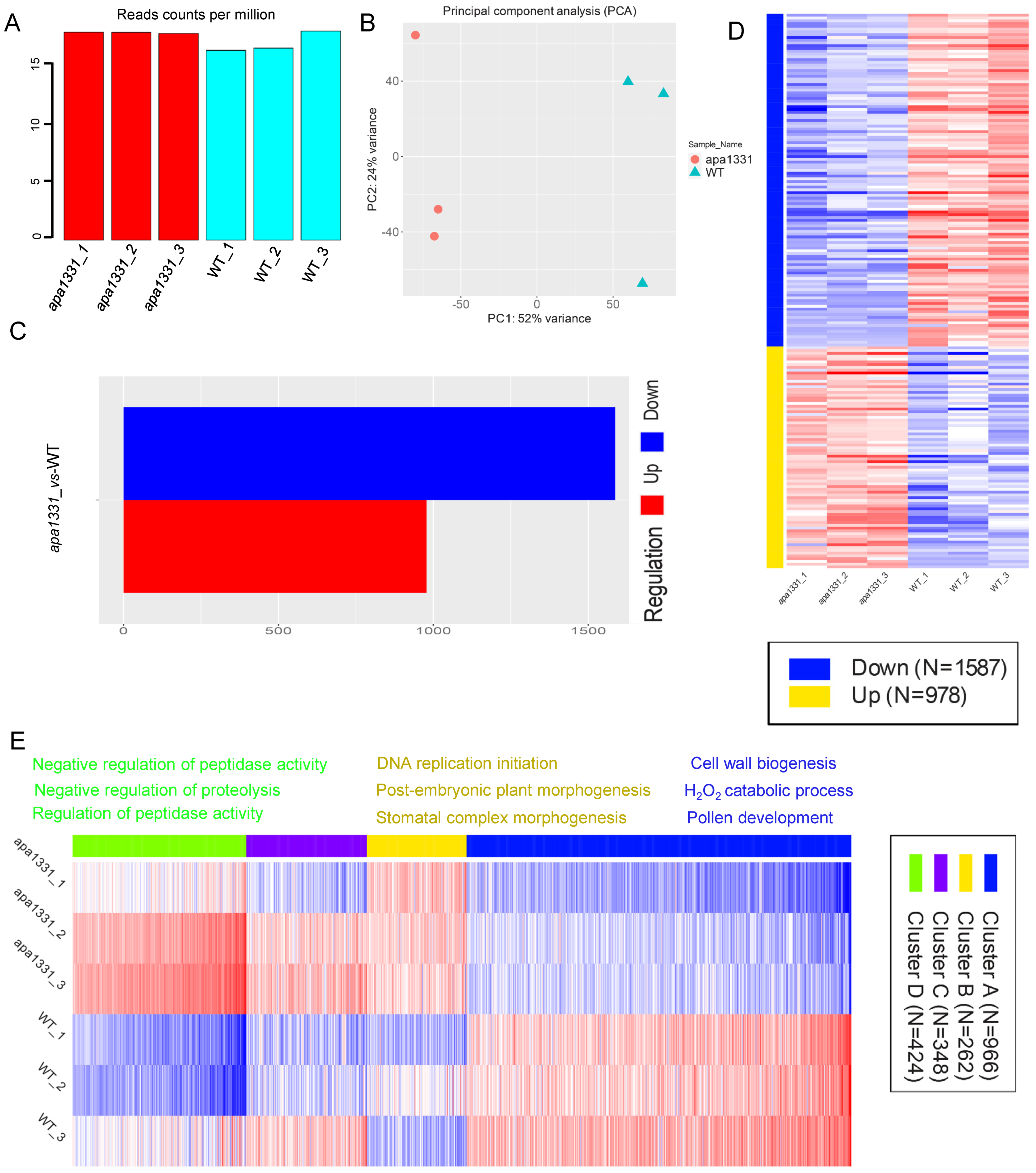
2.2. Identification of DEGs in WT and apa1331
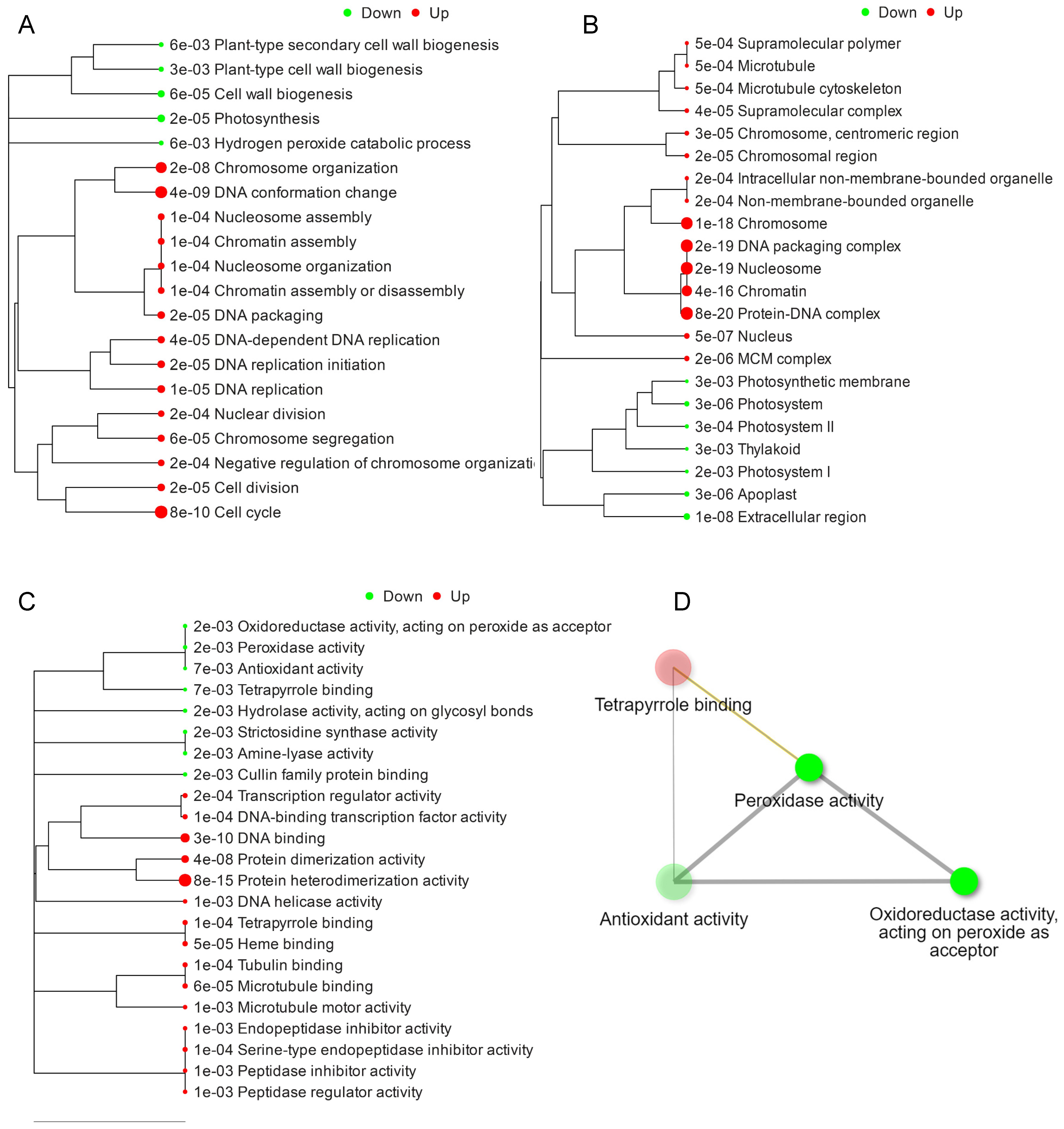
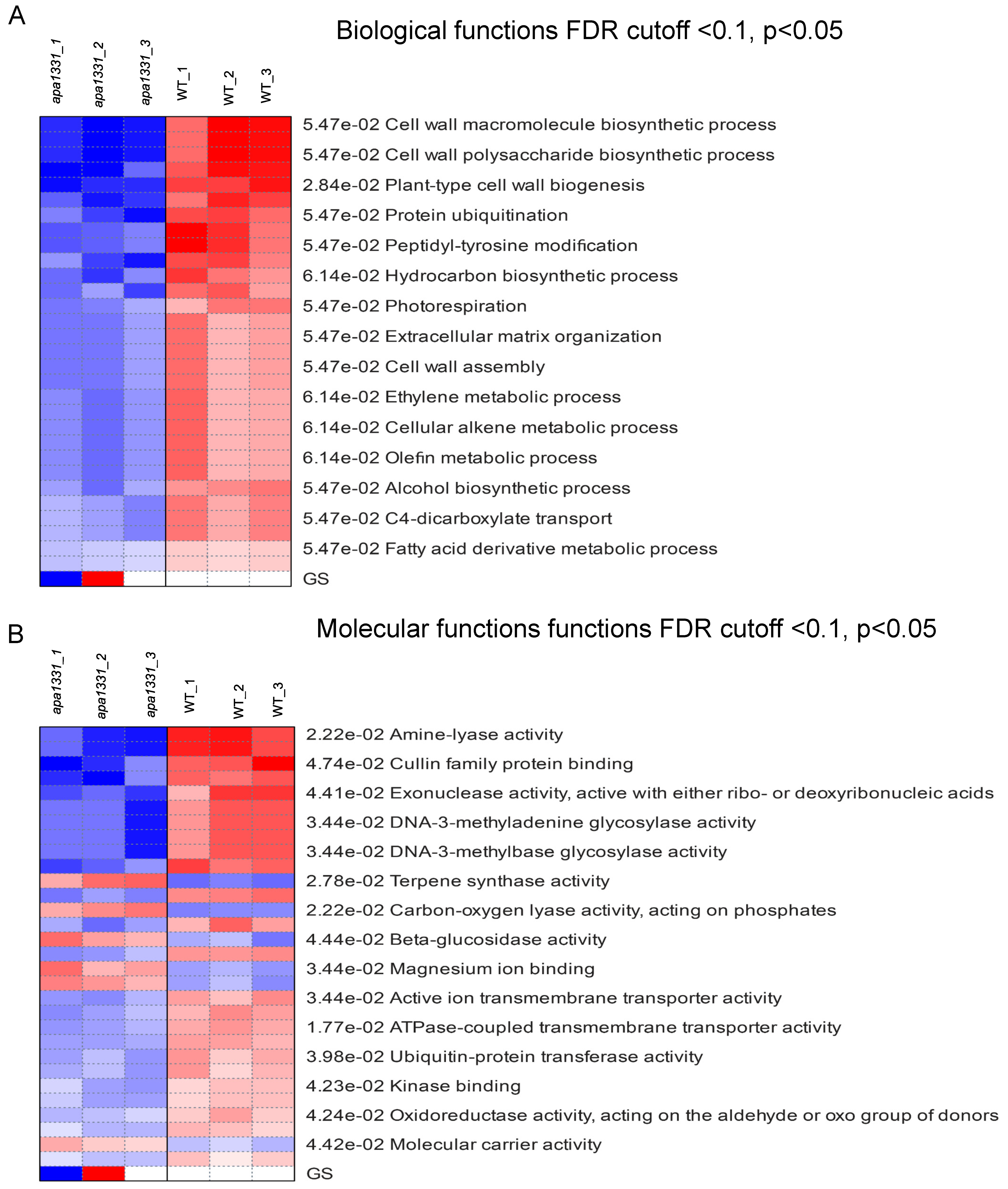
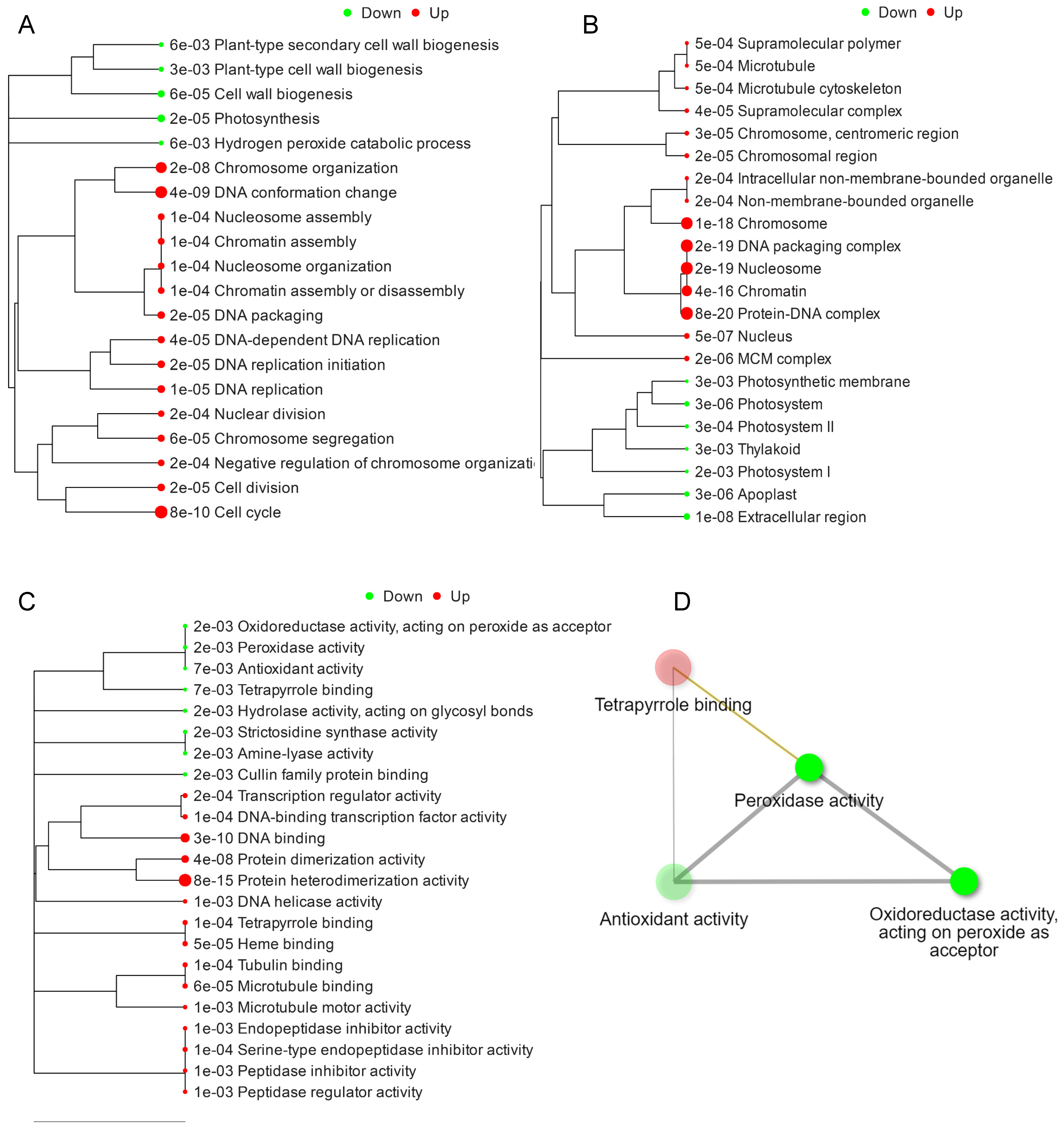
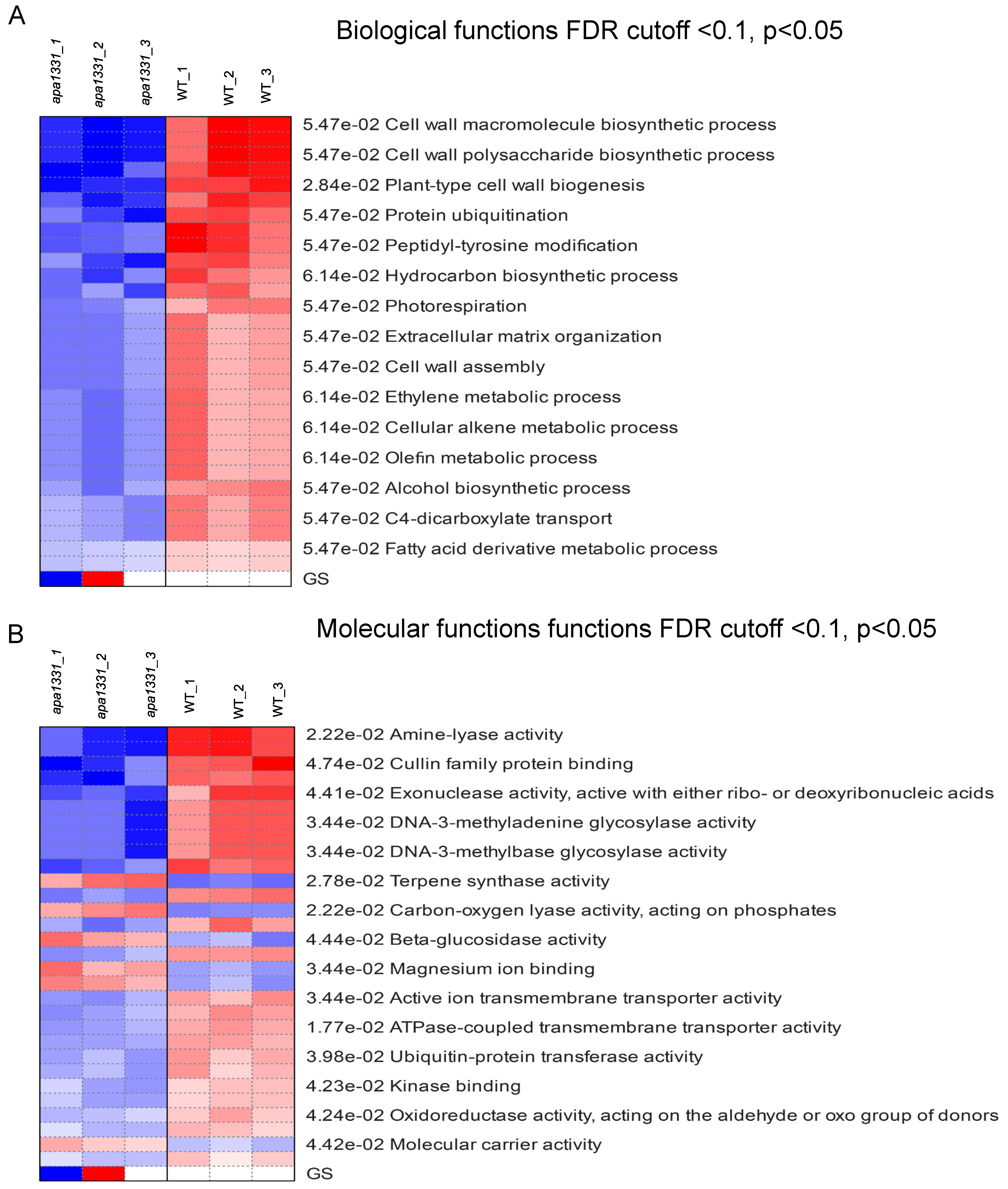
2.3. Hierarchical Clustering and GO Terms Analysis of DEGs
2.4. KEGG Pathway Expression Analysis of DEGs
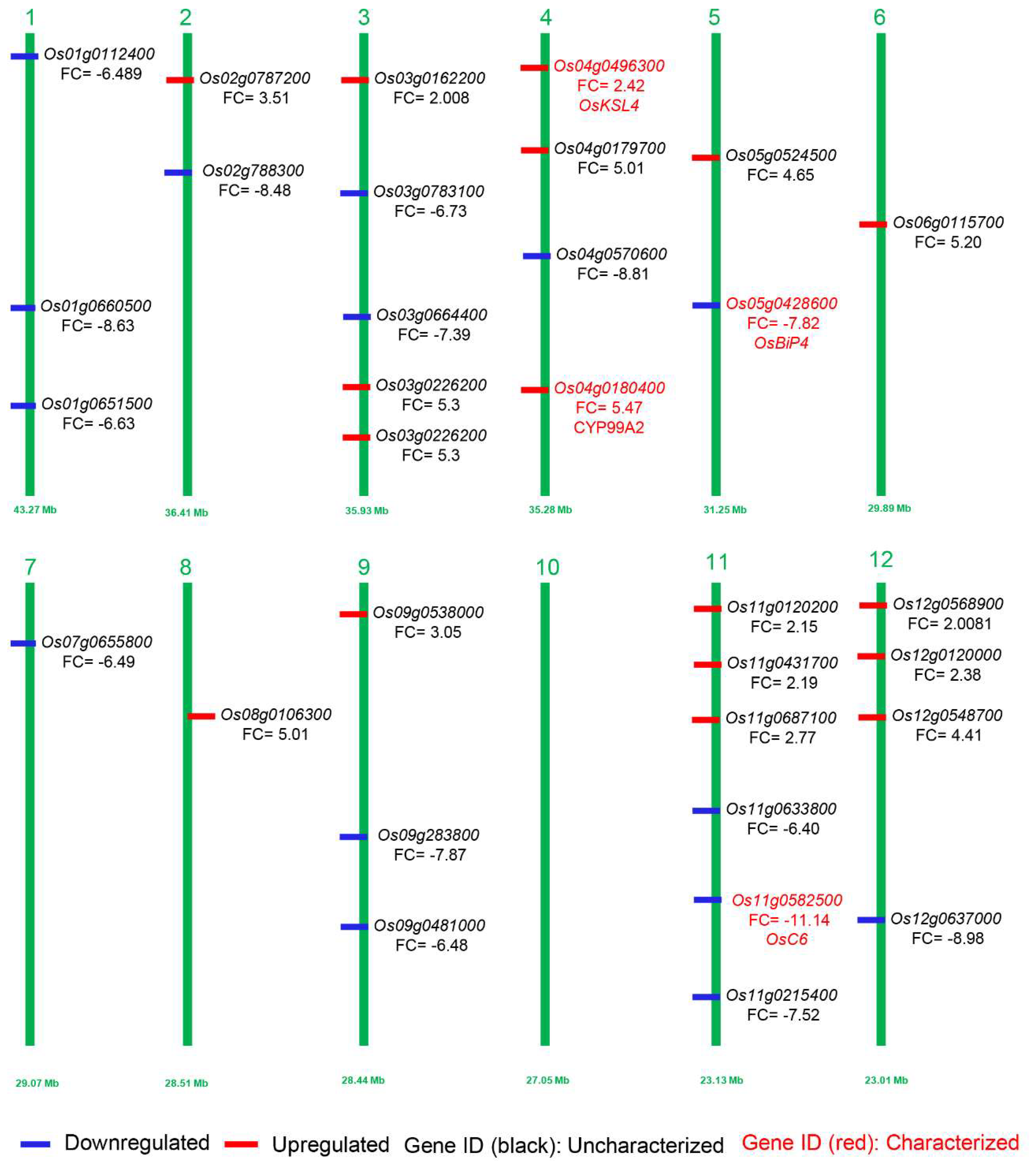
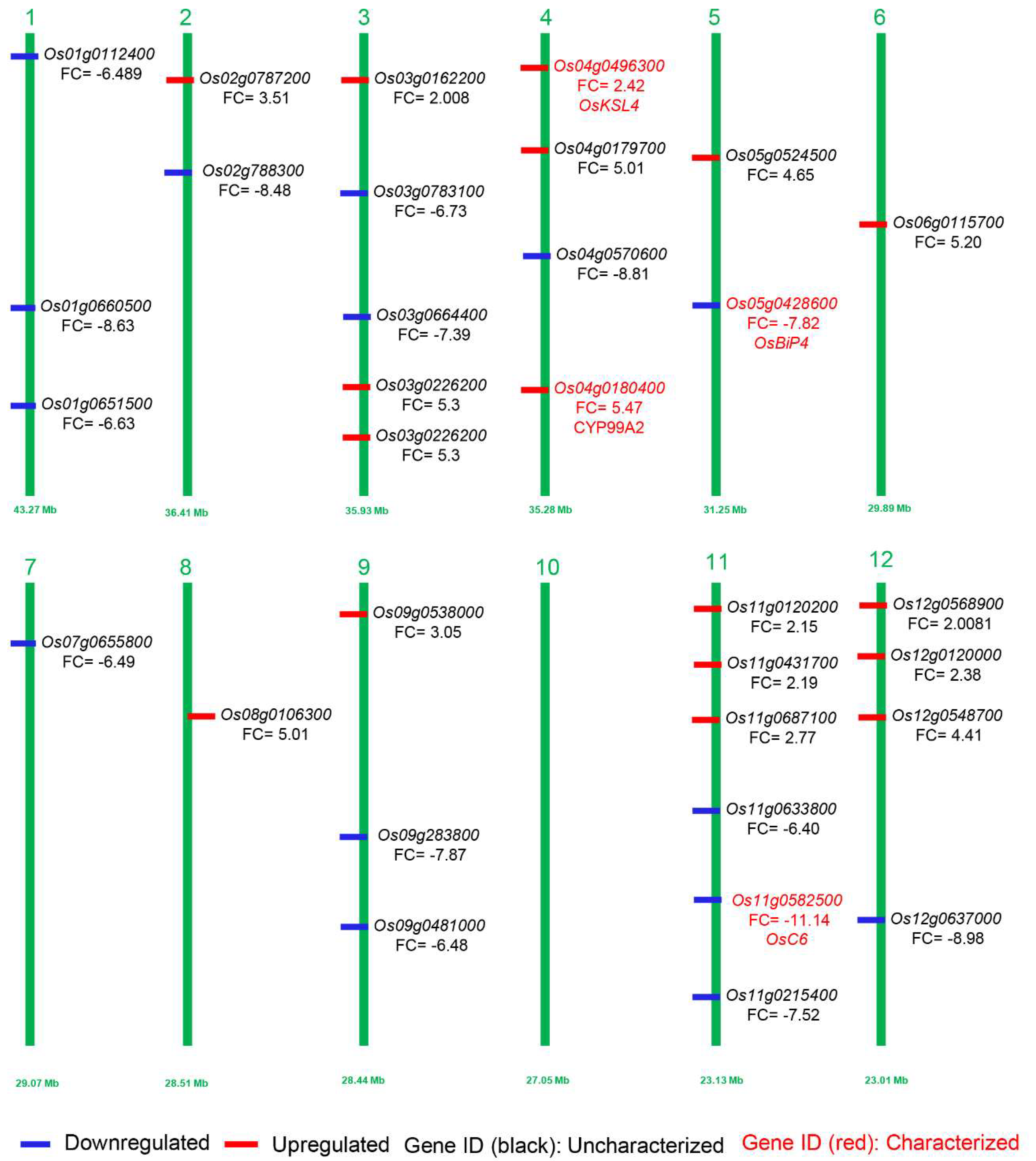
2.5. Transcriptome Response Revealed the Key Genes Involved in Panicle Development
2.6. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) Validation of Genes Expression
3. Discussion
3.1. Role of Phytohormones in ROS Homeostasis and Spikelet Abortion
3.2. The Transcriptomic Response of DEGs Highlighted the Role of PCD and Increased ROS in Panicle Abortion
3.3. Role of Antioxidants and Redox Changes in Inflorescence Development and Spikelet Abortion
3.4. Role of Non-Enzymatic Metabolites in Inflorescence Development and Spikelet Abortion
4. Materials and Methods
4.1. Experimental Material
4.2. Quantification of Endogenous Phytohormones
4.2.1. Samples Preparation
4.2.2. Quantification of GA
4.2.3. Quantification of IAA and ABA
4.2.4. Quantification of CTK and BR
4.3. RNA Extraction and Illumina Sequencing
4.4. Reads Mapping and Annotations
4.5. Differential Expression Genes (DEGs) Identification
4.6. Gene Function Annotation
4.7. RT-qPCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takeda, S.; Matsuoka, M. Genetic approaches to crop improvement: Responding to environmental and population changes. Nat. Rev. Genet. 2008, 9, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Zhang, Q. Genetic and molecular bases of rice yield. Annu. Rev. Plant Biol. 2010, 61, 421–442. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Matsuoka, M. Generating high-yielding varieties by genetic manipulation of plant architecture. Curr. Opin. Biotechnol. 2004, 15, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Xu, P.; Riaz, A.; Wu, X. Current Advances in Molecular Mechanisms and Physiological Basis of Panicle Degeneration in Rice. Int. J. Mol. Sci. 2019, 20, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Wu, T.; Zhang, H.; Xu, P.; Zafar, S.A.; Liao, Y.; Chen, X.; Zhou, H.; Liu, Y.; Wang, W.; et al. A putative SUBTILISIN-LIKE SERINE PROTEASE 1 (SUBSrP1) regulates anther cuticle biosynthesis and panicle development in rice. J. Adv. Res. 2022. [Google Scholar] [CrossRef]
- Peng, Y.; Hou, F.; Bai, Q.; Xu, P.; Liao, Y.; Zhang, H.; Gu, C.; Deng, X.; Wu, T.; Chen, X.; et al. Rice Calcineurin B-Like Protein-Interacting Protein Kinase 31 (OsCIPK31) Is Involved in the Development of Panicle Apical Spikelets. Front. Plant Sci. 2018, 9, 1661. [Google Scholar] [CrossRef]
- Pei, Y.; Deng, Y.; Zhang, H.; Zhang, Z.; Liu, J.; Chen, Z.; Cai, D.; Li, K.; Du, Y.; Zang, J. EAR APICAL DEGENERATION1 regulates maize ear development by maintaining malate supply for apical inflorescence. Plant Cell 2022, 34, 2222–2241. [Google Scholar] [CrossRef]
- Heng, Y.; Wu, C.; Long, Y.; Luo, S.; Ma, J.; Chen, J.; Liu, J.; Zhang, H.; Ren, Y.; Wang, M. OsALMT7 Maintains Panicle Size and Grain Yield in Rice by Mediating Malate Transport. Plant Cell 2018, 30, 889–906. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Zhu, X.; Wang, Q.; Zhang, J.; Chen, H.; Dong, G.; Zhu, L.; Zheng, H.; Xie, Q.; Nian, J.; et al. Rice TUTOU1 Encodes a Suppressor of cAMP Receptor-Like Protein That Is Important for Actin Organization and Panicle Development. Plant Physiol. 2015, 169, 1179–1191. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Zhang, R.; Huang, G.; Li, Y.; Melaku, G.; Zhang, S.; Chen, H.; Zhao, Y.; Zhang, J.; Zhang, Y. Developing superior alleles of yield genes in rice by artificial mutagenesis using the CRISPR/Cas9 system. Crop J. 2018, 6, 475–481. [Google Scholar] [CrossRef]
- Zafar, S.A.; Patil, S.B.; Uzair, M.; Fang, J.; Zhao, J.; Guo, T.; Yuan, S.; Uzair, M.; Luo, Q.; Shi, J. DEGENERATED PANICLE AND PARTIAL STERILITY 1 (DPS 1) encodes a cystathionine β-synthase domain containing protein required for anther cuticle and panicle development in rice. New Phytol. 2020, 225, 356–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevanthi, A.; Kandwal, P.; Kale, P.B.; Prakash, C.; Ramkumar, M.; Yadav, N.; Mahato, A.K.; Sureshkumar, V.; Behera, M.; Deshmukh, R.K. Whole genome characterization of a few EMS-induced mutants of upland rice variety Nagina 22 reveals a staggeringly high frequency of SNPs which show high phenotypic plasticity towards the wild-type. Front. Plant Sci. 2018, 9, 1179. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Chen, Z.; Lu, J.; Xu, C.; Xie, G.; Li, Y.; Deng, X.W.; He, H.; Tang, X. Simultaneous identification of multiple causal mutations in rice. Front. Plant Sci. 2017, 7, 2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, J.; Wang, L.; Zhu, L.; Bian, L.; Zhang, B.; Chen, X.; Huang, G.; Zhang, X.; Fan, J.; Cao, L. A rice single cell transcriptomic atlas defines the developmental trajectories of rice floret and inflorescence meristems. New Phytol. 2022, 234, 494–512. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Pan, Y.; Zhao, X.; Zhu, L.; Fu, B.; Li, Z. Genome-wide temporal-spatial gene expression profiling of drought responsiveness in rice. BMC Genom. 2011, 12, 149. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.; Moon, S.; Lee, Y.-S.; Zhu, L.; Liang, W.; Zhang, D.; Jung, K.-H.; An, G. Defective tapetum cell death 1 (DTC1) regulates ROS levels by binding to metallothionein during tapetum degeneration. Plant Physiol. 2016, 170, 1611–1623. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Shi, X.; Li, S.; Zhang, L.; Song, X. Oxidative stress and aberrant programmed cell death are associated with pollen abortion in isonuclear alloplasmic male-sterile wheat. Front. Plant Sci. 2018, 9, 595. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.; Shulaev, V.; Mittler, R. Reactive oxygen signaling and abiotic stress. Physiol. Plant. 2008, 133, 481–489. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, Y.; Wang, Z.; Yang, J. Polyamines and ethylene in rice young panicles in response to soil drought during panicle differentiation. Plant Growth Regul. 2017, 82, 491–503. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Boote, K.J.; Allen, L.H. Longevity and temperature response of pollen as affected by elevated growth temperature and carbon dioxide in peanut and grain sorghum. Environ. Exp. Bot. 2011, 70, 51–57. [Google Scholar] [CrossRef]
- Zheng, S.; Dong, J.; Lu, J.; Li, J.; Jiang, D.; Yu, H.; Ye, S.; Bu, W.; Liu, Z.; Zhou, H. A cytosolic pentatricopeptide repeat protein is essential for tapetal plastid development by regulating OsGLK1 transcript levels in rice. New Phytol. 2022, 234, 1678–1695. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Dai, X.; Qu, H.; Men, Q.; Liu, J.; Yu, L.; Gu, M.; Xu, G. The rice phosphate transporter OsPHT1; 7 plays a dual role in phosphorus redistribution and anther development. Plant Physiol. 2022, 188, 2272–2288. [Google Scholar] [CrossRef]
- Ishimaru, T.; Hirose, T.; Matsuda, T.; Goto, A.; Takahashi, K.; Sasaki, H.; Terao, T.; Ishii, R.-i.; Ohsugi, R.; Yamagishi, T. Expression patterns of genes encoding carbohydrate-metabolizing enzymes and their relationship to grain filling in rice (Oryza sativa L.): Comparison of caryopses located at different positions in a panicle. Plant Cell Physiol. 2005, 46, 620–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganeshaiah, K.; Uma Shaanker, R. Seed and fruit abortion as a process of self organization among developing sinks. Physiol. Plant. 1994, 91, 81–89. [Google Scholar] [CrossRef]
- Lee, B.; Martin, P.; Bangerth, F. Phytohormone levels in the florets of a single wheat spikelet during pre-anthesis development and relationships to grain set. J. Exp. Bot. 1988, 39, 927–933. [Google Scholar] [CrossRef]
- Wu, C.; Cui, K.; Wang, W.; Li, Q.; Fahad, S.; Hu, Q.; Huang, J.; Nie, L.; Peng, S. Heat-induced phytohormone changes are associated with disrupted early reproductive development and reduced yield in rice. Sci. Rep. 2016, 6, 34978. [Google Scholar] [CrossRef]
- Zhang, D.; Luo, X.; Zhu, L. Cytological analysis and genetic control of rice anther development. J. Genet. Genom. 2011, 38, 379–390. [Google Scholar] [CrossRef]
- Jun, D.; Chang-En, T.; Cheng-Ye, L.; Yu-Wen, H.; Hong-Xian, L. Dynamic changes of endogenous plant hormones in rice grains in different parts of panicle at grain filling stage. J. Integr. Plant Biol. 1999, 41, 315–323. [Google Scholar]
- Zhou, B.; Chen, H.; Huang, X.; Li, N.; Hu, Z.; Gao, Z.; Lu, Y. Rudimentary leaf abortion with the development of panicle in litchi: Changes in ultrastructure, antioxidant enzymes and phytohormones. Sci. Hortic. 2008, 117, 288–296. [Google Scholar] [CrossRef]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Thomas, P.D. GO functional similarity clustering depends on similarity measure, clustering method, and annotation completeness. BMC Bioinform. 2019, 20, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; Van Arkel, G.; Pereira, A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafar, S.A.; Uzair, M.; Khan, M.R.; Patil, S.; Fang, J.; Zhao, J.; Lata Singla-Pareek, S.; Pareek, A.; Li, X. DPS1 regulates cuticle development and leaf senescence in rice. Food Energy Secur. 2021, 10, e273. [Google Scholar]
- Kim, S.-Y.; Volsky, D.J. PAGE: Parametric analysis of gene set enrichment. BMC Bioinform. 2005, 6, 144. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, S.; Zhu, W.; Hamilton, J.; Lin, H.; Campbell, M.; Childs, K.; Thibaud-Nissen, F.; Malek, R.L.; Lee, Y.; Zheng, L. The TIGR rice genome annotation resource: Improvements and new features. Nucleic Acids Res. 2007, 35, D883–D887. [Google Scholar] [CrossRef] [Green Version]
- Childs, K.L.; Davidson, R.M.; Buell, C.R. Gene coexpression network analysis as a source of functional annotation for rice genes. PLoS ONE 2011, 6, e22196. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Liang, W.; Yin, C.; Zong, J.; Gu, F.; Zhang, D. OsC6, Encoding a Lipid Transfer Protein, Is Required for Postmeiotic Anther Development In Rice. Plant Physiol. 2010, 154, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Wakasa, Y.; Hayashi, S.; Takaiwa, F. Expression of OsBiP4 and OsBiP5 is highly correlated with the endoplasmic reticulum stress response in rice. Planta 2012, 236, 1519–1527. [Google Scholar] [CrossRef]
- Okada, A.; Okada, K.; Miyamoto, K.; Koga, J.; Shibuya, N.; Nojiri, H.; Yamane, H. OsTGAP1, a bZIP transcription factor, coordinately regulates the inductive production of diterpenoid phytoalexins in rice. J. Biol. Chem. 2009, 284, 26510–26518. [Google Scholar] [CrossRef] [Green Version]
- Margis-Pinheiro, M.; Zhou, X.-R.; Zhu, Q.-H.; Dennis, E.S.; Upadhyaya, N.M. Isolation and characterization of a Ds-tagged rice (Oryza sativa L.) GA-responsive dwarf mutant defective in an early step of the gibberellin biosynthesis pathway. Plant Cell Rep. 2005, 23, 819–833. [Google Scholar] [CrossRef] [PubMed]
- Barbier, F.F.; Dun, E.A.; Beveridge, C.A. Apical dominance. Curr. Biol. 2017, 27, R864–R865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaliang, W.; Yikai, Z.; Qinghua, S.; Huizhe, C.; Jing, X.; Guohui, H.; Yanhua, C.; Xiaodan, W.; Junke, W.; Zihao, Y. Decrement of sugar consumption in rice young panicle under high temperature aggravates spikelet number reduction. Rice Sci. 2020, 27, 44–55. [Google Scholar] [CrossRef]
- Zhang, C.; Feng, B.; Chen, T.; Zhang, X.; Tao, L.; Fu, G. Sugars, antioxidant enzymes and IAA mediate salicylic acid to prevent rice spikelet degeneration caused by heat stress. Plant Growth Regul. 2017, 83, 313–323. [Google Scholar] [CrossRef]
- Alqudah, A.M.; Sharma, R.; Börner, A. Insight into the genetic contribution of maximum yield potential, spikelet development and abortion in barley. Plants People Planet 2021, 3, 721–736. [Google Scholar] [CrossRef]
- Ahmadi, A.; Baker, D. Effects of abscisic acid (ABA) on grain filling processes in wheat. Plant Growth Regul. 1999, 28, 187–197. [Google Scholar] [CrossRef]
- Kwak, J.M.; Mori, I.C.; Pei, Z.-M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Liu, K.; Wang, Z.; Liu, L. Abscisic acid and ethylene interact in rice spikelets in response to water stress during meiosis. J. Plant Growth Regul. 2007, 26, 318–328. [Google Scholar] [CrossRef]
- Deveshwar, P.; Bovill, W.D.; Sharma, R.; Able, J.A.; Kapoor, S. Analysis of anther transcriptomes to identify genes contributing to meiosis and male gametophyte development in rice. BMC Plant Biol. 2011, 11, 78. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Y.; Zhang, Q.; Cui, Y.; Xiang, J.; Chen, H.; Hu, G.; Chen, Y.; Wang, X.; Zhu, D. Comparative transcriptome analysis of panicle development under heat stress in two rice (Oryza sativa L.) cultivars differing in heat tolerance. PeerJ 2019, 7, e7595. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Yang, H.; Wei, Z.; Ma, H.; Ge, X. Rice male development under drought stress: Phenotypic changes and stage-dependent transcriptomic reprogramming. Mol. Plant 2013, 6, 1630–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockshin, R.A.; Zakeri, Z. Apoptosis, autophagy, and more. Int. J. Biochem. Cell Biol. 2004, 36, 2405–2419. [Google Scholar] [CrossRef]
- Van Doorn, W.G.; Beers, E.P.; Dangl, J.L.; Franklintong, V.E.; Gallois, P.; Haranishimura, I.; Jones, A.M.; Kawaiyamada, M.; Lam, E.; Mundy, J. Morphological classification of plant cell deaths. Cell Death Differ. 2011, 18, 1241–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Breusegem, F.; Dat, J.F. Reactive Oxygen Species in Plant Cell Death. Plant Physiol. 2006, 141, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Sun, A.; Chen, S.; Chen, L.; Guo, F. SPL6 represses signalling outputs of ER stress in control of panicle cell death in rice. Nat. Plants 2018, 4, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.-L.; Zheng, X.-L.; Zhou, C.-Y.; Kanwar, M.K.; Zhou, J. Functions of redox signaling in pollen development and stress response. Antioxidants 2022, 11, 287. [Google Scholar] [CrossRef]
- Ji, C.; Li, H.; Chen, L.; Xie, M.; Wang, F.; Chen, Y.; Liu, Y.-G. A novel rice bHLH transcription factor, DTD, acts coordinately with TDR in controlling tapetum function and pollen development. Mol. Plant 2013, 6, 1715–1718. [Google Scholar] [CrossRef] [Green Version]
- Larkin, R.M. Tetrapyrrole signaling in plants. Front. Plant Sci. 2016, 7, 1586. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.H.; Suen, D.F. Iron insufficiency in floral buds impairs pollen development by disrupting tapetum function. Plant J. 2021, 108, 244–267. [Google Scholar] [CrossRef]
- Hernández, I.; Munné-Bosch, S. Linking phosphorus availability with photo-oxidative stress in plants. J. Exp. Bot. 2015, 66, 2889–2900. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Stress-triggered redox signalling: What’s in pROSpect? Plant Cell Environ. 2016, 39, 951–964. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.; Biancucci, M.; Lonoce, C.; Costantino, P.; Trovato, M. Proline is required for male gametophyte development in Arabidopsis. BMC Plant Biol. 2012, 12, 236. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Chourey, P.S.; Boote, K.J.; Allen, L.H., Jr. Short-term high temperature growth conditions during vegetative-to-reproductive phase transition irreversibly compromise cell wall invertase-mediated sucrose catalysis and microspore meiosis in grain sorghum (Sorghum bicolor). J. Plant Physiol. 2010, 167, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.S.; Shu, S.; Wang, Y.; Hasan, M.; El-Yazied, A.A.; Alabdallah, N.M.; Hajjar, D.; Altaf, M.A.; Sun, J.; Guo, S. Melatonin pretreatment confers heat tolerance and repression of heat-induced senescence in tomato through the modulation of ABA-and GA-mediated pathways. Front. Plant Sci. 2021, 12, 381. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-Y.; Li, X.; Zhao, D.; Deng-Wang, M.-Y.; Dai, C.-C. Reactive oxygen species and hormone signaling cascades in endophytic bacterium induced essential oil accumulation in Atractylodes lancea. Planta 2016, 244, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G. Ensembl 2018. Nucleic Acids Res. 2018, 46, D754–D761. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Wang, X.-J. GOEAST: A web-based software toolkit for Gene Ontology enrichment analysis. Nucleic Acids Res. 2008, 36, W358–W363. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Ali, A.; Wang, J.; Song, J.; Fang, Y.; Zhou, T.; Luo, Y.; Zhang, H.; Chen, X.; Liao, Y. A homologous gene of OsREL2/ASP1, ASP-LSL regulates pleiotropic phenotype including long sterile lemma in rice. BMC Plant Biol. 2021, 21, 390. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RAPD | MSU | FC | P- Adjusted |
Late Inflorescences (FPKM) |
Early Inflorescence (FPKM) |
Anther (FPKM) | GO |
|---|---|---|---|---|---|---|---|
| Os11g0582500 | LOC_Os11g37280 | −11.148 | 5.58 × 10−14 | 273.82 | 7.2958 | 19.027 | lipid binding, transport, plasma membrane |
| Os12g0637000 | LOC_Os12g44010 | −8.9894 | 3.73 × 10−13 | 8.3933 | 0.8633 | 4.3885 | cell wall, hyrdorlase activity, cytosol |
| Os04g0570600 | LOC_Os04g48210 | −8.8109 | 1.00 × 10−11 | 0.4118 | 6.4236 | 4.8603 | catalytic activity, metabolic processes, oxygen binding |
| Os01g0660500 | LOC_Os01g47100 | −8.6318 | 6.12 × 10−16 | 0 | 0.7213 | 5.8671 | nucleotide process, nuclease, nucleic acid binding |
| Os02g0788300 | LOC_Os02g54690 | −8.4887 | 1.42 × 10−14 | 0 | 0.6296 | 0.6264 | translation, nucelic acid binding, cytoplasm |
| Os12g0242700 | LOC_Os12g13930 | −8.4829 | 4.58 × 10−19 | 0.4769 | 15.97 | 2.6067 | plastid, transferase, lipidic metabolic process |
| Os09g0273800 | LOC_Os09g10200 | −7.8273 | 1.50 × 10−10 | 0 | 0.8435 | 0 | cell cycle, cytoskeleton, nucleus |
| Os05g0428600 | LOC_Os05g35400 | −7.5769 | 1.77 × 10−14 | 0.2241 | 1.7673 | 0.5022 | endoplasmic reticulum, response to biotic stress, cellular component |
| Os11g0215400 | LOC_Os11g10910 | −7.5261 | 7.58 × 10−29 | 0 | 10.882 | 0 | protein metabolic process, hydrolase activity, chloroplast |
| Os03g0664400 | LOC_Os03g46150 | −7.3955 | 4.39 × 10−5 | 0 | 7.2273 | 0 | lipid binding, transport, membrane |
| Os01g0112400 | LOC_Os01g02190 | −6.8985 | 0.000386131 | 0 | 47.986 | 0 | transport, transporter activity, cellular process |
| Os03g0783100 | LOC_Os03g56974 | −6.7322 | 8.00 × 10−8 | 0 | 3.2233 | 0 | topoisomerase function, nuclear antigen |
| Os01g0651500 | LOC_Os01g46270 | −6.6382 | 3.98 × 10−7 | 13.876 | 0 | 0 | lipidic process, transferase activity, protein binding |
| Os07g0655800 | LOC_Os07g46210 | −6.4993 | 1.56 × 10−72 | 0 | 63.094 | 0.1016 | ATPase activity, LTP family protein, protease inhibition |
| Os09g0481000 | LOC_Os09g30320 | −6.4816 | 6.41 × 10−6 | 704.44 | 62.081 | 178.28 | BURP domain-containing protein |
| Os11g0633800 | LOC_Os11g41560 | −6.4084 | 1.52 × 10−5 | 3.31 | 1.9748 | 4.2554 | F-box domain-containing protein |
| RAPD | MSU | FC | P-adjusted | Late Inflorescences (FPKM) | Early Inflorescence (FPKM) | Anther (FPKM) | GO |
|---|---|---|---|---|---|---|---|
| Os03g0162200 | LOC_Os03g06670 | 2.008056 | 2.68 × 10−5 | 10.1154 | 108.937 | 2.40101 | flower development, DNA binding, response to external stimulus |
| Os12g0568900 | LOC_Os12g38120 | 2.089498 | 1.86 × 10−9 | 2.32593 | 66.3759 | 0 | response to biotic stimulus, membrane |
| Os11g0120200 | LOC_Os11g02740 | 2.15798 | 5.63 × 10−5 | 1.48597 | 6.96247 | 0.79251 | cytosol, biological processes |
| Os11g0431700 | LOC_Os11g24374 | 2.299897 | 3.29 × 10−10 | 0.637085 | 1.49044 | 0 | cell, protein metabolic processes, hydrolase activity |
| Os12g0120000 | LOC_Os12g02710 | 2.383414 | 5.15 × 10−8 | 1.29036 | 6.28534 | 0.768418 | molecular functions, cytosol, biological process |
| Os04g0496300 | LOC_Os04g41900 | 2.429537 | 3.13 × 10−9 | 10.2279 | 114.047 | 3.76954 | molecular functions, cellular components, biological process |
| Os11g0687100 | LOC_Os11g45990 | 2.770047 | 0.000458 | 3.23511 | 22.0661 | 0 | protein modification, binding, catalytic activity |
| Os09g0538000 | LOC_Os09g36700 | 3.053863 | 2.19 × 10−22 | 90.9664 | 36.714 | 0.399433 | cell, nuclease activity, protein modification |
| Os02g0787200 | LOC_Os02g54590 | 3.510367 | 4.94 × 10−5 | 0.516972 | 0.439793 | 94.1342 | metabolic processes, cellular processes, response to stress |
| Os12g0548700 | LOC_Os12g36240 | 4.411432 | 7.2 × 10−5 | 0 | 0 | 0.835161 | response to stress, metabolic stress, cell wall |
| Os04g0179700 | LOC_Os04g10060 | 5.012307 | 6.83 × 10−5 | 0.498632 | 0 | 0 | catalytic activity, plastid, lipid metabolic processes. |
| Os08g0106300 | LOC_Os08g01520 | 5.014211 | 4.17 × 10−9 | 0 | 0.479214 | 0 | binding, catalytic activity, oxygen binding |
| Os06g0115700 | LOC_Os06g02530 | 5.207817 | 6.28 × 10−6 | 8.49399 | 37.2876 | 1.03739 | molecular function, biological process |
| Os03g0226200 | LOC_Os03g12510 | 5.306009 | 3.32 × 10−6 | 0.871437 | 0 | 2.50797 | cell wall, cytosol, oxygen binding |
| Os04g0180400 | LOC_Os04g10160 | 5.479893 | 4.09 × 10−5 | 0.492373 | 0 | 0 | metabolic process, cytosol, oxygen binding |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.; Wu, T.; Xu, Z.; Riaz, A.; Alqudah, A.M.; Iqbal, M.Z.; Zhang, H.; Liao, Y.; Chen, X.; Liu, Y.; et al. Phytohormones and Transcriptome Analyses Revealed the Dynamics Involved in Spikelet Abortion and Inflorescence Development in Rice. Int. J. Mol. Sci. 2022, 23, 7887. https://doi.org/10.3390/ijms23147887
Ali A, Wu T, Xu Z, Riaz A, Alqudah AM, Iqbal MZ, Zhang H, Liao Y, Chen X, Liu Y, et al. Phytohormones and Transcriptome Analyses Revealed the Dynamics Involved in Spikelet Abortion and Inflorescence Development in Rice. International Journal of Molecular Sciences. 2022; 23(14):7887. https://doi.org/10.3390/ijms23147887
Chicago/Turabian StyleAli, Asif, Tingkai Wu, Zhengjun Xu, Asad Riaz, Ahmad M. Alqudah, Muhammad Zafar Iqbal, Hongyu Zhang, Yongxiang Liao, Xiaoqiong Chen, Yutong Liu, and et al. 2022. "Phytohormones and Transcriptome Analyses Revealed the Dynamics Involved in Spikelet Abortion and Inflorescence Development in Rice" International Journal of Molecular Sciences 23, no. 14: 7887. https://doi.org/10.3390/ijms23147887
APA StyleAli, A., Wu, T., Xu, Z., Riaz, A., Alqudah, A. M., Iqbal, M. Z., Zhang, H., Liao, Y., Chen, X., Liu, Y., Mujtaba, T., Zhou, H., Wang, W., Xu, P., & Wu, X. (2022). Phytohormones and Transcriptome Analyses Revealed the Dynamics Involved in Spikelet Abortion and Inflorescence Development in Rice. International Journal of Molecular Sciences, 23(14), 7887. https://doi.org/10.3390/ijms23147887