
The MexJK Multidrug Efflux Pump Is Not Involved in Acquired or Intrinsic Antibiotic Resistance in Pseudomonas aeruginosa, but Modulates the Bacterial Quorum Sensing Response


Abstract
:1. Introduction
2. Results and Discussion
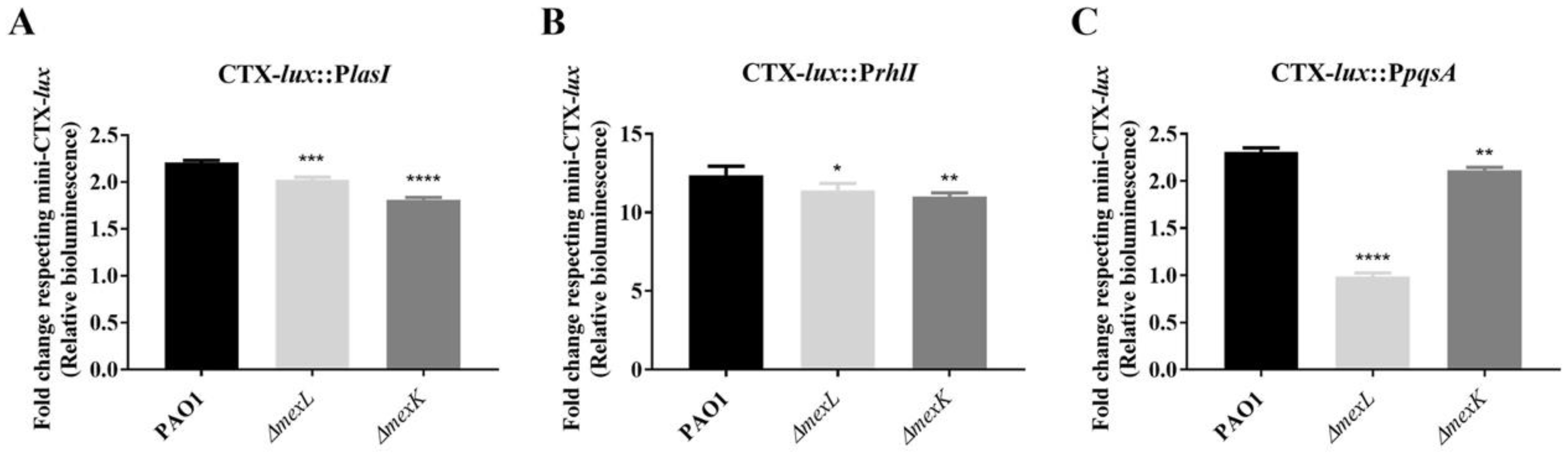
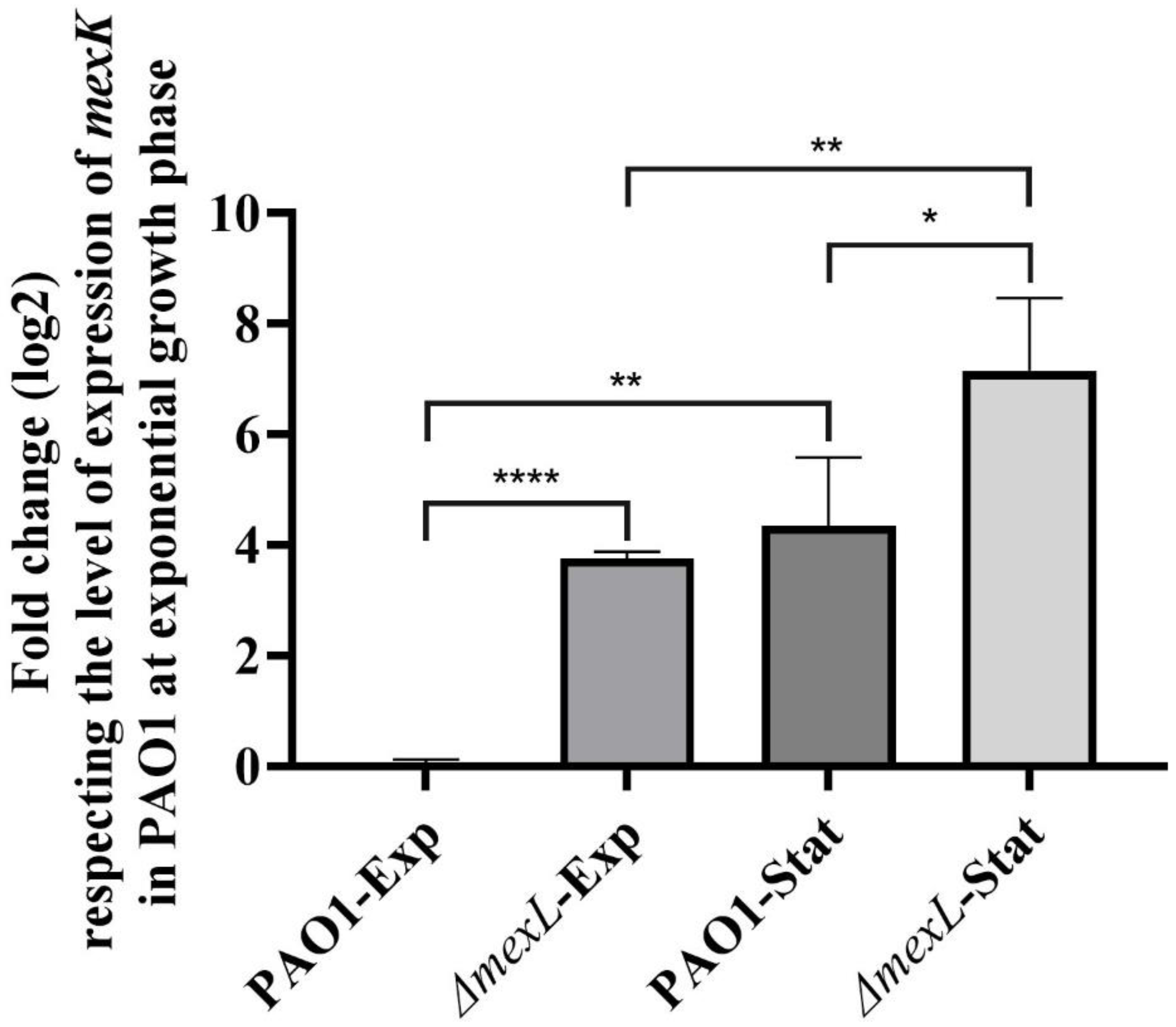
2.1. The Expression of MexJK Is Regulated by Both MexL and Growth Phase
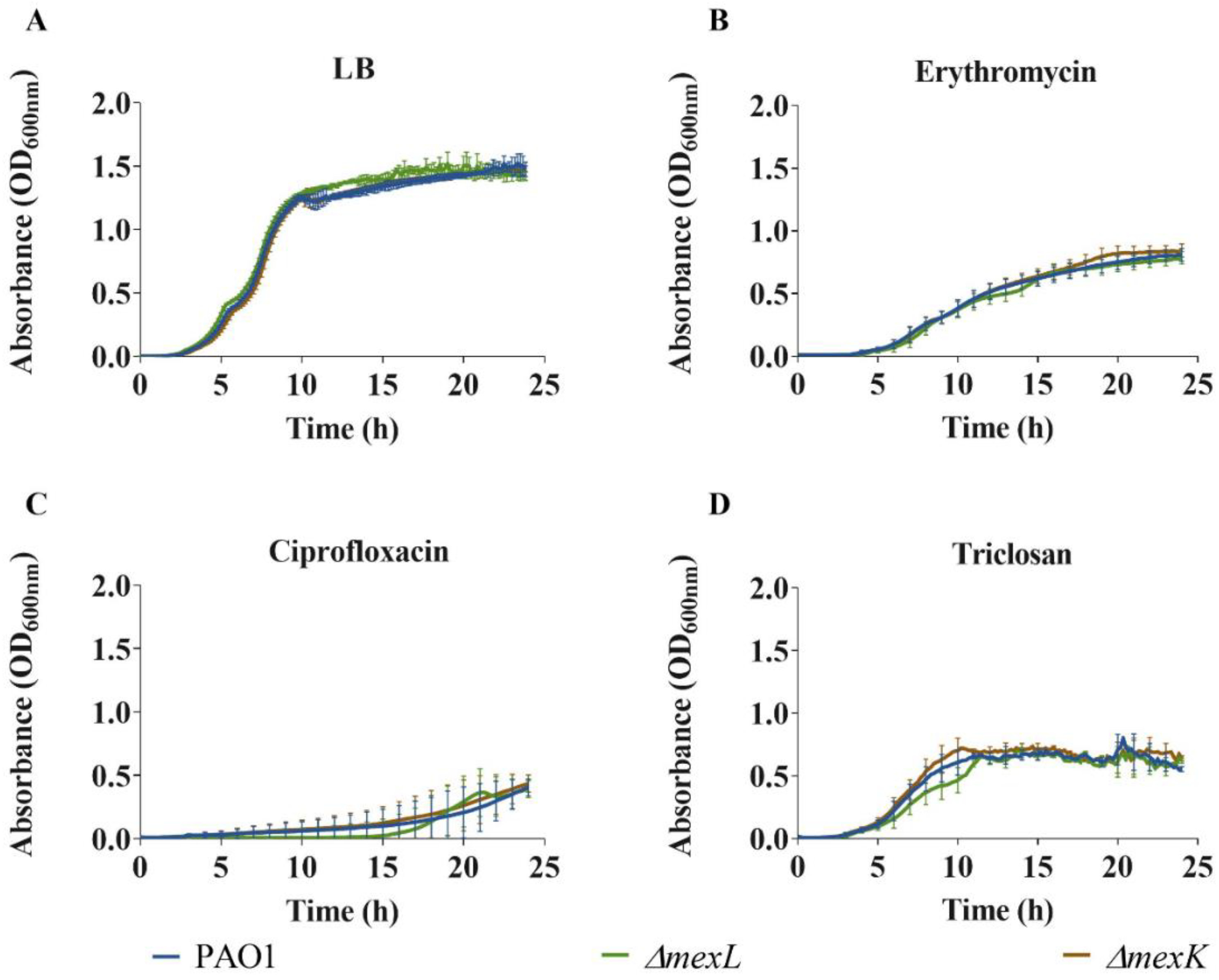
2.2. Overexpression of MexJK Efflux System Does Not Change P. aeruginosa Antibiotics’ Susceptibility Profile
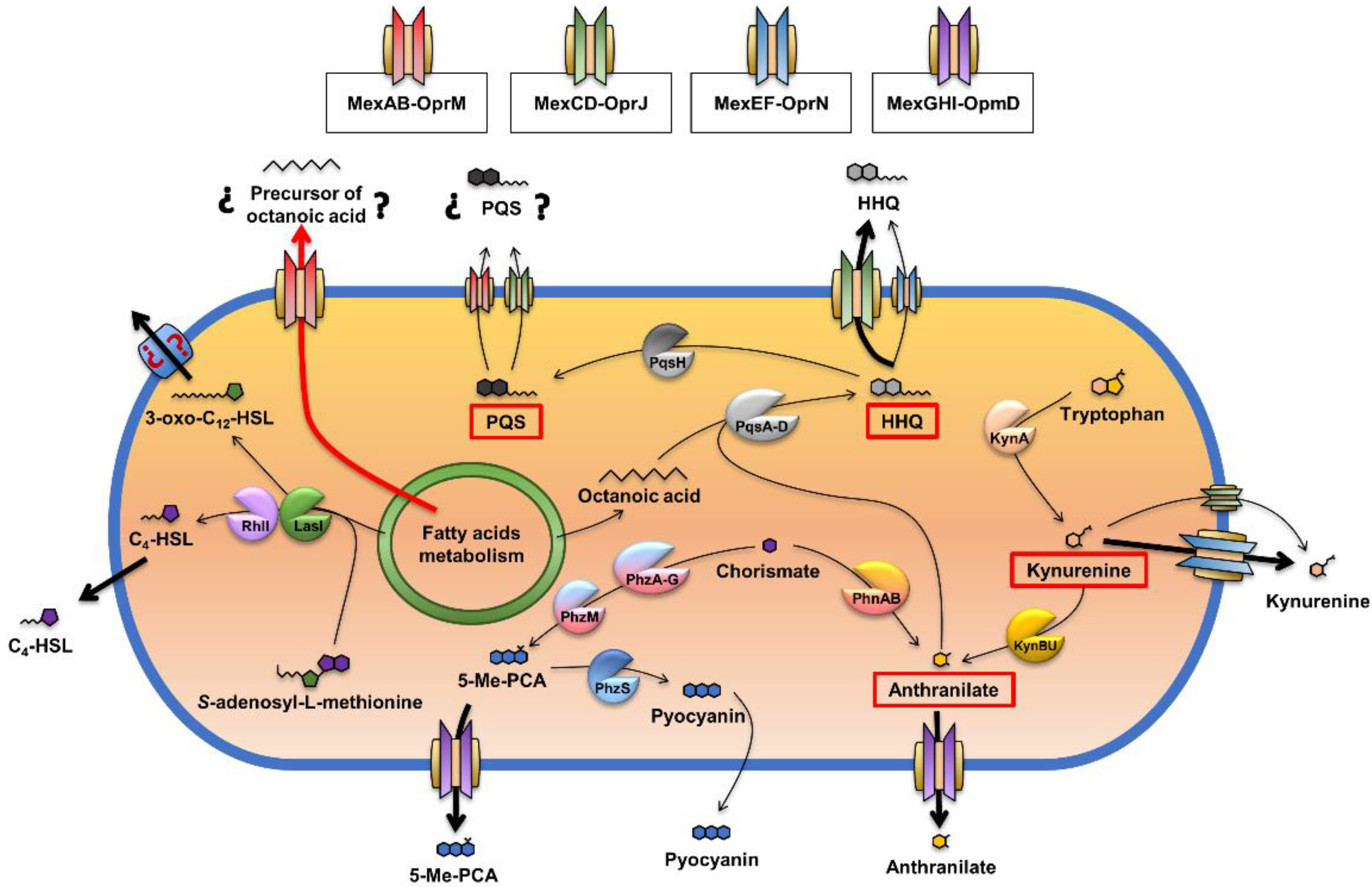
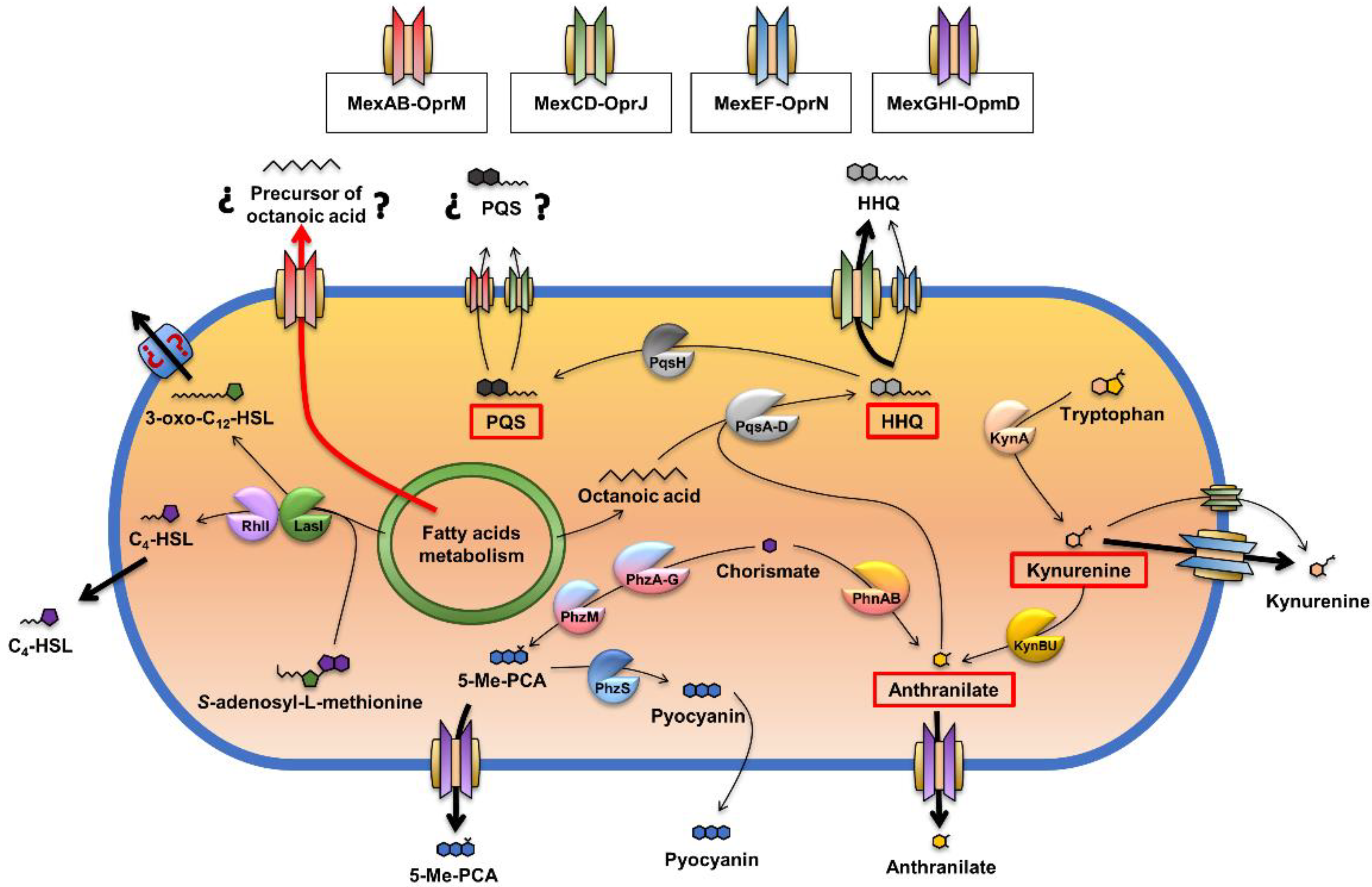
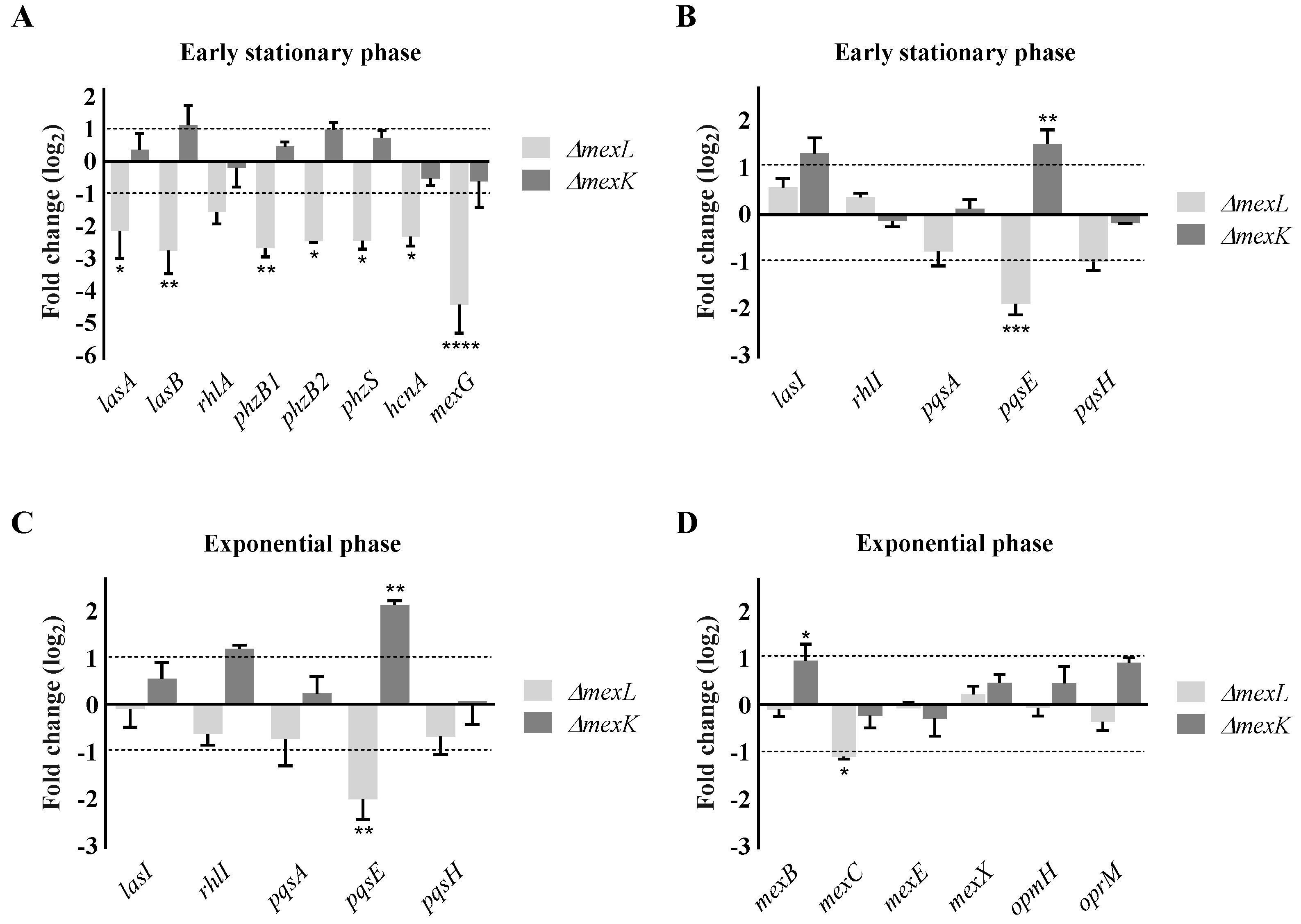
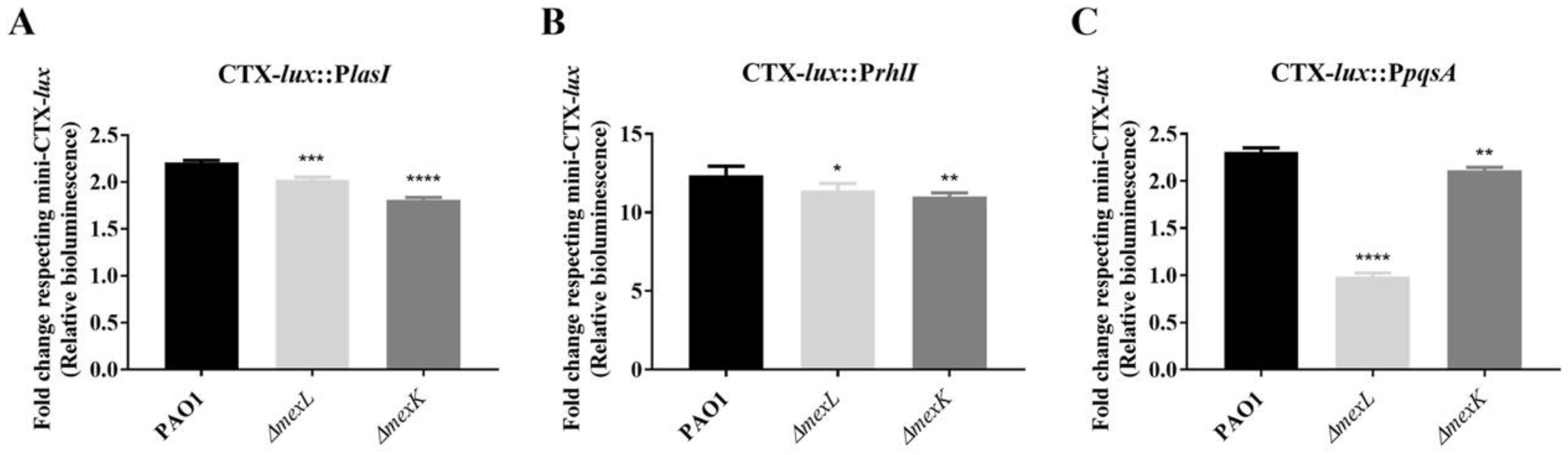
2.3. The Overexpression of the MexJK Efflux System Modifies the Expression of Several Genes Related to the P. aeruginosa QS Response
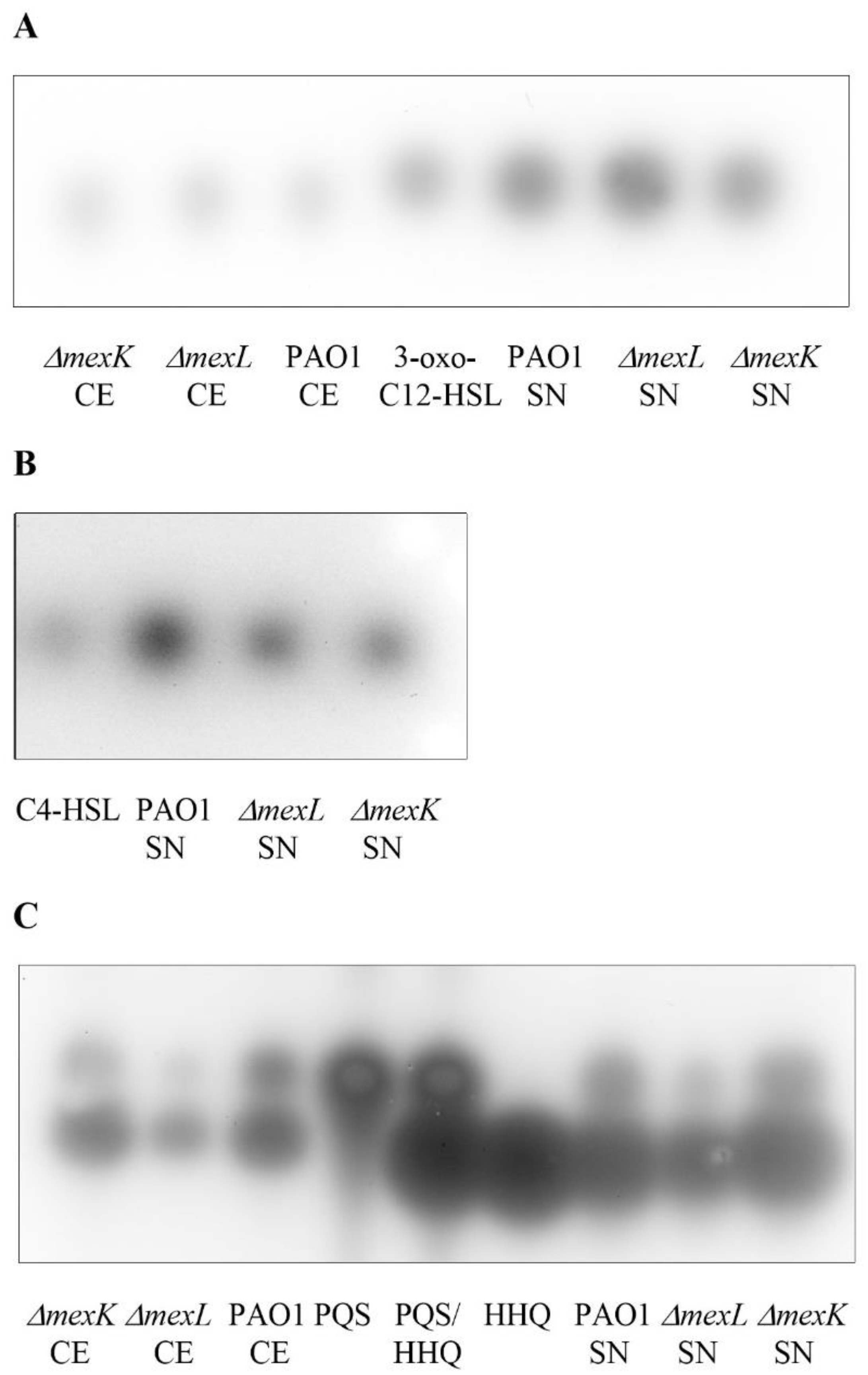
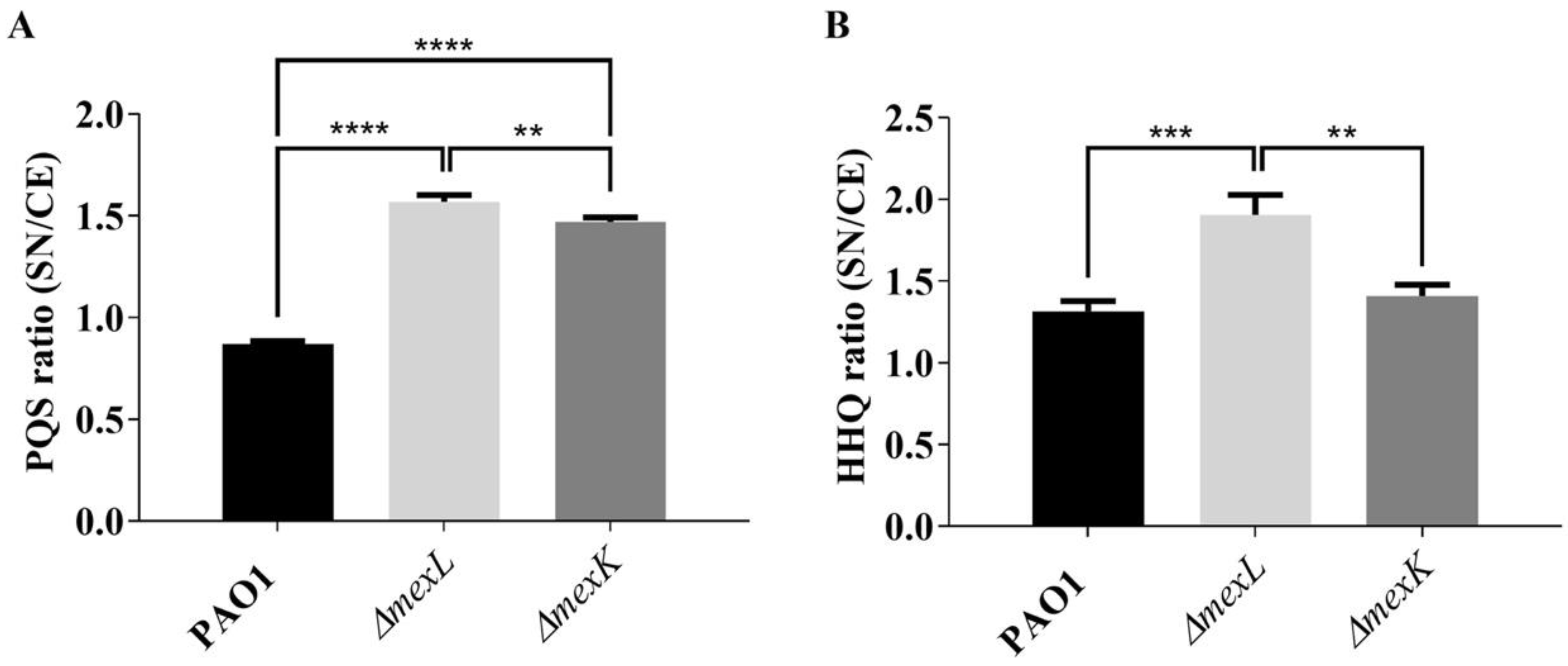
2.4. The Overexpression of the MexJK Efflux System Is Associated with a Decreased Production of PQS and HHQ Autoinducer Compounds
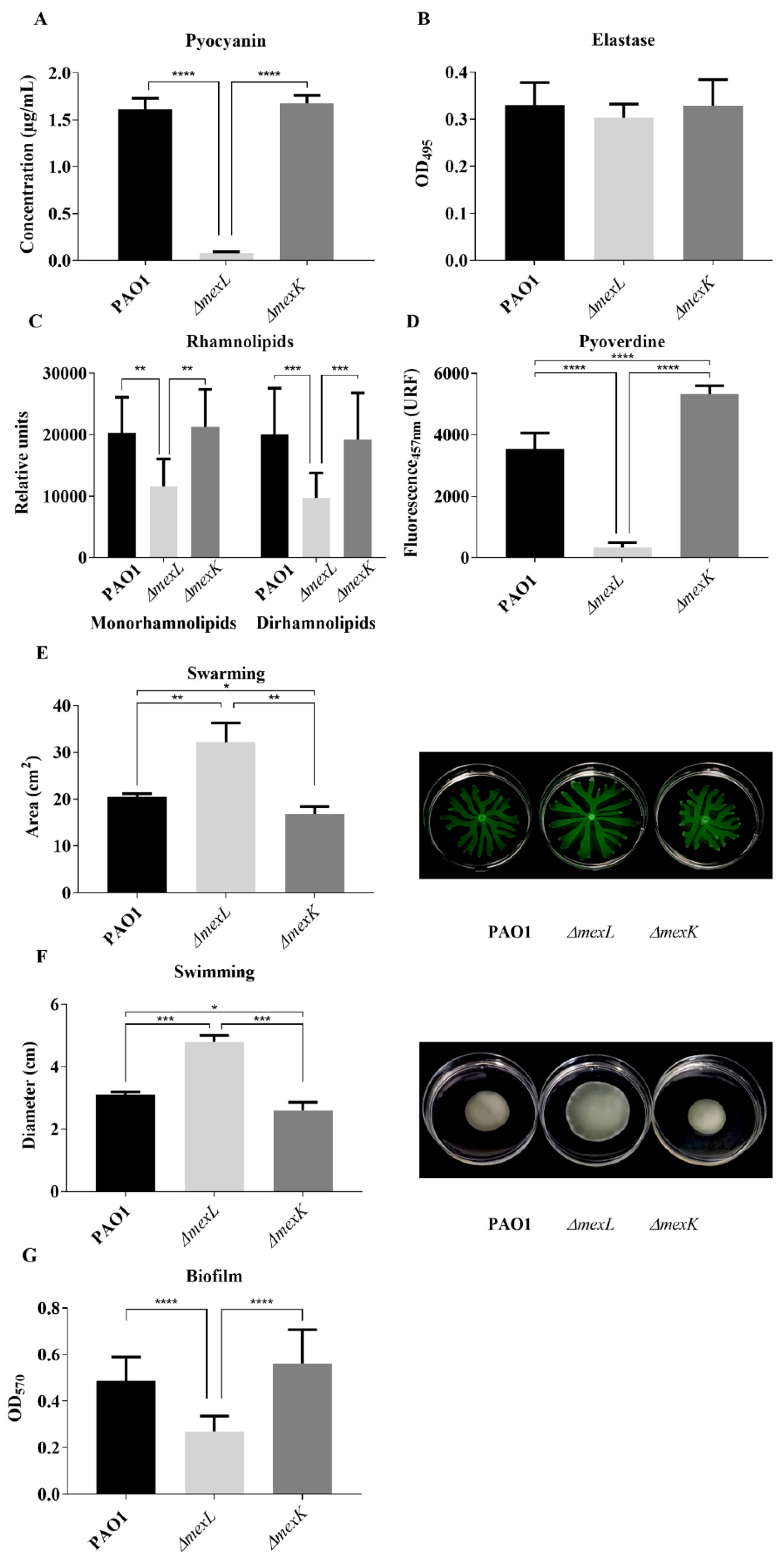
2.5. Differential Activity of MexJK Efflux System Alters the Production of QS-Regulated Virulence Factors
3. Materials and Methods
3.1. Bacterial Strains, Plasmids, and Primers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains/Plasmids | Description | Reference/Origin |
|---|---|---|
| Pseudomonas aeruginosa | ||
| P. aeruginosa PAO1-Lausanne (PAO1) | Wild-type PAO1-L strain | Lab collection |
| P. aeruginosa ΔmexL | PAO1 strain which overproduces the MexJK efflux system by partial deletion of the mexL gene | Present work |
| P. aeruginosa ΔmexK | PAO1 strain with an inactive MexJK efflux system by partial deletion of the mexK gene | Present work |
| PAO1 CTX::PpqsA-lux::pqsA (PqsR-based biosensor) | Biosensor strain used for detecting AQs produced by P. aeruginosa strains | [72] |
| P. aeruginosa PAO1-L miniCTX-lux | PAO1 strain with the construction miniCTX-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa ΔmexL miniCTX-lux | ΔmexL strain with the construction miniCTX-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa ΔmexK miniCTX-lux | ΔmexK strain with the construction miniCTX-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa PAO1-L CTX-lux::PlasI | PAO1 strain with the construction miniCTX::PlasI-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa ΔmexL CTX-lux::PlasI | ΔmexL strain with the construction miniCTX::PlasI-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa ΔmexK CTX-lux::PlasI | ΔmexK strain with the construction miniCTX::PlasI-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa PAO1-L CTX-lux::PrhlI | PAO1 strain with the construction miniCTX::PrhlI-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa ΔmexL CTX-lux::PrhlI | ΔmexL strain with the construction miniCTX::PrhlI-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa ΔmexK CTX-lux::PrhlI | ΔmexK strain with the construction miniCTX::PrhlI-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa PAO1-L CTX-lux::PpqsA | PAO1 strain with the construction miniCTX::PpqsA-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa ΔmexL CTX-lux::PpqsA | ΔmexL strain with the construction miniCTX::PpqsA-lux inserted in a neutral site of the chromosome | Present work |
| P. aeruginosa ΔmexK CTX-lux::PpqsA | ΔmexK strain with the construction miniCTX::PpqsA-lux inserted in a neutral site of the chromosome | Present work |
| Escherichia coli | ||
| S17 λpir | Strain used for transferring plasmids in conjugation assays: F− thi pro hsdR hsdM+ recA RP42-Tc::Mu-Km::Tn7 | [76] |
| S17 λpir (miniCTX-lux) | S17 λpir strain used for transferring the plasmid Mini-CTX-lux in conjugation assays | [72] |
| S17 λpir (miniCTX::PlasI-lux) | S17 λpir strain used for transferring the plasmid miniCTX::PlasI-lux in conjugation assays | Borrowed from Miguel Cámara |
| S17 λpir (miniCTX::PrhlI-lux) | S17 λpir strain used for transferring the plasmid miniCTX::PrhlI-lux in conjugation assays | [24] |
| S17 λpir (miniCTX::PpqsA-lux) | S17 λpir strain used for transferring the plasmid miniCTX::PpqsA-lux in conjugation assays | [72] |
| S17 λpir (pEX18-Ap-ΔmexL) | S17 λpir strain used for transferring the plasmid pEX18-Ap_ΔmexL in conjugation assays | Present work |
| S17 λpir (pEX18-Ap-ΔmexK) | S17 λpir strain used for transferring the plasmid pEX18-Ap_ΔmexK in conjugation assays | Present work |
| One Shot OmniMax™ 2 T1 | Host strain used for the maintenance of cloning plasmids: F′ {proAB lacIq lacZΔM15 Tn10(TetR) Δ(ccdAB) mcrA, Δ(mrr,hsdRMS-mcrBC) ɸ80(lacZ)ΔM15 Δ(lacZYA-argF)U169 endA1 recA1 supE44 thi-1 gyrA96 relA1 tonA panD | Invitrogen |
| OmniMax™ (pEX18-Ap-ΔmexL) | One Shot OmniMax™ 2 T1 harboring the plasmid pEX18-Ap_ΔmexL | Present work |
| OmniMax™ (pEX18-Ap-ΔmexK) | One Shot OmniMax™ 2 T1 harboring the plasmid pEX18-Ap_ΔmexK | Present work |
| JM109-pSB1142 (LasR-based biosensor) | Biosensor strain used for detecting 3-oxo-C12-HSL produced by P. aeruginosa strains | [77] |
| JM109-pSB536 (RhlR-based biosensor) | Biosensor strain used for detecting C4-HSL produced by P. aeruginosa strains | [78] |
| Plasmids | ||
| miniCTX-lux | Reporter plasmid with the luxCDABE operon. TcR | [79] |
| miniCTX::PlasI-lux | Reporter plasmid with the luxCDABE operon under the control of PlasI promoter. TcR | Borrowed from Miguel Cámara |
| miniCTX::PrhlI-lux | Reporter plasmid with the luxCDABE operon under the control of PrhlI promoter. TcR | [24] |
| miniCTX::PpqsA-lux | Reporter plasmid with the luxCDABE operon under the control of PpqsA promoter. TcR | [72] |
| pGEM-T Easy | Commercial plasmid “pGEM-T Easy Vector” used for cloning optimization of PCR products. AmpR | Promega |
| pGEM-T Easy-ΔmexL | Commercial plasmid “pGEM-T Easy Vector” (Promega) used for cloning optimization of the flanking region of mexL gene. AmpR | Present work |
| pGEM-T Easy-ΔmexK | Commercial plasmid “pGEM-T Easy Vector” used for cloning optimization of the flanking region of mexK gene. AmpR | Present work |
| pEX18-Ap | Plasmid with conjugative properties used for deleting genes in P. aeruginosa by homologous recombination. AmpR | [80] |
| pEX18-Ap-ΔmexL | Plasmid with conjugative properties containing “Up and down mexL insert” used for deleting mexL gene in P. aeruginosa by homologous recombination. AmpR | Present work |
| pEX18-Ap-ΔmexK | Plasmid with conjugative properties containing “Up and down mexK insert” used for deleting mexK gene in P. aeruginosa by homologous recombination. AmpR | Present work |
3.2. Growth Media, Culture Conditions, and Antibiotic Susceptibility Testing
3.3. Generation of ΔmexL and ΔmexK Mutant Strains
3.4. RNA Preparation and RT-qPCR
3.5. Detection of QS Molecules
3.6. Analysis of the Activation Kinetics of the Promoters PlasI, PrhlI, and PpqsA throughout the Growth Cycle
3.7. Analysis of the Production of QS-Regulated Virulence Factors
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reynolds, D.; Kollef, M. The Epidemiology and Pathogenesis and Treatment of Pseudomonas aeruginosa Infections: An Update. Drugs 2021, 81, 2117–2131. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Solano, L.; Macia, M.D.; Fajardo, A.; Oliver, A.; Martinez, J.L. Chronic Pseudomonas aeruginosa infection in chronic obstructive pulmonary disease. Clin. Infect. Dis. 2008, 47, 1526–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talwalkar, J.S.; Murray, T.S. The Approach to Pseudomonas aeruginosa in Cystic Fibrosis. Clin. Chest Med. 2016, 37, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Sanz-García, F.; Gil-Gil, T.; Laborda, P.; Ochoa-Sánchez, L.E.; Martínez, J.L.; Hernando-Amado, S. Coming from the Wild: Multidrug Resistant Opportunistic Pathogens Presenting a Primary, Not Human-Linked, Environmental Habitat. Int. J. Mol. Sci. 2021, 22, 15. [Google Scholar]
- Laborda, P.; Sanz-García, F.; Hernando-Amado, S.; Martínez, J.L. Pseudomonas aeruginosa: An antibiotic resilient pathogen with environmental origin. Curr. Opin. Microbiol. 2021, 64, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Breidenstein, E.B.; de la Fuente-Nunez, C.; Hancock, R.E. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- Tummler, B.; Wiehlmann, L.; Klockgether, J.; Cramer, N. Advances in understanding Pseudomonas. F1000Prime Rep. 2014, 6, 9. [Google Scholar] [CrossRef]
- Santajit, S.; Indrawattana, N. Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. BioMed Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [Green Version]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2009, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet. Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Schweizer, H.P. Efflux as a mechanism of resistance to antimicrobials in Pseudomonas aeruginosa and related bacteria: Unanswered questions. Genet. Mol. Res. 2003, 2, 48–62. [Google Scholar] [PubMed]
- Nikaido, E.; Shirosaka, I.; Yamaguchi, A.; Nishino, K. Regulation of the AcrAB multidrug efflux pump in Salmonella enterica serovar Typhimurium in response to indole and paraquat. Microbiology 2011, 157 Pt 3, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernando-Amado, S.; Blanco, P.; Alcalde-Rico, M.; Corona, F.; Reales-Calderon, J.A.; Sanchez, M.B.; Martinez, J.L. Multidrug efflux pumps as main players in intrinsic and acquired resistance to antimicrobials. Drug Resist. Updates 2016, 28, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Laborda, P.; Alcalde-Rico, M.; Blanco, P.; Martinez, J.L.; Hernando-Amado, S. Novel Inducers of the Expression of Multidrug Efflux Pumps That Trigger Pseudomonas aeruginosa Transient Antibiotic Resistance. Antimicrob. Agents Chemother. 2019, 63, 11. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Corona, F.; Martinez, J.L. Biolog Phenotype Microarray Is a Tool for the Identification of Multidrug Resistance Efflux Pump Inducers. Antimicrob. Agents Chemother. 2018, 62, 11. [Google Scholar] [CrossRef] [Green Version]
- Blanco, P.; Corona, F.; Sanchez, M.B.; Martinez, J.L. Vitamin K3 Induces the Expression of the Stenotrophomonas maltophilia SmeVWX Multidrug Efflux Pump. Antimicrob. Agents Chemother. 2017, 61, 5. [Google Scholar] [CrossRef] [Green Version]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial Multidrug Efflux Pumps: Much More Than Antibiotic Resistance Determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.L.; Sanchez, M.B.; Martinez-Solano, L.; Hernandez, A.; Garmendia, L.; Fajardo, A.; Alvarez-Ortega, C. Functional role of bacterial multidrug efflux pumps in microbial natural ecosystems. FEMS Microbiol. Rev. 2009, 33, 430–449. [Google Scholar] [CrossRef]
- Alcalde-Rico, M.; Hernando-Amado, S.; Blanco, P.; Martinez, J.L. Multidrug Efflux Pumps at the Crossroad between Antibiotic Resistance and Bacterial Virulence. Front. Microbiol. 2016, 7, 1483. [Google Scholar] [CrossRef] [Green Version]
- Poole, K. Stress responses as determinants of antimicrobial resistance in Gram-negative bacteria. Trends Microbiol. 2012, 20, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Warner, D.M.; Folster, J.P.; Shafer, W.M.; Jerse, A.E. Regulation of the MtrC-MtrD-MtrE efflux-pump system modulates the in vivo fitness of Neisseria gonorrhoeae. J. Infect. Dis. 2007, 196, 1804–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcalde-Rico, M.; Olivares-Pacheco, J.; Alvarez-Ortega, C.; Camara, M.; Martinez, J.L. Role of the Multidrug Resistance Efflux Pump MexCD-OprJ in the Pseudomonas aeruginosa Quorum Sensing Response. Front. Microbiol. 2018, 9, 2752. [Google Scholar] [CrossRef] [PubMed]
- Alcalde-Rico, M.; Olivares-Pacheco, J.; Halliday, N.; Camara, M.; Martinez, J.L. The impaired quorum sensing response of Pseudomonas aeruginosa MexAB-OprM efflux pump overexpressing mutants is not due to non-physiological efflux of 3-oxo-C12-HSL. Env. Microbiol 2020, 22, 5167–5188. [Google Scholar] [CrossRef]
- Lamarche, M.G.; Deziel, E. MexEF-OprN Efflux Pump Exports the Pseudomonas Quinolone Signal (PQS) Precursor HHQ (4-hydroxy-2-heptylquinoline). PLoS ONE 2011, 6, e24310. [Google Scholar] [CrossRef]
- Aedekerk, S.; Diggle, S.P.; Song, Z.; Høiby, N.; Cornelis, P.; Williams, P.; Cámara, M. The MexGHI-OpmD multidrug efflux pump controls growth, antibiotic susceptibility and virulence in Pseudomonas aeruginosa via 4-quinolone-dependent cell-to-cell communication. Microbiology 2005, 151, 1113–1125. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.; Camara, M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: A tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol. 2009, 12, 182–191. [Google Scholar] [CrossRef]
- Williams, P.; Winzer, K.; Chan, W.C.; Camara, M. Look who’s talking: Communication and quorum sensing in the bacterial world. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2007, 362, 1119–1134. [Google Scholar] [CrossRef] [Green Version]
- Williams, P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153 Pt 12, 3923–3938. [Google Scholar] [CrossRef] [Green Version]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Pearson, J.P.; Pesci, E.C.; Iglewski, B.H. Roles of Pseudomonas aeruginosa las and rhl quorum-sensing systems in control of elastase and rhamnolipid biosynthesis genes. J. Bacteriol. 1997, 179, 5756–5767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsek, M.R.; Greenberg, E.P. Quorum sensing signals in development of Pseudomonas aeruginosa biofilms. Methods Enzymol. 1999, 310, 43–55. [Google Scholar] [PubMed]
- Rumbaugh, K.P.; Griswold, J.A.; Iglewski, B.H.; Hamood, A.N. Contribution of quorum sensing to the virulence of Pseudomonas aeruginosa in burn wound infections. Infect. Immun. 1999, 67, 5854–5862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteley, M.; Lee, K.M.; Greenberg, E.P. Identification of genes controlled by quorum sensing in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1999, 96, 13904–13909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winstanley, C.; Fothergill, J.L. The role of quorum sensing in chronic cystic fibrosis Pseudomonas aeruginosa infections. FEMS Microbiol. Lett. 2009, 290, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [Green Version]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Rasamiravaka, T.; El Jaziri, M. Quorum-Sensing Mechanisms and Bacterial Response to Antibiotics in P. aeruginosa. Curr. Microbiol. 2016, 73, 747–753. [Google Scholar] [CrossRef]
- Diggle, S.P.; Cornelis, P.; Williams, P.; Camara, M. 4-quinolone signalling in Pseudomonas aeruginosa: Old molecules, new perspectives. Int. J. Med. Microbiol. IJMM 2006, 296, 83–91. [Google Scholar] [CrossRef]
- Pesci, E.C.; Milbank, J.B.; Pearson, J.P.; McKnight, S.; Kende, A.S.; Greenberg, E.P.; Iglewski, B.H. Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1999, 96, 11229–11234. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Zhang, L. The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 2015, 6, 26–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Park, H.-Y.; Park, S.J.; Park, S.-J.; Kim, S.-K.; Ha, C.; Im, S.-J.; Lee, J.-H. Growth phase-differential quorum sensing regulation of anthranilate metabolism in Pseudomonas aeruginosa. Mol. Cells 2011, 32, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Deziel, E.; Gopalan, S.; Tampakaki, A.P.; Lepine, F.; Padfield, K.E.; Saucier, M.; Xiao, G.; Rahme, L.G. The contribution of MvfR to Pseudomonas aeruginosa pathogenesis and quorum sensing circuitry regulation: Multiple quorum sensing-regulated genes are modulated without affecting lasRI, rhlRI or the production of N-acyl-L-homoserine lactones. Mol. Microbiol. 2005, 55, 998–1014. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Gan, J.; Zhao, J.; Kong, W.; Zhang, J.; Zhu, M.; Li, F.; Song, Y.; Qin, J.; Liang, H. Crystal structure of Pseudomonas aeruginosa RsaL bound to promoter DNA reaffirms its role as a global regulator involved in quorum-sensing. Nucleic Acids Res. 2017, 45, 699–710. [Google Scholar] [CrossRef] [Green Version]
- Groleau, M.C.; de Oliveira Pereira, T.; Dekimpe, V.; Déziel, E. PqsE Is Essential for RhlR-Dependent Quorum Sensing Regulation in Pseudomonas aeruginosa. mSystems 2020, 5, 3. [Google Scholar] [CrossRef]
- Letizia, M.; Mellini, M.; Fortuna, A.; Visca, P.; Imperi, F.; Leoni, L.; Rampioni, G. PqsE Expands and Differentially Modulates the RhlR Quorum Sensing Regulon in Pseudomonas aeruginosa. Microbiol. Spectr. 2022, 10, e0096122. [Google Scholar] [CrossRef]
- Evans, K.; Passador, L.; Srikumar, R.; Tsang, E.; Nezezon, J.; Poole, K. Influence of the MexAB-OprM multidrug efflux system on quorum sensing in Pseudomonas aeruginosa. J. Bacteriol. 1998, 180, 5443–5447. [Google Scholar] [CrossRef] [Green Version]
- Pearson, J.P.; Van Delden, C.; Iglewski, B.H. Active efflux and diffusion are involved in transport of Pseudomonas aeruginosa cell-to-cell signals. J. Bacteriol. 1999, 181, 1203–1210. [Google Scholar] [CrossRef] [Green Version]
- Olivares, J.; Alvarez-Ortega, C.; Linares, J.F.; Rojo, F.; Köhler, T.; Martínez, J.L. Overproduction of the multidrug efflux pump MexEF-OprN does not impair Pseudomonas aeruginosa fitness in competition tests, but produces specific changes in bacterial regulatory networks. Environ. Microbiol. 2012, 14, 1968–1981. [Google Scholar] [CrossRef]
- Sakhtah, H.; Koyama, L.; Zhang, Y.; Morales, D.K.; Fields, B.L.; Price-Whelan, A.; Hogan, D.A.; Shepard, K.; Dietrich, L.E. The Pseudomonas aeruginosa efflux pump MexGHI-OpmD transports a natural phenazine that controls gene expression and biofilm development. Proc. Natl. Acad. Sci. USA 2016, 113, E3538–E3547. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Nikaido, H.; Poole, K. Role of mexA-mexB-oprM in antibiotic efflux in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1995, 39, 1948–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziha-Zarifi, I.; Llanes, C.; Kohler, T.; Pechere, J.C.; Plesiat, P. In vivo emergence of multidrug-resistant mutants of Pseudomonas aeruginosa overexpressing the active efflux system MexA-MexB-OprM. Antimicrob. Agents Chemother. 1999, 43, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Substrate specificities of MexAB-OprM, MexCD-OprJ, and MexXY-oprM efflux pumps in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef] [Green Version]
- Poole, K.; Gotoh, N.; Tsujimoto, H.; Zhao, Q.; Wada, A.; Yamasaki, T.; Neshat, S.; Yamagishi, J.; Li, X.Z.; Nishino, T. Overexpression of the mexC-mexD-oprJ efflux operon in nfxB-type multidrug-resistant strains of Pseudomonas aeruginosa. Mol. Microbiol. 1996, 21, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Kohler, T.; Michea-Hamzehpour, M.; Henze, U.; Gotoh, N.; Curty, L.K.; Pechere, J.C. Characterization of MexE-MexF-OprN, a positively regulated multidrug efflux system of Pseudomonas aeruginosa. Mol. Microbiol. 1997, 23, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Mine, T.; Morita, Y.; Kataoka, A.; Mizushima, T.; Tsuchiya, T. Expression in Escherichia coli of a new multidrug efflux pump, MexXY, from Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1999, 43, 415–417. [Google Scholar] [CrossRef] [Green Version]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Contribution of the MexX-MexY-oprM efflux system to intrinsic resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 2242–2246. [Google Scholar] [CrossRef] [Green Version]
- Chuanchuen, R.; Murata, T.; Gotoh, N.; Schweizer, H.P. Substrate-dependent utilization of OprM or OpmH by the Pseudomonas aeruginosa MexJK efflux pump. Antimicrob. Agents Chemother. 2005, 49, 2133–2136. [Google Scholar] [CrossRef] [Green Version]
- Chuanchuen, R.; Narasaki, C.T.; Schweizer, H.P. The MexJK efflux pump of Pseudomonas aeruginosa requires OprM for antibiotic efflux but not for efflux of triclosan. J. Bacteriol. 2002, 184, 5036–5044. [Google Scholar] [CrossRef] [Green Version]
- Chuanchuen, R.; Gaynor, J.B.; Karkhoff-Schweizer, R.; Schweizer, H.P. Molecular characterization of MexL, the transcriptional repressor of the mexJK multidrug efflux operon in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2005, 49, 1844–1851. [Google Scholar] [CrossRef] [Green Version]
- Schuster, M.; Lostroh, C.P.; Ogi, T.; Greenberg, E.P. Identification, timing, and signal specificity of Pseudomonas aeruginosa quorum-controlled genes: A transcriptome analysis. J. Bacteriol. 2003, 185, 2066–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, M.; Greenberg, E.P. Early activation of quorum sensing in Pseudomonas aeruginosa reveals the architecture of a complex regulon. BMC Genom. 2007, 8, 287. [Google Scholar] [CrossRef] [PubMed]
- Rampioni, G.; Schuster, M.; Greenberg, E.P.; Bertani, I.; Grasso, M.; Venturi, V.; Zennaro, E.; Leoni, L. RsaL provides quorum sensing homeostasis and functions as a global regulator of gene expression in Pseudomonas aeruginosa. Mol. Microbiol. 2007, 66, 1557–1565. [Google Scholar] [CrossRef]
- Kasper, S.H.; Bonocora, R.P.; Wade, J.T.; Musah, R.A.; Cady, N.C. Chemical Inhibition of Kynureninase Reduces Pseudomonas aeruginosa Quorum Sensing and Virulence Factor Expression. ACS Chem. Biol. 2016, 11, 1106–1117. [Google Scholar] [CrossRef] [PubMed]
- Poonsuk, K.; Tribuddharat, C.; Chuanchuen, R. Simultaneous overexpression of multidrug efflux pumps in Pseudomonas aeruginosa non-cystic fibrosis clinical isolates. Can. J. Microbiol. 2014, 60, 437–443. [Google Scholar] [CrossRef]
- Rampioni, G.; Pustelny, C.; Fletcher, M.P.; Wright, V.J.; Bruce, M.; Rumbaugh, K.P.; Heeb, S.; Camara, M.; Williams, P. Transcriptomic analysis reveals a global alkyl-quinolone-independent regulatory role for PqsE in facilitating the environmental adaptation of Pseudomonas aeruginosa to plant and animal hosts. Environ. Microbiol. 2010, 12, 1659–1673. [Google Scholar]
- Mima, T.; Joshi, S.; Gomez-Escalada, M.; Schweizer, H.P. Identification and Characterization of TriABC-OpmH, a Triclosan Efflux Pump of Pseudomonas aeruginosa Requiring Two Membrane Fusion Proteins. J. Bacteriol. 2007, 189, 7600–7609. [Google Scholar] [CrossRef] [Green Version]
- Hocquet, D.; Nordmann, P.; El Garch, F.; Cabanne, L.; Plesiat, P. Involvement of the MexXY-OprM efflux system in emergence of cefepime resistance in clinical strains of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2006, 50, 1347–1351. [Google Scholar] [CrossRef] [Green Version]
- Vandeplassche, E.; Sass, A.; Lemarcq, A.; Dandekar, A.A.; Coenye, T.; Crabbé, A. In vitro evolution of Pseudomonas aeruginosa AA2 biofilms in the presence of cystic fibrosis lung microbiome members. Sci. Rep. 2019, 9, 12859. [Google Scholar] [CrossRef] [Green Version]
- Rampioni, G.; Falcone, M.; Heeb, S.; Frangipani, E.; Fletcher, M.P.; Dubern, J.F.; Visca, P.; Leoni, L.; Camara, M.; Williams, P. Unravelling the Genome-Wide Contributions of Specific 2-Alkyl-4-Quinolones and PqsE to Quorum Sensing in Pseudomonas aeruginosa. PLoS Pathog. 2016, 12, e1006029. [Google Scholar] [CrossRef]
- Alcalde-Rico, M.; Martínez, J.L. Methods for Measuring the Production of Quorum Sensing Signal Molecules. In Antibiotic Resistance Protocols; Gillespie, S.H., Ed.; Springer: New York, NY, USA, 2018; pp. 1–15. [Google Scholar]
- Fletcher, M.P.; Diggle, S.P.; Camara, M.; Williams, P. Biosensor-based assays for PQS, HHQ and related 2-alkyl-4-quinolone quorum sensing signal molecules. Nat. Protoc. 2007, 2, 1254–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, D.G.; Merritt, J.H.; Hampton, T.H.; Hodgkinson, J.T.; Janecek, M.; Spring, D.R.; Welch, M.; O’Toole, G.A. 2-Heptyl-4-quinolone, a precursor of the Pseudomonas quinolone signal molecule, modulates swarming motility in Pseudomonas aeruginosa. J. Bacteriol. 2011, 193, 6770–6780. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.M.; Neely, A.; Stintzi, A.; Georges, C.; Holder, I.A. Pyoverdin is essential for virulence of Pseudomonas aeruginosa. Infect. Immun. 1996, 64, 518–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voulhoux, R.; Filloux, A.; Schalk, I.J. Pyoverdine-mediated iron uptake in Pseudomonas aeruginosa: The Tat system is required for PvdN but not for FpvA transport. J. Bacteriol. 2006, 188, 3317–3323. [Google Scholar] [CrossRef] [Green Version]
- Khanam, S.; Guragain, M.; Lenaburg, D.L.; Kubat, R.; Patrauchan, M.A. Calcium induces tobramycin resistance in Pseudomonas aeruginosa by regulating RND efflux pumps. Cell Calcium 2017, 61, 32–43. [Google Scholar] [CrossRef]
- Simon, R.; O’Connell, M.; Labes, M.; Pühler, A. Plasmid vectors for the genetic analysis and manipulation of rhizobia and other gram-negative bacteria. Methods Enzymol. 1986, 118, 640–659. [Google Scholar]
- Winson, M.K.; Swift, S.; Fish, L.; Throup, J.P.; Jorgensen, F.; Chhabra, S.R.; Bycroft, B.W.; Williams, P.; Stewart, G.S. Construction and analysis of luxCDABE-based plasmid sensors for investigating N-acyl homoserine lactone-mediated quorum sensing. FEMS Microbiol. Lett. 1998, 163, 185–192. [Google Scholar] [CrossRef]
- Swift, S.; Karlyshev, A.V.; Fish, L.; Durant, E.L.; Winson, M.K.; Chhabra, S.R.; Williams, P.; Macintyre, S.; Stewart, G.S. Quorum sensing in Aeromonas hydrophila and Aeromonas salmonicida: Identification of the LuxRI homologs AhyRI and AsaRI and their cognate N-acylhomoserine lactone signal molecules. J. Bacteriol. 1997, 179, 5271–5281. [Google Scholar] [CrossRef] [Green Version]
- Becher, A.; Schweizer, H.P. Integration-proficient Pseudomonas aeruginosa vectors for isolation of single-copy chromosomal lacZ and lux gene fusions. BioTechniques 2000, 29, 948–950, 952. [Google Scholar] [CrossRef] [Green Version]
- Hoang, T.T.; Karkhoff-Schweizer, R.R.; Kutchma, A.J.; Schweizer, H.P. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: Application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 1998, 212, 77–86. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.T.; Kutchma, A.J.; Becher, A.; Schweizer, H.P. Integration-proficient plasmids for Pseudomonas aeruginosa: Site-specific integration and use for engineering of reporter and expression strains. Plasmid 2000, 43, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Essar, D.W.; Eberly, L.; Hadero, A.; Crawford, I.P. Identification and characterization of genes for a second anthranilate synthase in Pseudomonas aeruginosa: Interchangeability of the two anthranilate synthases and evolutionary implications. J. Bacteriol. 1990, 172, 884–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, E.; Safrin, M. Elastinolytic and proteolytic enzymes. Methods Mol. Biol. 2014, 1149, 135–169. [Google Scholar] [PubMed]
- Wittgens, A.; Tiso, T.; Arndt, T.T.; Wenk, P.; Hemmerich, J.; Muller, C.; Wichmann, R.; Kupper, B.; Zwick, M.; Wilhelm, S.; et al. Growth independent rhamnolipid production from glucose using the non-pathogenic Pseudomonas putida KT2440. Microb. Cell Factories 2011, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Hoegy, F.; Mislin, G.L.; Schalk, I.J. Pyoverdine and pyochelin measurements. Methods Mol. Biol. 2014, 1149, 293–301. [Google Scholar]
- Linares, J.F.; Moreno, R.; Fajardo, A.; Martinez-Solano, L.; Escalante, R.; Rojo, F.; Martinez, J.L. The global regulator Crc modulates metabolism, susceptibility to antibiotics and virulence in Pseudomonas aeruginosa. Env. Microbiol. 2010, 12, 3196–3212. [Google Scholar] [CrossRef]
| Antibiotic | Strain | Antibiotic | Strain | ||||
|---|---|---|---|---|---|---|---|
| PAO1 | ΔmexL | ΔmexK | PAO1 | ΔmexL | ΔmexK | ||
| Amikacin | 3 | 1.5 | 1.5 | Cefuroxime | >256 | >256 | >256 |
| Gentamicin | 1 | 1 | 1.5 | Cephalothin | >256 | >256 | >256 |
| Kanamycin | 24 | 32 | 32 | Ampicillin | >256 | >256 | >256 |
| Streptomycin | 24 | 16 | 16 | Aztreonam | 1.5 | 1.5 | 1.5 |
| Tobramycin | 1 | 0.75 | 0.75 | Imipenem | 6 | 4 | 4 |
| Tetracycline | 12 | 8 | 6 | Ertapenem | 16 | 16 | 16 |
| Tigecycline | 12 | 8 | 12 | Meropenem | 1 | 1 | 1 |
| Nalidixic acid | 128 | 96 | 96 | Piperacillin | 3 | 2 | 3 |
| Ciprofloxacin | 0.064 | 0.047 | 0.064 | Oxacillin | >256 | >256 | >256 |
| Gatifloxacin | 0.25 | 0.25 | 0.19 | Colistin | 2 | 2 | 2 |
| Levofloxacin | 0.25 | 0.25 | 0.25 | Polymyxin B | 1 | 1 | 1.5 |
| Moxifloxacin | 1.5 | 1.5 | 2 | Chloramphenicol | 64 | 64 | 64 |
| Norfloxacin | 0.38 | 0.38 | 0.38 | Erythromycin | >256 | >256 | >256 |
| Ofloxacin | 0.75 | 0.75 | 0.75 | Fosfomycin | 256 | 256 | 256 |
| Cefepime | 0.75 | 0.75 | 0.75 | Rifampicin | 12 | 12 | 12 |
| Cefoxitin | >256 | >256 | >256 | Trimethoprim-Sulfamethoxazole | 1.5 | 2 | 1.5 |
| Cefotaxime | 12 | 8 | 8 | Ceftazidime–Avibactam | 0.75 | 0.75 | 0.75 |
| Ceftazidime | 0.5 | 0.5 | 0.5 | ||||
| Name | Sequence | Description |
|---|---|---|
| CTX-Fwd | 5′-GTCATGCTCTTCTCTAATGCGTG-3′ | Check the insertion of mini-CTX-lux in the chromosome of P. aeruginosa strains |
| CTX-Rev | 5′-GCGTAATACGACTCACTATAGGGC-3′ | |
| rplU Fwd | 5′-CGCAGTGATTGTTACCGGTG-3′ | Check DNA contamination of RNA samples |
| rplU Rev | 5′-AGGCCTGAATGCCGGTGATC-3′ | |
| EcoRI_mexL_Fwd | 5′- CCCGAATTCCTGGGAATGGCTGACCAGGT-3′ | Amplification of flanking area “Up” around mexL gene |
| mexL_int_Rev | 5′-CGTCGCGCCTGAGCTGCCGCGCTTATACAATTGAAA-3′ | |
| mexL_int_Fwd | 5′-TTTCAATTGTATAAGCGCGGCAGCTCAGGCGCGACG-3′ | Amplification of flanking area “Down” around mexL gene |
| BamHI_mexL_Rev | 5′-CCCGGATCCTGCTCGCGCGGCTACGC-3′ | |
| mexL_Fwd_Check | 5′-CGAGGAACAGGGAGGAAAAC-3′ | Check mexL deletion |
| mexL_Rev_Check | 5′-GGCGCCTACTTCCCCTTC-3′ | |
| EcoRI_mexK_Fw | 5′-CCCGAATTCCGAGGTGCTGATCGGCCTGC-3′ | Amplification of flanking area “Up” around mexK gene |
| mexK_int_Rev | 5′-GTCCCTTCTCCCGTCAGGGCGACTACTCCTTGGCCG-3′ | |
| mexK_int_Fwd | 5′-CGGCCAAGGAGTAGTCGCCCTGACGGGAGAAGGGAC-3′ | Amplification of flanking area “Down” around mexK gene |
| BamHI_mexK_Rev | 5′-CCCGGATCCAGCTGATGAAGCAGTTCGGC-3′ | |
| mexK_Fwd_Check | 5′-GGTGCTCGAAGGCCTGAA-3′ | Check mexK deletion |
| mexK_Rev_Check | 5′-AACGTCGAGGGCTATGTCAC-3′ | |
| mexK Fwd | 5′-GGTGCTCGAAGGCCTGAA-3′ | RT-qPCR |
| mexK Rev | 5′-AACGTCGAGGGCTATGTCAC-3′ | |
| hcnB Fwd | 5′-GAACGCCGAGAATCCCATCT-3′ | RT-qPCR |
| hcnB Rev | 5′-CATCGCCGGGCTGAAGAT-3′ | |
| lasA Fwd | 5′-ATGGACCAGATCCAGGTGAG-3′ | RT-qPCR |
| lasA Rev | 5′-CGTTGTCGTAGTTGCTGGTG-3′ | |
| lasB Fwd | 5′-ATCGGCAAGTACACCTACGG-3′ | RT-qPCR |
| lasB Rev | 5′-ACCAGTCCCGGTACAGTTTG-3′ | |
| lasI Fwd | 5′-CTACAGCCTGCAGAACGACA-3′ | RT-qPCR |
| lasI Rev | 5′-ATCTGGGTCTTGGCATTGAG-3′ | |
| mexB Fwd | 5′-TGAACAGCGTGTTCGAACTGG-3′ | RT-qPCR |
| mexB Rev | 5′-CACTTCGACATTACGAATCCC-3′ | |
| mexC Fwd | 5′-GAAGCGCTTCGAGGAGGG-3′ | RT-qPCR |
| mexC Rev | 5′-CAGCCAGCAGGACTTCGATA-3′ | |
| mexE Fwd | 5′-GGACTTCCTCGACAACCAGG-3′ | RT-qPCR |
| mexE Rev | 5′-AGAAGTTCGTGCTGGTCCTG-3′ | |
| mexG Fwd | 5′-GGCGAAGCTGTTCGACTATC-3′ | RT-qPCR |
| mexG Rev | 5′-AGAAGGTGTGGACGATGAGG-3′ | |
| mexX Fwd | 5′-GTCGCCCTATTCCTGCTGG-3′ | RT-qPCR |
| mexX Rev | 5′-GTCACCCGTCGCCTGTAC-3′ | |
| opmH Fwd | 5′-GTGCGCGACTACAACAACAG-3′ | RT-qPCR |
| opmH Rev | 5′-GCCGAGCAGTTACAGAGCAA-3′ | |
| oprM Fwd | 5′-TCAACCTGCCGATCTTCACC-3′ | RT-qPCR |
| oprM Rev | 5′-GCGACGAGTACTACCAGCTC-3′ | |
| phnB Fwd | 5′-CACTCGCTGGTGGTCAGTC-3′ | RT-qPCR |
| phnB Rev | 5′-AGAGTAGAGCGTTCTCCAGCA-3′ | |
| phzB1 Fwd | 5′-AACGAACTTCGCGAAAAGAA-3′ | RT-qPCR |
| phzB1 Rev | 5′-TTTGTCTTTGCCACGAATGA-3′ | |
| phzB2 Fwd | 5′-GCGAGACGGTGGTCAAGTAT-3′ | RT-qPCR |
| phzB2 Rev | 5′-AATCCGGGAAGCATTTCAG-3′ | |
| phzS Fwd | 5′-CAAGTCGCTGGTGAACTGG-3′ | RT-qPCR |
| phzS Rev | 5′-CGGGTACTGCAGGATCAACT-3′ | |
| pqsA Fwd | 5′-CAATACACCTCGGGTTCCAC-3′ | RT-qPCR |
| pqsA Rev | 5′-TGAACCAGGGAAAGAACAGG-3′ | |
| pqsE Fwd | 5′-TGGTGTTCGACGACATGGAG-3′ | RT-qPCR |
| pqsE Rev | 5′-AATCCCTCGACGAACTGAGC-3′ | |
| pqsH Fwd | 5′-ATGTCTACGCGACCCTGAAG-3′ | RT-qPCR |
| pqsH Rev | 5′-AACTCCTCGAGGTCGTTGTG-3′ | |
| rhlA Fwd | 5′-CGAGGTCAATCACCTGGTCT-3′ | RT-qPCR |
| rhlA Rev | 5′-GACGGTCTCGTTGAGCAGAT-3′ | |
| rhlI Fwd | 5′-CTCTCTGAATCGCTGGAAGG-3′ | RT-qPCR |
| rhlI Rev | 5′-GACGTCCTTGAGCAGGTAGG-3′ | |
| rpsL Fwd | 5′-GCAAGCGCATGGTCGACAAGA-3′ | RT-qPCR (housekeeping) |
| rpsL Rev | 5′-CGCTGTGCTCTTGCAGGTTGTGA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amieva, R.; Gil-Gil, T.; Martínez, J.L.; Alcalde-Rico, M. The MexJK Multidrug Efflux Pump Is Not Involved in Acquired or Intrinsic Antibiotic Resistance in Pseudomonas aeruginosa, but Modulates the Bacterial Quorum Sensing Response. Int. J. Mol. Sci. 2022, 23, 7492. https://doi.org/10.3390/ijms23147492
Amieva R, Gil-Gil T, Martínez JL, Alcalde-Rico M. The MexJK Multidrug Efflux Pump Is Not Involved in Acquired or Intrinsic Antibiotic Resistance in Pseudomonas aeruginosa, but Modulates the Bacterial Quorum Sensing Response. International Journal of Molecular Sciences. 2022; 23(14):7492. https://doi.org/10.3390/ijms23147492
Chicago/Turabian StyleAmieva, Rafael, Teresa Gil-Gil, José Luis Martínez, and Manuel Alcalde-Rico. 2022. "The MexJK Multidrug Efflux Pump Is Not Involved in Acquired or Intrinsic Antibiotic Resistance in Pseudomonas aeruginosa, but Modulates the Bacterial Quorum Sensing Response" International Journal of Molecular Sciences 23, no. 14: 7492. https://doi.org/10.3390/ijms23147492
APA StyleAmieva, R., Gil-Gil, T., Martínez, J. L., & Alcalde-Rico, M. (2022). The MexJK Multidrug Efflux Pump Is Not Involved in Acquired or Intrinsic Antibiotic Resistance in Pseudomonas aeruginosa, but Modulates the Bacterial Quorum Sensing Response. International Journal of Molecular Sciences, 23(14), 7492. https://doi.org/10.3390/ijms23147492