
Dynamic Changes in the Gene Expression Patterns and Lipid Profiles in the Developing and Maturing Meibomian Glands



Abstract
:1. Introduction
2. Results
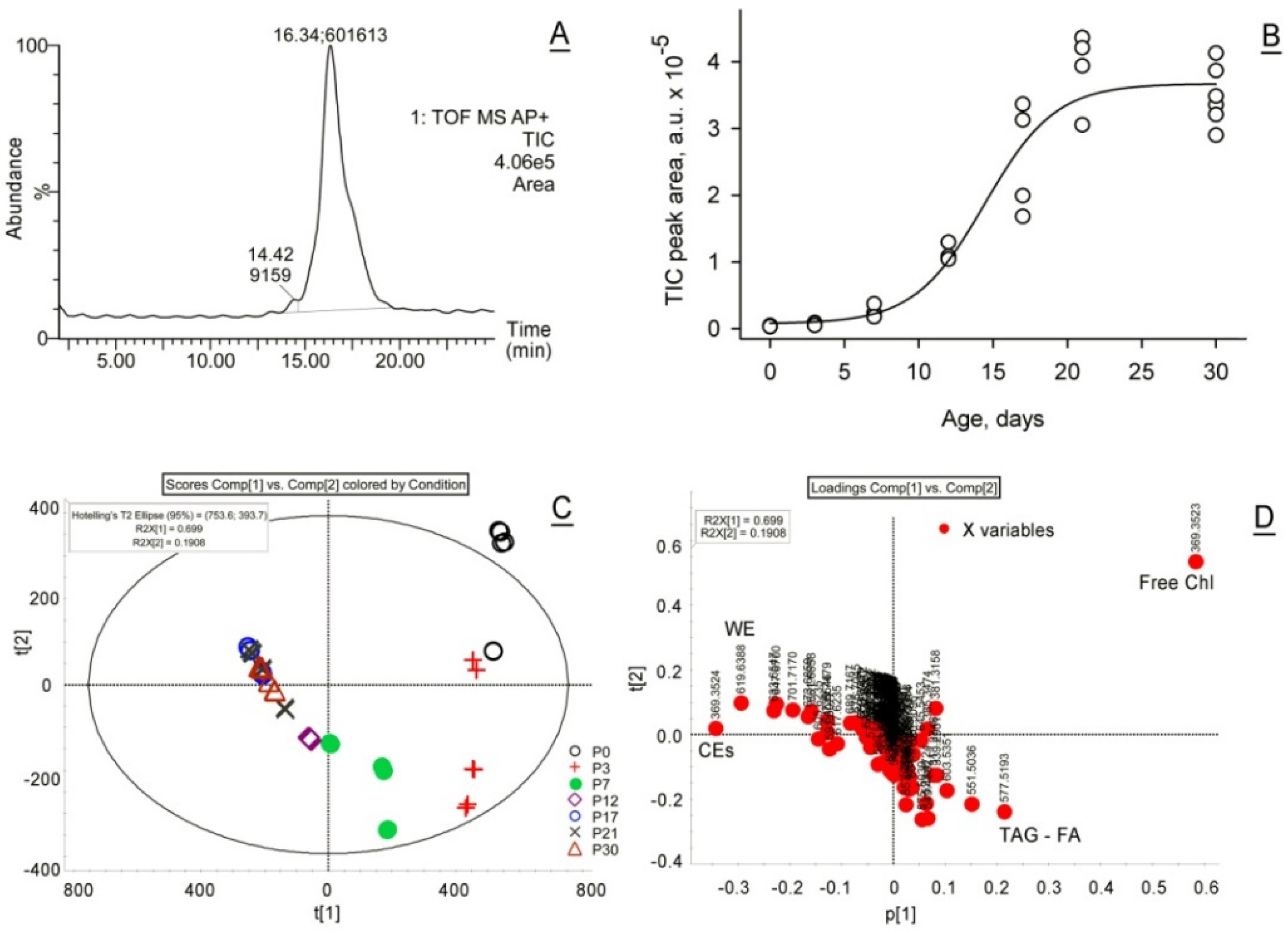
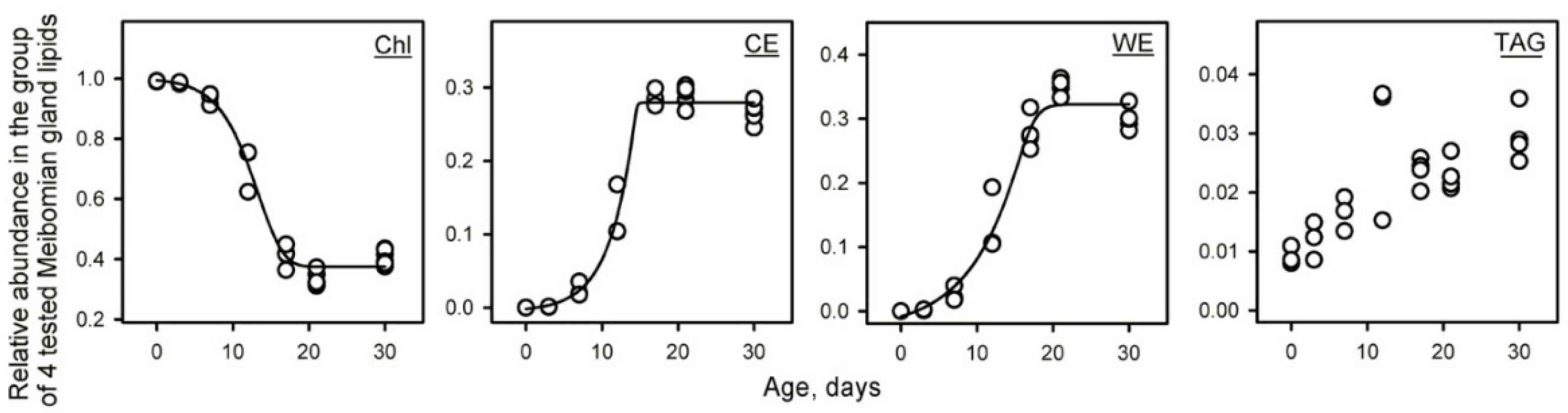
2.1. Lipidomic Analyses of Developing and Maturing MGs
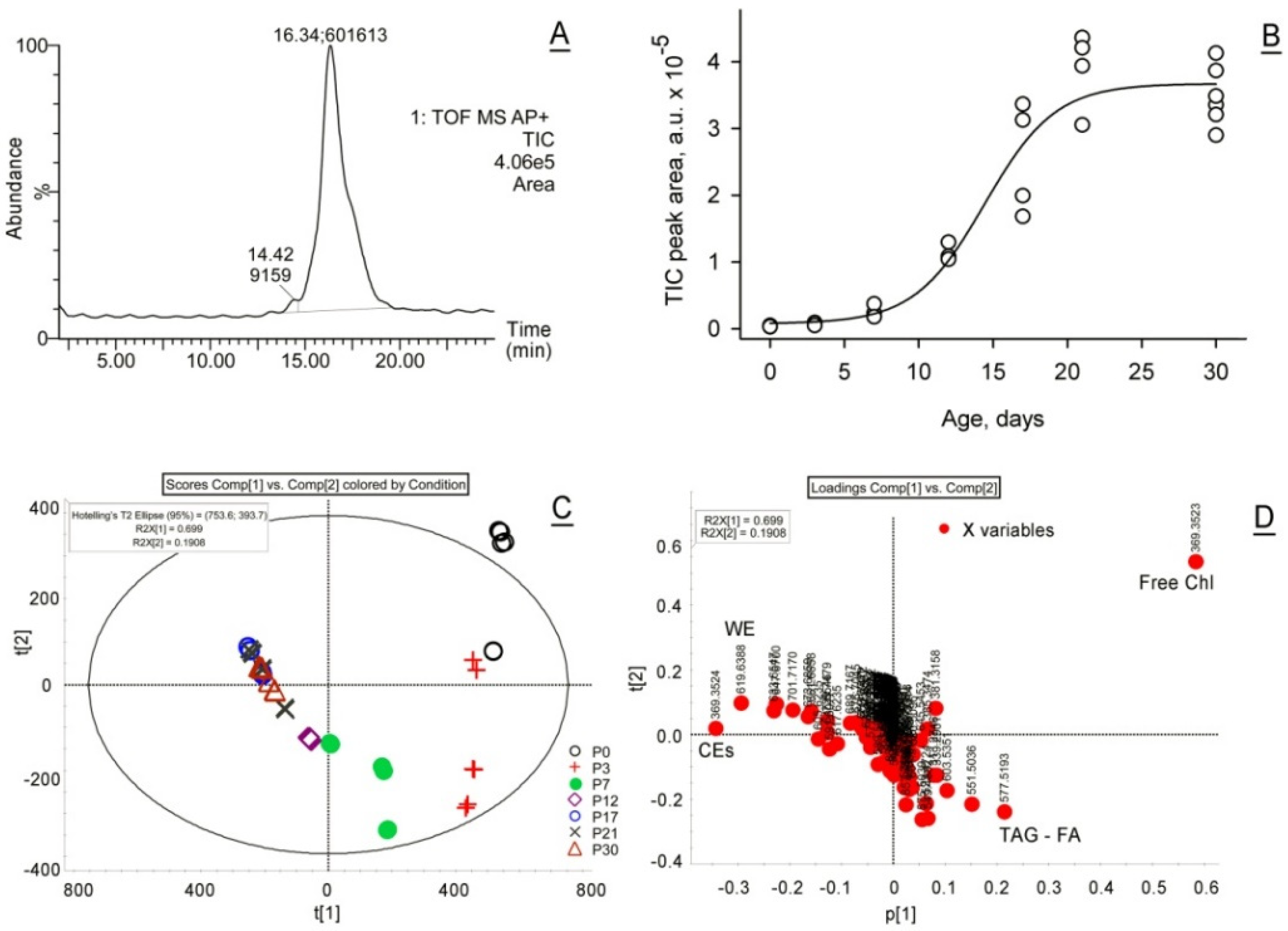
2.1.1. Unbiased, Untargeted Analyses of MG Lipids
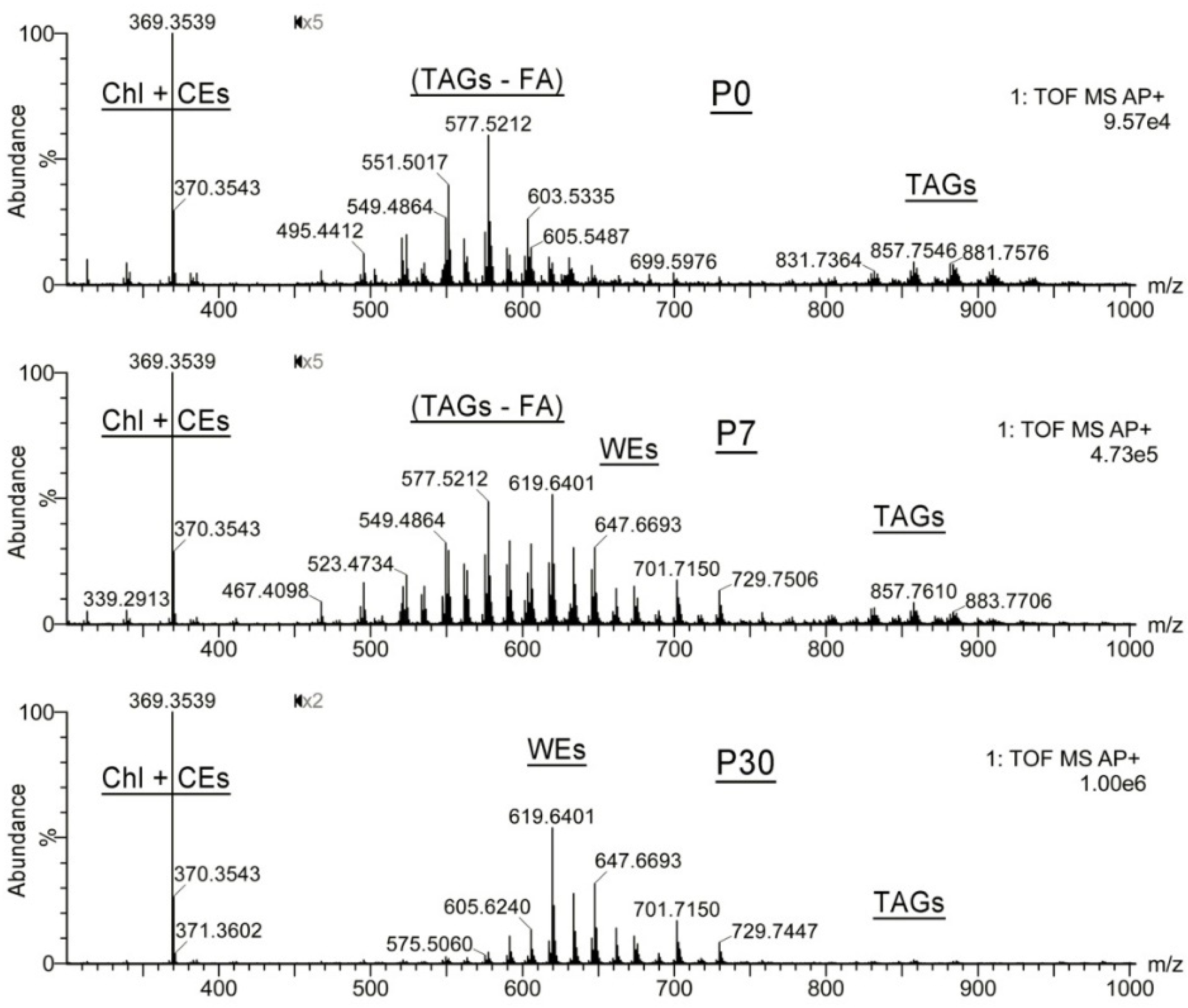
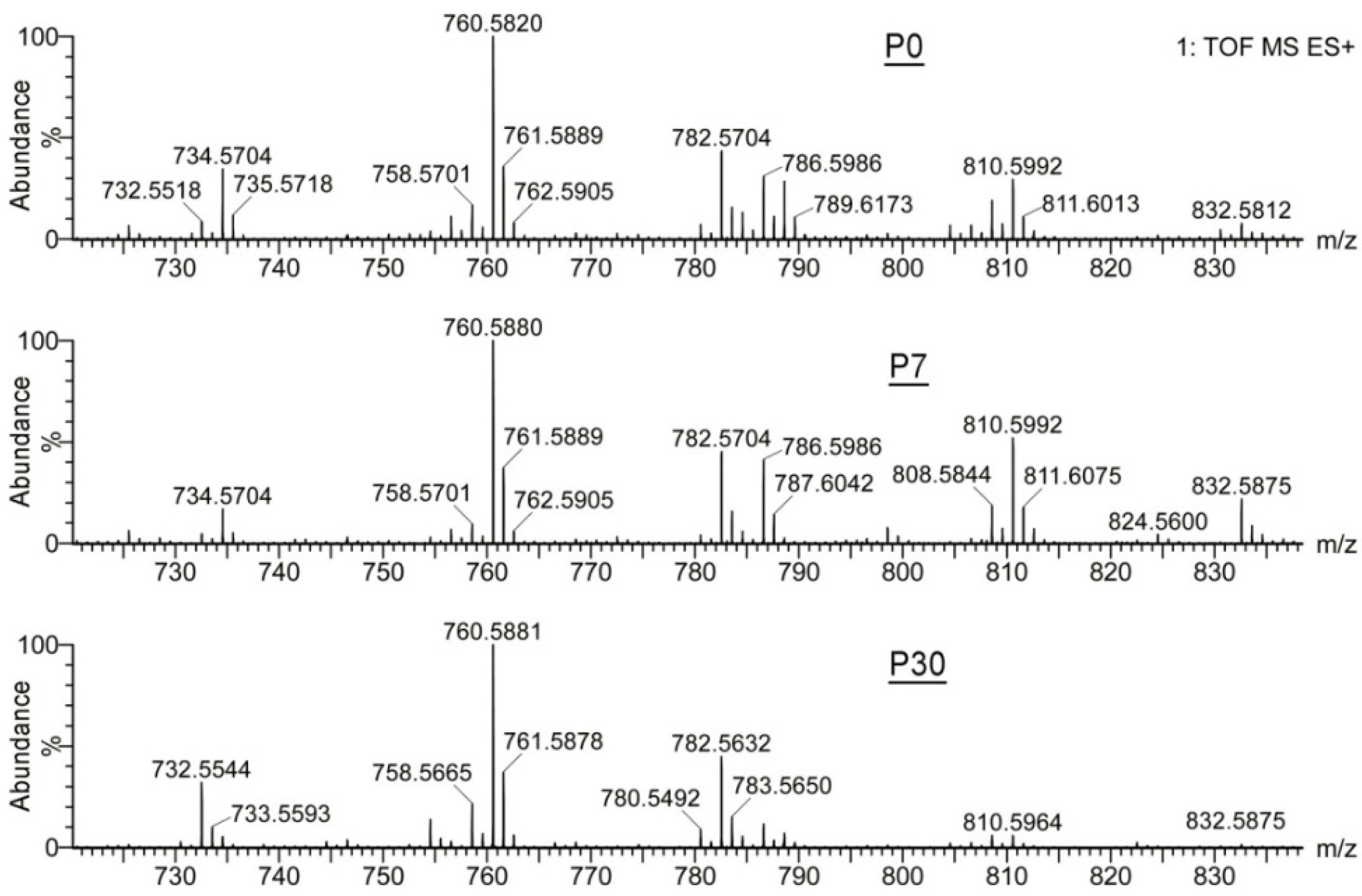
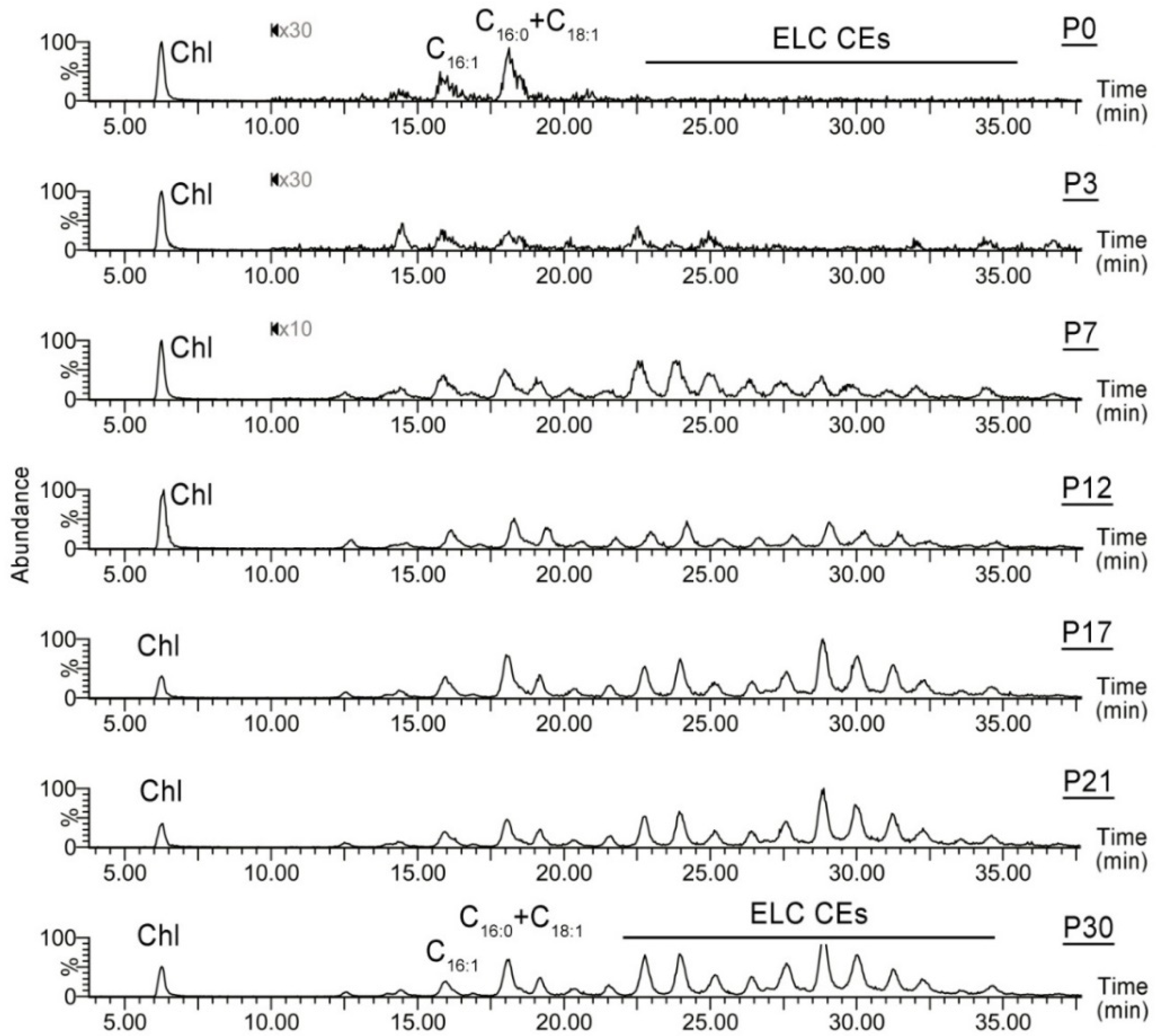
2.1.2. Observation Mass Spectra of Meibomian Lipids
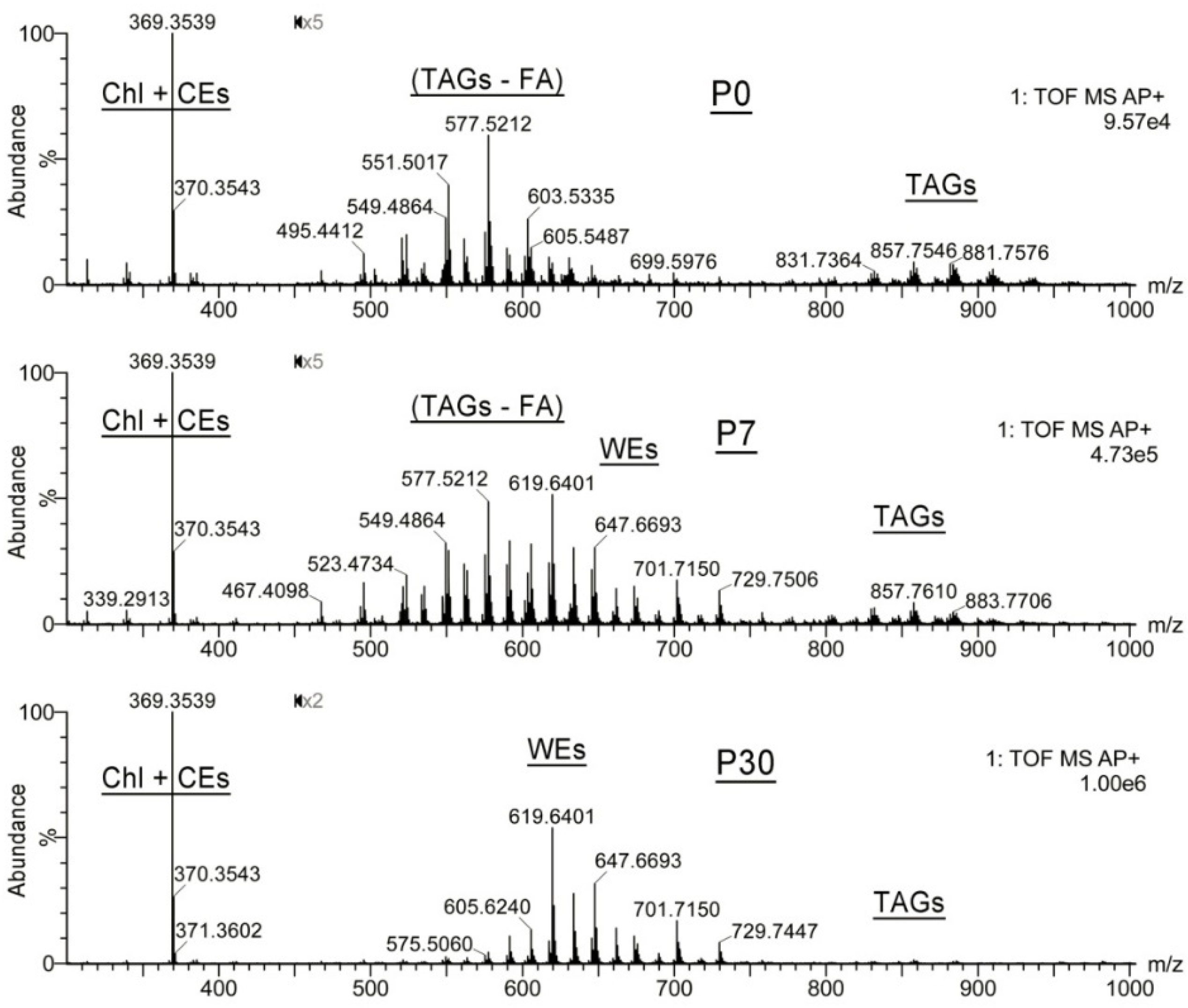
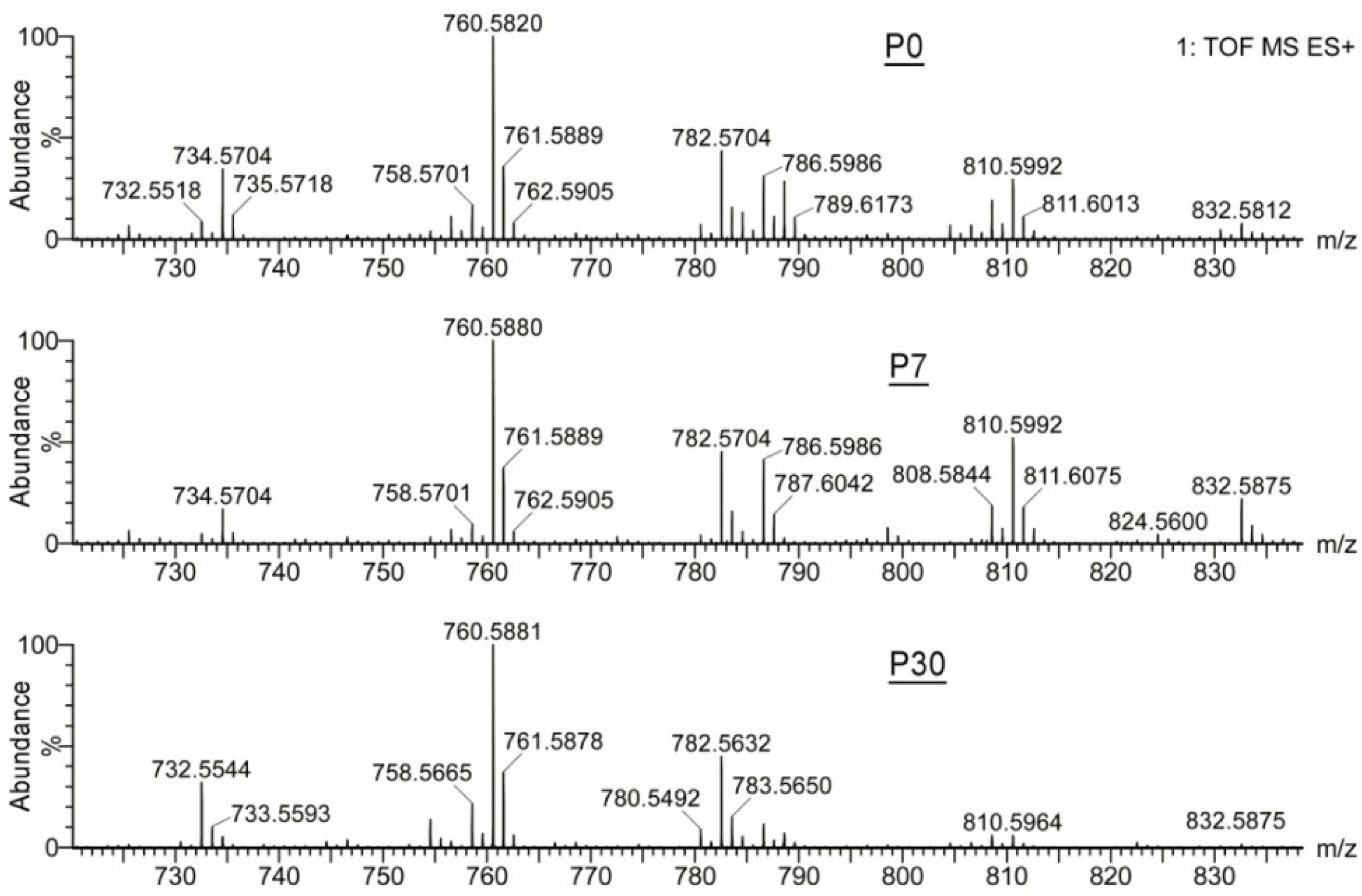
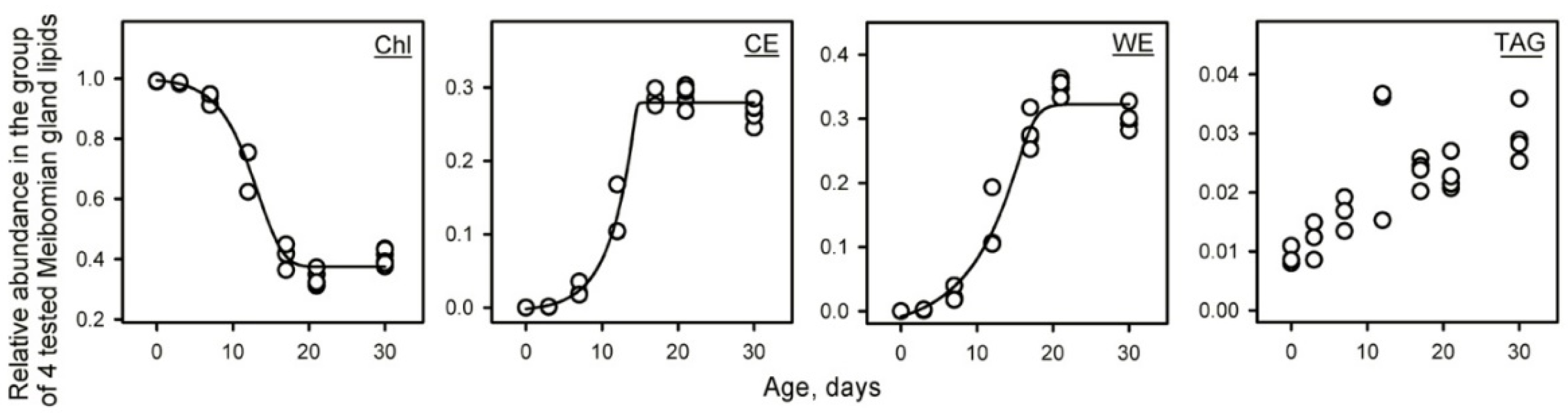
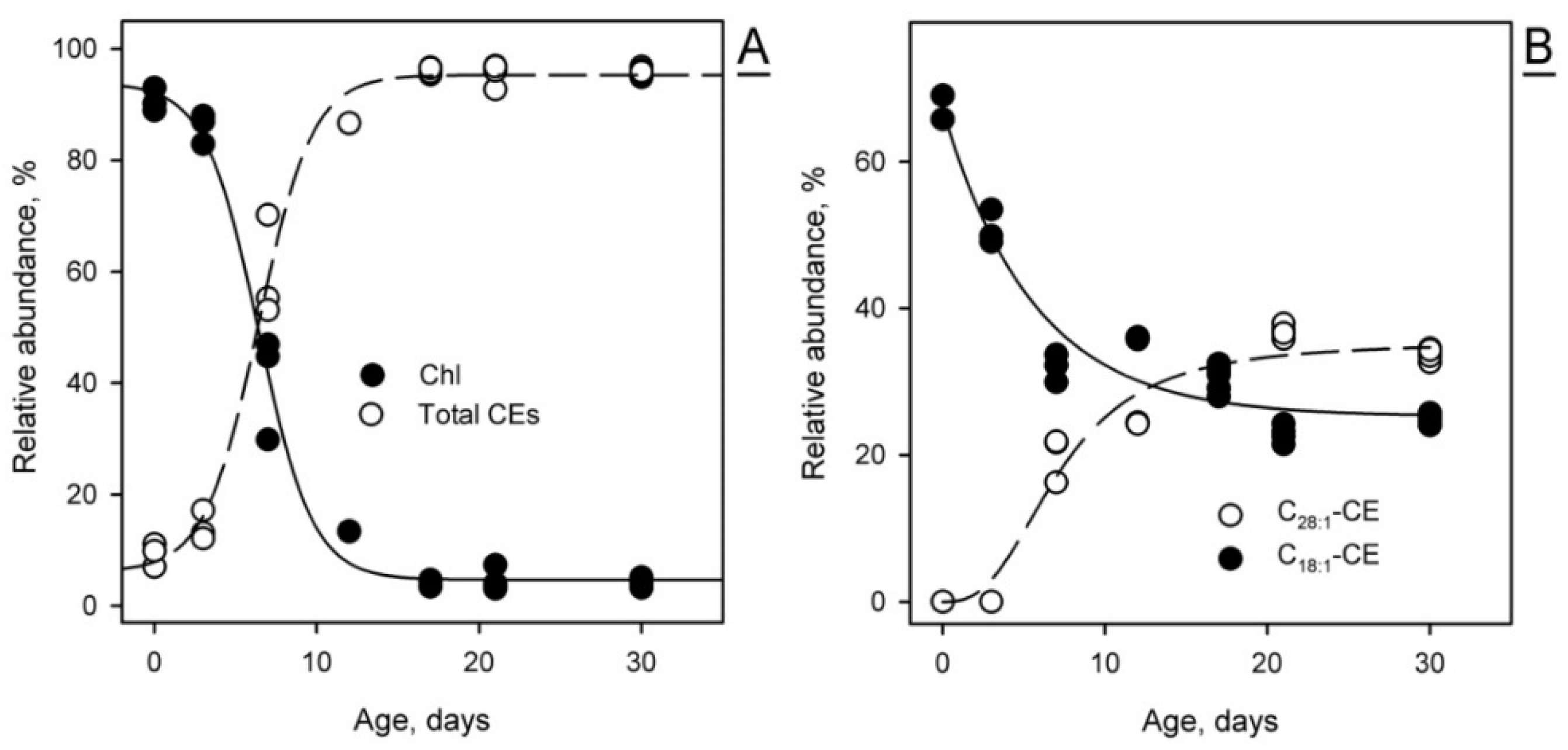
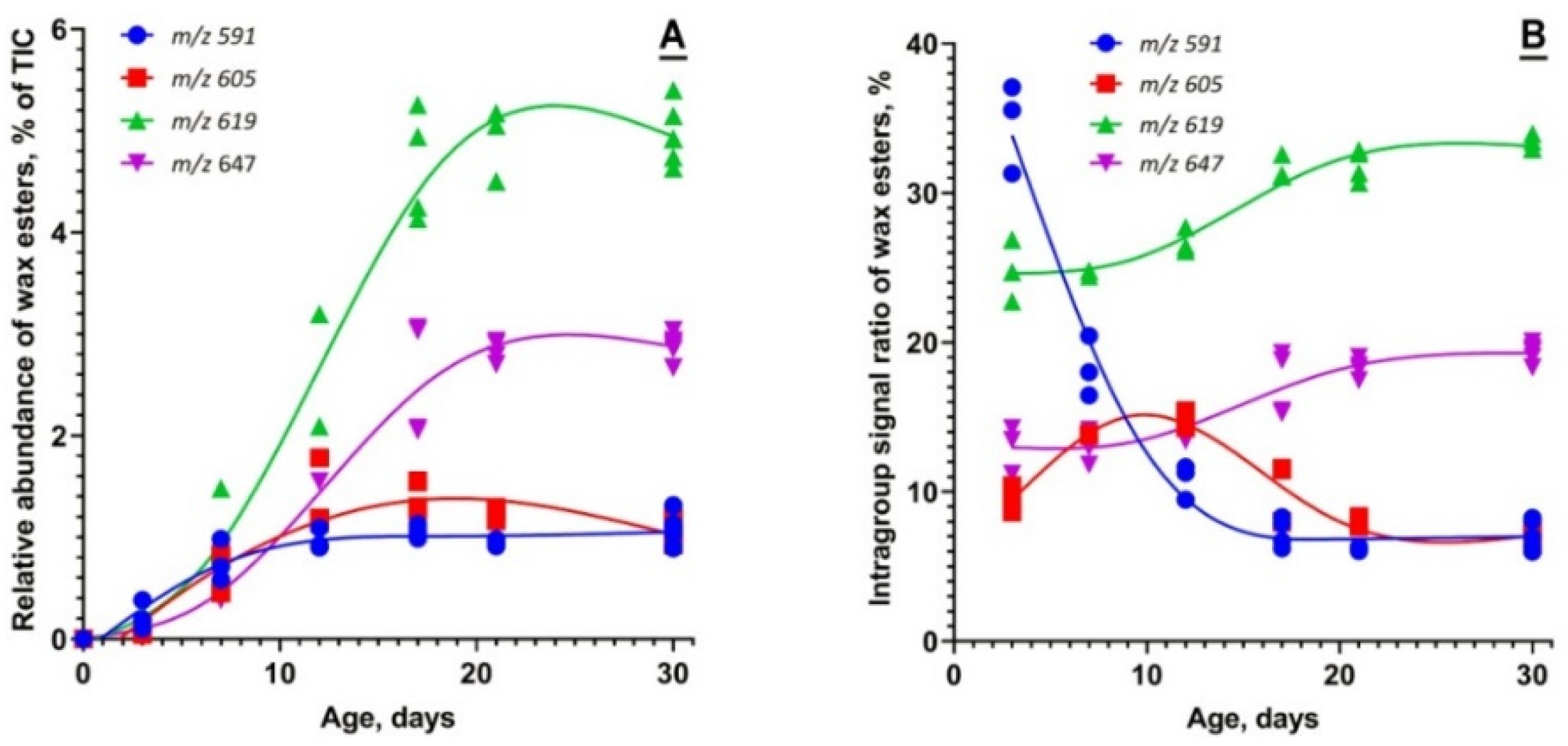
2.1.3. Targeted Analysis of Meibomian Lipids
2.2. mRNA Microarray Analyses of Mouse Tarsal Plate Transcriptomes
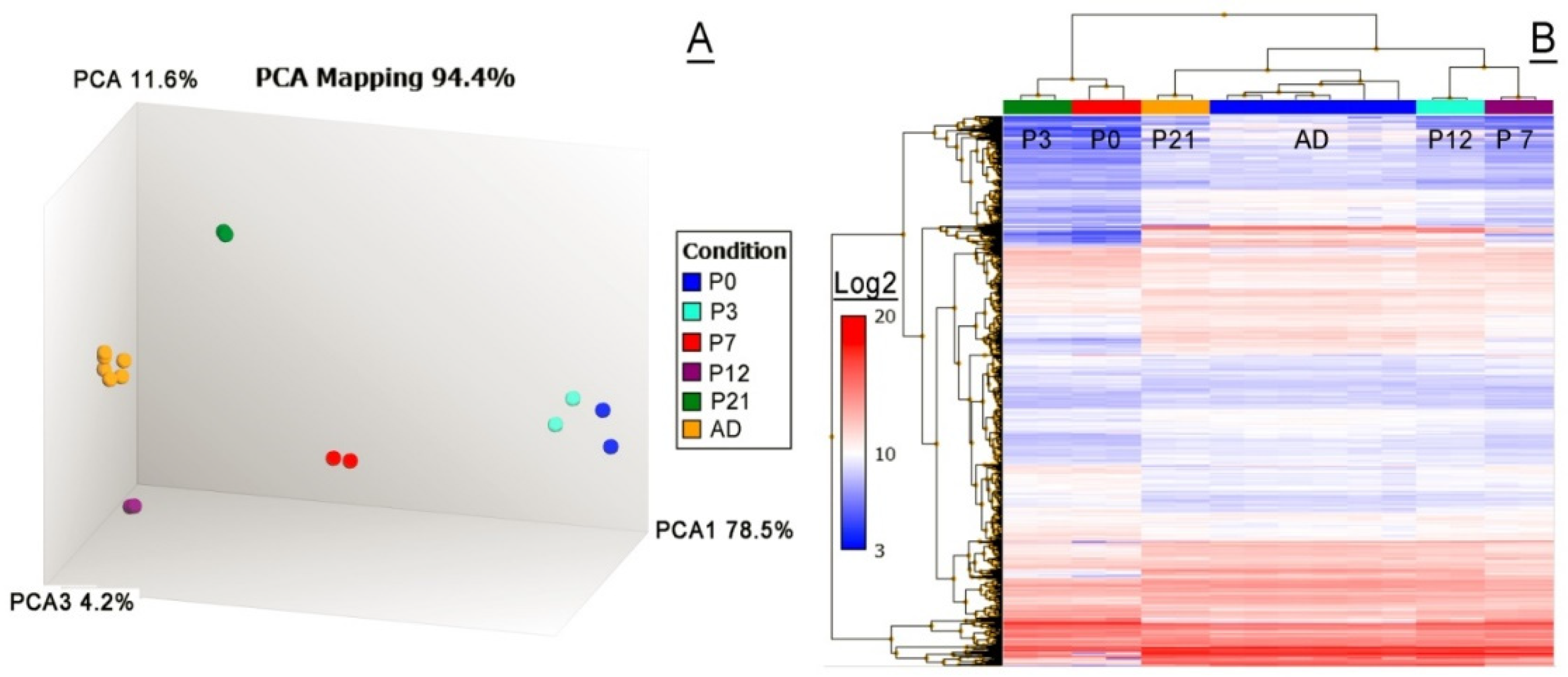
2.2.1. Untargeted Transcriptomic Analyses
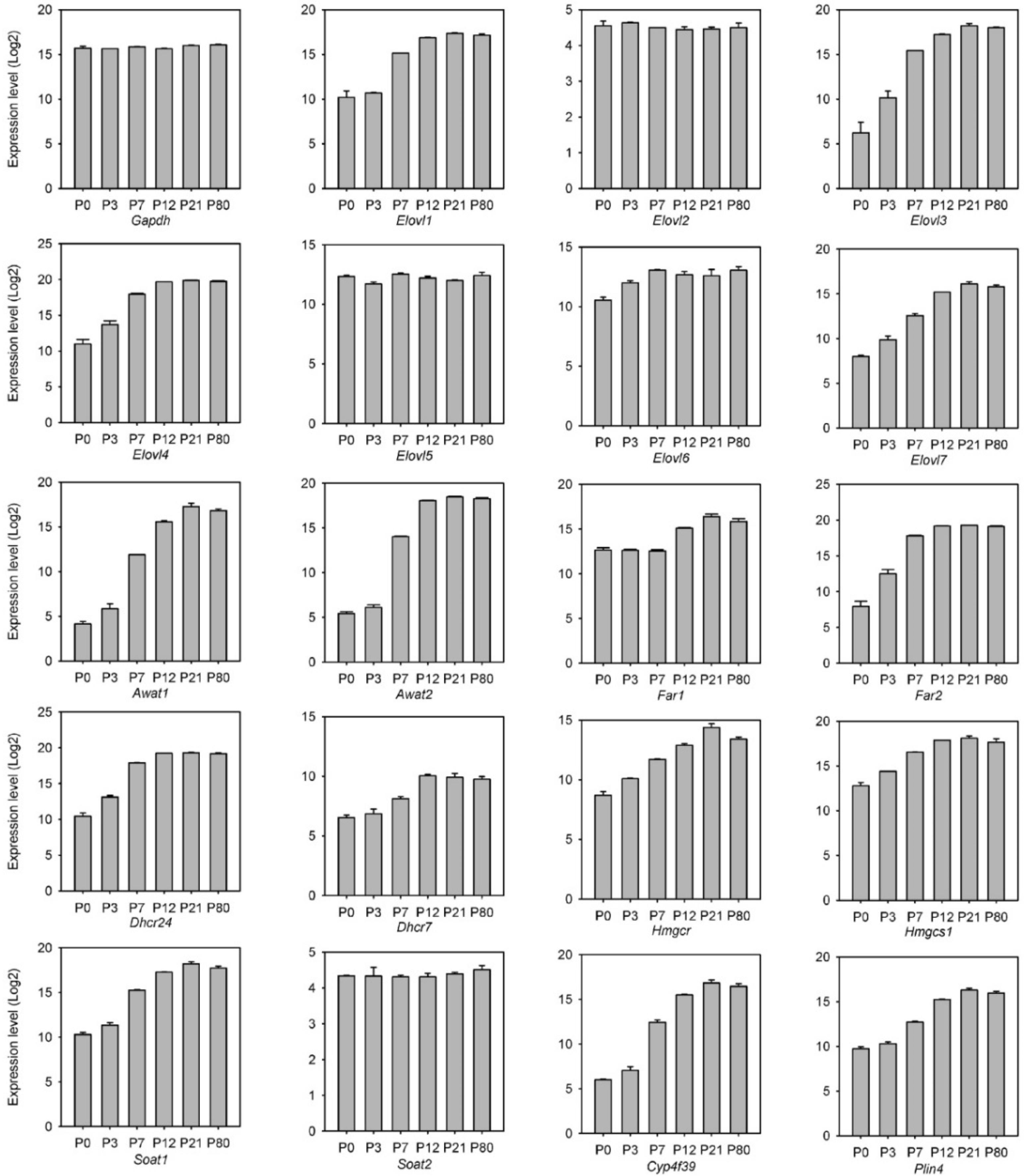
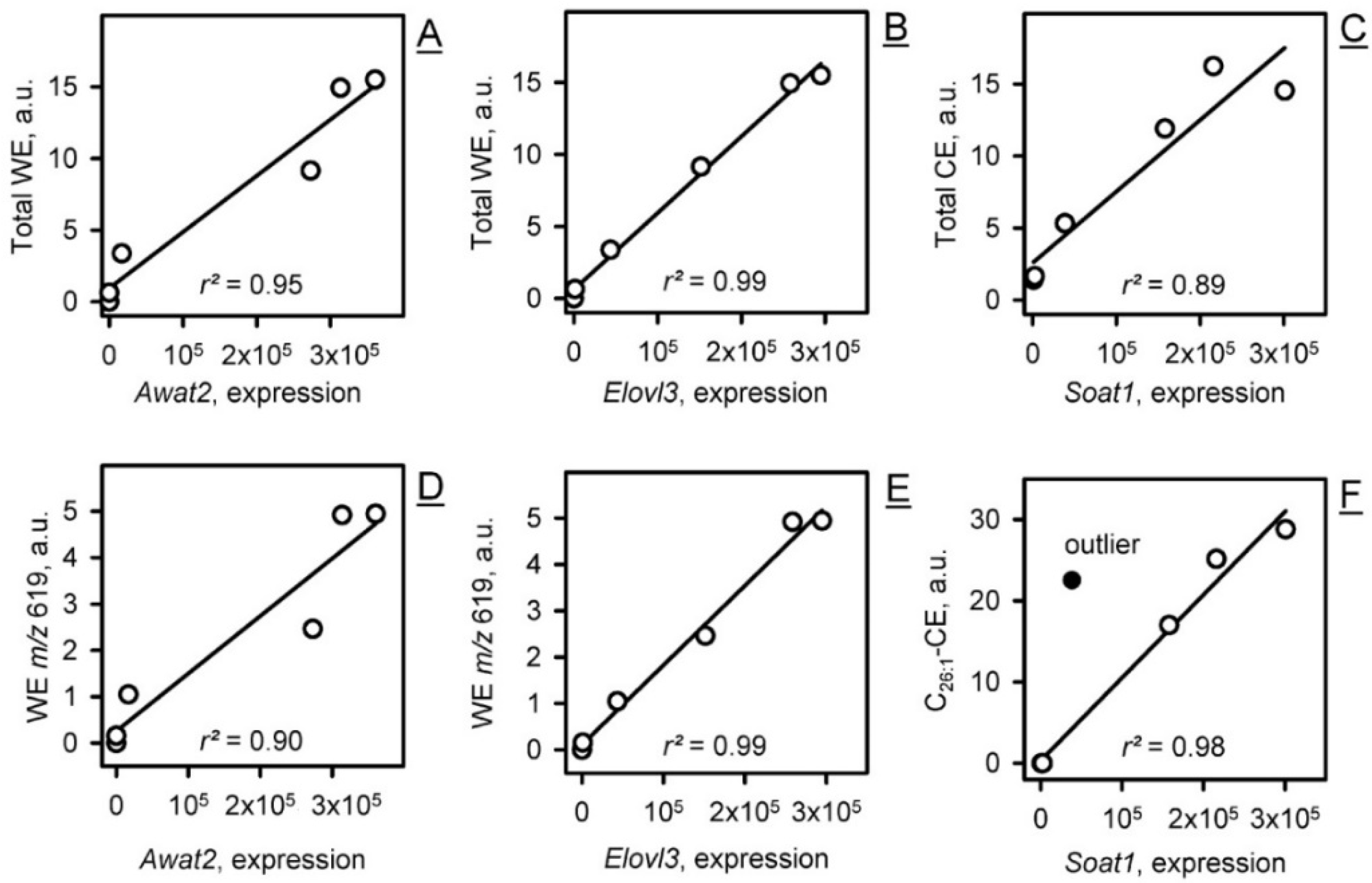
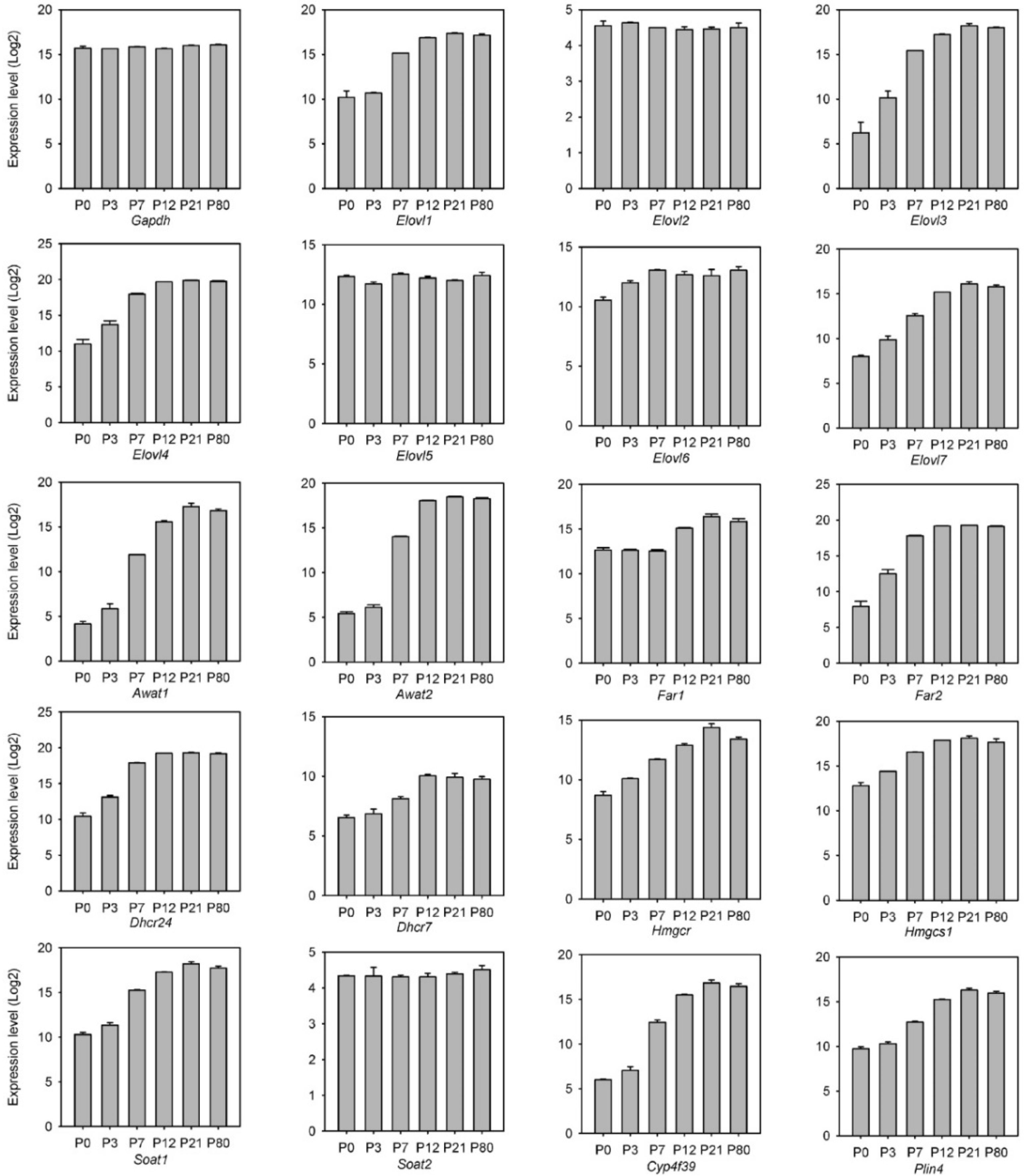
2.2.2. Transcriptomic Analysis of the Genes of Meibogenesis
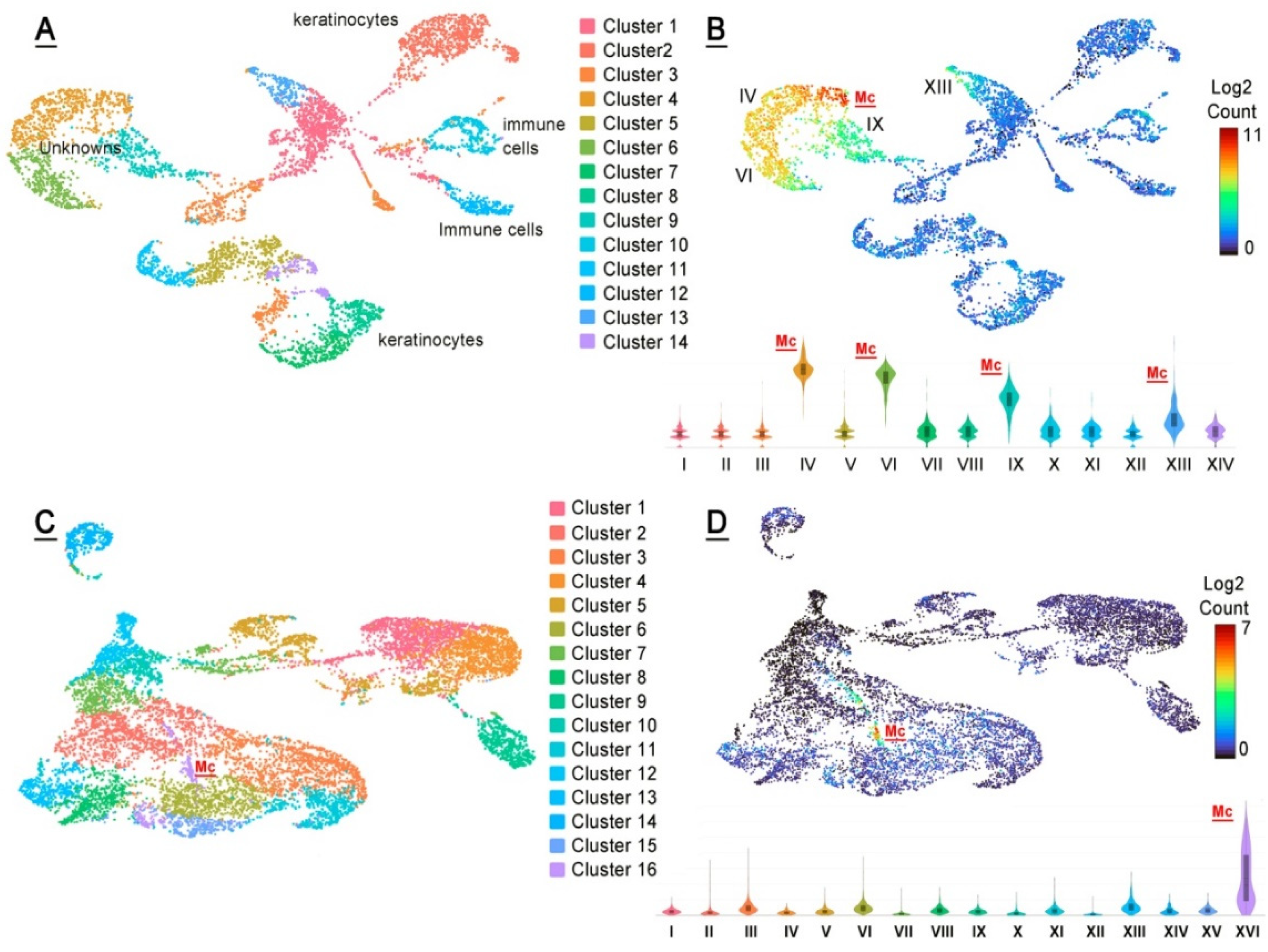
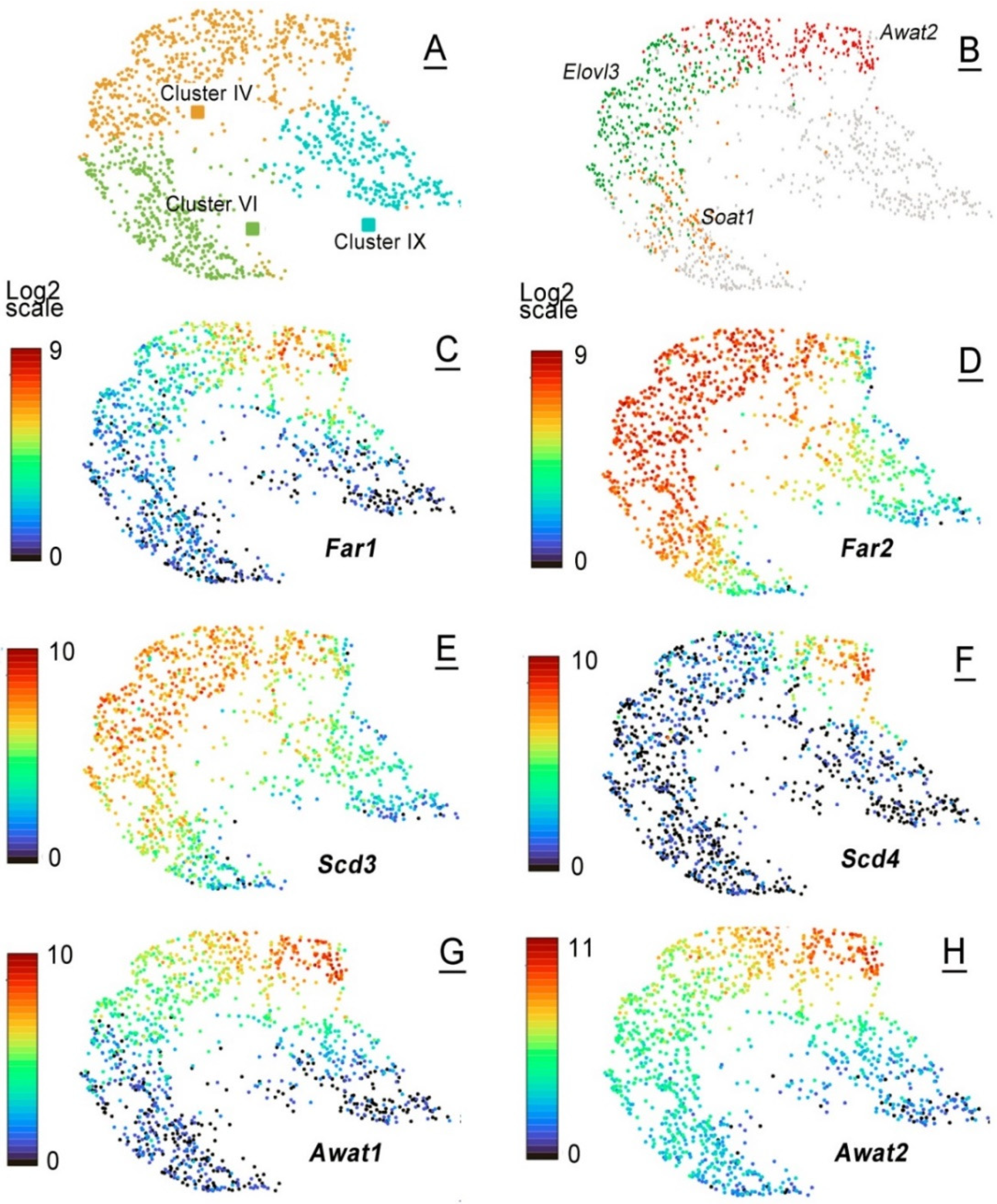
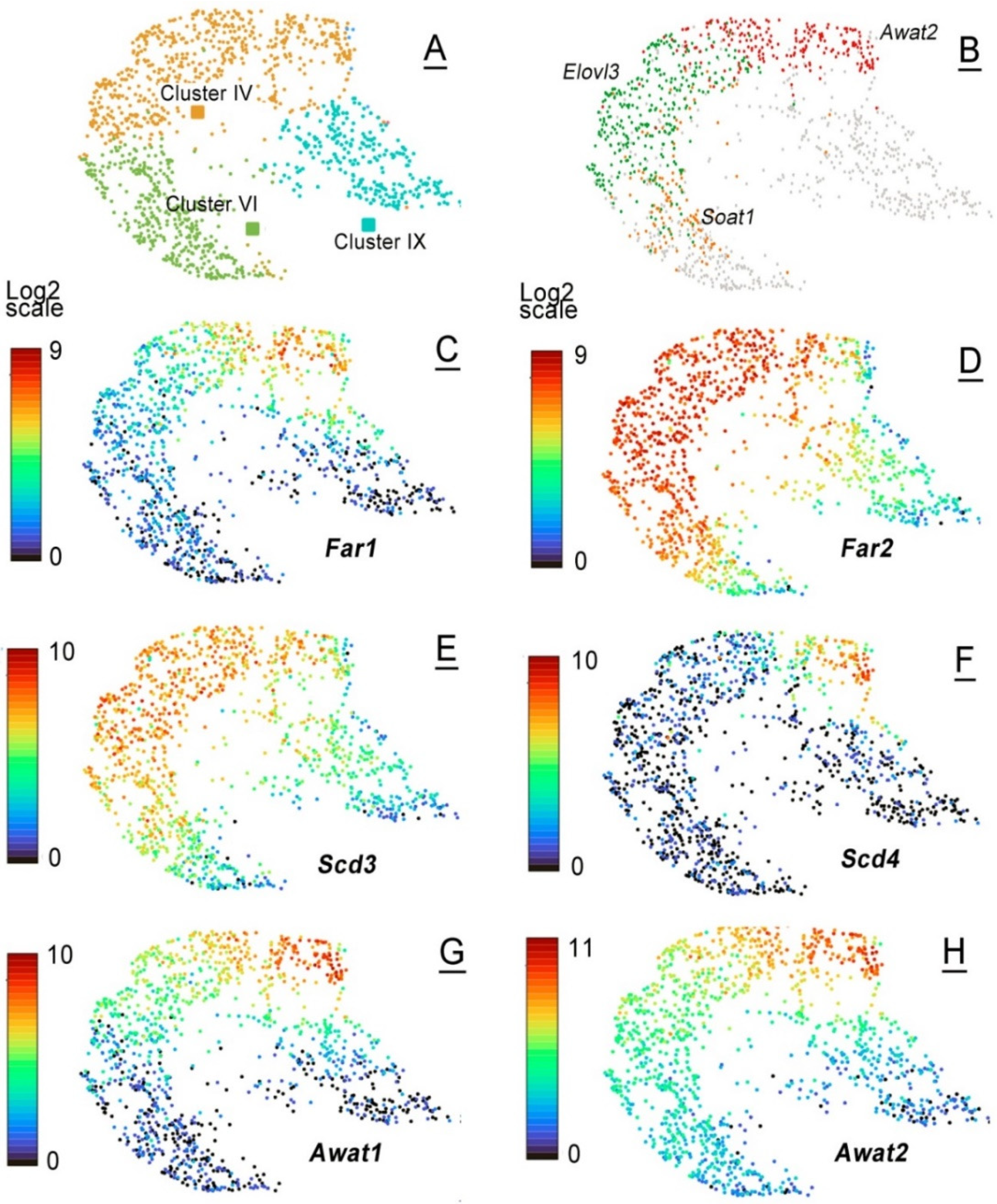
2.3. Single-Cell mRNA Analyses of Mouse MGs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Cluster IV (AD), MNA | Cluster VI (AD), MNA | Cluster IX (AD), MNA | Cluster XIII (AD), MNA | Cluster XVI (P7), MNA |
|---|---|---|---|---|---|
| Scd1 | 217.4 | 197.7 | 197.7 | 107.6 | 34.0 |
| Elovl1 | 100.4 | 41.2 | 40.1 | 57.9 | 2.8 |
| Elovl4 | 52.2 | 19.0 | 16.1 | 44.4 | 3.1 |
| Far2 | 38.7 | 23.1 | 18.3 | 16.7 | 5.8 |
| Msmo1 | 37.9 | 35.2 | 24.3 | 19.4 | 14.3 |
| Awat2 | 33.3 | 1.9 | 5.0 | 53.2 | 0.8 |
| Dhcr24 | 31.1 | 77.5 | 73.5 | 12.6 | 15.6 |
| Scd3 | 30.6 | 12.6 | 14.8 | 15.3 | 3.5 |
| Elovl3 | 30.5 | 16.4 | 13.0 | 25.3 | 8.5 |
| Soat1 | 25.2 | 22.3 | 21.6 | 8.1 | 10.4 |
| Hacd2 | 23.7 | 17.1 | 10.0 | 16.2 | 4.2 |
| Plin2 | 20.9 | 9.5 | 10.6 | 35.7 | 4.3 |
| Awat1 | 18.3 | 0.3 | 2.6 | 18.2 | 0.6 |
| Acot1 | 16.8 | 6.3 | 3.8 | 13.1 | 3.0 |
| Sdr16c5 | 15.3 | 11.3 | 8.5 | 10.3 | 4.0 |
| Dgat2 | 14.7 | 15.3 | 9.0 | 6.6 | 2.7 |
| Hacl1 | 13.1 | 6.3 | 5.2 | 10.5 | 2.9 |
| Hmgcs1 | 10.4 | 27.6 | 23.1 | 6.6 | 15.2 |
| Dhrs7 | 10.4 | 8.0 | 5.6 | 8.2 | 8.0 |
| Scd4 | 6.6 | 0.1 | 1.8 | 15.8 | <0.01 |
| Far1 | 6.4 | 0.3 | 2.8 | 4.7 | 0.2 |
| Phyh | 5.9 | 1.7 | 1.2 | 4.3 | 0.2 |
| Hsd17b2 | 5.2 | 0.6 | 0.9 | 4.4 | 0.8 |
| Acadm | 5.2 | 5.0 | 2.8 | 2.7 | 1.7 |
| Acaa1b | 5.0 | 0.5 | 1.4 | 8.1 | 0.6 |
| Hsd3b6 | 3.7 | 0.4 | 0.8 | 2.1 | 0.6 |
| Fasn | 3.1 | 8.2 | 8.5 | 1.2 | 3.7 |
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals
4.3. Transcriptomic Analyses of Mouse Tissues
4.4. Single Cell RNA Sequencing
4.5. Lipidomic Analyses of Mouse Tarsal Plates
4.6. Data Computation and Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Butovich, I.A.; Bhat, N.; Wojtowicz, J.C. Comparative Transcriptomic and Lipidomic Analyses of Human Male and Female Meibomian Glands Reveal Common Signature Genes of Meibogenesis. Int. J. Mol. Sci. 2019, 20, 4539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A.; McMahon, A.; Wojtowicz, J.C.; Bhat, N.; Wilkerson, A. Effects of sex (or lack thereof) on meibogenesis in mice (Mus musculus): Comparative evaluation of lipidomes and transcriptomes of male and female tarsal plates. Ocul. Surf. 2019, 17, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.J.; Liu, Q.; Chen, X.K.; Xiong, H.B.; Ke, N.; Chen, L. Influence of Vitamin A deficiency on the transcriptomic profile of rat meibomian glands. Int. Ophthalmol. 2021, 41, 1455–1465. [Google Scholar] [CrossRef]
- Butovich, I.A.; McMahon, A.; Wojtowicz, J.C.; Lin, F.; Mancini, R.; Itani, K. Dissecting lipid metabolism in meibomian glands of humans and mice: An integrative study reveals a network of metabolic reactions not duplicated in other tissues. Biochim. Biophys. Acta 2016, 1861, 538–553. [Google Scholar] [CrossRef] [Green Version]
- Swamynathan, S.K. Ocular surface development and gene expression. J. Ophthalmol. 2013, 2013, 103947. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A. Meibomian glands, meibum, and meibogenesis. Exp. Eye Res. 2017, 163, 2–16. [Google Scholar] [CrossRef]
- Sassa, T.; Tadaki, M.; Kiyonari, H.; Kihara, A. Very long-chain tear film lipids produced by fatty acid elongase ELOVL1 prevent dry eye disease in mice. FASEB J. 2018, 32, 2966–2978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, A.; Lu, H.; Butovich, I.A. A role for ELOVL4 in the mouse meibomian gland and sebocyte cell biology. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2832–2840. [Google Scholar]
- Butovich, I.A.; Wilkerson, A.; Bhat, N.; McMahon, A.; Yuksel, S. On the pivotal role of Elovl3/ELOVL3 in meibogenesis and ocular physiology of mice. FASEB J. 2019, 33, 10034–10048. [Google Scholar] [CrossRef]
- Wilkerson, A.; Bhat, N.; McMahon, A.; Butovich, I.A. Elongase of very long chain fatty acids-3 (ELOVL3) is critical for meibum biosynthesis in mice. In ARVO-2019; Investigative Ophthalmology & Visual Science: Vancouver, BC, Canada, 2019; Volume 60, p. 4194. [Google Scholar]
- Tanno, H.; Sassa, T.; Sawai, M.; Kihara, A. Production of branched-chain very-long-chain fatty acids by fatty acid elongases and their tissue distribution in mammals. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158842. [Google Scholar] [CrossRef]
- Wilkerson, A.; Bhat, N.; Quoc Hai Pham, H.; Yuksel, S.; Butovich, I. Physiological effects of inactivation and the roles of Elovl3/ELOVL3 in maintaining ocular homeostasis. FASEB J. 2021, 35, e21327. [Google Scholar] [CrossRef] [PubMed]
- McMahon, A.; Yuksel, S.; Bhat, N.; Pham, H.; Wilkerson, A.; Butovich, I.A. Inactivation of Awat2 in mice causes loss of wax ester lipids from meibum. Investig. Ophthalmol. Vis. Sci. 2020, 61, 2632. [Google Scholar]
- Widjaja-Adhi, M.A.K.; Silvaroli, J.A.; Chelstowska, S.; Trischman, T.; Bederman, I.; Sayegh, R.; Golczak, M. Deficiency in Acyl-CoA:Wax Alcohol Acyltransferase 2 causes evaporative dry eye disease by abolishing biosynthesis of wax esters. FASEB J. 2020, 34, 13792–13808. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, K.; Sawai-Ogawa, M.; Kihara, A. Formation of fatty alcohols-components of meibum lipids-by the fatty acyl-CoA reductase FAR2 is essential for dry eye prevention. FASEB J. 2022, 36, e22216. [Google Scholar] [CrossRef]
- Miyamoto, M.; Sassa, T.; Sawai, M.; Kihara, A. Lipid polarity gradient formed by omega-hydroxy lipids in tear film prevents dry eye disease. Elife 2020, 9, e53582. [Google Scholar] [CrossRef]
- McMahon, A.; Yuksel, S.; Wilkerson, A.; Bhat, N.; Butovich, I.A. Defining the role of ELOVL6 in meibogenesis in mice. In ARVO-2019; Investigative Ophthalmology & Visual Science: Vancouver, BC, Canada, 2019; Volume 60, p. 4302. [Google Scholar]
- Butovich, I.A.; Suzuki, T. Effects of Aging on Human Meibum. Investig. Ophthalmol. Vis. Sci. 2021, 62, 23. [Google Scholar] [CrossRef]
- Nien, C.J.; Massei, S.; Lin, G.; Nabavi, C.; Tao, J.; Brown, D.J.; Paugh, J.R.; Jester, J.V. Effects of age and dysfunction on human meibomian glands. Arch. Ophthalmol. 2011, 129, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Parfitt, G.J.; Brown, D.J.; Jester, J.V. Transcriptome analysis of aging mouse meibomian glands. Mol. Vis. 2016, 22, 518–527. [Google Scholar]
- Nien, C.J.; Paugh, J.R.; Massei, S.; Wahlert, A.J.; Kao, W.W.; Jester, J.V. Age-related changes in the meibomian gland. Exp. Eye Res. 2009, 89, 1021–1027. [Google Scholar] [CrossRef] [Green Version]
- Arava, Y.; Wang, Y.; Storey, J.D.; Liu, C.L.; Brown, P.O.; Herschlag, D. Genome-wide analysis of mRNA translation profiles in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2003, 100, 3889–3894. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A.; Lu, H.; McMahon, A.; Eule, J.C. Toward an animal model of the human tear film: Biochemical comparison of the mouse, canine, rabbit, and human meibomian lipidomes. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6881–6896. [Google Scholar] [CrossRef] [PubMed]
- Rho, C.R.; Kim, S.W.; Lane, S.; Gao, F.; Kim, J.; Xie, Y.; Brown, D.J.; Skowronska-Krawczyk, D.; Jester, J.V. Expression of Acyl-CoA wax-alcohol acyltransferase 2 (AWAT2) by human and rabbit meibomian glands and meibocytes. Ocul. Surf. 2022, 23, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Sawai, M.; Watanabe, K.; Tanaka, K.; Kinoshita, W.; Otsuka, K.; Miyamoto, M.; Sassa, T.; Kihara, A. Diverse meibum lipids produced by Awat1 and Awat2 are important for stabilizing tear film and protecting the ocular surface. iScience 2021, 24, 102478. [Google Scholar] [CrossRef]
- Sundberg, J.P.; Shen, T.; Fiehn, O.; Rice, R.H.; Silva, K.A.; Kennedy, V.E.; Gott, N.E.; Dionne, L.A.; Bechtold, L.S.; Murray, S.A.; et al. Sebaceous gland abnormalities in fatty acyl CoA reductase 2 (Far2) null mice result in primary cicatricial alopecia. PLoS ONE 2018, 13, e0205775. [Google Scholar] [CrossRef] [PubMed]
- Butovich, I.A.; Wilkerson, A.; Yuksel, S. Depletion of Cholesteryl Esters Causes Meibomian Gland Dysfunction-Like Symptoms in a Soat1-Null Mouse Model. Int. J. Mol. Sci. 2021, 22, 1583. [Google Scholar] [CrossRef] [PubMed]
- Ooi, K.G.; Rao, A.; Goh, J.S.; Gracie, G.; Cherepanoff, S.; Madigan, M.C.; Watson, S.L. HMG-CoA reductase expression in human eyelid tissue and in a human meibomian gland epithelial cell line. Graefes Arch. Clin. Exp. Ophthalmol. 2019, 257, 785–790. [Google Scholar] [CrossRef]
- Torocsik, D.; Kovacs, D.; Poliska, S.; Szentkereszty-Kovacs, Z.; Lovaszi, M.; Hegyi, K.; Szegedi, A.; Zouboulis, C.C.; Stahle, M. Genome wide analysis of TLR1/2- and TLR4-activated SZ95 sebocytes reveals a complex immune-competence and identifies serum amyloid A as a marker for activated sebaceous glands. PLoS ONE 2018, 13, e0198323. [Google Scholar] [CrossRef]
- Cheng, J.B.; Russell, D.W. Mammalian wax biosynthesis. I. Identification of two fatty acyl-Coenzyme A reductases with different substrate specificities and tissue distributions. J. Biol. Chem. 2004, 279, 37789–37797. [Google Scholar] [CrossRef] [Green Version]
- Jakobsson, A.; Westerberg, R.; Jacobsson, A. Fatty acid elongases in mammals: Their regulation and roles in metabolism. Prog. Lipid Res. 2006, 45, 237–249. [Google Scholar] [CrossRef]
- Kihara, A. Very long-chain fatty acids: Elongation, physiology and related disorders. J. Biochem. 2012, 152, 387–395. [Google Scholar] [CrossRef]
- Butovich, I.A.; Suzuki, T.; Wojtowicz, J.; Bhat, N.; Yuksel, S. Comprehensive profiling of Asian and Caucasian meibomian gland secretions reveals similar lipidomic signatures regardless of ethnicity. Sci. Rep. 2020, 10, 14510. [Google Scholar] [CrossRef] [PubMed]
- Butovich, I.A.; Wilkerson, A.; Yuksel, S. Differential effects of dietary cholesterol and triglycerides on the lipid homeostasis in Meibomian glands. J. Steroid Biochem. Mol. Biol. 2021, 211, 105894. [Google Scholar] [CrossRef] [PubMed]
- Butovich, I.A.; Suzuki, T. Delineating a novel metabolic high triglycerides-low waxes syndrome that affects lipid homeostasis in meibomian and sebaceous glands. Exp. Eye Res. 2020, 199, 108189. [Google Scholar] [CrossRef] [PubMed]
- Boughton, B.; Mackenna, R.M.; Wheatley, V.R.; Wormall, A. Studies of sebum. VIII. Observations on the squalene and cholesterol content and the possible functions of squalene in human sebum. Biochem. J. 1957, 66, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Esler, W.P.; Tesz, G.J.; Hellerstein, M.K.; Beysen, C.; Sivamani, R.; Turner, S.M.; Watkins, S.M.; Amor, P.A.; Carvajal-Gonzalez, S.; Geoly, F.J.; et al. Human sebum requires de novo lipogenesis, which is increased in acne vulgaris and suppressed by acetyl-CoA carboxylase inhibition. Sci. Transl. Med. 2019, 11, eaau8465. [Google Scholar] [CrossRef]
- Robosky, L.C.; Wade, K.; Woolson, D.; Baker, J.D.; Manning, M.L.; Gage, D.A.; Reily, M.D. Quantitative evaluation of sebum lipid components with nuclear magnetic resonance. J. Lipid Res. 2008, 49, 686–692. [Google Scholar] [CrossRef] [Green Version]
- Lam, S.M.; Tong, L.; Yong, S.S.; Li, B.; Chaurasia, S.S.; Shui, G.; Wenk, M.R. Meibum lipid composition in Asians with dry eye disease. PLoS ONE 2011, 6, e24339. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Green, K.B.; Nichols, K.K. Compositional Analysis of Wax Esters in Human Meibomian Gland Secretions by Direct Infusion Electrospray Ionization Mass Spectrometry. Lipids 2016, 51, 1269–1287. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.H.; Kunnen, C.M.; Duchoslav, E.; Dolla, N.K.; Kelso, M.J.; Papas, E.B.; Lazon de la Jara, P.; Willcox, M.D.; Blanksby, S.J.; Mitchell, T.W. A comparison of patient matched meibum and tear lipidomes. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7417–7424. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butovich, I.A.; Wilkerson, A. Dynamic Changes in the Gene Expression Patterns and Lipid Profiles in the Developing and Maturing Meibomian Glands. Int. J. Mol. Sci. 2022, 23, 7884. https://doi.org/10.3390/ijms23147884
Butovich IA, Wilkerson A. Dynamic Changes in the Gene Expression Patterns and Lipid Profiles in the Developing and Maturing Meibomian Glands. International Journal of Molecular Sciences. 2022; 23(14):7884. https://doi.org/10.3390/ijms23147884
Chicago/Turabian StyleButovich, Igor A., and Amber Wilkerson. 2022. "Dynamic Changes in the Gene Expression Patterns and Lipid Profiles in the Developing and Maturing Meibomian Glands" International Journal of Molecular Sciences 23, no. 14: 7884. https://doi.org/10.3390/ijms23147884
APA StyleButovich, I. A., & Wilkerson, A. (2022). Dynamic Changes in the Gene Expression Patterns and Lipid Profiles in the Developing and Maturing Meibomian Glands. International Journal of Molecular Sciences, 23(14), 7884. https://doi.org/10.3390/ijms23147884