
The Role of Human Milk Lipids and Lipid Metabolites in Protecting the Infant against Non-Communicable Disease

 , , , and
, , , and 
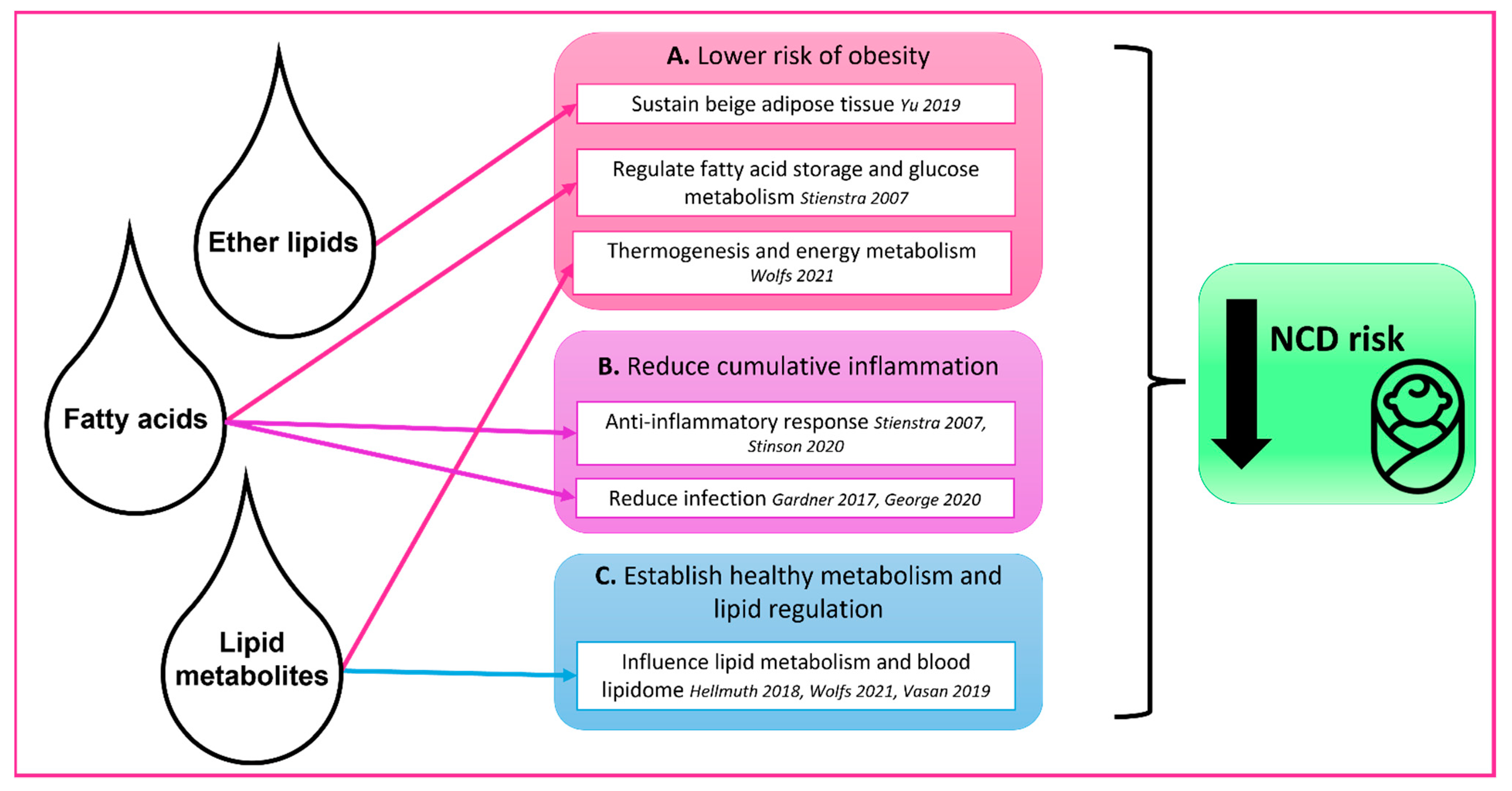
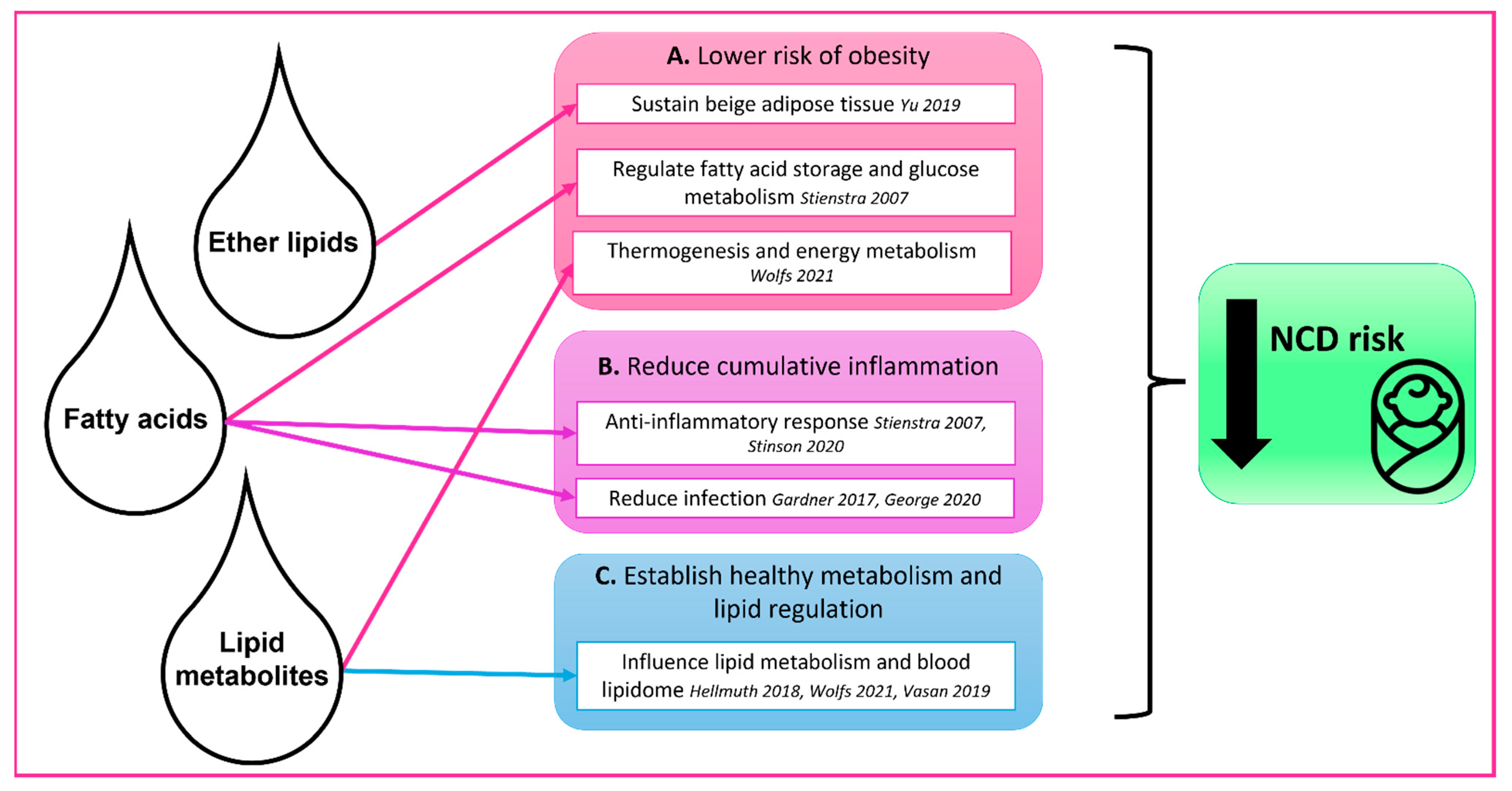{kind=link}
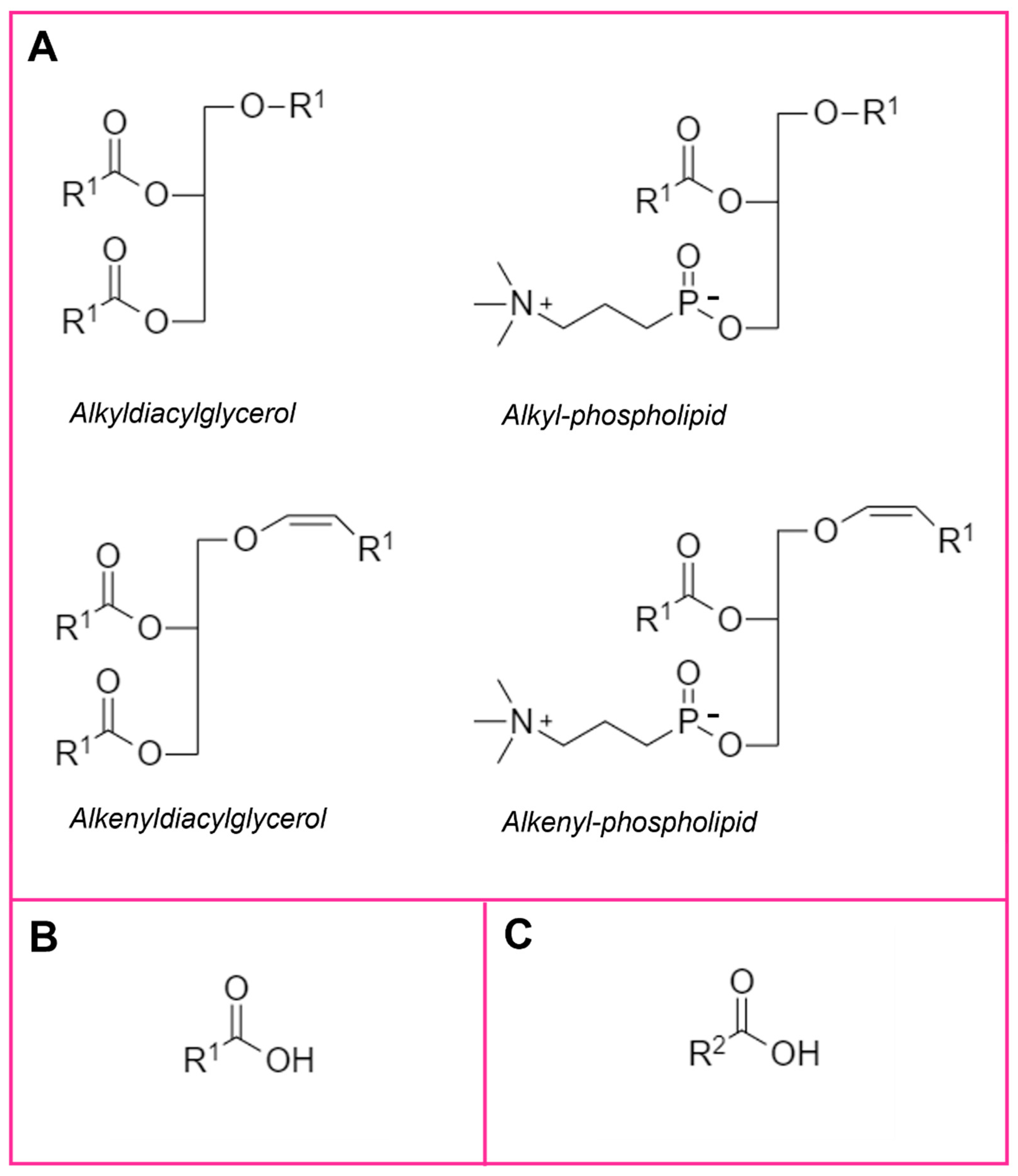
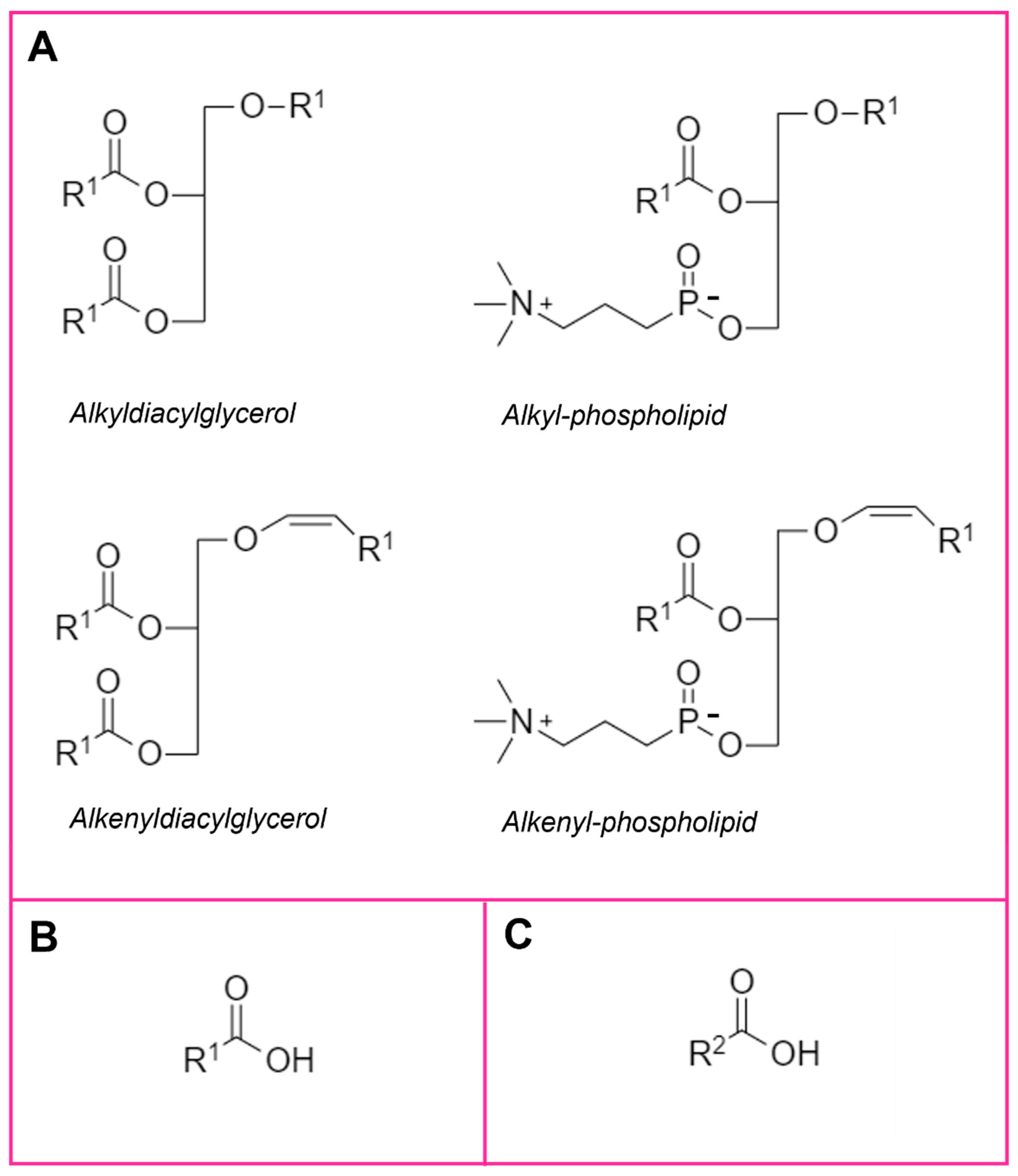{kind=link}
Abstract
:1. Introduction
2. Breastfeeding and Non-Communicable Disease Risk
2.1. Early Life Growth
2.2. Early Life Inflammation
2.3. Early Life Metabolism
3. The Emerging Field of Human Milk Lipidomics and Non-Communicable Disease Risk
4. Human Milk Ether Lipids
5. Human Milk Fatty Acids
6. Human Milk Lipid Metabolites
7. Future Directions
- ❖
- should be considered bioactive and included in early life studies
- ❖
- need to be described comprehensively, in diverse populations
- ❖
- need to be considered in combination with other biofluids, such as infant blood samples
- ❖
- metabotypes or scores need to be developed and interrogated
- ❖
- must be included within multidisciplinary research
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.; Wang, J. Modelling and prediction of global non-communicable diseases. BMC Public Health 2020, 20, 822. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Mendez, E.; Quintero-Fabian, S.; Fernandez-Mejia, C.; Lazo-de-la-Vega-Monroy, M.-L. Early-life programming of adipose tissue. Nutr. Res. Rev. 2020, 33, 244–259. [Google Scholar] [CrossRef] [PubMed]
- Geserick, M.; Vogel, M.; Gausche, R.; Lipek, T.; Spielau, U.; Keller, E.; Pfäffle, R.; Kiess, W.; Körner, A. Acceleration of BMI in Early Childhood and Risk of Sustained Obesity. N. Engl. J. Med. 2018, 379, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Victora, C.G.; Bahl, R.; Barros, A.J.; França, G.V.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C. Breastfeeding in the 21st century: Epidemiology, mechanisms, and lifelong effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Liu, L.; Zhu, Y.; Huang, G.; Wang, P.P. The association between breastfeeding and childhood obesity: A meta-analysis. BMC Public Health 2014, 14, 1267. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.B.; Vehling, L.; Chan, D.; Klopp, A.; Nickel, N.C.; McGavock, J.M.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Moraes, T.J.; et al. Infant Feeding and Weight Gain: Separating Breast Milk from Breastfeeding and Formula from Food. J. Pediatr. 2018, 142, e20181092. [Google Scholar] [CrossRef] [Green Version]
- Stienstra, R.; Duval, C.; Müller, M.; Kersten, S. PPARs, Obesity, and Inflammation. PPAR Res. 2007, 2007, 95974. [Google Scholar] [CrossRef] [Green Version]
- Frank, N.M.; Lynch, K.F.; Uusitalo, U.; Yang, J.; Lönnrot, M.; Virtanen, S.M.; Hyöty, H.; Norris, J.M.; Rewers, M.; Bautista, K.; et al. The relationship between breastfeeding and reported respiratory and gastrointestinal infection rates in young children. BMC Pediatr. 2019, 19, 339. [Google Scholar] [CrossRef]
- Prentice, P.; Koulman, A.; Matthews, L.; Acerini, C.L.; Ong, K.K.; Dunger, D.B. Lipidomic Analyses, Breast- and Formula-Feeding, and Growth in Infants. J. Pediatr. 2015, 166, 276–281.e6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, L.; Wang, L.; Pan, Y.; Hao, X.; Zhang, G.; Li, X.; Hussain, M. Comparative Lipidomics Analysis of Human Milk and Infant Formulas Using UHPLC-Q-TOF-MS. J. Agric. Food Chem. 2021, 69, 1146–1155. [Google Scholar] [CrossRef]
- Hewelt-Belka, W.; Garwolińska, D.; Młynarczyk, M.; Kot-Wasik, A. Comparative Lipidomic Study of Human Milk from Different Lactation Stages and Milk Formulas. Nutrients 2020, 12, 2165. [Google Scholar] [CrossRef] [PubMed]
- Burugupalli, S.; Smith, A.A.T.; Olshansky, G.; Huynh, K.; Giles, C.; Paul, S.; Nguyen, A.; Duong, T.; Mellett, N.; Cinel, M.; et al. Ontogeny of plasma lipid metabolism in pregnancy and early childhood: A longitudinal population study. eLife 2021, 11, e72779. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Dilbaz, S.; Coßmann, J.; Hoang, A.C.; Diedrich, V.; Herwig, A.; Harauma, A.; Hoshi, Y.; Moriguchi, T.; Landgraf, K.; et al. Breast milk alkylglycerols sustain beige adipocytes through adipose tissue macrophages. J. Clin. Investig. 2019, 129, 2485–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaacs, C.E.; Kashyap, S.; Heird, W.C.; Thormar, H. Antiviral and antibacterial lipids in human milk and infant formula feeds. Arch. Dis. Child. 1990, 65, 861–864. [Google Scholar] [CrossRef] [Green Version]
- Hellmuth, C.; Uhl, O.; Demmelmair, H.; Grunewald, M.; Auricchio, R.; Castillejo, G.; Korponay-Szabo, I.R.; Polanco, I.; Roca, M.; Vriezinga, S.L.; et al. The impact of human breast milk components on the infant metabolism. PLoS ONE 2018, 13, e0197713. [Google Scholar] [CrossRef] [PubMed]
- Wolfs, D.; Lynes, M.D.; Tseng, Y.H.; Pierce, S.; Bussberg, V.; Darkwah, A.; Tolstikov, V.; Narain, N.R.; Rudolph, M.C.; Kiebish, M.A.; et al. Brown Fat-Activating Lipokine 12,13-diHOME in Human Milk Is Associated with Infant Adiposity. J. Clin. Endocrinol. Metab. 2021, 106, e943–e956. [Google Scholar] [CrossRef]
- Stinson, L.F.; Gay, M.C.L.; Koleva, P.T.; Eggesbø, M.; Johnson, C.C.; Wegienka, G.; du Toit, E.; Shimojo, N.; Munblit, D.; Campbell, D.E.; et al. Human Milk from Atopic Mothers Has Lower Levels of Short Chain Fatty Acids. Front. Immunol. 2020, 11, 1427. [Google Scholar] [CrossRef]
- Gardner, A.S.; Rahman, I.A.; Lai, C.T.; Hepworth, A.; Trengove, N.; Hartmann, P.E.; Geddes, D.T. Changes in Fatty Acid Composition of Human Milk in Response to Cold-Like Symptoms in the Lactating Mother and Infant. Nutrients 2017, 9, 1034. [Google Scholar] [CrossRef]
- George, A.D.; Gay, M.C.L.; Wlodek, M.E.; Trengove, R.D.; Murray, K.; Geddes, D.T. Untargeted lipidomics using liquid chromatography-ion mobility-mass spectrometry reveals novel triacylglycerides in human milk. Sci. Rep. 2020, 10, 9255. [Google Scholar] [CrossRef]
- Vasan, S.K.; Noordam, R.; Gowri, M.S.; Neville, M.J.; Karpe, F.; Christodoulides, C. The proposed systemic thermogenic metabolites succinate and 12,13-diHOME are inversely associated with adiposity and related metabolic traits: Evidence from a large human cross-sectional study. Diabetologia 2019, 62, 2079–2087. [Google Scholar] [CrossRef] [Green Version]
- Rito, A.I.; Buoncristiano, M.; Spinelli, A.; Salanave, B.; Kunešová, M.; Hejgaard, T.; García Solano, M.; Fijałkowska, A.; Sturua, L.; Hyska, J.; et al. Association between Characteristics at Birth, Breastfeeding and Obesity in 22 Countries: The WHO European Childhood Obesity Surveillance Initiative—COSI 2015/2017. Obes. Facts 2019, 12, 226–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinckard, K.M.; Shettigar, V.K.; Wright, K.R.; Abay, E.; Baer, L.A.; Vidal, P.; Dewal, R.S.; Das, D.; Duarte-Sanmiguel, S.; Hernández-Saavedra, D.; et al. A Novel Endocrine Role for the BAT-Released Lipokine 12,13-diHOME to Mediate Cardiac Function. Circulation 2021, 143, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Horta, B.L.; Loret de Mola, C.; Victora, C.G. Long-term consequences of breastfeeding on cholesterol, obesity, systolic blood pressure and type 2 diabetes: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Rollins, N.C.; Bhandari, N.; Hajeebhoy, N.; Horton, S.; Lutter, C.K.; Martines, J.C.; Piwoz, E.G.; Richter, L.M.; Victora, C.G. Why invest, and what it will take to improve breastfeeding practices? Lancet 2016, 387, 491–504. [Google Scholar] [CrossRef]
- Mihrshahi, S.; Baur, L.A. What exposures in early life are risk factors for childhood obesity? J. Paediatr. Child Health 2018, 54, 1294–1298. [Google Scholar] [CrossRef]
- Dewey, K.G.; Peerson, J.M.; Brown, K.H.; Krebs, N.F.; Michaelsen, K.F.; Persson, L.A.; Salmenpera, L.; Whitehead, R.G.; Yeung, D.L. Growth of breast-fed infants deviates from current reference data: A pooled analysis of US, Canadian, and European data sets. World Health Organization Working Group on Infant Growth. Pediatrics 1995, 96 Pt 1, 495–503. [Google Scholar] [CrossRef]
- Carberry, A.E.; Colditz, P.B.; Lingwood, B.E. Body Composition from Birth to 4.5 Months in Infants Born to Non-Obese Women. Pediatric Res. 2010, 68, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Chomtho, S.; Wells, J.C.; Williams, J.E.; Davies, P.S.; Lucas, A.; Fewtrell, M.S. Infant growth and later body composition: Evidence from the 4-component model. Am. J. Clin. Nutr. 2008, 87, 1776–1784. [Google Scholar] [CrossRef]
- Prescott, S.L. Early-life environmental determinants of allergic diseases and the wider pandemic of inflammatory noncommunicable diseases. J. Allergy Clin. Immunol. 2013, 131, 23–30. [Google Scholar] [CrossRef]
- Olson, J.S.; Hayward, M.D. Breastfeeding, overweight status, and inflammation. Soc. Sci. Res. 2017, 64, 226–236. [Google Scholar] [CrossRef] [Green Version]
- McDade, T.W.; Metzger, M.W.; Chyu, L.; Duncan, G.J.; Garfield, C.; Adam, E.K. Long-term effects of birth weight and breastfeeding duration on inflammation in early adulthood. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridker, P.M. A Test in Context: High-Sensitivity C-Reactive Protein. J. Am. Coll. Cardiol. 2016, 67, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.J.; Williams, S.M.; Poulton, R. Breast feeding is related to C reactive protein concentration in adult women. J. Epidemiol. Community Health 2006, 60, 146–148. [Google Scholar] [CrossRef] [Green Version]
- Meikle, P.J.; Summers, S.A. Sphingolipids and phospholipids in insulin resistance and related metabolic disorders. Nat. Rev. Endocrinol. 2017, 13, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Beyene, H.B.; Olshansky, G.; Giles, C.; Huynh, K.; Cinel, M.; Mellett, N.A.; Smith, A.A.T.; Shaw, J.E.; Magliano, D.J.; Meikle, P.J. Lipidomic Signatures of Changes in Adiposity: A Large Prospective Study of 5849 Adults from the Australian Diabetes, Obesity and Lifestyle Study. Metabolites 2021, 11, 646. [Google Scholar] [CrossRef] [PubMed]
- Harit, D.; Faridi, M.M.; Aggarwal, A.; Sharma, S.B. Lipid profile of term infants on exclusive breastfeeding and mixed feeding: A comparative study. Eur. J. Clin. Nutr. 2008, 62, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallio, M.J.T.; Salmenperä, L.; Siimes, M.A.; Perheentupa, J.; Miettinen, T.A. Exclusive Breast-Feeding and Weaning: Effect on Serum Cholesterol and Lipoprotein Concentrations in Infants During the First Year of Life. Pediatrics 1992, 89, 663–666. [Google Scholar] [CrossRef]
- Hui, L.L.; Kwok, M.K.; Nelson, E.A.S.; Lee, S.L.; Leung, G.M.; Schooling, C.M. Breastfeeding in Infancy and Lipid Profile in Adolescence. Pediatrics 2019, 143, e20183075. [Google Scholar] [CrossRef]
- Koletzko, B. Human Milk Lipids. Ann. Nutr. Metab. 2016, 69 (Suppl. 2), 28–40. [Google Scholar] [CrossRef] [Green Version]
- Lopez, C.; Ménard, O. Human milk fat globules: Polar lipid composition and in situ structural investigations revealing the heterogeneous distribution of proteins and the lateral segregation of sphingomyelin in the biological membrane. Colloids Surf. B Biointerfaces 2011, 83, 29–41. [Google Scholar] [CrossRef]
- Alexandre-Gouabau, M.C.; Moyon, T.; Cariou, V.; Antignac, J.P.; Qannari, E.M.; Croyal, M.; Soumah, M.; Guitton, Y.; David-Sochard, A.; Billard, H.; et al. Breast Milk Lipidome Is Associated with Early Growth Trajectory in Preterm Infants. Nutrients 2018, 10, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, A.D.; Gay, M.C.L.; Selvalatchmanan, J.; Torta, F.; Bendt, A.K.; Wenk, M.R.; Murray, K.; Wlodek, M.E.; Geddes, D.T. Healthy Breastfeeding Infants Consume Different Quantities of Milk Fat Globule Membrane Lipids. Nutrients 2021, 13, 2951. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.M.; Brei, C.; Stecher, L.; Much, D.; Brunner, S.; Hauner, H. Associations between long-chain PUFAs in maternal blood, cord blood, and breast milk and offspring body composition up to 5 years: Follow-up from the INFAT study. Eur. J. Clin. Nutr. 2019, 73, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, M.C.; Young, B.E.; Lemas, D.J.; Palmer, C.E.; Hernandez, T.L.; Barbour, L.A.; Friedman, J.E.; Krebs, N.F.; MacLean, P.S. Early infant adipose deposition is positively associated with the n-6 to n-3 fatty acid ratio in human milk independent of maternal BMI. Int. J. Obes. 2017, 41, 510–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäkelä, J.; Linderborg, K.; Niinikoski, H.; Yang, B.; Lagström, H. Breast milk fatty acid composition differs between overweight and normal weight women: The STEPS Study. Eur. J. Nutr. 2013, 52, 727–735. [Google Scholar] [CrossRef]
- Paul, S.; Lancaster, G.I.; Meikle, P.J. Plasmalogens: A potential therapeutic target for neurodegenerative and cardiometabolic disease. Prog. Lipid Res. 2019, 74, 186–195. [Google Scholar] [CrossRef]
- Weir, J.M.; Wong, G.; Barlow, C.K.; Greeve, M.A.; Kowalczyk, A.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Jowett, J.B.M.; Shaw, J.; et al. Plasma lipid profiling in a large population-based cohort. J. Lipid Res. 2013, 54, 2898–2908. [Google Scholar] [CrossRef] [Green Version]
- Meikle, P.J.; Wong, G.; Tsorotes, D.; Barlow, C.K.; Weir, J.M.; Christopher, M.J.; MacIntosh, G.L.; Goudey, B.; Stern, L.; Kowalczyk, A.; et al. Plasma Lipidomic Analysis of Stable and Unstable Coronary Artery Disease. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2723–2732. [Google Scholar] [CrossRef] [Green Version]
- Meikle, P.J.; Wong, G.; Barlow, C.K.; Weir, J.M.; Greeve, M.A.; MacIntosh, G.L.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Kowalczyk, A.; et al. Plasma Lipid Profiling Shows Similar Associations with Prediabetes and Type 2 Diabetes. PLoS ONE 2013, 8, e74341. [Google Scholar] [CrossRef] [Green Version]
- Hallgren, B.; Niklasson, A.; Ställberg, G.; Thorin, H. On the occurrence of 1-O-alkylglycerols and 1-O-(2-methoxyalkyl)glycerols in human colostrum, human milk, cow’s milk, sheep’s milk, human red bone marrow, red cells, blood plasma and a uterine carcinoma. Acta Chem. Scand. Ser. B Org. Chem. Biochem. 1974, 28, 1029–1034. [Google Scholar] [CrossRef]
- Garcia, C.; Lutz, N.W.; Confort-Gouny, S.; Cozzone, P.J.; Armand, M.; Bernard, M. Phospholipid fingerprints of milk from different mammalians determined by 31P NMR: Towards specific interest in human health. Food Chem. 2012, 135, 1777–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Smith, A.A.T.; Culham, K.; Gunawan, K.A.; Weir, J.M.; Cinel, M.A.; Jayawardana, K.S.; Mellett, N.A.; Lee, M.K.; Murphy, A.J.; et al. Shark liver oil supplementation enriches endogenous plasmalogens and reduces markers of dyslipidemia and inflammation. J. Lipid Res. 2021, 62, 100092. [Google Scholar] [CrossRef] [PubMed]
- Chorostowska-Wynimko, J.; Krotkiewski, M.; Radomska-Leśniewska, D.; Sokolnicka, I.; Skopińska-Rózewska, E. The synergistic effect of lactic acid bacteria and alkylglycerols on humoral immunity in mice. Int. J. Tissue React. 2001, 23, 81–87. [Google Scholar]
- Simopoulos, A.P. Importance of the Omega-6/Omega-3 Balance in Health and Disease: Evolutionary Aspects of Diet. World Rev. Nutr. Diet. 2011, 102, 21. [Google Scholar]
- Logan, C.A.; Brandt, S.; Wabitsch, M.; Brenner, H.; Wiens, F.; Stahl, B.; Marosvölgyi, T.; Decsi, T.; Rothenbacher, D.; Genuneit, J. New approach shows no association between maternal milk fatty acid composition and childhood wheeze or asthma. Allergy 2017, 72, 1374–1383. [Google Scholar] [CrossRef]
- George, A.D.; Gay, M.C.L.; Wlodek, M.E.; Murray, K.; Geddes, D.T. The Fatty Acid Species and Quantity Consumed by the Breastfed Infant Are Important for Growth and Development. Nutrients 2021, 13, 4183. [Google Scholar] [CrossRef]
- Mook-Kanamori, D.O.; Steegers, E.A.P.; Uitterlinden, A.G.; Moll, H.A.; van Duijn, C.M.; Hofman, A.; Jaddoe, V.W.V. Breast-Feeding Modifies the Association of PPARγ2 Polymorphism Pro12Ala With Growth in Early Life: The Generation R Study. Diabetes 2009, 58, 992–998. [Google Scholar] [CrossRef] [Green Version]
- Rees, W.D.; McNeil, C.J.; Maloney, C.A. The Roles of PPARs in the Fetal Origins of Metabolic Health and Disease. PPAR Res. 2008, 2008, 459030. [Google Scholar] [CrossRef]
- Dunstan, J.A.; Roper, J.; Mitoulas, L.; Hartmann, P.E.; Simmer, K.; Prescott, S.L. The effect of supplementation with fish oil during pregnancy on breast milk immunoglobulin A, soluble CD14, cytokine levels and fatty acid composition. Clin. Exp. Allergy 2004, 34, 1237–1242. [Google Scholar] [CrossRef]
- Lynes, M.D.; Leiria, L.O.; Lundh, M.; Bartelt, A.; Shamsi, F.; Huang, T.L.; Takahashi, H.; Hirshman, M.F.; Schlein, C.; Lee, A.; et al. The cold-induced lipokine 12,13-diHOME promotes fatty acid transport into brown adipose tissue. Nat. Med. 2017, 23, 631–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levan, S.R.; Stamnes, K.A.; Lin, D.L.; Panzer, A.R.; Fukui, E.; McCauley, K.; Fujimura, K.E.; McKean, M.; Ownby, D.R.; Zoratti, E.M.; et al. Elevated faecal 12,13-diHOME concentration in neonates at high risk for asthma is produced by gut bacteria and impedes immune tolerance. Nat. Microbiol. 2019, 4, 1851–1861. [Google Scholar] [CrossRef] [PubMed]
- Lundström, S.L.; Yang, J.; Källberg, H.J.; Thunberg, S.; Gafvelin, G.; Haeggström, J.Z.; Grönneberg, R.; Grunewald, J.; van Hage, M.; Hammock, B.D.; et al. Allergic Asthmatics Show Divergent Lipid Mediator Profiles from Healthy Controls Both at Baseline and following Birch Pollen Provocation. PLoS ONE 2012, 7, e33780. [Google Scholar] [CrossRef] [PubMed]
- Rzehak, P.; Hellmuth, C.; Uhl, O.; Kirchberg, F.F.; Peissner, W.; Harder, U.; Grote, V.; Weber, M.; Xhonneux, A.; Langhendries, J.P.; et al. Rapid Growth and Childhood Obesity Are Strongly Associated with LysoPC(14:0). Ann. Nutr. Metab. 2014, 64, 294–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewey, K.G.; Güngör, D.; Donovan, S.M.; Madan, E.M.; Venkatramanan, S.; Davis, T.A.; Kleinman, R.E.; Taveras, E.M.; Bailey, R.L.; Novotny, R.; et al. Breastfeeding and risk of overweight in childhood and beyond: A systematic review with emphasis on sibling-pair and intervention studies. Am. J. Clin. Nutr. 2021, 114, 1774–1790. [Google Scholar] [CrossRef] [PubMed]
- Selvalatchmanan, J.; Rukmini, A.V.; Ji, S.; Triebl, A.; Gao, L.; Bendt, A.K.; Wenk, M.R.; Gooley, J.J.; Torta, F. Variability of Lipids in Human Milk. Metabolites 2021, 11, 104. [Google Scholar] [CrossRef]
- George, A.D.; Gay, M.C.L.; Murray, K.; Muhlhausler, B.S.; Wlodek, M.E.; Geddes, D.T. Human Milk Sampling Protocols Affect Estimation of Infant Lipid Intake. J. Nutr. 2020, 150, 2924–2930. [Google Scholar] [CrossRef]
- Lamb, R.L.; Haszard, J.J.; Little, H.M.J.; Franks, A.F.; Meeks, M.G. Macronutrient Composition of Donated Human Milk in a New Zealand Population. J. Hum. Lact. 2021, 37, 114–121. [Google Scholar] [CrossRef]
- George, A.D.; Gay, M.C.L.; Wlodek, M.E.; Geddes, D.T. The importance of infants’ lipid intake in human milk research. Nutr. Rev. 2021, 79, 1353–1361. [Google Scholar] [CrossRef]
- Furse, S.; Billing, G.; Snowden, S.G.; Smith, J.; Goldberg, G.; Koulman, A. Relationship between the lipid composition of maternal plasma and infant plasma through breast milk. Metab. Off. J. Metab. Soc. 2019, 15, 129. [Google Scholar] [CrossRef] [Green Version]
- Alexandre-Gouabau, M.C.; Moyon, T.; David-Sochard, A.; Fenaille, F.; Cholet, S.; Royer, A.L.; Guitton, Y.; Billard, H.; Darmaun, D.; Rozé, J.C.; et al. Comprehensive Preterm Breast Milk Metabotype Associated with Optimal Infant Early Growth Pattern. Nutrients 2019, 11, 528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Pan, J.; Li, Y.; Ju, M.; Zhang, W.; Lu, J.; Lv, J.; Li, K. Comprehensive Human Milk Patterns Are Related to Infant Growth and Allergy in the CHMP Study. Mol. Nutr. Food Res. 2021, 65, 2100011. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, A.D.; Burugupalli, S.; Paul, S.; Mansell, T.; Burgner, D.; Meikle, P.J. The Role of Human Milk Lipids and Lipid Metabolites in Protecting the Infant against Non-Communicable Disease. Int. J. Mol. Sci. 2022, 23, 7490. https://doi.org/10.3390/ijms23147490
George AD, Burugupalli S, Paul S, Mansell T, Burgner D, Meikle PJ. The Role of Human Milk Lipids and Lipid Metabolites in Protecting the Infant against Non-Communicable Disease. International Journal of Molecular Sciences. 2022; 23(14):7490. https://doi.org/10.3390/ijms23147490
Chicago/Turabian StyleGeorge, Alexandra D., Satvika Burugupalli, Sudip Paul, Toby Mansell, David Burgner, and Peter J. Meikle. 2022. "The Role of Human Milk Lipids and Lipid Metabolites in Protecting the Infant against Non-Communicable Disease" International Journal of Molecular Sciences 23, no. 14: 7490. https://doi.org/10.3390/ijms23147490
APA StyleGeorge, A. D., Burugupalli, S., Paul, S., Mansell, T., Burgner, D., & Meikle, P. J. (2022). The Role of Human Milk Lipids and Lipid Metabolites in Protecting the Infant against Non-Communicable Disease. International Journal of Molecular Sciences, 23(14), 7490. https://doi.org/10.3390/ijms23147490