
LIN28 Family in Testis: Control of Cell Renewal, Maturation, Fertility and Aging

 ,
,  , , and
, , and
Abstract
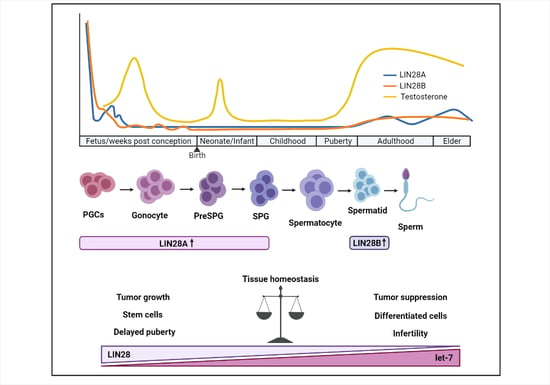
1. Introduction
2. Expression and Function of LIN28AB in Tissues
3. Structure and Subcellular Distribution of LIN28AB
4. Expression Dynamics of LIN28 Family in the Testis
4.1. LIN28AB Expression Dynamics across Fetal Testis Development
4.2. LIN28AB Expression Dynamics across Postnatal Testis Development
4.3. LIN28 in the Testicular Aging
5. Molecular Pathways of LIN28 Family
5.1. Network of LIN28AB Targets
5.2. Upstream Regulators of LIN28AB
6. LIN28AB in Testicular Disorders
7. The LIN28AB as a Molecular Target in Treating Injuries and Diseases
8. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CysCysHisCys | CCHC |
| CSD | cold-shock domain |
| CSP | cold shock proteins |
| CTSC | cardiac tissue-derived stem-like cell |
| DOX | doxorubicin |
| dpc | days post coitum |
| dpp | days postpartum |
| EDC | endocrine-disrupting chemicals |
| ESC | embryonic stem cell |
| GCNIS | germ cell neoplasia in situ |
| GCT | germ cell tumor |
| GOF | gain-of-function |
| hESC | human embryonic stem cell |
| HFD | high-fat diet |
| HSPC | hematopoietic stem and progenitor cell |
| IHC | immunohistochemical |
| IMP | importin |
| KS | Klinefelter’s syndrome |
| LOF | loss-of-function |
| MPW | masculinization programming window |
| NLS | nuclear localization signal |
| NoLS | nucleolar localization signal |
| OSCC | oral squamous cell carcinoma |
| PGC | primordial germ cell |
| preSPG | pre-spermatogonia |
| PSC | pluripotent stem cell |
| RPC | retinal progenitor cell |
| scRNA-seq | single-cell RNA sequencing |
| SPG | spermatogonia |
| SSC | spermatogonial stem cell |
| TGCT | testicular germ cell tumors |
| WG | weeks of gestation |
| WGA | weeks of gestational age |
| ZKD | zinc knuckles domain |
References
- Skakkebaek, N.E.; Rajpert-De Meyts, E.; Buck Louis, G.M.; Toppari, J.; Andersson, A.M.; Eisenberg, M.L.; Jensen, T.K.; Jørgensen, N.; Swan, S.H.; Sapra, K.J.; et al. Male Reproductive Disorders and Fertility Trends: Influences of Environment and Genetic Susceptibility. Physiol. Rev. 2015, 96, 55–97. [Google Scholar] [CrossRef]
- Rotgers, E.; Jørgensen, A.; Yao, H.H.C. At the Crossroads of Fate-Somatic Cell Lineage Specification in the Fetal Gonad. Endocr. Rev. 2018, 39, 739–759. [Google Scholar] [CrossRef] [PubMed]
- Mäkelä, J.A.; Koskenniemi, J.J.; Virtanen, H.E.; Toppari, J. Testis Development. Endocr. Rev. 2019, 40, 857–905. [Google Scholar] [CrossRef] [PubMed]
- Sangiao-Alvarellos, S.; Manfredi-Lozano, M.; Ruiz-Pino, F.; León, S.; Morales, C.; Cordido, F.; Gaytán, F.; Pinilla, L.; Tena-Sempere, M. Testicular Expression of the Lin28/Let-7 System: Hormonal Regulation and Changes during Postnatal Maturation and after Manipulations of Puberty. Sci. Rep. 2015, 5, 15683. [Google Scholar] [CrossRef] [PubMed]
- Moss, E.; Lee, R.; Ambros, V. Control of Developmental Timing by the Cold Shock Domain Protein Lin-28 and Its Regulation by the Lin-4 RNA. Cell 1997, 88, 637–646. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, Y.; Ito, H.; Watanabe, A.; Ge, X.; Kodama, T.; Aburatani, H. Identification and Characterization of Lin-28 Homolog B (LIN28B) in Human Hepatocellular Carcinoma. Gene 2006, 384, 51–61. [Google Scholar] [CrossRef]
- Nam, Y.; Chen, C.; Gregory, R.I.; Chou, J.J.; Sliz, P. Molecular Basis for Interaction of Let-7 MicroRNAs with Lin28. Cell 2011, 147, 1080–1091. [Google Scholar] [CrossRef]
- Darr, H.; Benvenisty, N. Genetic Analysis of the Role of the Reprogramming Gene LIN-28 in Human Embryonic Stem Cells. Stem. Cells 2009, 27, 352–362. [Google Scholar] [CrossRef]
- Rahkonen, N.; Stubb, A.; Malonzo, M.; Edelman, S.; Emani, M.R.; Närvä, E.; Lähdesmäki, H.; Ruohola-Baker, H.; Lahesmaa, R.; Lund, R. Mature Let-7 MiRNAs Fine Tune Expression of LIN28B in Pluripotent Human Embryonic Stem Cells. Stem. Cell Res. 2016, 17, 498–503. [Google Scholar] [CrossRef]
- Viswanathan, S.R.; Daley, G.Q. Lin28: A MicroRNA Regulator with a Macro Role. Cell 2010, 140, 445–449. [Google Scholar] [CrossRef]
- Hu, Z.; Ma, J.; Yue, H.; Li, X.; Wang, C.; Wang, L.; Sun, B.; Chen, Z.; Wang, L.; Gu, Y. A Role for Lin28a in Aging-Associated Decline of Adult Hippocampal Neurogenesis. bioRxiv 2022. [Google Scholar] [CrossRef]
- Kurian, J.; Yuko, A.E.; Kasatkin, N.; Rigaud, V.O.C.; Busch, K.; Harlamova, D.; Wagner, M.; Recchia, F.A.; Wang, H.; Mohsin, S.; et al. Uncoupling Protein 2-Mediated Metabolic Adaptations Define Cardiac Cell Function in the Heart during Transition from Young to Old Age. Stem. Cells Transl. Med. 2021, 10, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Nguyen, C.K.; Liu, X.; Kanellopoulou, C.; Muljo, S.A. Lin28b Reprograms Adult Bone Marrow Hematopoietic Progenitors to Mediate Fetal-like Lymphopoiesis. Science 2012, 335, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Vogt, E.J.; Meglicki, M.; Hartung, K.I.; Borsuk, E.; Behr, R. Importance of the Pluripotency Factor LIN28 in the Mammalian Nucleolus during Early Embryonic Development. Development 2012, 139, 4514–4523. [Google Scholar] [CrossRef]
- Seabrook, J.L.; Cantlon, J.D.; Cooney, A.J.; McWhorter, E.E.; Fromme, B.A.; Bouma, G.J.; Anthony, R.V.; Winger, Q.A. Role of LIN28A in Mouse and Human Trophoblast Cell Differentiation. Biol. Reprod. 2013, 89, 95. [Google Scholar] [CrossRef]
- Ali, A.; Anthony, R.V.; Bouma, G.J.; Winger, Q.A. LIN28-Let-7 Axis Regulates Genes in Immortalized Human Trophoblast Cells by Targeting the ARID3B-Complex. FASEB J. 2019, 33, 12348–12363. [Google Scholar] [CrossRef]
- Canfield, J.; Arlier, S.; Mong, E.F.; Lockhart, J.; VanWye, J.; Guzeloglu-Kayisli, O.; Schatz, F.; Magness, R.R.; Lockwood, C.J.; Tsibris, J.C.M.; et al. Decreased LIN28B in Preeclampsia Impairs Human Trophoblast Differentiation and Migration. FASEB J. 2019, 33, 2759–2769. [Google Scholar] [CrossRef]
- Sangiao-Alvarellos, S.; Manfredi-Lozano, M.; Ruiz-Pino, F.; Navarro, V.M.; Sánchez-Garrido, M.A.; Leon, S.; Dieguez, C.; Cordido, F.; Matagne, V.; Dissen, G.A.; et al. Changes in Hypothalamic Expression of the Lin28/Let-7 System and Related MicroRNAs during Postnatal Maturation and after Experimental Manipulations of Puberty. Endocrinology 2013, 154, 942–955. [Google Scholar] [CrossRef]
- Barbaux, S.; Gascoin-Lachambre, G.; Buffat, C.; Monnier, P.; Mondon, F.; Tonanny, M.B.; Pinard, A.; Auer, J.; Bessières, B.; Barlier, A.; et al. Genome-Wide Approach Reveals Novel Imprinted Genes Expressed in the Human Placenta. Epigenetics 2012, 7, 1079–1090. [Google Scholar] [CrossRef]
- Chan, H.W.; Lappas, M.; Yee, S.W.; Vaswani, K.; Mitchell, M.D.; Rice, G.E. The Expression of the Let-7 MiRNAs and Lin28 Signalling Pathway in Human Term Gestational Tissues. Placenta 2013, 34, 443–448. [Google Scholar] [CrossRef]
- Ali, A.; Bouma, G.J.; Anthony, R.V.; Winger, Q.A. The Role of LIN28-Let-7-ARID3B Pathway in Placental Development. Int. J. Mol. Sci. 2020, 21, 3637. [Google Scholar] [CrossRef] [PubMed]
- Gillis, A.J.M.; Stoop, H.; Biermann, K.; van Gurp, R.J.H.L.M.; Swartzman, E.; Cribbes, S.; Ferlinz, A.; Shannon, M.; Oosterhuis, J.W.; Looijenga, L.H.J. Expression and Interdependencies of Pluripotency Factors LIN28, OCT3/4, NANOG and SOX2 in Human Testicular Germ Cells and Tumours of the Testis. Int. J. Androl. 2011, 34, e160–e174. [Google Scholar] [CrossRef] [PubMed]
- Aeckerle, N.; Eildermann, K.; Drummer, C.; Ehmcke, J.; Schweyer, S.; Lerchl, A.; Bergmann, M.; Kliesch, S.; Gromoll, J.; Schlatt, S.; et al. The Pluripotency Factor LIN28 in Monkey and Human Testes: A Marker for Spermatogonial Stem Cells? Mol. Hum. Reprod. 2012, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Zhang, J.; Ratanasirintrawoot, S.; Chandrasekaran, S.; Wu, Z.; Ficarro, S.B.; Yu, C.; Ross, C.A.; Cacchiarelli, D.; Xia, Q.; Seligson, M.; et al. LIN28 Regulates Stem Cell Metabolism and Conversion to Primed Pluripotency. Cell Stem. Cell 2016, 19, 66–80. [Google Scholar] [CrossRef]
- Zhou, W.; Choi, M.; Margineantu, D.; Margaretha, L.; Hesson, J.; Cavanaugh, C.; Blau, C.A.; Horwitz, M.S.; Hockenbery, D.; Ware, C.; et al. HIF1α Induced Switch from Bivalent to Exclusively Glycolytic Metabolism during ESC-to-EpiSC HESC Transition. EMBO J. 2012, 31, 2103–2116. [Google Scholar] [CrossRef]
- Folmes, C.D.L.; Nelson, T.J.; Martinez-Fernandez, A.; Arrell, D.K.; Lindor, J.Z.; Dzeja, P.P.; Ikeda, Y.; Perez-Terzic, C.; Terzic, A. Somatic Oxidative Bioenergetics Transitions into Pluripotency-Dependent Glycolysis to Facilitate Nuclear Reprogramming. Cell Metab. 2011, 14, 264–271. [Google Scholar] [CrossRef]
- Widén, E.; Ripatti, S.; Cousminer, D.L.; Surakka, I.; Lappalainen, T.; Järvelin, M.R.; Eriksson, J.G.; Raitakari, O.; Salomaa, V.; Sovio, U.; et al. Distinct Variants at LIN28B Influence Growth in Height from Birth to Adulthood. Am. J. Hum. Genet. 2010, 86, 773–782. [Google Scholar] [CrossRef]
- Lanktree, M.B.; Guo, Y.; Murtaza, M.; Glessner, J.T.; Bailey, S.D.; Onland-Moret, N.C.; Lettre, G.; Ongen, H.; Rajagopalan, R.; Johnson, T.; et al. Meta-Analysis of Dense Genecentric Association Studies Reveals Common and Uncommon Variants Associated with Height. Am. J. Hum. Genet. 2011, 88, 6–18. [Google Scholar] [CrossRef]
- Lettre, G.; Jackson, A.U.; Gieger, C.; Schumacher, F.R.; Berndt, S.I.; Sanna, S.; Eyheramendy, S.; Voight, B.F.; Butler, J.L.; Guiducci, C.; et al. Identification of Ten Loci Associated with Height Highlights New Biological Pathways in Human Growth. Nat. Genet. 2008, 40, 584–591. [Google Scholar] [CrossRef]
- Medland, S.E.; Zayats, T.; Glaser, B.; Nyholt, D.R.; Gordon, S.D.; Wright, M.J.; Montgomery, G.W.; Campbell, M.J.; Henders, A.K.; Timpson, N.J.; et al. A Variant in LIN28B Is Associated with 2D:4D Finger-Length Ratio, a Putative Retrospective Biomarker of Prenatal Testosterone Exposure. Am. J. Hum. Genet. 2010, 86, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Leinonen, J.T.; Surakka, I.; Havulinna, A.S.; Kettunen, J.; Luoto, R.; Salomaa, V.; Widén, E. Association of LIN28B with Adult Adiposity-Related Traits in Females. PLoS ONE 2012, 7, e48785. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Kraft, P.; Chen, C.; Buring, J.E.; Paré, G.; Hankinson, S.E.; Chanock, S.J.; Ridker, P.M.; Hunter, D.J.; Chasman, D.I. Genome-Wide Association Studies Identify Loci Associated with Age at Menarche and Age at Natural Menopause. Nat. Genet. 2009, 41, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.K.; Elks, C.E.; Li, S.; Zhao, J.H.; Luan, J.; Andersen, L.B.; Bingham, S.A.; Brage, S.; Smith, G.D.; Ekelund, U.; et al. Genetic Variation in LIN28B Is Associated with the Timing of Puberty. Nat. Genet. 2009, 41, 729–733. [Google Scholar] [CrossRef]
- Perry, J.R.B.; Stolk, L.; Franceschini, N.; Lunetta, K.L.; Zhai, G.; McArdle, P.F.; Smith, A.V.; Aspelund, T.; Bandinelli, S.; Boerwinkle, E.; et al. Meta-Analysis of Genome-Wide Association Data Identifies Two Loci Influencing Age at Menarche. Nat. Genet. 2009, 41, 648–650. [Google Scholar] [CrossRef]
- Leinonen, J.T.; Chen, Y.C.; Tukiainen, T.; Panula, P.; Widén, E. Transient Modification of Lin28b Expression—Permanent Effects on Zebrafish Growth. Mol. Cell. Endocrinol. 2019, 479, 61–70. [Google Scholar] [CrossRef]
- Zhu, H.; Shah, S.; Shyh-Chang, N.; Shinoda, G.; Einhorn, W.S.; Viswanathan, S.R.; Takeuchi, A.; Grasemann, C.; Rinn, J.L.; Lopez, M.F.; et al. Lin28a Transgenic Mice Manifest Size and Puberty Phenotypes Identified in Human Genetic Association Studies. Nat. Genet. 2010, 42, 626–630. [Google Scholar] [CrossRef]
- Shyh-Chang, N.; Zhu, H.; Yvanka De Soysa, T.; Shinoda, G.; Seligson, M.T.; Tsanov, K.M.; Nguyen, L.; Asara, J.M.; Cantley, L.C.; Daley, G.Q. XLin28 Enhances Tissue Repair by Reprogramming Cellular Metabolism. Cell 2013, 155, 778. [Google Scholar] [CrossRef]
- Robinton, D.A.; Chal, J.; Lummertz da Rocha, E.; Han, A.; Yermalovich, A.V.; Oginuma, M.; Schlaeger, T.M.; Sousa, P.; Rodriguez, A.; Urbach, A.; et al. The Lin28/Let-7 Pathway Regulates the Mammalian Caudal Body Axis Elongation Program. Dev. Cell 2019, 48, 396–405.e3. [Google Scholar] [CrossRef]
- Dong, N.; Liu, Y.; Zhang, T.; Zhao, L.; Tian, J.; Ruan, J. Different Expression Patterns of Lin28 and Lin28b in Mouse Molar Development. Arch. Oral Biol. 2017, 82, 280–285. [Google Scholar] [CrossRef]
- Xia, X.; Teotia, P.; Ahmad, I. Lin28a Regulates Neurogliogenesis in Mammalian Retina through the Igf Signaling. Dev. Biol. 2018, 440, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Ambros, V. The C. Elegans Heterochronic Gene Lin-28 Coordinates the Timing of Hypodermal and Somatic Gonadal Programs for Hermaphrodite Reproductive System Morphogenesis. Development 2019, 146, dev164293. [Google Scholar] [CrossRef] [PubMed]
- West, R.C.; McWhorter, E.S.; Ali, A.; Goetzman, L.N.; Russ, J.E.; Gonzalez-Berrios, C.L.; Anthony, R.V.; Bouma, G.J.; Winger, Q.A. HMGA2 Is Regulated by LIN28 and BRCA1 in Human Placental Cells. Biol. Reprod. 2019, 100, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Santoro, G.; Lapucci, C.; Giannoccaro, M.; Caporilli, S.; Rusin, M.; Seidenari, A.; Ferrari, M.; Farina, A. Diagnostics Abnormal Circulating Maternal MiRNA Expression Is Associated with a Low (<4%) Cell-Free DNA Fetal Fraction. Diagnostics 2021, 11, 2108. [Google Scholar] [CrossRef]
- Ali, A.; Stenglein, M.D.; Spencer, T.E.; Bouma, G.J.; Anthony, R.V.; Winger, Q.A. Trophectoderm-Specific Knockdown of LIN28 Decreases Expression of Genes Necessary for Cell Proliferation and Reduces Elongation of Sheep Conceptus. Int. J. Mol. Sci. 2020, 21, 2549. [Google Scholar] [CrossRef]
- Kingsley, P.D.; Palis, J. GRP2 Proteins Contain Both CCHC Zinc Fingers and a Cold Shock Domain. Plant Cell 1994, 6, 1522–1523. [Google Scholar] [CrossRef]
- Mihailovich, M.; Militti, C.; Gabaldón, T.; Gebauer, F. Eukaryotic Cold Shock Domain Proteins: Highly Versatile Regulators of Gene Expression. BioEssays 2010, 32, 109–118. [Google Scholar] [CrossRef]
- Piskounova, E.; Polytarchou, C.; Thornton, J.E.; Lapierre, R.J.; Pothoulakis, C.; Hagan, J.P.; Iliopoulos, D.; Gregory, R.I. Lin28A and Lin28B Inhibit Let-7 MicroRNA Biogenesis by Distinct Mechanisms. Cell 2011, 147, 1066–1079. [Google Scholar] [CrossRef]
- Hafner, M.; Max, K.E.A.; Bandaru, P.; Morozov, P.; Gerstberger, S.; Brown, M.; Molina, H.; Tuschl, T. Identification of MRNAs Bound and Regulated by Human LIN28 Proteins and Molecular Requirements for RNA Recognition. RNA 2013, 19, 613–626. [Google Scholar] [CrossRef]
- Molenaar, J.J.; Domingo-Fernández, R.; Ebus, M.E.; Lindner, S.; Koster, J.; Drabek, K.; Mestdagh, P.; Van Sluis, P.; Valentijn, L.J.; Van Nes, J.; et al. LIN28B Induces Neuroblastoma and Enhances MYCN Levels via Let-7 Suppression. Nat. Genet. 2012, 44, 1199–1206. [Google Scholar] [CrossRef]
- Balzer, E.; Moss, E.G. Localization of the Developmental Timing Regulator Lin28 to MRNP Complexes, P-Bodies and Stress Granules. RNA Biol. 2007, 4, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Childs, A.J.; Kinnell, H.L.; He, J.; Anderson, R.A. LIN28 Is Selectively Expressed by Primordial and Pre-Meiotic Germ Cells in the Human Fetal Ovary. Stem. Cells Dev. 2012, 21, 2343–2349. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, H.; Okada, Y.; Imai, T.; Iwanami, A.; Mischel, P.S.; Okano, H. Musashi1 Cooperates in Abnormal Cell Lineage Protein 28 (Lin28)-Mediated Let-7 Family MicroRNA Biogenesis in Early Neural Differentiation. J. Biol. Chem. 2011, 286, 16121–16130. [Google Scholar] [CrossRef] [PubMed]
- Lott, K.; Cingolani, G. The Importin β Binding Domain as a Master Regulator of Nucleocytoplasmic Transport. Biochim. Biophys. Acta—Mol. Cell Res. 2011, 1813, 1578–1592. [Google Scholar] [CrossRef]
- Pumroy, R.A.; Cingolani, G. Diversification of Importin-α Isoforms in Cellular Trafficking And. Biochem. J. 2015, 466, 13–28. [Google Scholar] [CrossRef]
- Nathaniel, B.; Whiley, P.A.F.; Miyamoto, Y.; Loveland, K.L. Importins: Diverse Roles in Male Fertility. Semin. Cell Dev. Biol. 2022, 121, 82–98. [Google Scholar] [CrossRef]
- Kim, S.; Lee, H.; Han, K.; Kim, S.C.; Choi, Y.; Park, S.; Bak, G.; Lee, Y.; Choi, J.K.; Kim, T.; et al. SET7/9 Methylation of the Pluripotency Factor LIN28A Is a Nucleolar Localization Mechanism That Blocks Let-7 Biogenesis in Human ESCs. Cell Stem. Cell 2015, 15, 735–749. [Google Scholar] [CrossRef]
- Cardoso-Moreira, M.; Halbert, J.; Valloton, D.; Velten, B.; Chen, C.; Shao, Y.; Liechti, A.; Ascenção, K.; Rummel, C.; Ovchinnikova, S.; et al. Gene Expression across Mammalian Organ Development. Nature 2019, 571, 505–509. [Google Scholar] [CrossRef]
- Home < Expression Atlas < EMBL-EBI. Available online: https://www.ebi.ac.uk/gxa/home (accessed on 2 May 2022).
- Felici, M.; Farini, D.; Dolci, S. In or Out Stemness: Comparing Growth Factor Signalling in Mouse Embryonic Stem Cells and Primordial Germ Cells. Curr. Stem. Cell Res. Ther. 2009, 4, 87–97. [Google Scholar] [CrossRef]
- Kispert, A.; Gossler, A. Introduction to Early Mouse Development; Elsevier Ltd.: Amsterdam, The Netherlands, 2004. [Google Scholar] [CrossRef]
- Guo, F.; Yan, L.; Guo, H.; Li, L.; Hu, B.; Zhao, Y.; Yong, J.; Hu, Y.; Wang, X.; Wei, Y.; et al. The Transcriptome and DNA Methylome Landscapes of Human Primordial Germ Cells. Cell 2015, 161, 1437–1452. [Google Scholar] [CrossRef]
- De Felici, M. Origin, Migration, and Proliferation of Human Primordial Germ Cells. In Oogenesis; Springer: Berlin/Heidelberg, Germany, 2013; pp. 19–37. [Google Scholar] [CrossRef]
- Sonne, S.B.; Almstrup, K.; Dalgaard, M.; Juncker, A.S.; Edsgard, D.; Ruban, L.; Harrison, N.J.; Schwager, C.; Abdollahi, A.; Huber, P.E.; et al. Analysis of Gene Expression Profiles of Microdissected Cell Populations Indicates That Testicular Carcinoma in Situ Is an Arrested Gonocyte. Cancer Res. 2009, 69, 5241–5250. [Google Scholar] [CrossRef] [PubMed]
- Buljubašić, R.; Buljubašić, M.; Bojanac, A.K.; Ulamec, M.; Vlahović, M.; Ježek, D.; Bulić-Jakuš, F.; Sinčić, N. Epigenetics and Testicular Germ Cell Tumors. Gene 2018, 661, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, T.L.; Esnal, A.; Robinson, L.L.L.; Anderson, R.A.; Saunders, P.T.K. Immunohistochemical Profiling of Germ Cells Within the Human Fetal Testis: Identification of Three Subpopulations. Biol. Reprod. 2004, 71, 2012–2021. [Google Scholar] [CrossRef]
- Guo, J.; Sosa, E.; Chitiashvili, T.; Nie, X.; Rojas, E.J.; Oliver, E.; Plath, K.; Hotaling, J.M.; Stukenborg, J.B.; Clark, A.T.; et al. Single-Cell Analysis of the Developing Human Testis Reveals Somatic Niche Cell Specification and Fetal Germline Stem Cell Establishment. Cell Stem. Cell 2021, 28, 764–778.e4. [Google Scholar] [CrossRef]
- Li, L.; Dong, J.; Yan, L.; Yong, J.; Liu, X.; Hu, Y.; Fan, X.; Wu, X.; Guo, H.; Wang, X.; et al. Single-Cell RNA-Seq Analysis Maps Development of Human Germline Cells and Gonadal Niche Interactions. Cell Stem. Cell 2017, 20, 858–873.e4. [Google Scholar] [CrossRef] [PubMed]
- Picut, C.A.; Ziejewski, M.K.; Stanislaus, D. Comparative Aspects of Pre- and Postnatal Development of the Male Reproductive System. Birth Defects Res. 2018, 110, 190–227. [Google Scholar] [CrossRef]
- Shinoda, G.; De Soysa, T.Y.; Seligson, M.T.; Yabuuchi, A.; Fujiwara, Y.; Huang, P.Y.; Hagan, J.P.; Gregory, R.I.; Moss, E.G.; Daley, G.Q. Lin28a Regulates Germ Cell Pool Size and Fertility. Stem Cells 2013, 31, 1001–1009. [Google Scholar] [CrossRef]
- Hammoud, S.S.; Low, D.H.P.; Yi, C.; Lee, C.L.; Oatley, J.M.; Payne, C.J.; Carrell, D.T.; Guccione, E.; Cairns, B.R. Transcription and Imprinting Dynamics in Developing Postnatal Male Germline Stem Cells. Genes Dev. 2015, 29, 2312–2324. [Google Scholar] [CrossRef]
- Gaytan, F.; Sangiao-Alvarellos, S.; Manfredi-Lozano, M.; García-Galiano, D.; Ruiz-Pino, F.; Romero-Ruiz, A.; León, S.; Morales, C.; Cordido, F.; Pinilla, L.; et al. Distinct Expression Patterns Predict Differential Roles of the Mirna-Binding Proteins, Lin28 and Lin28b, in the Mouse Testis: Studies during Postnatal Development and in a Model of Hypogonadotropic Hypogonadism. Endocrinology 2013, 154, 1321–1336. [Google Scholar] [CrossRef]
- Sohni, A.; Tan, K.; Song, H.W.; Burow, D.; de Rooij, D.G.; Laurent, L.; Hsieh, T.C.; Rabah, R.; Hammoud, S.S.; Vicini, E.; et al. The Neonatal and Adult Human Testis Defined at the Single-Cell Level. Cell Rep. 2019, 26, 1501–1517.e4. [Google Scholar] [CrossRef]
- Yan, W.; Ma, L.; Burns, K.H.; Matzuk, M.M. HILS1 Is a Spermatid-Specific Linker Histone H1-like Protein Implicated in Chromatin Remodeling during Mammalian Spermiogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 10546–10551. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Baba, T. Gene Expression in Spermiogenesis. Cell. Mol. Life Sci. 2005, 62, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Jun-Hao, E.T.; Gupta, R.R.; Shyh-Chang, N. Lin28 and Let-7 in the Metabolic Physiology of Aging. Trends Endocrinol. Metab. 2016, 27, 132–141. [Google Scholar] [CrossRef]
- Corre, C.; Shinoda, G.; Zhu, H.; Cousminer, D.L.; Crossman, C.; Bellissimo, C.; Goldenberg, A.; Daley, G.Q.; Palmert, M.R. Sex-Specific Regulation of Weight and Puberty by the Lin28/Let-7 Axis. J. Endocrinol. 2016, 228, 179–191. [Google Scholar] [CrossRef]
- Sreejith, P.; Jang, W.; To, V.; Jo, Y.H.; Biteau, B.; Kim, C. Lin28 Is a Critical Factor in the Function and Aging of Drosophila Testis Stem Cell Niche. Aging 2019, 11, 855–873. [Google Scholar] [CrossRef]
- Wang, D.; Hou, L.; Nakamura, S.; Su, M.; Li, F.; Chen, W.; Yan, Y.; Green, C.D.; Chen, D.; Zhang, H.; et al. LIN-28 Balances Longevity and Germline Stem Cell Number in Caenorhabditis Elegans through Let-7/AKT/DAF-16 Axis. Aging Cell 2017, 16, 113–124. [Google Scholar] [CrossRef]
- Leitch, H.G.; Smith, A. The Mammalian Germline as a Pluripotency Cycle. Development 2013, 140, 2495–2501. [Google Scholar] [CrossRef]
- Heo, I.; Joo, C.; Kim, Y.K.; Ha, M.; Yoon, M.J.; Cho, J.; Yeom, K.H.; Han, J.; Kim, V.N. TUT4 in Concert with Lin28 Suppresses MicroRNA Biogenesis through Pre-MicroRNA Uridylation. Cell 2009, 138, 696–708. [Google Scholar] [CrossRef]
- Jin, J.; Jing, W.; Lei, X.X.; Feng, C.; Peng, S.; Boris-Lawrie, K.; Huang, Y. Evidence That Lin28 Stimulates Translation by Recruiting RNA Helicase A to Polysomes. Nucleic Acids Res. 2011, 39, 3724–3734. [Google Scholar] [CrossRef]
- Wilbert, M.L.; Huelga, S.C.; Kapeli, K.; Stark, T.J.; Liang, T.Y.; Chen, S.X.; Yan, B.Y.; Nathanson, J.L.; Hutt, K.R.; Lovci, M.T.; et al. LIN28 Binds Messenger RNAs at GGAGA Motifs and Regulates Splicing Factor Abundance. Mol. Cell 2012, 48, 195–206. [Google Scholar] [CrossRef]
- Polesskaya, A.; Cuvellier, S.; Naguibneva, I.; Duquet, A.; Moss, E.G.; Harel-Bellan, A. Lin-28 Binds IGF-2 MRNA and Participates in Skeletal Myogenesis by Increasing Translation Efficiency. Genes Dev. 2007, 21, 1125–1138. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.I.H.; Lee, J.H.; Kim, S.Y.; Cho, K.O. Expression of Exogenous LIN28 Contributes to Proliferation and Survival of Mouse Primary Cortical Neurons in Vitro. Neuroscience 2013, 248, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Bateman, J.M.; McNeill, H. Insulin/IGF Signalling in Neurogenesis. Cell. Mol. Life Sci. 2006, 63, 1701–1705. [Google Scholar] [CrossRef] [PubMed]
- Zha, J.; Lackner, M.R. Targeting the Insulin-like Growth Factor Receptor-1R Pathway for Cancer Therapy. Clin. Cancer Res. 2010, 16, 2512–2517. [Google Scholar] [CrossRef]
- Balzer, E.; Heine, C.; Jiang, Q.; Lee, V.M.; Moss, E.G. LIN28 Alters Cell Fate Succession and Acts Independently of the Let-7 MicroRNA during Neurogliogenesis In Vitro. Development 2010, 137, 891–900. [Google Scholar] [CrossRef]
- Zhu, H.; Ng, S.C.; Segr, A.V.; Shinoda, G.; Shah, S.P.; Einhorn, W.S.; Takeuchi, A.; Engreitz, J.M.; Hagan, J.P.; Kharas, M.G.; et al. The Lin28/Let-7 Axis Regulates Glucose Metabolism. Cell 2011, 147, 81–94. [Google Scholar] [CrossRef]
- Zhang, Y.; Williams, P.R.; Jacobi, A.; Wang, C.; Goel, A.; Hirano, A.A.; Brecha, N.C.; Kerschensteiner, D.; He, Z. Elevating Growth Factor Responsiveness and Axon Regeneration by Modulating Presynaptic Inputs. Neuron 2019, 103, 39–51.e5. [Google Scholar] [CrossRef]
- Bingsen, X.U.; Zhang, K.; Huang, Y. Lin28 Modulates Cell Growth and Associates with a Subset of Cell Cycle Regulator MRNAs in Mouse Embryonic Stem Cells. RNA 2009, 15, 357–361. [Google Scholar] [CrossRef]
- Xu, B.; Huang, Y. Histone H2a MRNA Interacts with Lin28 and Contains a Lin28-Dependent Posttranscriptional Regulatory Element. Nucleic Acids Res. 2009, 37, 4256–4263. [Google Scholar] [CrossRef]
- Qiu, C.; Ma, Y.; Wang, J.; Peng, S.; Huang, Y. Lin28-Mediated Post-Transcriptional Regulation of Oct4 Expression in Human Embryonic Stem Cells. Nucleic Acids Res. 2009, 38, 1240–1248. [Google Scholar] [CrossRef]
- Cho, J.; Chang, H.; Kwon, S.C.; Kim, B.; Kim, Y.; Choe, J.; Ha, M.; Kim, Y.K.; Kim, V.N. LIN28A Is a Suppressor of ER-Associated Translation in Embryonic Stem Cells. Cell 2012, 151, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Chen, L.L.; Lei, X.X.; Yang, L.; Lin, H.; Carmichael, G.G.; Huang, Y. Genome-Wide Studies Reveal That Lin28 Enhances the Translation of Genes Important for Growth and Survival of Human Embryonic Stem Cells. Stem. Cells 2011, 29, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, L.; Wang, S.; Yang, F.; Wang, M.; Li, L.; Wu, X. LIN28A Binds to Meiotic Gene Transcripts and Modulates Their Translation in Male Germ Cells. J. Cell Sci. 2020, 133. [Google Scholar] [CrossRef] [PubMed]
- Graf, R.; Munschauer, M.; Mastrobuoni, G.; Mayr, F.; Heinemann, U.; Kempa, S.; Rajewsky, N.; Landthaler, M. Identification of LIN28B-Bound MRNAs Reveals Features of Target Recognition and Regulation. RNA Biol. 2013, 10, 1146–1159. [Google Scholar] [CrossRef]
- Shinoda, G.; Shyh-Chang, N.; Yvanka De Soysa, T.; Zhu, H.; Seligson, M.T.; Shah, S.P.; Abo-Sido, N.; Yabuuchi, A.; Hagan, J.P.; Gregory, R.I. Fetal Deficiency of Lin28 Programs Life-Long Aberrations in Growth and Glucose Metabolism. Stem. Cells 2013, 31, 1563–1573. [Google Scholar] [CrossRef]
- Mayr, F.; Schütz, A.; Döge, N.; Heinemann, U. The Lin28 Cold-Shock Domain Remodels Pre-Let-7 MicroRNA. Nucleic Acids Res. 2012, 40, 7492–7506. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, X.; Greshock, J.; Shen, L.; Yang, X. Genomic DNA Copy-Number Alterations of the Let-7 Family in Human Cancers. PLoS ONE 2012, 7, 44399. [Google Scholar] [CrossRef]
- Johnson, S.M.; Grosshans, H.; Shingara, J.; Byrom, M.; Jarvis, R.; Cheng, A.; Labourier, E.; Reinert, K.L.; Brown, D.; Slack, F.J. RAS Is Regulated by the Let-7 MicroRNA Family. Cell 2005, 120, 635–647. [Google Scholar] [CrossRef]
- Lee, Y.S.; Dutta, A. The Tumor Suppressor MicroRNA Let-7 Represses the HMGA2 Oncogene. Genes Dev. 2007, 21, 1025–1030. [Google Scholar] [CrossRef]
- Worringer, K.A.; Rand, T.A.; Hayashi, Y.; Sami, S.; Takahashi, K.; Tanabe, K.; Narita, M.; Srivastava, D.; Yamanaka, S. The Let-7/LIN-41 Pathway Regulates Reprogramming to Human Induced Pluripotent Stem Cells by Controlling Expression of Prodifferentiation Genes. Cell Stem. Cell 2014, 14, 40–52. [Google Scholar] [CrossRef]
- Newman, M.A.; Thomson, J.M.; Hammond, S.M. Lin-28 Interaction with the Let-7 Precursor Loop Mediates Regulated MicroRNA Processing. RNA 2008, 14, 1539–1549. [Google Scholar] [CrossRef] [PubMed]
- Martinez, N.J.; Gregory, R.I. MicroRNA Gene Regulatory Pathways in the Establishment and Maintenance of ESC Identity. Cell Stem. Cell 2010, 7, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.J.A.; Olson, E.N. Control of Glucose Homeostasis and Insulin Sensitivity by the Let-7 Family of MicroRNAs. Proc. Natl. Acad. Sci. USA 2011, 108, 21075–21080. [Google Scholar] [CrossRef]
- King, C.E.; Wang, L.; Winograd, R.; Madison, B.B.; Mongroo, P.S.; Johnstone, C.N.; Rustgi, A.K. LIN28B Fosters Colon Cancer Migration, Invasion and Transformation through Let-7-Dependent and-Independent Mechanisms. Oncogene 2011, 30, 4185–4193. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Katsaros, D.; Shaverdashvili, K.; Qian, B.; Wu, Y.; de la Longrais, I.A.R.; Preti, M.; Menato, G.; Yu, H. Pluripotent Factor Lin-28 and Its Homologue Lin-28b in Epithelial Ovarian Cancer and Their Associations with Disease Outcomes and Expression of Let-7a and IGF-II. Eur. J. Cancer 2009, 45, 2212–2218. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, M.; Miki, Y.; Masuda, M.; Hata, S.; Shibahara, Y.; Hirakawa, H.; Suzuki, T.; Sasano, H. LIN28: A Regulator of Tumor-Suppressing Activity of Let-7 MicroRNA in Human Breast Cancer. J. Steroid Biochem. Mol. Biol. 2012, 131, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Allan, R.W.; Cheng, L.; Peng, Y.; Guo, C.C.; Dahiya, N.; Akhi, S.; Li, J. RNA-Binding Protein LIN28 Is a Marker for Testicular Germ Cell Tumors. Hum. Pathol. 2011, 42, 710–718. [Google Scholar] [CrossRef]
- West, J.A.; Viswanathan, S.R.; Yabuuchi, A.; Cunniff, K.; Takeuchi, A.; Park, I.H.; Sero, J.E.; Zhu, H.; Perez-Atayde, A.; Frazier, A.L.; et al. A Role for Lin28 in Primordial Germ-Cell Development and Germ-Cell Malignancy. Nature 2009, 460, 909–913. [Google Scholar] [CrossRef]
- Hamano, R.; Miyata, H.; Yamasaki, M.; Sugimura, K.; Tanaka, K.; Kurokawa, Y.; Nakajima, K.; Takiguchi, S.; Fujiwara, Y.; Mori, M.; et al. High Expression of Lin28 Is Associated with Tumour Aggressiveness and Poor Prognosis of Patients in Oesophagus Cancer. Br. J. Cancer 2012, 106, 1415–1423. [Google Scholar] [CrossRef]
- Hovestadt, V.; Jones, D.T.W.; Picelli, S.; Wang, W.; Kool, M.; Northcott, P.A.; Sultan, M.; Stachurski, K.; Ryzhova, M.; Warnatz, H.J.; et al. Decoding the Regulatory Landscape of Medulloblastoma Using DNA Methylation Sequencing. Nature 2014, 510, 537–541. [Google Scholar] [CrossRef]
- Manier, S.; Powers, J.T.; Sacco, A.; Glavey, S.V.; Huynh, D.; Reagan, M.R.; Salem, K.Z.; Moschetta, M.; Shi, J.; Mishima, Y.; et al. The LIN28B/Let-7 Axis Is a Novel Therapeutic Pathway in Multiple Myeloma; Nature Publishing Group: Berlin, Germany, 2017; Volume 31. [Google Scholar] [CrossRef]
- Zhou, J.; Chan, Z.L.; Bi, C.; Lu, X.; Chong, P.S.Y.; Chooi, J.Y.; Cheong, L.L.; Liu, S.C.; Ching, Y.Q.; Zhou, Y.; et al. LIN28B Activation by PRL-3 Promotes Leukemogenesis and a Stem Cell-like Transcriptional Program in AML. Mol. Cancer Res. 2017, 15, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, C.; Sun, L.; Huang, D.; Li, T.; He, X.; Wu, G.; Yang, Z.; Zhong, X.; Song, L.; et al. Lin28/Let-7 Axis Regulates Aerobic Glycolysis and Cancer Progression via PDK1. Nat. Commun. 2014, 5, 5212. [Google Scholar] [CrossRef] [PubMed]
- Yuko, A.E.; Carvalho Rigaud, V.O.; Kurian, J.; Lee, J.H.; Kasatkin, N.; Behanan, M.; Wang, T.; Luchesse, A.M.; Mohsin, S.; Koch, W.J.; et al. LIN28a Induced Metabolic and Redox Regulation Promotes Cardiac Cell Survival in the Heart after Ischemic Injury. Redox Biol. 2021, 47, 102162. [Google Scholar] [CrossRef] [PubMed]
- Mayr, F.; Heinemann, U. Mechanisms of Lin28-Mediated MiRNA and MRNA Regulation-a Structural and Functional Perspective. Int. J. Mol. Sci. 2013, 14, 16532–16553. [Google Scholar] [CrossRef]
- Loughlin, F.E.; Gebert, L.F.R.; Towbin, H.; Brunschweiger, A.; Hall, J.; Allain, F.H.T. Structural Basis of Pre-Let-7 MiRNA Recognition by the Zinc Knuckles of Pluripotency Factor Lin28. Nat. Struct. Mol. Biol. 2012, 19, 84–91. [Google Scholar] [CrossRef]
- Piskounova, E.; Viswanathan, S.R.; Janas, M.; LaPierre, R.J.; Daley, G.Q.; Sliz, P.; Gregory, R.I. Determinants of MicroRNA Processing Inhibition by the Developmentally Regulated RNA-Binding Protein Lin28. J. Biol. Chem. 2008, 283, 21310–21314. [Google Scholar] [CrossRef]
- Hagan, J.P.; Piskounova, E.; Gregory, R.I. Lin28 Recruits the TUTase Zcchc11 to Inhibit Let-7 Maturation in Mouse Embryonic Stem Cells. Nat. Struct. Mol. Biol. 2009, 16, 1021–1025. [Google Scholar] [CrossRef]
- Heo, I.; Joo, C.; Cho, J.; Ha, M.; Han, J.; Kim, V.N. Lin28 Mediates the Terminal Uridylation of Let-7 Precursor MicroRNA. Mol. Cell 2008, 32, 276–284. [Google Scholar] [CrossRef]
- Thornton, J.E.; Chang, H.M.; Piskounova, E.; Gregory, R.I. Lin28-Mediated Control of Let-7 MicroRNA Expression by Alternative TUTases Zcchc11 (TUT4) and Zcchc6 (TUT7). RNA 2012, 18, 1875–1885. [Google Scholar] [CrossRef]
- Wang, L.; Nam, Y.; Lee, A.K.; Yu, C.; Roth, K.; Chen, C.; Ransey, E.M.; Sliz, P. LIN28 Zinc Knuckle Domain Is Required and Sufficient to Induce Let-7 Oligouridylation. Cell Rep. 2017, 18, 2664–2675. [Google Scholar] [CrossRef]
- Kolenda, T.; Przybyla, W.; Teresiak, A.; Mackiewicz, A.; Lamperska, K.M. The Mystery of Let-7d—A Small RNA with Great Power. Wspolczesna Onkol. 2014, 18, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Ustianenko, D.; Chiu, H.S.; Treiber, T.; Weyn-Vanhentenryck, S.M.; Treiber, N.; Meister, G.; Sumazin, P.; Zhang, C. LIN28 Selectively Modulates a Subclass of Let-7 MicroRNAs. Mol. Cell 2018, 71, 271–283.e5. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Bian, S.; Li, J.; He, J.; Chen, H.; Ge, L.; Jiao, Z.; Zhang, Y.; Peng, W.; Du, F.; et al. MeCP2 Suppresses LIN28A Expression via Binding to Its Methylated-CpG Islands in Pancreatic Cancer Cells. Oncotarget 2016, 7, 14476–14485. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, Y.; Yang, J.; Gu, Y.; Zhang, E.; Yuan, W.; Wang, C.; Jin, G.; Ma, H.; Hu, Z. Hypomethylation-Activated Cancer-Testis Gene LIN28B Promotes Cell Proliferation and Metastasis in Gastric Cancer. Gene 2022, 813, 146115. [Google Scholar] [CrossRef]
- Wang, L.X.; Wang, J.; Qu, T.T.; Zhang, Y.; Shen, Y.F. Reversible Acetylation of Lin28 Mediated by PCAF and SIRT1. Biochim. Biophys. Acta—Mol. Cell Res. 2014, 1843, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Tsanov, K.M.; Pearson, D.S.; Wu, Z.; Han, A.; Triboulet, R.; Seligson, M.T.; Powers, J.T.; Osborne, J.K.; Kane, S.; Gygi, S.P.; et al. LIN28 Phosphorylation by MAPK/ERK Couples Signalling to the Post-Transcriptional Control of Pluripotency. Nat. Cell Biol. 2017, 19, 60–67. [Google Scholar] [CrossRef]
- Haq, S.; Das, S.; Kim, D.H.; Chandrasekaran, A.P.; Hong, S.H.; Kim, K.S.; Ramakrishna, S. The Stability and Oncogenic Function of LIN28A Are Regulated by USP28. Biochim. Biophys. Acta—Mol. Basis Dis. 2019, 1865, 599–610. [Google Scholar] [CrossRef]
- Kim, C.W.; Vo, M.T.; Kim, H.K.; Lee, H.H.; Yoon, N.A.; Lee, B.J.; Min, Y.J.; Joo, W.D.; Cha, H.J.; Park, J.W.; et al. Ectopic Over-Expression of Tristetraprolin in Human Cancer Cells Promotes Biogenesis of Let-7 by down-Regulation of Lin28. Nucleic Acids Res. 2012, 40, 3856–3869. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, H.J.; Yoon, N.A.; Lee, W.H.; Min, Y.J.; Ko, B.K.; Lee, B.J.; Lee, A.; Cha, H.J.; Cho, W.J.; et al. Tumor Suppressor P53 Plays a Key Role in Induction of Both Tristetraprolin and Let-7 in Human Cancer Cells. Nucleic Acids Res. 2013, 41, 5614–5625. [Google Scholar] [CrossRef]
- Chen, T.; Chen, C.; Wu, H.; Chen, X.; Xie, R.; Wang, F.; Sun, H.; Chen, L. Overexpression of P53 Accelerates Puberty in High-Fat Diet–Fed Mice through Lin28/Let-7 System. Exp. Biol. Med. 2021, 246, 66–71. [Google Scholar] [CrossRef]
- Tocchini, C.; Ciosk, R. TRIM-NHL Proteins in Development and Disease. Semin. Cell Dev. Biol. 2015, 47–48, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Lipkowitz, S.; Weissman, A.M. RINGs of Good and Evil: RING Finger Ubiquitin Ligases at the Crossroads of Tumour Suppression and Oncogenesis. Nat. Rev. Cancer 2011, 11, 629–643. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Cho, S.; Sun Kim, M.; Choi, K.; Cho, J.Y.; Gwak, H.S.; Kim, Y.J.; Yoo, H.; Lee, S.H.; Park, J.B.; et al. The Ubiquitin Ligase Human TRIM71 Regulates Let-7 MicroRNA Biogenesis via Modulation of Lin28B Protein. Biochim. Biophys. Acta—Gene Regul. Mech. 2014, 1839, 374–386. [Google Scholar] [CrossRef]
- Yin, J.; Kim, T.-H.; Park, N.; Shin, D.; Choi, H.I.; Cho, S.; Park, J.B.; Kim, J.H.; Bae, J. TRIM71 Suppresses Tumorigenesis via Modulation of Lin28B-Let-7-HMGA2 Signaling. Oncotarget 2016, 7, 79854–79868. [Google Scholar] [CrossRef]
- Singh, R.; Junghare, V.; Hazra, S.; Singh, U.; Sengar, G.S.; Raja, T.V.; Kumar, S.; Tyagi, S.; Das, A.K.; Kumar, A.; et al. Database on Spermatozoa Transcriptogram of Catagorised Frieswal Crossbred (Holstein Friesian X Sahiwal) Bulls. Theriogenology 2019, 129, 130–145. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Li, N.; Liang, S.; Huang, Q.; Coukos, G.; Zhang, L. Identification of MicroRNAs Regulating Reprogramming Factor LIN28 in Embryonic Stem Cells and Cancer Cells. J. Biol. Chem. 2010, 285, 41961–41971. [Google Scholar] [CrossRef]
- Wu, L.; Belasco, J.G. Micro-RNA Regulation of the Mammalian. Society 2005, 25, 9198–9208. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.; Gao, L.; McClellan, S.; Finan, M.A.; Butler, T.W.; Owen, L.B.; Piazza, G.A.; Xi, Y. MiR-181 Mediates Cell Differentiation by Interrupting the Lin28 and Let-7 Feedback Circuit. Cell Death Differ. 2012, 19, 378–386. [Google Scholar] [CrossRef]
- Murray, M.J.; Saini, H.K.; Siegler, C.A.; Hanning, J.E.; Barker, E.M.; Van Dongen, S.; Ward, D.M.; Raby, K.L.; Groves, I.J.; Scarpini, C.G.; et al. LIN28 Expression in Malignant Germ Cell Tumors Downregulates Let-7 and Increases Oncogene Levels. Cancer Res. 2013, 73, 4872–4884. [Google Scholar] [CrossRef]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The LIN28/Let-7 Pathway in Cancer. Front. Genet. 2017, 8, 31. [Google Scholar] [CrossRef]
- Murray, M.J.; Nicholson, J.C.; Coleman, N. Biology of Childhood Germ Cell Tumours, Focussing on the Significance of MicroRNAs. Andrology 2015, 3, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Buaas, F.W.; Sharma, M.; Snyder, E.; Rooij, D.G.; Braun, R.E.; William Buaas, F.; De Rooij, D.G. LIN28A Marks the Spermatogonial Progenitor Population and Regulates Its Cyclic Expansion. Stem. Cells 2014, 32, 860–873. [Google Scholar] [CrossRef]
- Werler, S.; Demond, H.; Damm, O.S.; Ehmcke, J.; Middendorff, R.; Gromoll, J.; Wistuba, J. Germ Cell Loss Is Associated with Fading Lin28a Expression in a Mouse Model for Klinefelter’s Syndrome. Reproduction 2014, 147, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Wu, X.; Kaestner, K.H.; Wang, P.J. The Pluripotency Factor LIN28 Marks Undifferentiated Spermatogonia in Mouse. BMC Dev. Biol. 2009, 9, 38. [Google Scholar] [CrossRef]
- Sèdes, L.; Desdoits-Lethimonier, C.; Rouaisnel, B.; Holota, H.; Thirouard, L.; Lesne, L.; Damon-Soubeyrand, C.; Martinot, E.; Saru, J.P.; Mazaud-Guittot, S.; et al. Crosstalk between BPA and FXRα Signaling Pathways Lead to Alterations of Undifferentiated Germ Cell Homeostasis and Male Fertility Disorders. Stem. Cell Rep. 2018, 11, 944–958. [Google Scholar] [CrossRef]
- Brieño-Enríquez, M.A.; García-López, J.; Cárdenas, D.B.; Guibert, S.; Cleroux, E.; Děd, L.; Hourcade, J.D.D.; Pěknicová, J.; Weber, M.; Del Mazo, J. Exposure to Endocrine Disruptor Induces Transgenerational Epigenetic Deregulation of MicroRNAs in Primordial Germ Cells. PLoS ONE 2015, 10, e0124296. [Google Scholar] [CrossRef]
- Rochester, J.R. Bisphenol A and Human Health: A Review of the Literature. Reprod. Toxicol. 2013, 42, 132–155. [Google Scholar] [CrossRef]
- Anway, M.D.; Cupp, A.S.; Uzumcu, N.; Skinner, M.K. Toxicology: Epigenetic Transgenerational Actions of Endocrine Disruptors and Male Fertility. Science 2005, 308, 1466–1469. [Google Scholar] [CrossRef]
- Matsuura, I.; Saitoh, T.; Ashina, M.; Wako, Y.; Iwata, H.; Toyota, N.; Ishizuka, Y.; Namiki, M.; Hoshino, N.; Tsuchitani, M.; et al. Evaluation of a Two-Generation Reproduction Toxicity Study Adding Endopoints to Detect Endocrine Disrupting Activity Using Vinclozolin. J. Toxicol. Sci. 2005, 30, 163–188. [Google Scholar] [CrossRef]
- Spiller, C.M.; Bowles, J. Germ Cell Neoplasia in Situ: The Precursor Cell for Invasive Germ Cell Tumors of the Testis. Int. J. Biochem. Cell Biol. 2017, 86, 22–25. [Google Scholar] [CrossRef]
- Lv, K.; Liu, L.; Wang, L.; Yu, J.; Liu, X.; Cheng, Y.; Dong, M.; Teng, R.; Wu, L.; Fu, P.; et al. Lin28 Mediates Paclitaxel Resistance by Modulating P21, Rb and Let-7a MiRNA in Breast Cancer Cells. PLoS ONE 2012, 7, e40008. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yuan, C.; Lv, K.; Xie, S.; Fu, P.; Liu, X.; Chen, Y.; Qin, C.; Deng, W.; Hu, W. Lin28 Mediates Radiation Resistance of Breast Cancer Cells via Regulation of Caspase, H2AX and Let-7 Signaling. PLoS ONE 2013, 8, e67373. [Google Scholar] [CrossRef]
- Jee-Sun Oh, M.S.; Jae-Jin Kim, B.S.; Ju-Yeon Byun, M.S.; In-Ah Kim, M.D. Lin28-let7 modulates radiosensitivity of human cancer cells with activation of k-ras. Int. J. Radiat. Oncol. * Biol. * Phys. 2010, 76, 5–8. [Google Scholar] [CrossRef]
- Qin, R.; Zhou, J.; Chen, C.; Xu, T.; Yan, Y.; Ma, Y.; Zheng, Z.; Shen, Y.; Lu, Y.; Fu, D.; et al. LIN28 Is Involved in Glioma Carcinogenesis and Predicts Outcomes of Glioblastoma Multiforme Patients. PLoS ONE 2014, 9, e86446. [Google Scholar] [CrossRef]
- Kong, D.; Banerjee, S.; Ahmad, A.; Li, Y.; Wang, Z.; Sethi, S.; Sarkar, F.H. Epithelial to Mesenchymal Transition Is Mechanistically Linked with Stem Cell Signatures in Prostate Cancer Cells. PLoS ONE 2010, 5, e12445. [Google Scholar] [CrossRef]
- Zhang, H.; Zong, Y.; Qiu, G.; Jia, R.; Xu, X.; Wang, F.; Wu, D. Silencing Lin28 Promotes Apoptosis in Colorectal Cancer Cells by Upregulating Let-7c Targeting of Antiapoptotic BCL2L1. Mol. Med. Rep. 2018, 17, 5143–5149. [Google Scholar] [CrossRef]
- Chen, Y.; Xie, C.; Zheng, X.; Nie, X.; Wang, Z.; Liu, H.; Zhao, Y. LIN28/Let-7/PD-L1 Pathway as a Target for Cancer Immunotherapy. Cancer Immunol. Res. 2019, 7, 487–497. [Google Scholar] [CrossRef]
- Sikora, K.; Evan, G.; Stewart, J.; Watson, J.V. Detection of the C-Myc Oncogene Product in Testicular Cancer. Br. J. Cancer 1985, 52, 171–176. [Google Scholar] [CrossRef]
- Roos, M.; Pradère, U.; Ngondo, R.P.; Behera, A.; Allegrini, S.; Civenni, G.; Zagalak, J.A.; Marchand, J.R.; Menzi, M.; Towbin, H.; et al. A Small-Molecule Inhibitor of Lin28. ACS Chem. Biol. 2016, 11, 2773–2781. [Google Scholar] [CrossRef]
- Wang, L.; Rowe, R.G.; Jaimes, A.; Yu, C.; Nam, Y.; Pearson, D.S.; Zhang, J.; Xie, X.; Marion, W.; Heffron, G.J.; et al. Small-Molecule Inhibitors Disrupt Let-7 Oligouridylation and Release the Selective Blockade of Let-7 Processing by LIN28. Cell Rep. 2018, 23, 3091–3101. [Google Scholar] [CrossRef]
- Chen, H.; Sa, G.; Li, L.; He, S.; Wu, T. In Vitro and in Vivo Synergistic Anti-Tumor Effect of LIN28 Inhibitor and Metformin in Oral Squamous Cell Carcinoma. Eur. J. Pharmacol. 2021, 891, 173757. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.H.; Kim, T.H.; Jo, A.Y.; Chang, M.Y.; Park, C.H.; Kim, S.M.; Song, J.J.; Oh, S.M.; Yi, S.H.; Kim, H.H.; et al. LIN28A Enhances the Therapeutic Potential of Cultured Neural Stem Cells in a Parkinson’s Disease Model. Brain 2016, 139, 2722–2739. [Google Scholar] [CrossRef] [PubMed]
- Nathan, F.M.; Ohtake, Y.; Wang, S.; Jiang, X.; Sami, A.; Guo, H.; Zhou, F.Q.; Li, S. Upregulating Lin28a Promotes Axon Regeneration in Adult Mice with Optic Nerve and Spinal Cord Injury. Mol. Ther. 2020, 28, 1902–1917. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LIN28 | Role | Model | Reference |
|---|---|---|---|
| LIN28AB | stem cell self-renewal 1 cell reprograming 2 | mouse 1,2 and human 1 | [24] 1 [25] 1,2 |
| ESCs differentiation | human | [8,9] | |
| body size 1 onset of puberty 2 | transgenic mouse 1,2 human 2 | [33] 1,2 [35] 1,2 [37] 1,2 [34] 2 [36] 2 | |
| tail development | mouse | [39] | |
| tooth development | mouse | [40] | |
| trophoblast proliferation | human 1 and sheep 2 | [16] 1 [43] 1 [45] 2 | |
| developmental timing | C. elegans | [5] | |
| LIN28A paralog | heart development | mouse | [12] |
| neurogliogenesis | Rat and mouse | [41] | |
| ovulation | C. elegans | [42] | |
| trophoblast differentiation | human | [15] | |
| mesodermal and neural cell fate | mouse | [39] | |
| hippocampal neurogenesis | mouse | [11] | |
| germline stem cells self-renewal | human | [22] 1 [23] 1 | |
| body height | human | [28,29,30] | |
| LIN28B paralog | finger length ratio | human | [31] |
| adiposity | human | [32] | |
| placenta development | human | [43,44] | |
| hematopoietic maturation | mouse | [13]; | |
| age at menarche | human | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krsnik, D.; Marić, T.; Bulić-Jakuš, F.; Sinčić, N.; Bojanac, A.K. LIN28 Family in Testis: Control of Cell Renewal, Maturation, Fertility and Aging. Int. J. Mol. Sci. 2022, 23, 7245. https://doi.org/10.3390/ijms23137245
Krsnik D, Marić T, Bulić-Jakuš F, Sinčić N, Bojanac AK. LIN28 Family in Testis: Control of Cell Renewal, Maturation, Fertility and Aging. International Journal of Molecular Sciences. 2022; 23(13):7245. https://doi.org/10.3390/ijms23137245
Chicago/Turabian StyleKrsnik, Dajana, Tihana Marić, Floriana Bulić-Jakuš, Nino Sinčić, and Ana Katušić Bojanac. 2022. "LIN28 Family in Testis: Control of Cell Renewal, Maturation, Fertility and Aging" International Journal of Molecular Sciences 23, no. 13: 7245. https://doi.org/10.3390/ijms23137245
APA StyleKrsnik, D., Marić, T., Bulić-Jakuš, F., Sinčić, N., & Bojanac, A. K. (2022). LIN28 Family in Testis: Control of Cell Renewal, Maturation, Fertility and Aging. International Journal of Molecular Sciences, 23(13), 7245. https://doi.org/10.3390/ijms23137245