
Variation of Structural and Dynamical Flexibility of Myelin Basic Protein in Response to Guanidinium Chloride

 , , , ,
, , , ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
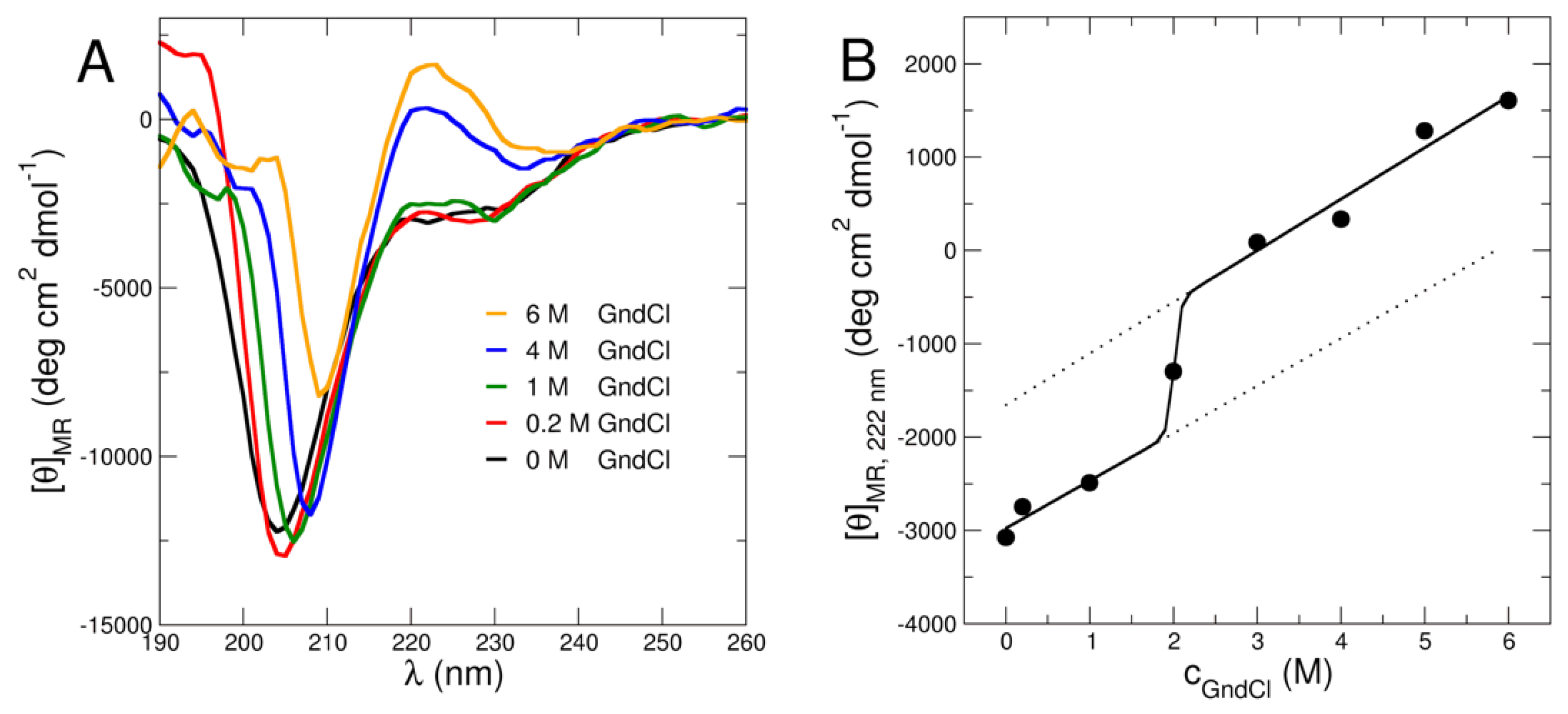
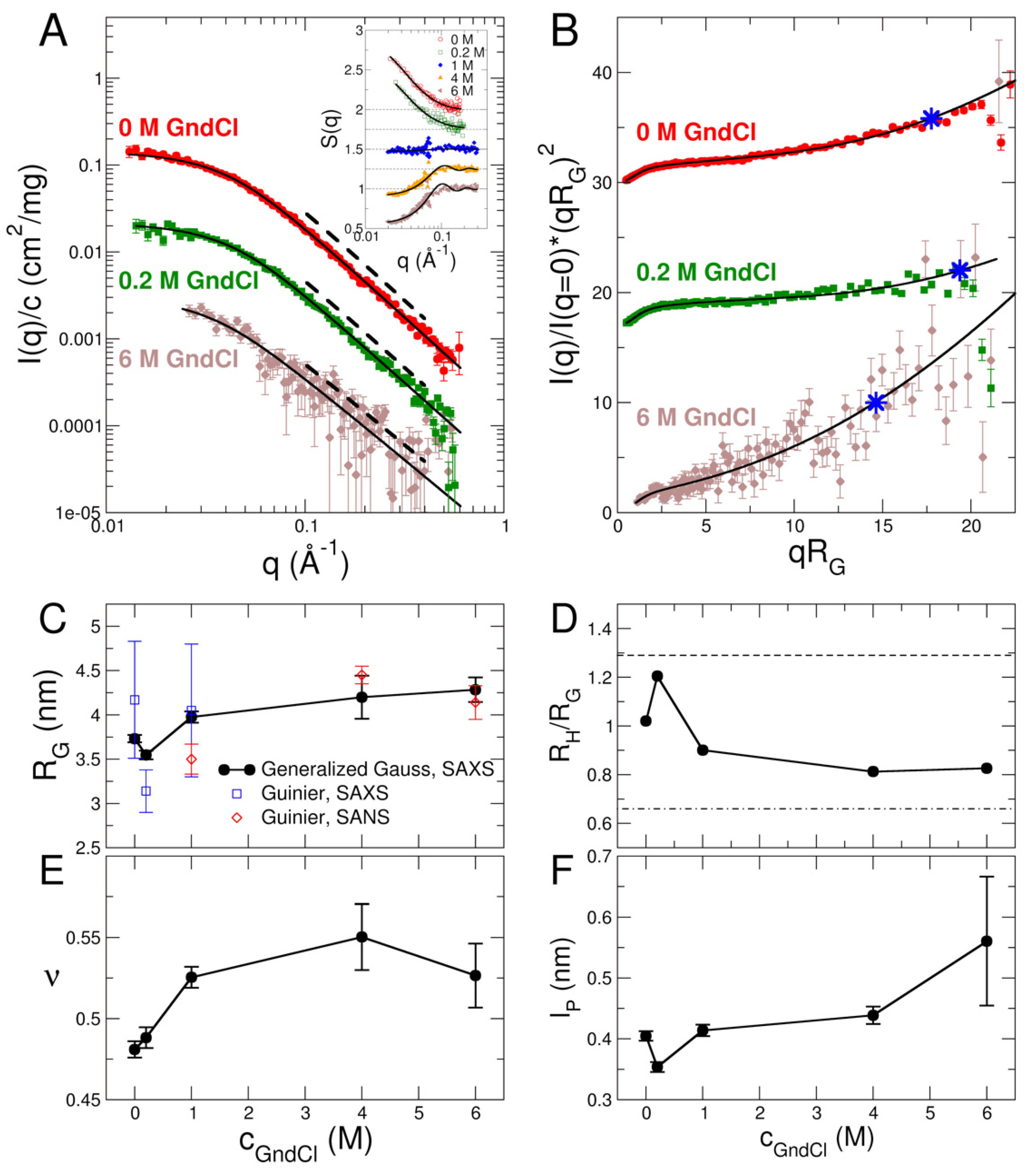
2.1. Structural Investigation Using CD and SAXS/SANS Experiments
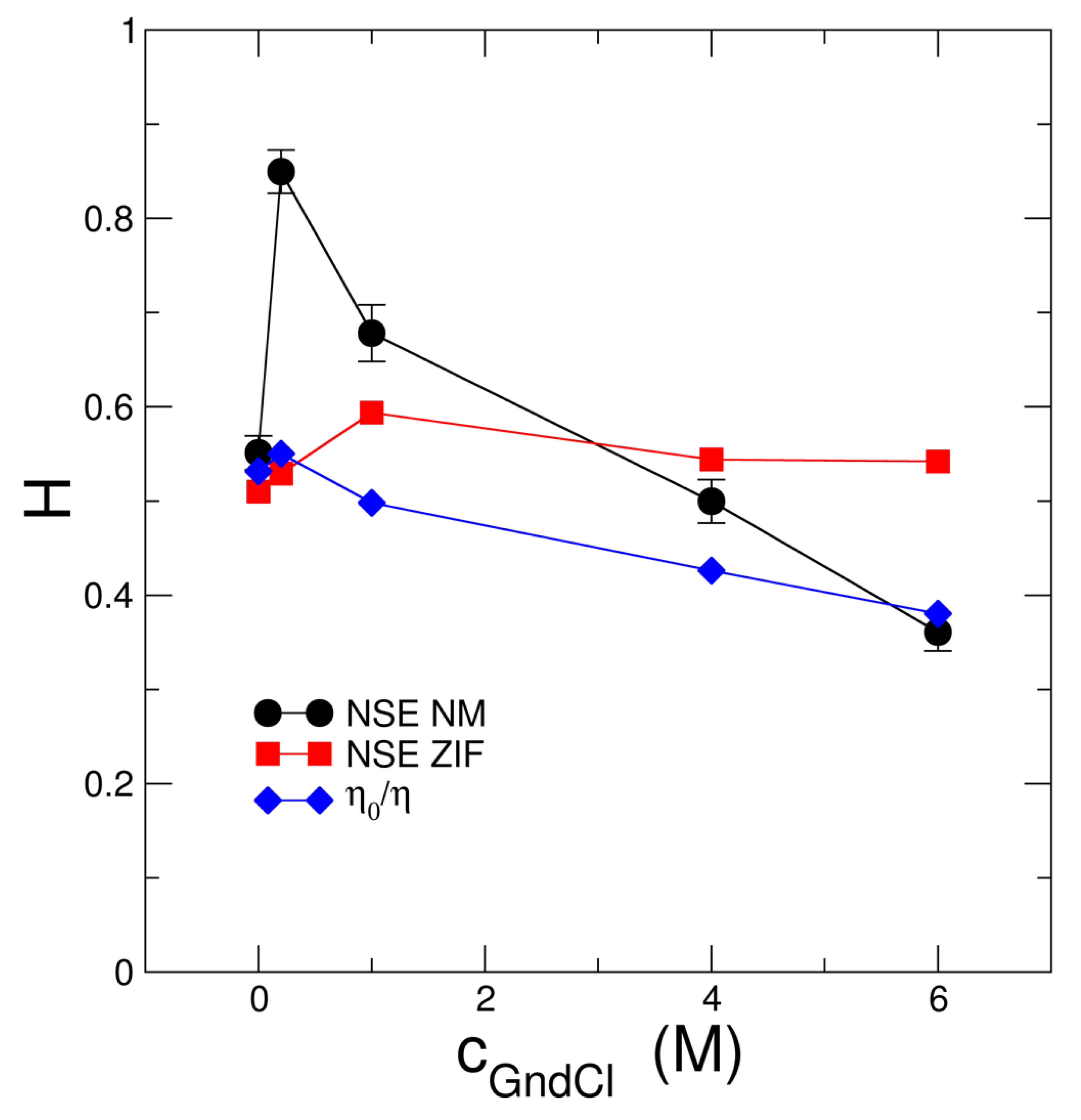
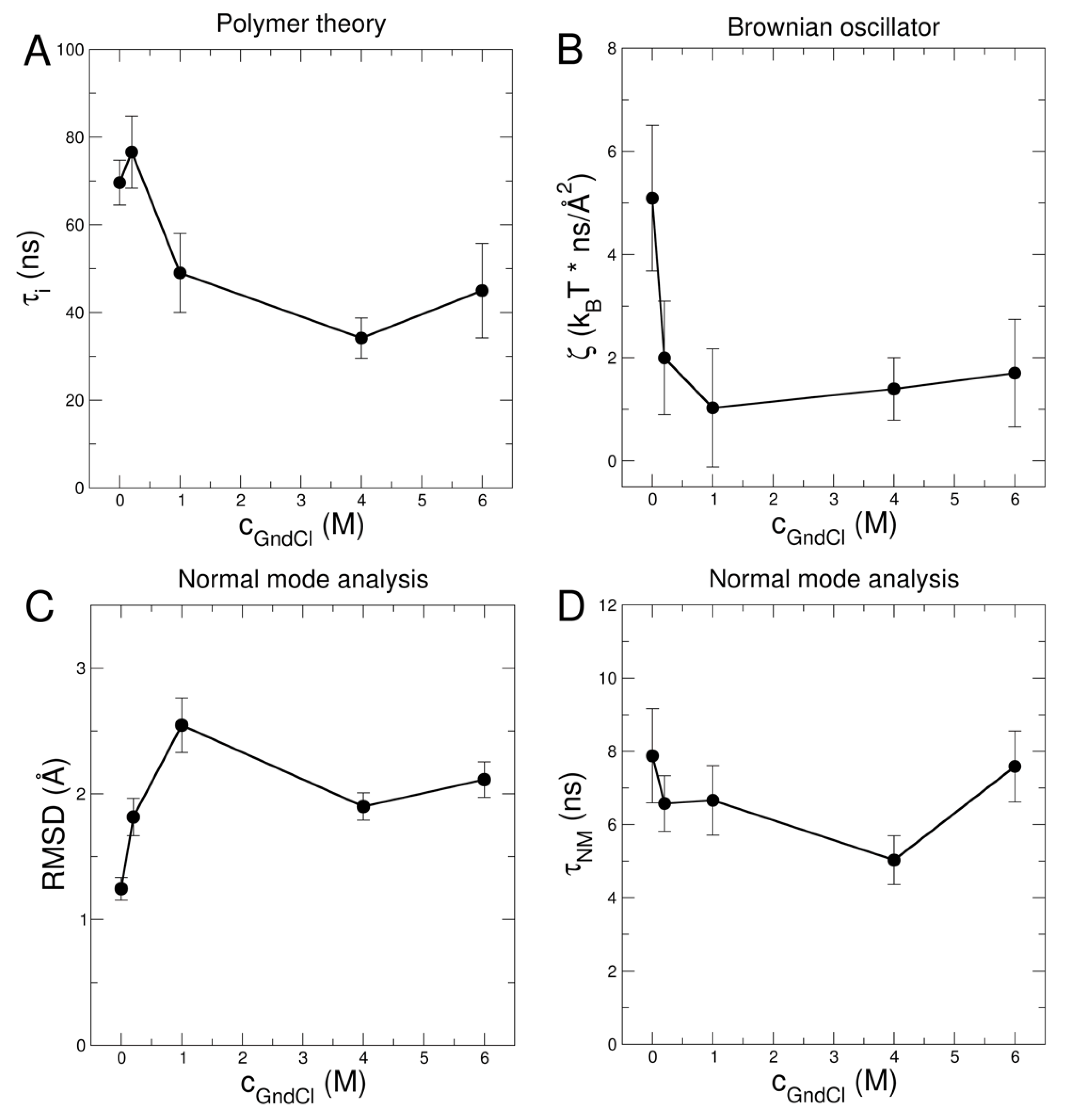
2.2. Investigation of Protein Dynamics Using NSE
3. Conclusions
4. Materials & Methods
4.1. Sample Preparation
4.2. Experimental Methods and Data Analysis
4.2.1. CD Experiments
4.2.2. DLS and Viscosity Measurements
4.2.3. SAXS/SANS Experiments and Data Analysis
4.2.4. NSE Spectroscopy Experiments
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oldfield, C.J.; Dunker, A.K. Intrinsically Disordered Proteins and Intrinsically Disordered Protein Regions. Annu. Rev. Biochem. 2014, 83, 553–584. [Google Scholar] [CrossRef] [PubMed]
- Dyson, H.J.; Wright, P.E. Intrinsically Unstructured Proteins and Their Functions. Nat. Rev. Mol. Cell Biol. 2005, 6, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.L.; Uversky, V.N. Intrinsic Disorder and Posttranslational Modifications: The Darker Side of the Biological Dark Matter. Front. Genet. 2018, 9, 158. [Google Scholar] [CrossRef]
- Uversky, V.N. Intrinsically Disordered Proteins and Their “Mysterious” (Meta)Physics. Front. Phys. 2019, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Burger, V.; Gurry, T.; Stultz, C. Intrinsically Disordered Proteins: Where Computation Meets Experiment. Polymers 2014, 6, 2684–2719. [Google Scholar] [CrossRef] [Green Version]
- Soranno, A.; Buchli, B.; Nettels, D.; Cheng, R.R.; Müller-Späth, S.; Pfeil, S.H.; Hoffmann, A.; Lipman, E.A.; Makarov, D.E.; Schuler, B. Quantifying Internal Friction in Unfolded and Intrinsically Disordered Proteins with Single-Molecule Spectroscopy. Proc. Natl. Acad. Sci. USA 2012, 109, 17800–17806. [Google Scholar] [CrossRef] [Green Version]
- Echeverria, I.; Makarov, D.E.; Papoian, G.A. Concerted Dihedral Rotations Give Rise to Internal Friction in Unfolded Proteins. J. Am. Chem. Soc. 2014, 136, 8708–8713. [Google Scholar] [CrossRef]
- Das, D.; Arora, L.; Mukhopadhyay, S. Short-Range Backbone Dihedral Rotations Modulate Internal Friction in Intrinsically Disordered Proteins. J. Am. Chem. Soc. 2022, 144, 1739–1747. [Google Scholar] [CrossRef]
- Das, D.; Arora, L.; Mukhopadhyay, S. Fluorescence Depolarization Kinetics Captures Short-Range Backbone Dihedral Rotations and Long-Range Correlated Dynamics of an Intrinsically Disordered Protein. J. Phys. Chem. B 2021, 125, 9708–9718. [Google Scholar] [CrossRef]
- Müller-Späth, S.; Soranno, A.; Hirschfeld, V.; Hofmann, H.; Rüegger, S.; Reymond, L.; Nettels, D.; Schuler, B. Charge Interactions Can Dominate the Dimensions of Intrinsically Disordered Proteins. Proc. Natl. Acad. Sci. USA 2010, 107, 14609–14614. [Google Scholar] [CrossRef] [Green Version]
- Shou, K.; Bremer, A.; Rindfleisch, T.; Knox-Brown, P.; Hirai, M.; Rekas, A.; Garvey, C.J.; Hincha, D.K.; Stadler, A.M.; Thalhammer, A. Conformational Selection of the Intrinsically Disordered Plant Stress Protein COR15A in Response to Solution Osmolarity-an X-ray and Light Scattering Study. Phys. Chem. Chem. Phys. 2019, 21, 18727–18740. [Google Scholar] [CrossRef] [PubMed]
- Theillet, F.X.; Binolfi, A.; Frembgen-Kesner, T.; Hingorani, K.; Sarkar, M.; Kyne, C.; Li, C.; Crowley, P.B.; Gierasch, L.; Pielak, G.J.; et al. Physicochemical Properties of Cells and Their Effects on Intrinsically Disordered Proteins (IDPs). Chem. Rev. 2014, 114, 6661–6714. [Google Scholar] [CrossRef] [PubMed]
- Träger, J.; Widder, K.; Kerth, A.; Harauz, G.; Hinderberger, D. Effect of Cholesterol and Myelin Basic Protein (MBP) Content on Lipid Monolayers Mimicking the Cytoplasmic Membrane of Myelin. Cells 2020, 9, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raasakka, A.; Kursula, P. Flexible Players within the Sheaths: The Intrinsically Disordered Proteins of Myelin in Health and Disease. Cells 2020, 9, 470. [Google Scholar] [CrossRef] [Green Version]
- Libich, D.S.; Harauz, G. Backbone Dynamics of the 18.5 KDa Isoform of Myelin Basic Protein Reveals Transient α-Helices and a Calmodulin-Binding Site. Biophys. J. 2008, 94, 4847–4866. [Google Scholar] [CrossRef] [Green Version]
- Vassall, K.A.; Bessonov, K.; de Avila, M.; Polverini, E.; Harauz, G. The Effects of Threonine Phosphorylation on the Stability and Dynamics of the Central Molecular Switch Region of 18.5-KDa Myelin Basic Protein. PLoS ONE 2013, 8, e0131653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebes, L.F.; Zand, R.; Phillips, W.D. Solution Behavior, Circular Dichroism and 220 MHz PMR Studies of the Bovine Myelin Basic Protein. Biochim. Et Biophys. Acta (BBA) Protein. Struct. 1975, 405, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Harauz, G.; Ishiyama, N.; Hill, C.M.; Bates, I.R.; Libich, D.S.; Fares, C. Myelin Basic Protein-Diverse Conformational States of an Intrinsically Unstructured Protein and Its Roles in Myelin Assembly and Multiple Sclerosis. Micron 2004, 35, 503–542. [Google Scholar] [CrossRef]
- Wang, C.; Neugebauer, U.; Bürck, J.; Myllykoski, M.; Baumgärtel, P.; Popp, J.; Kursula, P. Charge Isomers of Myelin Basic Protein: Structure and Interactions with Membranes, Nucleotide Analogues, and Calmodulin. PLoS ONE 2011, 6, e19915. [Google Scholar] [CrossRef]
- Raasakka, A.; Ruskamo, S.; Kowal, J.; Barker, R.; Baumann, A.; Martel, A.; Tuusa, J.; Myllykoski, M.; Bürck, J.; Ulrich, A.S.; et al. Membrane Association Landscape of Myelin Basic Protein Portrays Formation of the Myelin Major Dense Line. Sci. Rep. 2017, 7, 4974. [Google Scholar] [CrossRef]
- Stingaciu, L.R.; Biehl, R.; Changwoo, D.; Richter, D.; Stadler, A.M. Reduced Internal Friction by Osmolyte Interaction in Intrinsically Disordered Myelin Basic Protein. J. Phys. Chem. Lett. 2020, 11, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Stadler, A.M.; Stingaciu, L.; Radulescu, A.; Holderer, O.; Monkenbusch, M.; Biehl, R.; Richter, D. Internal Nanosecond Dynamics in the Intrinsically Disordered Myelin Basic Protein. J. Am. Chem. Soc. 2014, 136, 6987–6994. [Google Scholar] [CrossRef] [PubMed]
- Hassani, A.N.; Haris, L.; Appel, M.; Seydel, T.; Stadler, A.M.; Kneller, G.R. Multiscale Relaxation Dynamics and Diffusion of Myelin Basic Protein in Solution Studied by Quasielastic Neutron Scattering. J. Chem. Phys. 2022, 156, 025102. [Google Scholar] [CrossRef] [PubMed]
- Krugmann, B.; Koutsioubas, A.; Haris, L.; Micciulla, S.; Lairez, D.; Radulescu, A.; Förster, S.; Stadler, A.M. Adhesion Process of Biomimetic Myelin Membranes Triggered by Myelin Basic Protein. Front. Chem. 2021, 9, 259. [Google Scholar] [CrossRef]
- Krugmann, B.; Radulescu, A.; Appavou, M.-S.; Koutsioubas, A.; Stingaciu, L.R.; Dulle, M.; Förster, S.; Stadler, A.M. Membrane Stiffness and Myelin Basic Protein Binding Strength as Molecular Origin of Multiple Sclerosis. Sci. Rep. 2020, 10, 16691. [Google Scholar] [CrossRef]
- Fischer, J.; Radulescu, A.; Falus, P.; Richter, D.; Biehl, R. Structure and Dynamics of Ribonuclease A during Thermal Unfolding: The Failure of the Zimm Model. J. Phys. Chem. B 2021, 125, 780–788. [Google Scholar] [CrossRef]
- Farago, B.; Nicholl, I.D.; Wang, S.; Cheng, X.; Callaway, D.J.E.; Bu, Z. Activated Nanoscale Actin-Binding Domain Motion in the Catenin–Cadherin Complex Revealed by Neutron Spin Echo Spectroscopy. Proc. Natl. Acad. Sci. USA 2021, 118, e2025012118. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. Protein Secondary Structure Analyses from Circular Dichroism Spectroscopy: Methods and Reference Databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Miles, A.J.; Ramalli, S.G.; Wallace, B.A. DichroWeb, a Website for Calculating Protein Secondary Structure from Circular Dichroism Spectroscopic Data. Protein. Sci. 2022, 31, 37–46. [Google Scholar] [CrossRef]
- Polverini, E.; Rangaraj, G.; Libich, D.S.; Boggs, J.M.; Harauz, G. Binding of the Proline-Rich Segment of Myelin Basic Protein to SH3 Domains: Spectroscopic, Microarray, and Modeling Studies of Ligand Conformation and Effects of Posttranslational Modifications. Biochemistry 2008, 47, 267–282. [Google Scholar] [CrossRef]
- Adzhubei, A.A.; Sternberg, M.J.E.; Makarov, A.A. Polyproline-II Helix in Proteins: Structure and Function. J. Mol. Biol. 2013, 425, 2100–2132. [Google Scholar] [CrossRef] [PubMed]
- Chemes, L.B.; Alonso, L.G.; Noval, M.G.; de Prat-Gay, G. Circular Dichroism Techniques for the Analysis of Intrinsically Disordered Proteins and Domains. Methods Mol. Biol. 2012, 895, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Tiffany, M.L.; Krimm, S. Extended Conformations of Polypeptides and Proteins in Urea and Guanidine Hydrochloride. Biopolymers 1973, 12, 575–587. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, N.J. Using Circular Dichroism Spectra to Estimate Protein Secondary Structure. Nat. Protoc. 2007, 1, 2876–2890. [Google Scholar] [CrossRef] [PubMed]
- Radulescu, A.; Szekely, N.K.; Appavou, M.-S. KWS-2: Small Angle Scattering Diffractometer. J. Large-Scale Res. Facil. JLSRF 2015, 1, A29. [Google Scholar] [CrossRef] [Green Version]
- Houston, J.E.; Brandl, G.; Drochner, M.; Kemmerling, G.; Engels, R.; Papagiannopoulos, A.; Sarter, M.; Stadler, A.; Radulescu, A. The High-Intensity Option of the SANS Diffractometer KWS-2 at JCNS—Characterization and Performance of the New Multi-Megahertz Detection System. J. Appl. Crystallogr. 2018, 51, 323–336. [Google Scholar] [CrossRef]
- Radulescu, A.; Pipich, V.; Frielinghaus, H.; Appavou, M.S. KWS-2, the High Intensity /Wide Q-Range Small-Angle Neutron Diffractometer for Soft-Matter and Biology at FRM II. Proc. J. Phys. Conf. Ser. 2012, 351, 012026. [Google Scholar] [CrossRef] [Green Version]
- Camilloni, C.; Rocco, A.G.; Eberini, I.; Gianazza, E.; Broglia, R.A.; Tiana, G. Urea and Guanidinium Chloride Denature Protein L in Different Ways in Molecular Dynamics Simulations. Biophys. J. 2008, 94, 4654–4661. [Google Scholar] [CrossRef] [Green Version]
- Heyda, J.; Okur, H.I.; Hladílková, J.; Rembert, K.B.; Hunn, W.; Yang, T.; Dzubiella, J.; Jungwirth, P.; Cremer, P.S. Guanidinium Can Both Cause and Prevent the Hydrophobic Collapse of Biomacromolecules. J. Am. Chem. Soc. 2017, 139, 863–870. [Google Scholar] [CrossRef]
- Mehrnejad, F.; Khadem-Maaref, M.; Ghahremanpour, M.M.; Doustdar, F. Mechanisms of Amphipathic Helical Peptide Denaturation by Guanidinium Chloride and Urea: A Molecular Dynamics Simulation Study. J. Comput. Aided Mol. Des. 2010, 24, 829–841. [Google Scholar] [CrossRef]
- Borgia, A.; Zheng, W.; Buholzer, K.; Borgia, M.B.; Schüler, A.; Hofmann, H.; Soranno, A.; Nettels, D.; Gast, K.; Grishaev, A.; et al. Consistent View of Polypeptide Chain Expansion in Chemical Denaturants from Multiple Experimental Methods. J. Am. Chem. Soc. 2016, 138, 11714–11726. [Google Scholar] [CrossRef] [Green Version]
- Kohn, J.E.; Millett, I.S.; Jacob, J.; Zagrovic, B.; Dillon, T.M.; Cingel, N.; Dothager, R.S.; Seifert, S.; Thiyagarajan, P.; Sosnick, T.R.; et al. Random-Coil Behavior and the Dimensions of Chemically Unfolded Proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 12491–12496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- le Guillou, J.C.; Zinn-Justin, J. Critical Exponents for the N-Vector Model in Three Dimensions from Field Theory. Phys. Rev. Lett. 1977, 39, 95. [Google Scholar] [CrossRef]
- Pedersen, J.S.; Schurtenberger, P. Scattering Functions of Semiflexible Polymers with and without Excluded Volume Effects. Macromolecules 1996, 29, 7602–7612. [Google Scholar] [CrossRef]
- Receveur-Brechot, V.; Durand, D. How Random Are Intrinsically Disordered Proteins? A Small Angle Scattering Perspective. Curr. Protein Pept. Sci. 2012, 13, 55–75. [Google Scholar] [CrossRef] [PubMed]
- Kratky, O. Das Studium Gelöster Fadenmoleküle Mittels Der Röntgenkleinwinkelmethode. Colloid Polym. Sci. 1962, 182, 7–24. [Google Scholar] [CrossRef]
- Doi, M.; Edwards, S.F. The Theory of Polymer Dynamics; Birman, J., Edwards, S.F., Llewellyn Smith, C.H., Rees, M., Eds.; Clarendon Press: Oxford, UK, 1986. [Google Scholar]
- Stadler, A.M.; Koza, M.M.; Fitter, J. Determination of Conformational Entropy of Fully and Partially Folded Conformations of Holo- and Apomyoglobin. J. Phys. Chem. B 2015, 119, 72–82. [Google Scholar] [CrossRef]
- Tria, G.; Mertens, H.D.T.; Kachala, M.; Svergun, D.I. Advanced Ensemble Modelling of Flexible Macromolecules Using X-ray Solution Scattering. IUCrJ 2015, 2, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Biehl, R.; Monkenbusch, M.; Richter, D. Exploring Internal Protein Dynamics by Neutron Spin Echo Spectroscopy. Soft Matter 2011, 7, 1299–1307. [Google Scholar] [CrossRef] [Green Version]
- Biehl, R.; Richter, D. Slow Internal Protein Dynamics in Solution. J. Phys. Condensed Matter 2014, 26, 503103. [Google Scholar] [CrossRef]
- Farago, B.; Falus, P.; Hoffmann, I.; Gradzielski, M.; Thomas, F.; Gomez, C. The IN15 Upgrade. Neutron News 2015, 26, 15–17. [Google Scholar] [CrossRef]
- Pasini, S.; Holderer, O.; Kozielewski, T.; Richter, D.; Monkenbusch, M. J-NSE-Phoenix, a Neutron Spin-Echo Spectrometer with Optimized Superconducting Precession Coils at the MLZ in Garching. Rev. Sci. Instrum. 2019, 90, 043107. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.R.; Hawk, A.T.; Makarov, D.E. Exploring the Role of Internal Friction in the Dynamics of Unfolded Proteins Using Simple Polymer Models. J. Chem. Phys. 2013, 138, 074112. [Google Scholar] [CrossRef] [PubMed]
- Khatri, B.S.; McLeish, T.C.B. Rouse Model with Internal Friction: A Coarse Grained Framework for Single Biopolymer Dynamics. Macromolecules 2007, 40, 6770–6777. [Google Scholar] [CrossRef]
- Ameseder, F.; Radulescu, A.; Holderer, O.; Falus, P.; Richter, D.; Stadler, A.M. Relevance of Internal Friction and Structural Constraints for the Dynamics of Denatured Bovine Serum Albumin. J. Phys. Chem. Lett. 2018, 9, 2469–2473. [Google Scholar] [CrossRef]
- Monkenbusch, M.; Allgaier, J.; Richter, D.; Stellbrink, J.; Fetters, L.J.; Greiner, A. Nonflexible Coils in Solution: A Neutron Spin-Echo Investigation of Alkyl-Substituted Polynorbornenes in Tetrahydrofuran. Macromolecules 2006, 39, 9473–9479. [Google Scholar] [CrossRef]
- Arbe, A.; Pomposo, J.A.; Moreno, A.J.; LoVerso, F.; González-Burgos, M.; Asenjo-Sanz, I.; Iturrospe, A.; Radulescu, A.; Ivanova, O.; Colmenero, J. Structure and Dynamics of Single-Chain Nano-Particles in Solution. Polymer 2016, 105, 532–544. [Google Scholar] [CrossRef]
- González-Burgos, M.; Asenjo-Sanz, I.; Pomposo, J.A.; Radulescu, A.; Ivanova, O.; Pasini, S.; Arbe, A.; Colmenero, J. Structure and Dynamics of Irreversible Single-Chain Nanoparticles in Dilute Solution. A Neutron Scattering Investigation. Macromolecules 2020, 53, 8068–8082. [Google Scholar] [CrossRef]
- Banchio, A.J.; Heinen, M.; Holmqvist, P.; Nägele, G. Short- and Long-Time Diffusion and Dynamic Scaling in Suspensions of Charged Colloidal Particles. J. Chem. Phys. 2018, 148, 134902. [Google Scholar] [CrossRef] [Green Version]
- Bucciarelli, S.; Myung, J.S.; Farago, B.; Das, S.; Vliegenthart, G.A.; Holderer, O.; Winkler, R.G.; Schurtenberger, P.; Gompper, G.; Stradner, A. Dramatic Influence of Patchy Attractions on Short-Time Protein Diffusion under Crowded Conditions. Sci. Adv. 2016, 2, e1601432. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Kang, W.; Wang, J.; Wang, W. Temperature Dependence of Internal Friction of Peptides. J. Phys. Chem. B 2021, 125, 2821–2832. [Google Scholar] [CrossRef] [PubMed]
- García De La Torre, J.; Huertas, M.L.; Carrasco, B. Calculation of Hydrodynamic Properties of Globular Proteins from Their Atomic-Level Structure. Biophys. J. 2000, 78, 719–730. [Google Scholar] [CrossRef] [Green Version]
- Atilgan, A.R.; Durell, S.R.; Jernigan, R.L.; Demirel, M.C.; Keskin, O.; Bahar, I. Anisotropy of Fluctuation Dynamics of Proteins with an Elastic Network Model. Biophys. J. 2001, 80, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Balacescu, L.; Schrader, T.E.; Radulescu, A.; Zolnierczuk, P.; Holderer, O.; Pasini, S.; Fitter, J.; Stadler, A.M. Transition between Protein-like and Polymer-like Dynamic Behavior: Internal Friction in Unfolded Apomyoglobin Depends on Denaturing Conditions. Sci. Rep. 2020, 10, 1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammouda, B. Small-Angle Scattering From Branched Polymers. Macromol. Theory Simul. 2012, 21, 372–381. [Google Scholar] [CrossRef]
- Chen, W.R.; Butler, P.D.; Magid, L.J. Incorporating Intermicellar Interactions in the Fitting of SANS Data from Cationic Wormlike Micelles. Langmuir 2006, 22, 6539–6548. [Google Scholar] [CrossRef]
- Available online: http://www.sasview.org/ (accessed on 1 June 2022).
- Percus, J.K.; Yevick, G.J. Analysis of Classical Statistical Mechanics by Means of Collective Coordinates. Phys. Rev. 1958, 110, 1–13. [Google Scholar] [CrossRef]
- Hayter, J.B.; Penfold, J. An Analytic Structure Factor for Macroion Solutions. Mol. Phys. 1981, 42, 109–118. [Google Scholar] [CrossRef]
- Hansen, J.P.; Hayter, J.B. A Rescaled MSA Structure Factor for Dilute Charged Colloidal Dispersions. Mol. Phys. 1982, 46, 651–656. [Google Scholar] [CrossRef]
- Zolnierczuk, P.A.; Holderer, O.; Pasini, S.; Kozielewski, T.; Stingaciud, L.R.; Monkenbuschc, M. Efficient Data Extraction from Neutron Time-of-Flight Spin-Echo Raw Data. J. Appl. Cryst. Lography 2019, 52, 1022–1034. [Google Scholar] [CrossRef]
- Balacescu, L.; Vögl, F.; Staringer, S.; Ossovyi, V.; Brandl, G.; Lumma, N.; Feilbach, H.; Holderer, O.; Pasini, S.; Stadler, A.; et al. In Situ Dynamic Light Scattering Complementing Neutron Spin Echo Measurements on Protein Samples. J. Surf. Investig. 2020, 14, S185–S189. [Google Scholar] [CrossRef]
- Biehl, R. Jscatter, a Program for Evaluation and Analysis of Experimental Data. PLoS ONE 2019, 14, e0218789. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haris, L.; Biehl, R.; Dulle, M.; Radulescu, A.; Holderer, O.; Hoffmann, I.; Stadler, A.M. Variation of Structural and Dynamical Flexibility of Myelin Basic Protein in Response to Guanidinium Chloride. Int. J. Mol. Sci. 2022, 23, 6969. https://doi.org/10.3390/ijms23136969
Haris L, Biehl R, Dulle M, Radulescu A, Holderer O, Hoffmann I, Stadler AM. Variation of Structural and Dynamical Flexibility of Myelin Basic Protein in Response to Guanidinium Chloride. International Journal of Molecular Sciences. 2022; 23(13):6969. https://doi.org/10.3390/ijms23136969
Chicago/Turabian StyleHaris, Luman, Ralf Biehl, Martin Dulle, Aurel Radulescu, Olaf Holderer, Ingo Hoffmann, and Andreas M. Stadler. 2022. "Variation of Structural and Dynamical Flexibility of Myelin Basic Protein in Response to Guanidinium Chloride" International Journal of Molecular Sciences 23, no. 13: 6969. https://doi.org/10.3390/ijms23136969
APA StyleHaris, L., Biehl, R., Dulle, M., Radulescu, A., Holderer, O., Hoffmann, I., & Stadler, A. M. (2022). Variation of Structural and Dynamical Flexibility of Myelin Basic Protein in Response to Guanidinium Chloride. International Journal of Molecular Sciences, 23(13), 6969. https://doi.org/10.3390/ijms23136969