
Copper-Binding Domain Variation in a Novel Murine Lysyl Oxidase Model Produces Structurally Inferior Aortic Elastic Fibers Whose Failure Is Modified by Age, Sex, and Blood Pressure

 ,
,  , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
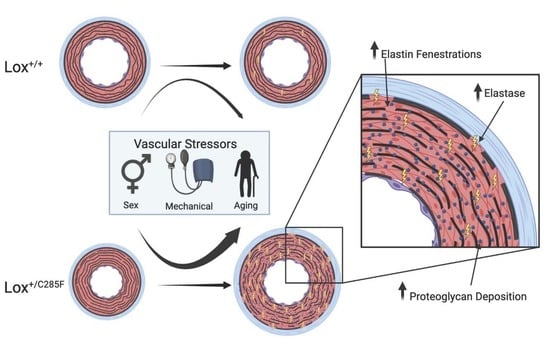
1. Introduction
2. Results
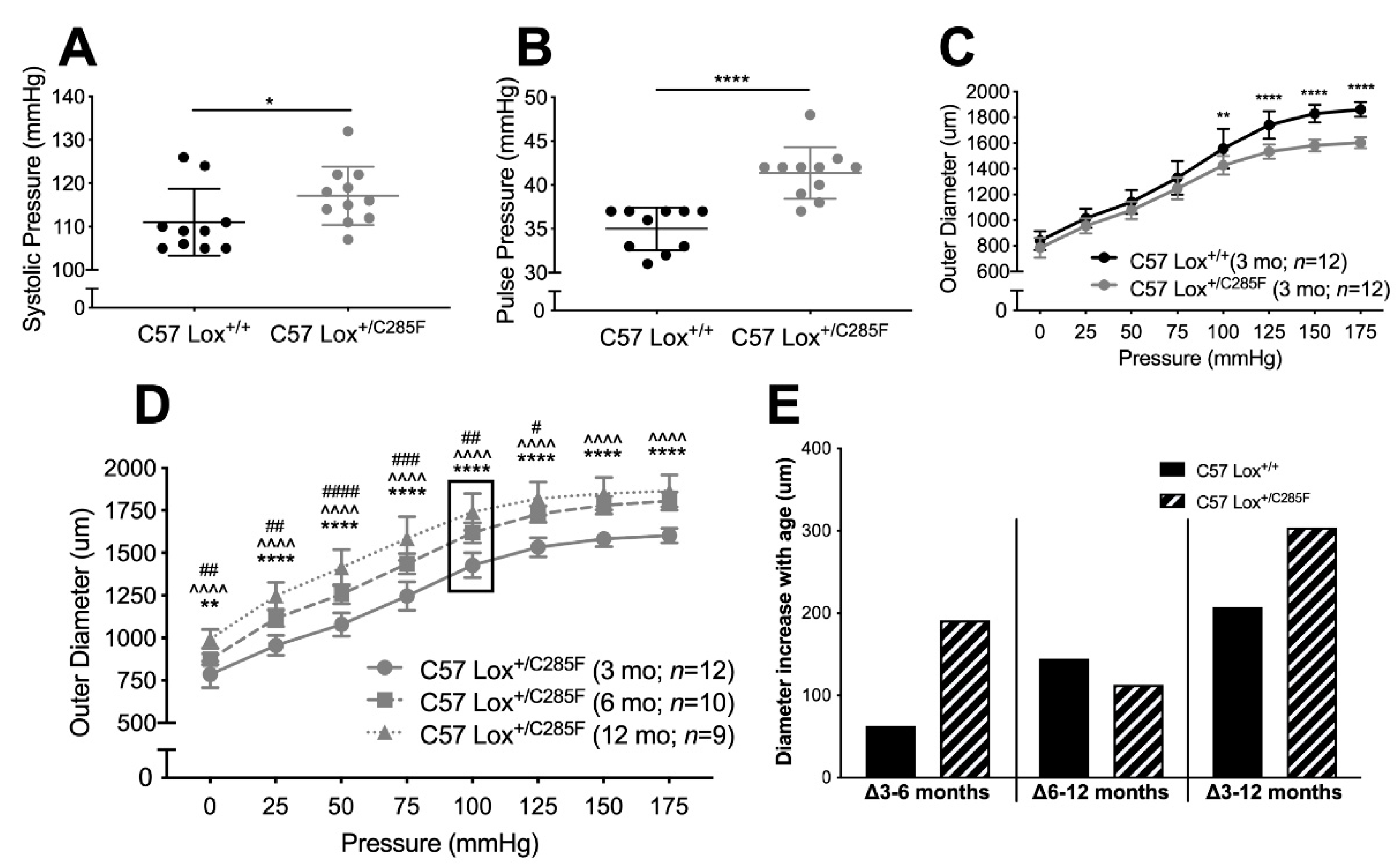
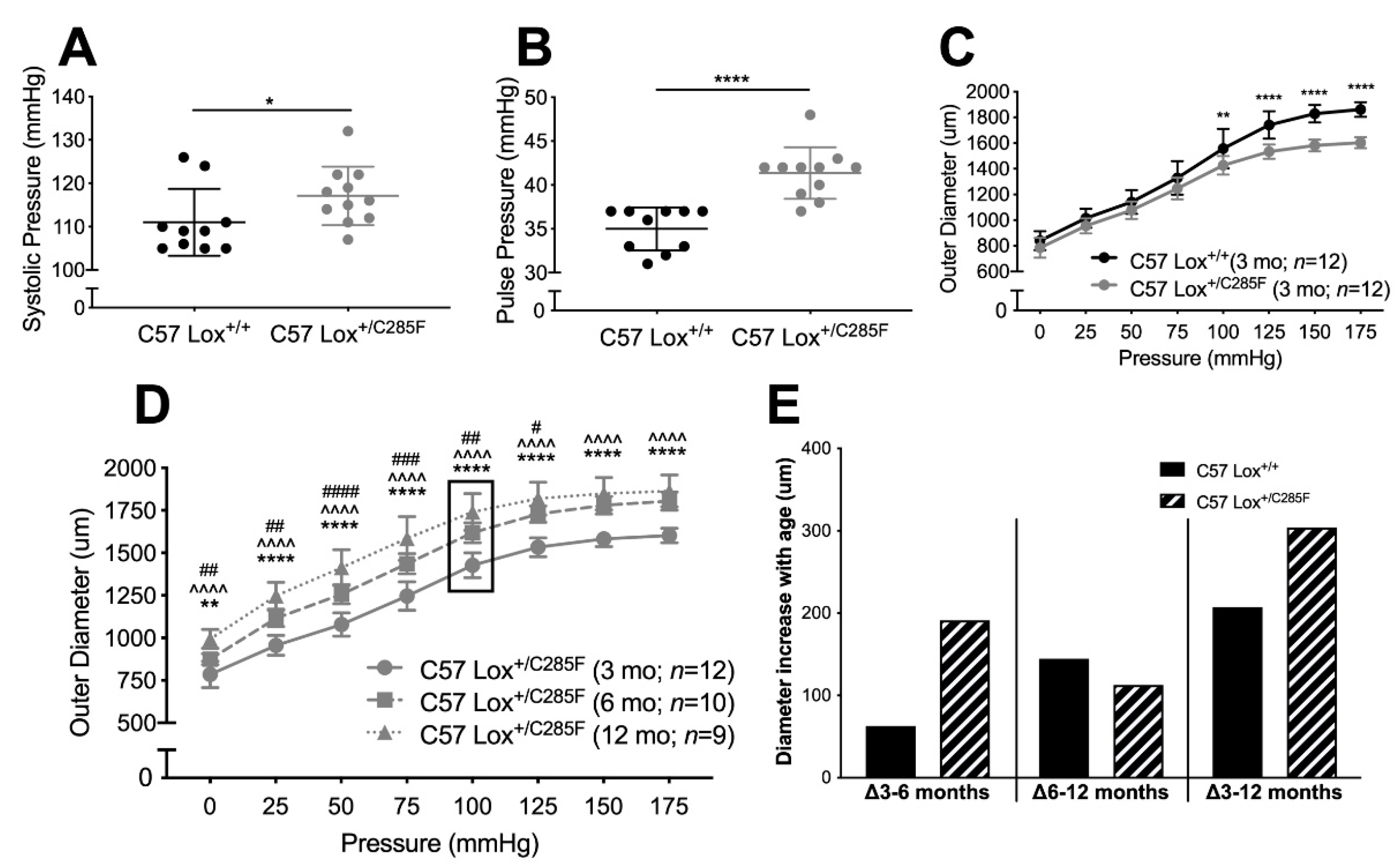
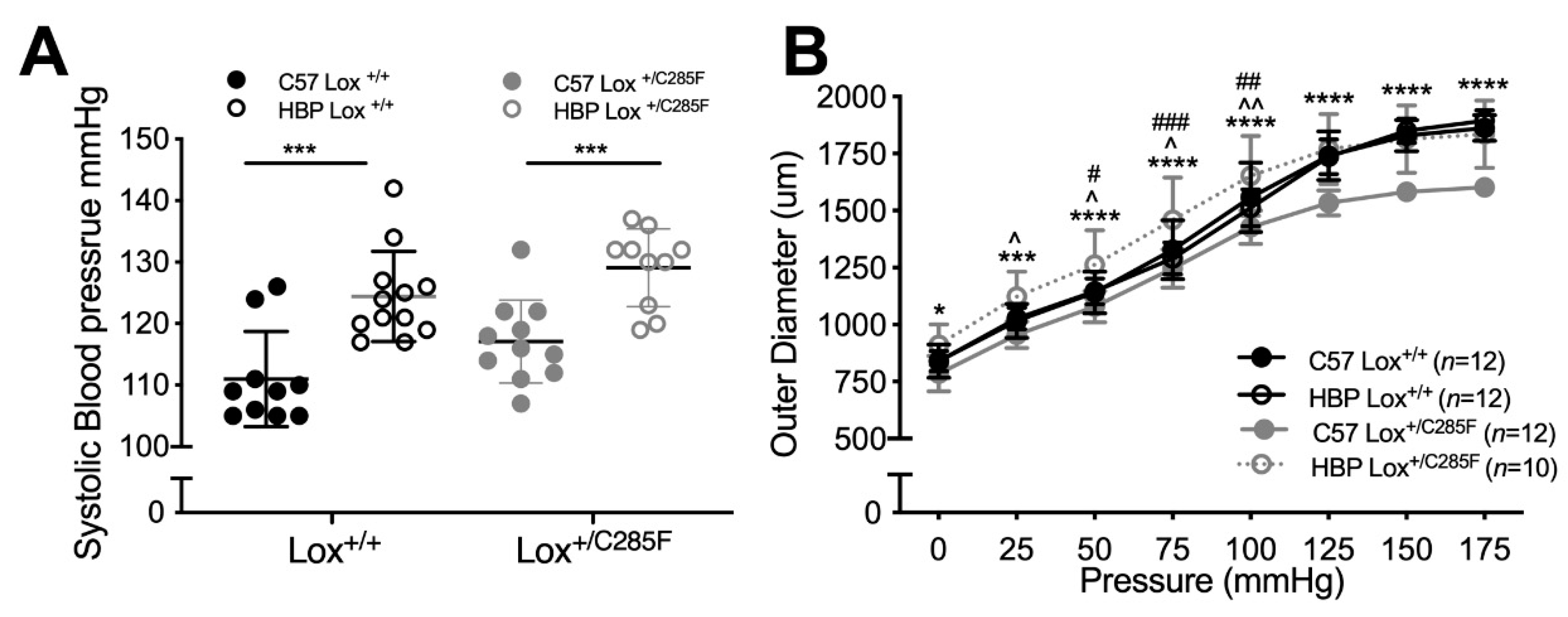
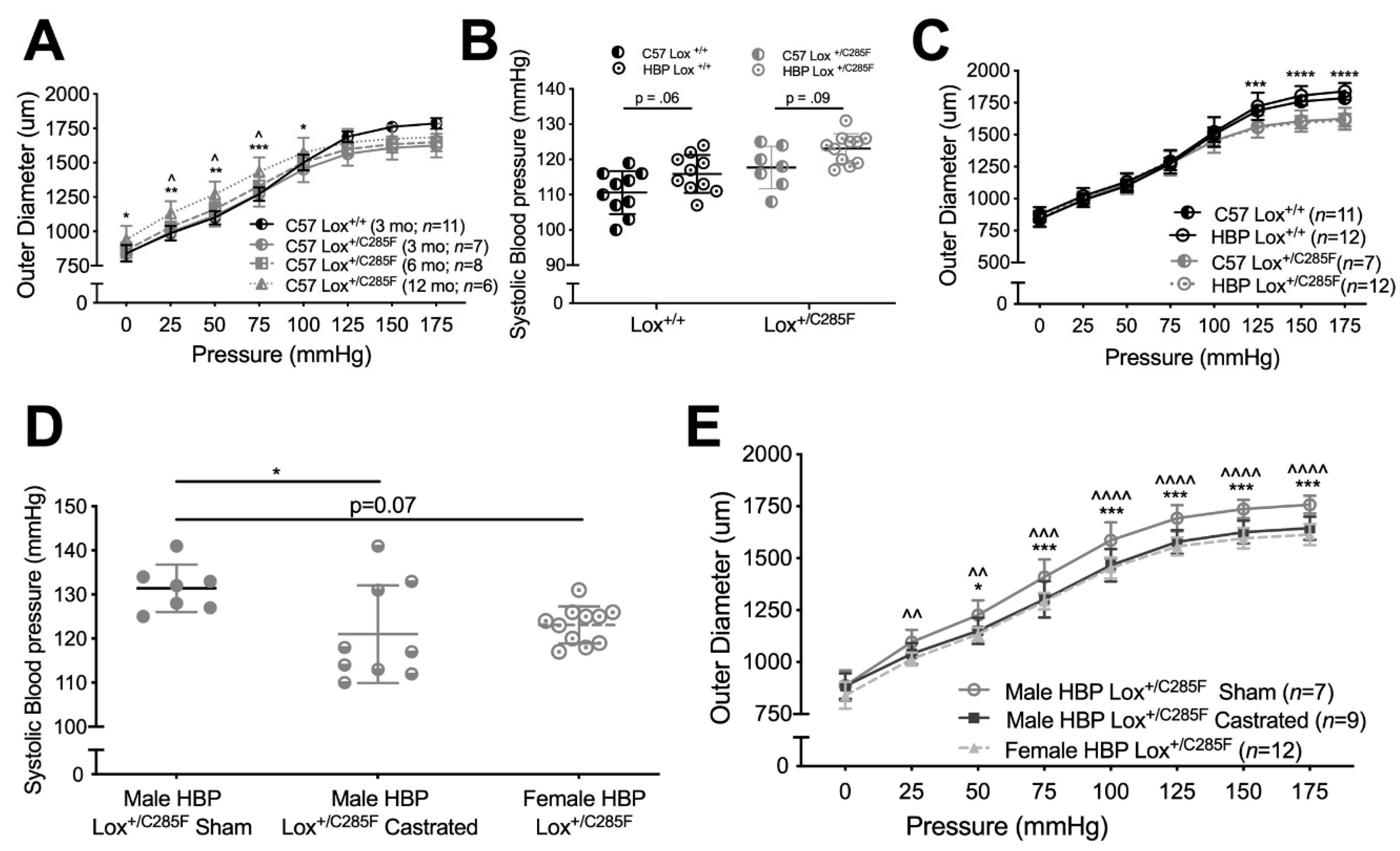
2.1. Cardiovascular Properties of Young Male Lox+/C285F Mice
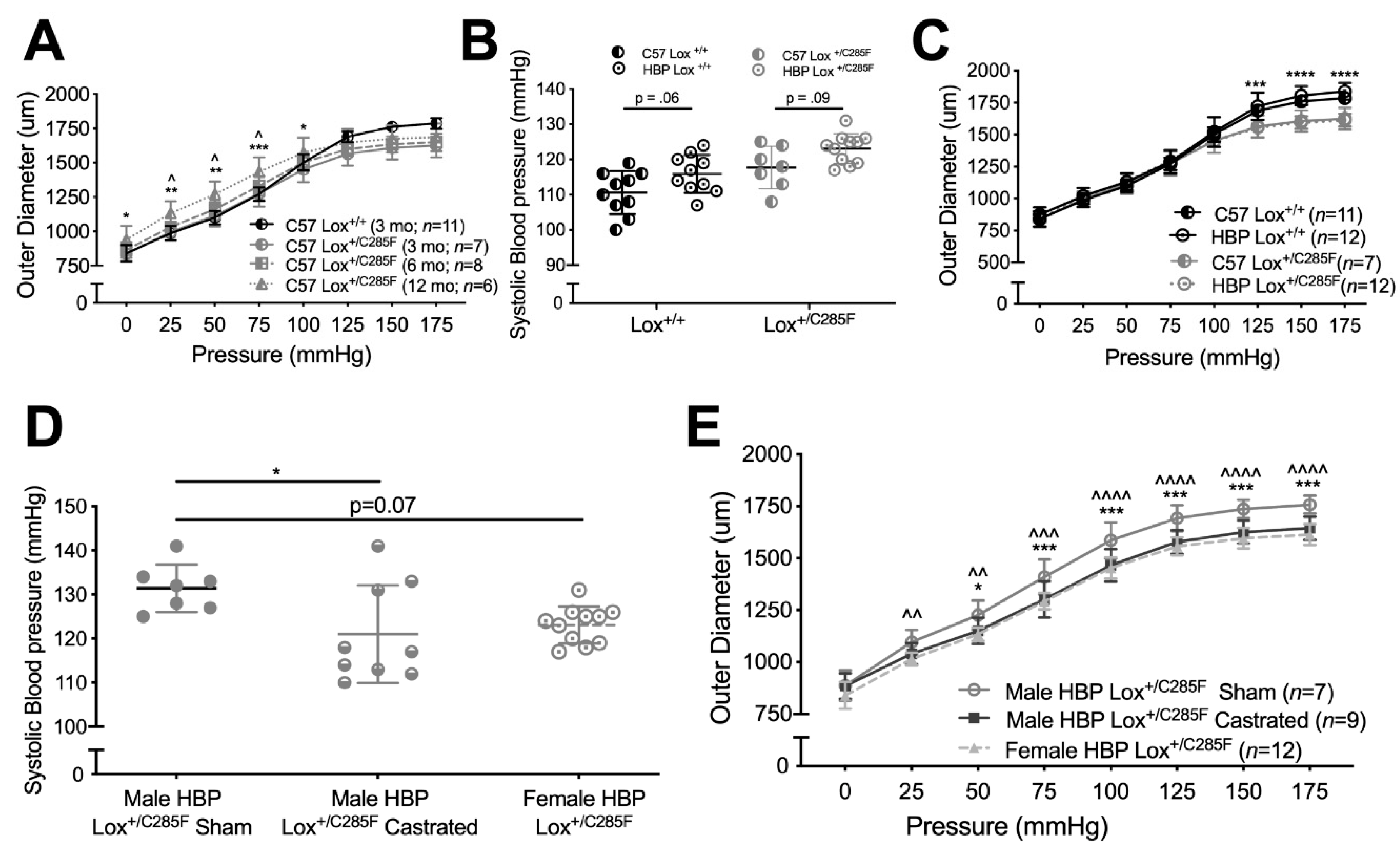
2.2. Lox+/C285F Dilation Rate Is Modified by Sex and Blood Pressure
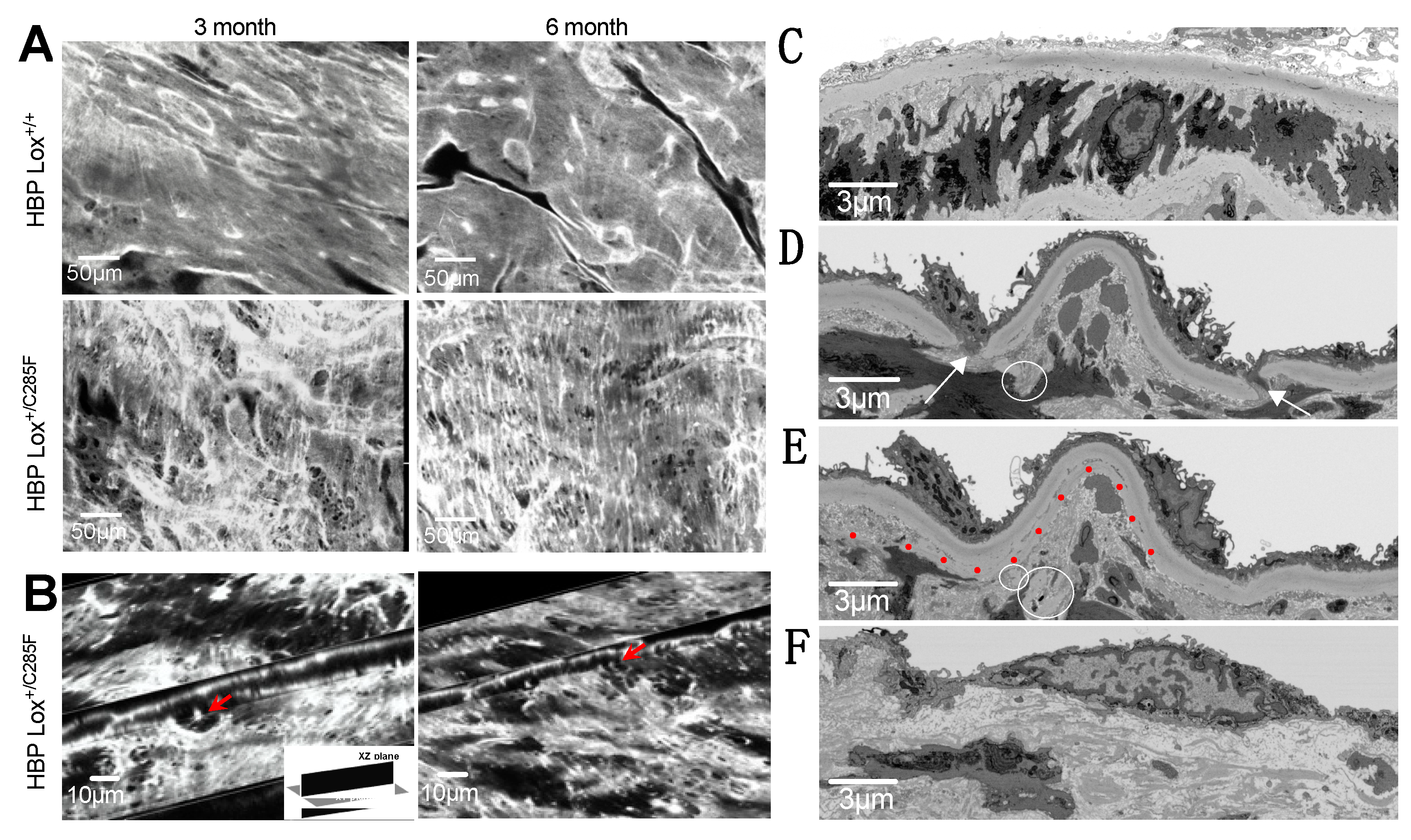
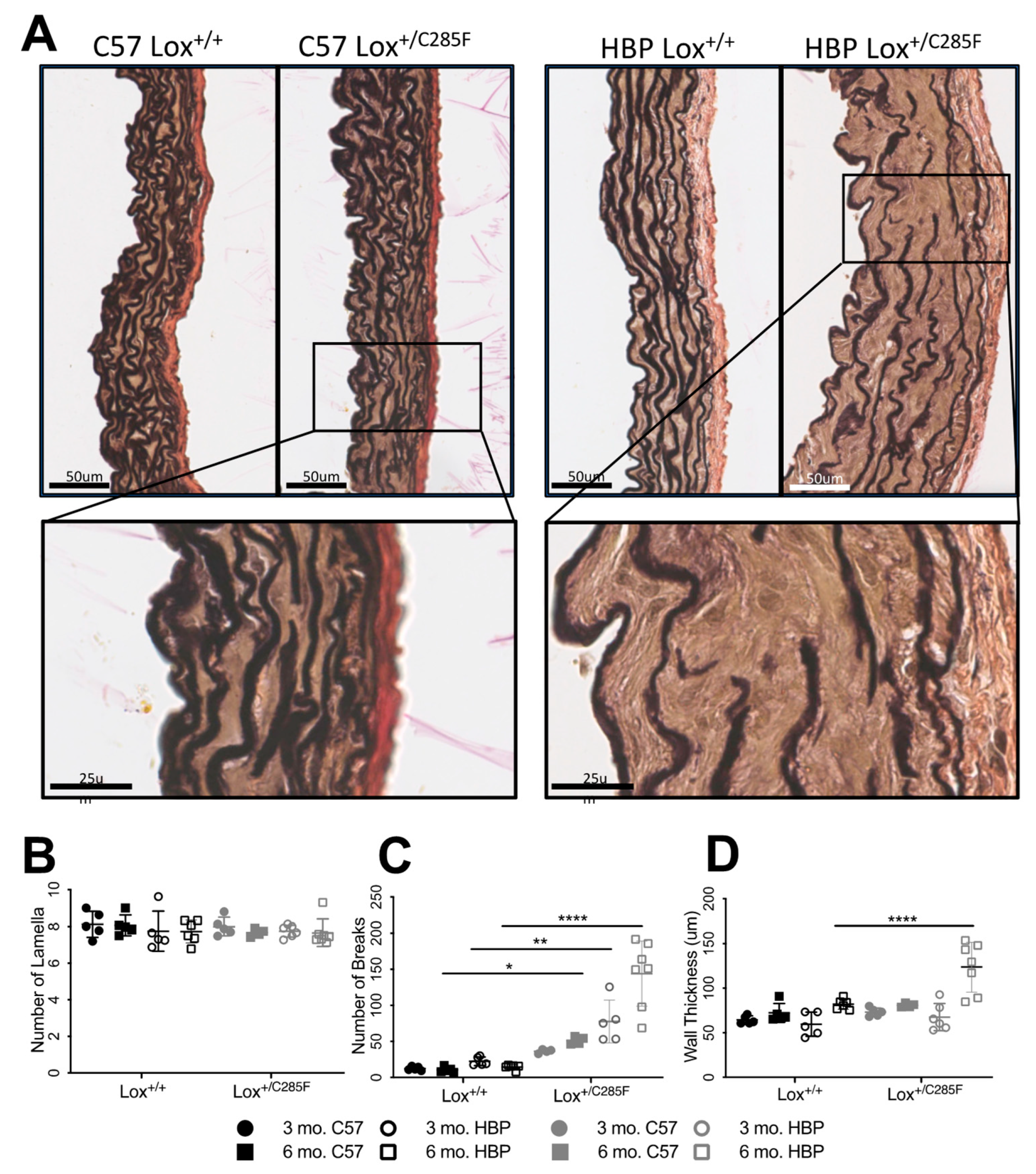
2.3. Increased Number of Breaks and Irregular Elastic Lamellar Sheets in Lox+/C285F Mice
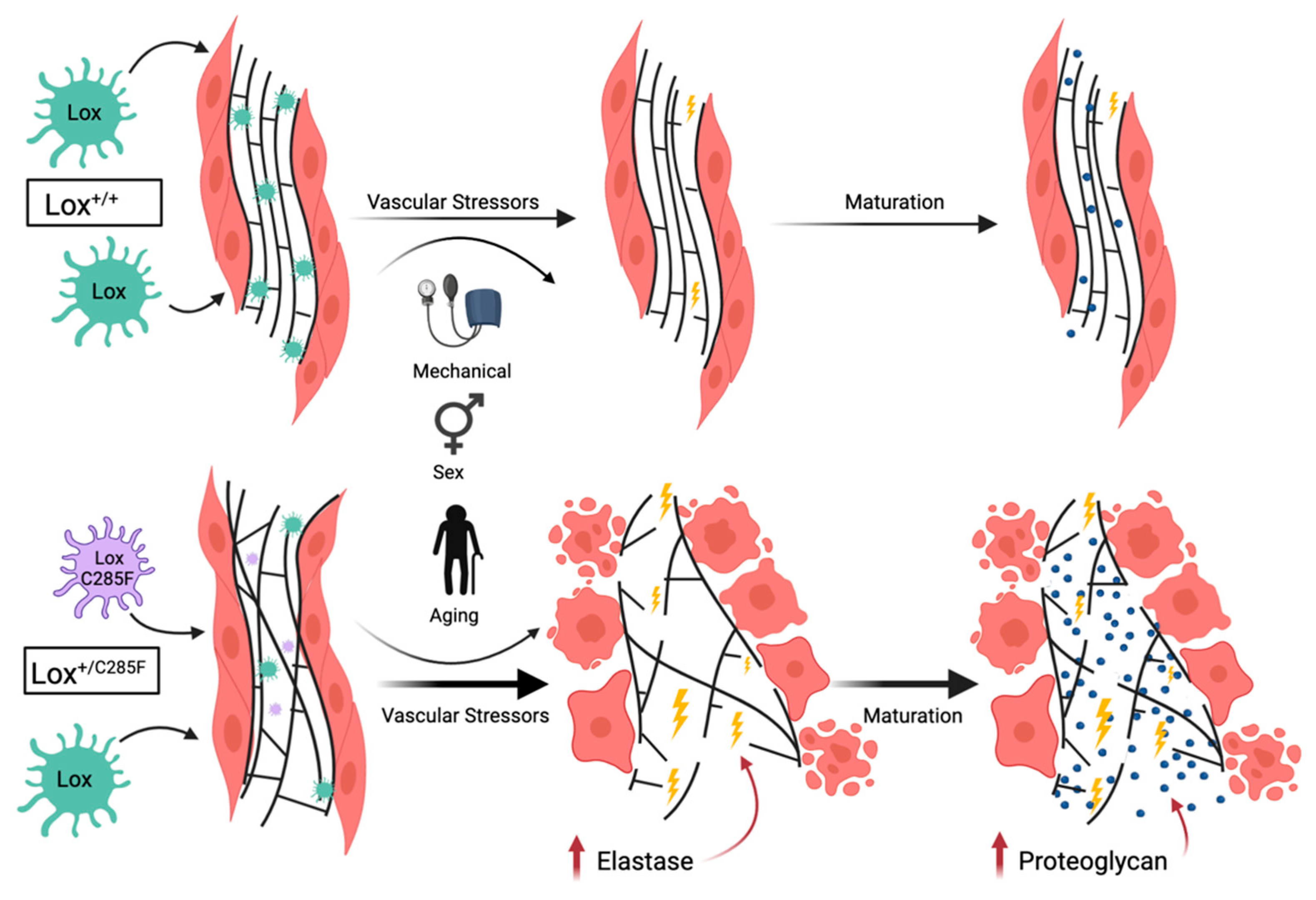
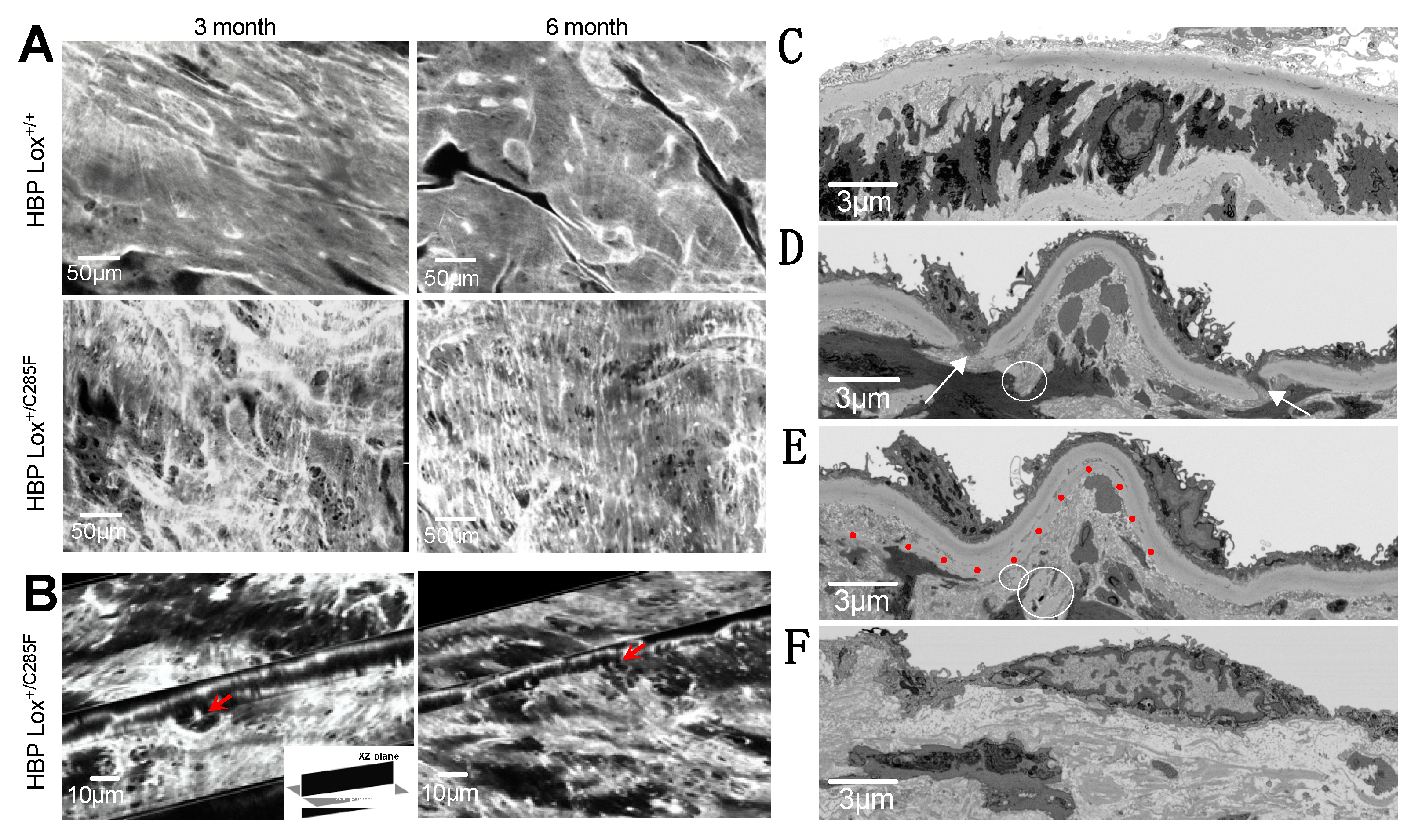
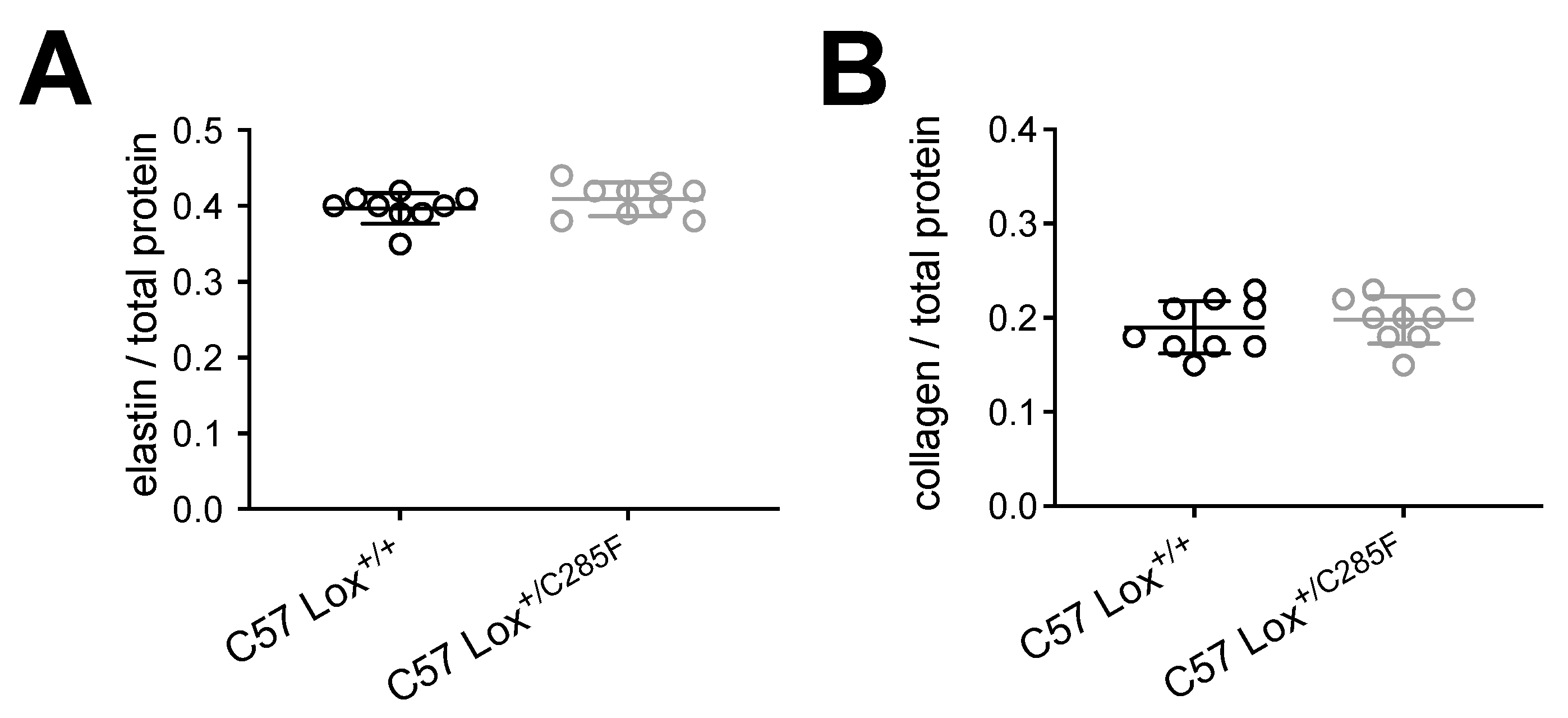
2.4. Mechanism of Lox-Mediated Disease
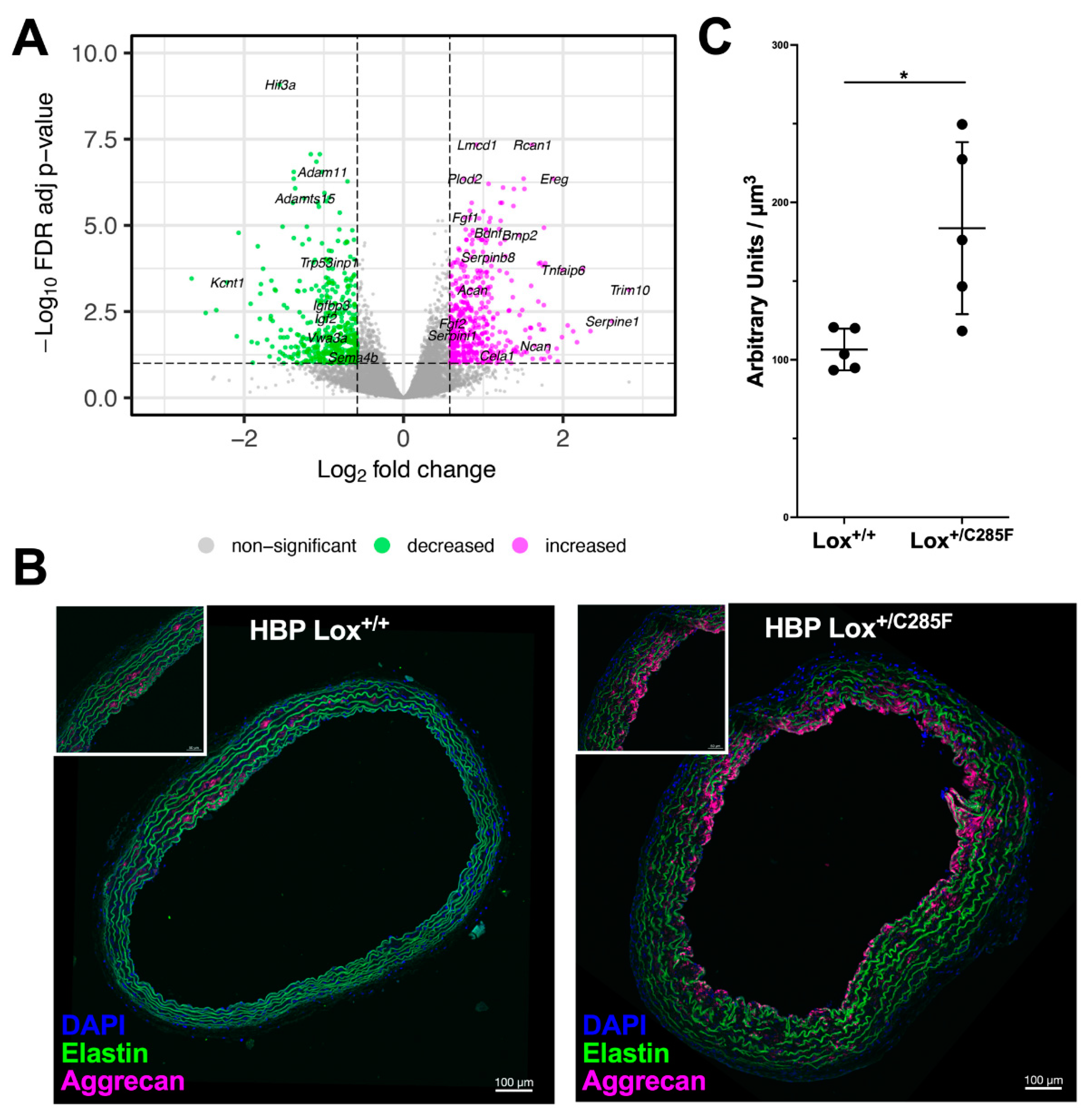
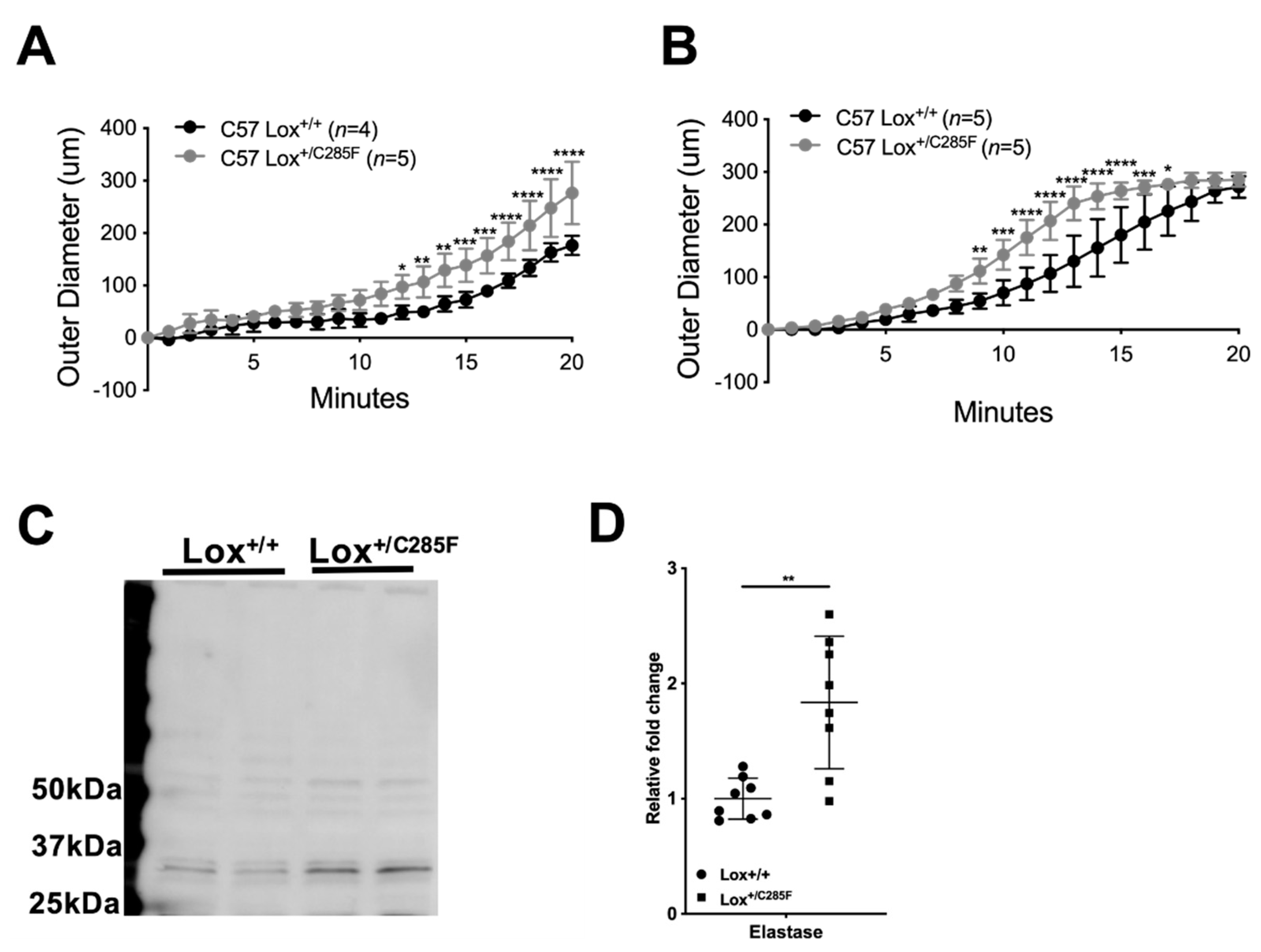
2.5. Lox+/C285F Vessels Are Susceptible to Elastolytic Damage and Show Increased Elastase Production
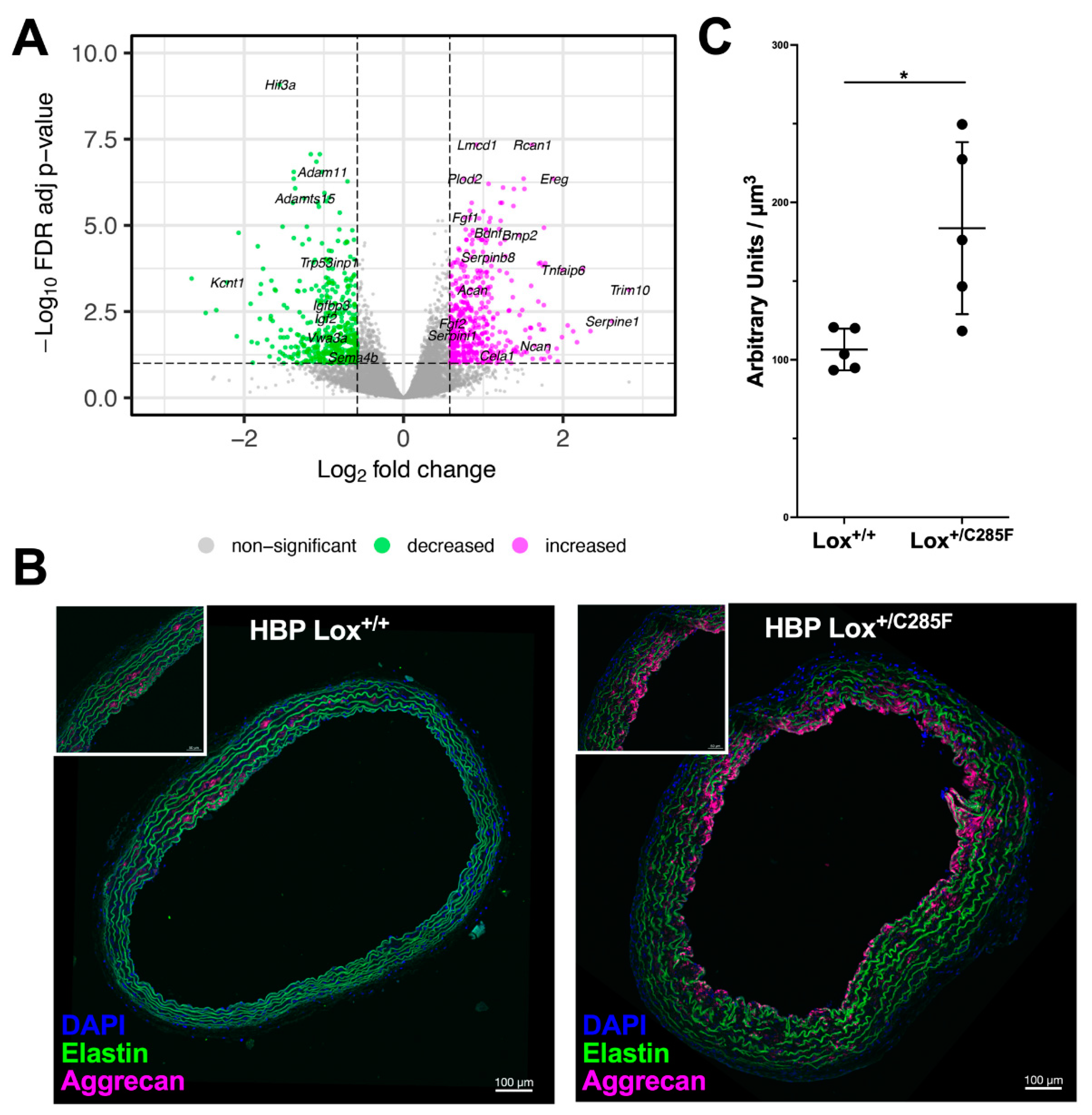
2.6. Lox+/C285F Gene Expression Pattern Reveals Increased Matrix Remodeling
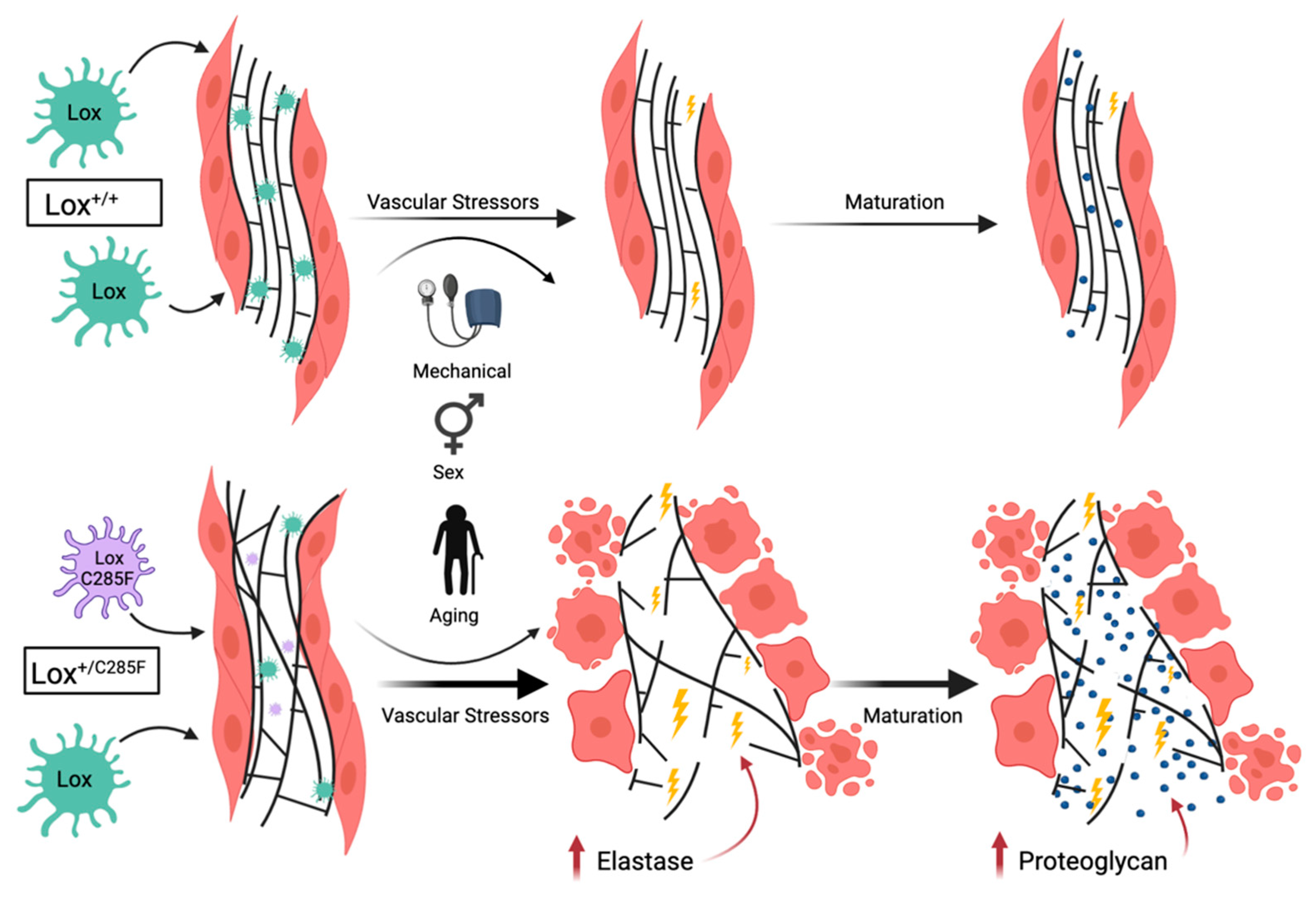
3. Discussion
4. Materials and Methods
4.1. Mouse Strains and Breeding
4.2. Systemic Blood Pressure and Heart Rate Measurement
4.3. Castration
4.4. Histology
4.5. Two-Photon Microscopy Imaging and Analysis
4.6. Focused Ion Beam-Scanning Electron Microscopy (FIB-SEM)
4.7. Advanced Imaging and Analysis
4.8. Biochemical Analysis on Elastin and Collagen Contents
4.9. RNA Preparation for QPCR and RNA Seq
4.10. Lysyl Oxidase Enzyme Activity Assay
4.11. Isolation and Protein Extraction from Mouse Aorta for Western Blot
4.12. Isolation of Mouse Embryonic Fibroblasts (MEFs)
4.13. Protein Extraction from MEFs
4.14. Western Blotting
4.15. Pressure–Diameter Testing with and without Elastase Treatment
4.16. Immunofluorescence
4.17. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kagan, H.M.; Li, W. Lysyl oxidase: Properties, specificity, and biological roles inside and outside of the cell. J. Cell Biochem. 2003, 88, 660–672. [Google Scholar] [CrossRef]
- Rodríguez, C.; Martínez-González, J.; Raposo, B.; Alcudia, J.F.; Guadall, A.; Badimon, L. Regulation of lysyl oxidase in vascular cells: Lysyl oxidase as a new player in cardiovascular diseases. Cardiovasc. Res. 2008, 79, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmelzer, C.E.H.; Hedtke, T.; Heinz, A. Unique molecular networks: Formation and role of elastin cross-links. IUBMB Life 2020, 72, 842–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, H.M.; Vaysberg, M.; Mikels, A.; McCauley, S.; Velayo, A.C.; Garcia, C.; Smith, V. Modulation of lysyl oxidase-like 2 enzymatic activity by an allosteric antibody inhibitor. J. Biol. Chem. 2010, 285, 20964–20974. [Google Scholar] [CrossRef] [Green Version]
- Lopez, K.M.; Greenaway, F.T. Identification of the copper-binding ligands of lysyl oxidase. J. Neural Transm. 2011, 118, 1101–1109. [Google Scholar] [CrossRef]
- Chen, X.; Greenaway, F.T. Identification of the disulfide bonds of lysyl oxidase. J. Neural Transm. 2011, 118, 1111–1114. [Google Scholar] [CrossRef]
- Hornstra, I.K.; Birge, S.; Starcher, B.; Bailey, A.J.; Mecham, R.P.; Shapiro, S.D. Lysyl oxidase is required for vascular and diaphragmatic development in mice. J. Biol. Chem. 2003, 278, 14387–14393. [Google Scholar] [CrossRef] [Green Version]
- Lee, V.S.; Halabi, C.M.; Hoffman, E.P.; Carmichael, N.; Leshchiner, I.; Lian, C.G.; Bierhals, A.J.; Vuzman, D.; Brigham Genomic, M.; Mecham, R.P.; et al. Loss of function mutation in LOX causes thoracic aortic aneurysm and dissection in humans. Proc. Natl. Acad. Sci. USA 2016, 113, 8759–8764. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.C.; Regalado, E.S.; Gong, L.; Duan, X.; Santos-Cortez, R.L.; Arnaud, P.; Ren, Z.; Cai, B.; Hostetler, E.M.; Moran, R.; et al. LOX Mutations Predispose to Thoracic Aortic Aneurysms and Dissections. Circ. Res. 2016, 118, 928–934. [Google Scholar] [CrossRef] [Green Version]
- Renard, M.; Francis, C.; Ghosh, R.; Scott, A.F.; Witmer, P.D.; Ades, L.C.; Andelfinger, G.U.; Arnaud, P.; Boileau, C.; Callewaert, B.L.; et al. Clinical Validity of Genes for Heritable Thoracic Aortic Aneurysm and Dissection. J. Am. Coll. Cardiol. 2018, 72, 605–615. [Google Scholar] [CrossRef]
- Van Gucht, I.; Krebsova, A.; Diness, B.R.; Laga, S.; Adlam, D.; Kempers, M.; Samani, N.J.; Webb, T.R.; Baranowska, A.A.; Van Den Heuvel, L.; et al. Novel LOX Variants in Five Families with Aortic/Arterial Aneurysm and Dissection with Variable Connective Tissue Findings. Int. J. Mol. Sci. 2021, 22, 7111. [Google Scholar] [CrossRef] [PubMed]
- Renner, S.; Schüler, H.; Alawi, M.; Kolbe, V.; Rybczynski, M.; Woitschach, R.; Sheikhzadeh, S.; Stark, V.C.; Olfe, J.; Roser, E.; et al. Next-generation sequencing of 32 genes associated with hereditary aortopathies and related disorders of connective tissue in a cohort of 199 patients. Genet. Med. 2019, 21, 1832–1841. [Google Scholar] [CrossRef]
- Cirnu, A.; Kolokotronis, K.; Walz, K.; Kilinç, A.; Janz, A.; Williams, T.; Busch, A.; Rost, S.; Gerull, B. Novel Mutation in LOX Associates With a Complex Aneurysmal Vascular and Cardiac Phenotype. Circ. Genom. Precis Med. 2021, 14, e003217. [Google Scholar] [CrossRef] [PubMed]
- Biesecker, L.G.; Adam, M.P.; Alkuraya, F.S.; Amemiya, A.R.; Bamshad, M.J.; Beck, A.E.; Bennett, J.T.; Bird, L.M.; Carey, J.C.; Chung, B.; et al. A dyadic approach to the delineation of diagnostic entities in clinical genomics. Am. J. Hum. Genet. 2021, 108, 8–15. [Google Scholar] [CrossRef]
- Lo, C. Information submitted by the NHLBI Cardiovascular Development Consortium (CvDC), Bench to Bassinet Program (B2B/CvDC) MGI Direct Data Submission (B2B/CvDC). In MGI: J:175213. 2011. Available online: http://www.informatics.jax.org/allele/MGI:5313524 (accessed on 22 April 2022).
- Krebs, C.J.; Krawetz, S.A. Lysyl oxidase copper-talon complex: A model. Biochim. Biophys. Acta 1993, 1202, 7–12. [Google Scholar] [CrossRef]
- Kanematsu, Y.; Kanematsu, M.; Kurihara, C.; Tsou, T.L.; Nuki, Y.; Liang, E.I.; Makino, H.; Hashimoto, T. Pharmacologically induced thoracic and abdominal aortic aneurysms in mice. Hypertension 2010, 55, 1267–1274. [Google Scholar] [CrossRef]
- Lee, V.S.; Halabi, C.M.; Broekelmann, T.J.; Trackman, P.C.; Stitziel, N.O.; Mecham, R.P. Intracellular retention of mutant lysyl oxidase leads to aortic dilation in response to increased hemodynamic stress. JCI Insight. 2019, 5, e127748. [Google Scholar] [CrossRef]
- Trackman, P.C.; Bais, M.V. Measurement of lysyl oxidase activity from small tissue samples and cell cultures. Methods Cell Biol. 2018, 143, 147–156. [Google Scholar] [CrossRef]
- Halabi, C.M.; Broekelmann, T.J.; Knutsen, R.H.; Ye, L.; Mecham, R.P.; Kozel, B.A. Chronic antihypertensive treatment improves pulse pressure but not large artery mechanics in a mouse model of congenital vascular stiffness. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1008–H1016. [Google Scholar] [CrossRef]
- Wolinsky, H.; Glagov, S. Structural Basis for the Static Mechanical Properties of the Aortic Media. Circ. Res. 1964, 14, 400–413. [Google Scholar] [CrossRef] [Green Version]
- Naba, A.; Clauser, K.R.; Hoersch, S.; Liu, H.; Carr, S.A.; Hynes, R.O. The matrisome: In silico definition and in vivo characterization by proteomics of normal and tumor extracellular matrices. Mol. Cell Proteom. 2012, 11, M111.014647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cikach, F.S.; Koch, C.D.; Mead, T.J.; Galatioto, J.; Willard, B.B.; Emerton, K.B.; Eagleton, M.J.; Blackstone, E.H.; Ramirez, F.; Roselli, E.E.; et al. Massive aggrecan and versican accumulation in thoracic aortic aneurysm and dissection. JCI Insight. 2018, 3, e97167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozel, B.A.; Knutsen, R.H.; Ye, L.; Ciliberto, C.H.; Broekelmann, T.J.; Mecham, R.P. Genetic modifiers of cardiovascular phenotype caused by elastin haploinsufficiency act by extrinsic noncomplementation. J. Biol. Chem. 2011, 286, 44926–44936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramanath, V.S.; Oh, J.K.; Sundt, T.M., 3rd; Eagle, K.A. Acute aortic syndromes and thoracic aortic aneurysm. Mayo Clin. Proc. 2009, 84, 465–481. [Google Scholar] [CrossRef] [Green Version]
- Lacro, R.V.; Dietz, H.C.; Sleeper, L.A.; Yetman, A.T.; Bradley, T.J.; Colan, S.D.; Pearson, G.D.; Selamet Tierney, E.S.; Levine, J.C.; Atz, A.M.; et al. Atenolol versus losartan in children and young adults with Marfan’s syndrome. N. Engl. J. Med. 2014, 371, 2061–2071. [Google Scholar] [CrossRef] [Green Version]
- Hiratzka, L.F.; Bakris, G.L.; Beckman, J.A.; Bersin, R.M.; Carr, V.F.; Casey, D.E., Jr.; Eagle, K.A.; Hermann, L.K.; Isselbacher, E.M.; Kazerooni, E.A.; et al. 2010 ACCF/AHA/AATS/ACR/ASA/SCA/SCAI/SIR/STS/SVM guidelines for the diagnosis and management of patients with Thoracic Aortic Disease: A report of the American College of Cardiology Foundation/American Heart Association Task Force on Practice Guidelines, American Association for Thoracic Surgery, American College of Radiology, American Stroke Association, Society of Cardiovascular Anesthesiologists, Society for Cardiovascular Angiography and Interventions, Society of Interventional Radiology, Society of Thoracic Surgeons, and Society for Vascular Medicine. Circulation 2010, 121, e266–e369. [Google Scholar] [CrossRef] [Green Version]
- Alsiraj, Y.; Thatcher, S.E.; Blalock, E.; Fleenor, B.; Daugherty, A.; Cassis, L.A. Sex Chromosome Complement Defines Diffuse Versus Focal Angiotensin II-Induced Aortic Pathology. Arter. Thromb. Vasc. Biol. 2018, 38, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Bowen, C.J.; Calderón Giadrosic, J.F.; Burger, Z.; Rykiel, G.; Davis, E.C.; Helmers, M.R.; Benke, K.; Gallo MacFarlane, E.; Dietz, H.C. Targetable cellular signaling events mediate vascular pathology in vascular Ehlers-Danlos syndrome. J. Clin. Investig. 2020, 130, 686–698. [Google Scholar] [CrossRef] [Green Version]
- Vallet, S.D.; Gueroult, M.; Belloy, N.; Dauchez, M.; Ricard-Blum, S. A Three-Dimensional Model of Human Lysyl Oxidase, a Cross-Linking Enzyme. ACS Omega 2019, 4, 8495–8505. [Google Scholar] [CrossRef]
- Thomassin, L.; Werneck, C.C.; Broekelmann, T.J.; Gleyzal, C.; Hornstra, I.K.; Mecham, R.P.; Sommer, P. The Pro-regions of lysyl oxidase and lysyl oxidase-like 1 are required for deposition onto elastic fibers. J. Biol. Chem. 2005, 280, 42848–42855. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Zhang, Y.; Kim, B.; Ge, G.; Annis, D.S.; Mosher, D.F.; Greenspan, D.S. Fibronectin binds and enhances the activity of bone morphogenetic protein 1. J. Biol. Chem. 2009, 284, 25879–25888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogelgren, B.; Polgár, N.; Szauter, K.M.; Ujfaludi, Z.; Laczkó, R.; Fong, K.S.; Csiszar, K. Cellular fibronectin binds to lysyl oxidase with high affinity and is critical for its proteolytic activation. J. Biol. Chem. 2005, 280, 24690–24697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Guimet, J.; Andilla, J.; Loza-Alvarez, P.; Egea, G. High-Resolution Morphological Approach to Analyse Elastic Laminae Injuries of the Ascending Aorta in a Murine Model of Marfan Syndrome. Sci. Rep. 2017, 7, 1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.A.; Spinale, F.G.; Ikonomidis, J.S. Transforming growth factor-beta signaling in thoracic aortic aneurysm development: A paradox in pathogenesis. J. Vasc. Res. 2009, 46, 119–137. [Google Scholar] [CrossRef] [Green Version]
- Loeys, B.L.; Schwarze, U.; Holm, T.; Callewaert, B.L.; Thomas, G.H.; Pannu, H.; De Backer, J.F.; Oswald, G.L.; Symoens, S.; Manouvrier, S.; et al. Aneurysm syndromes caused by mutations in the TGF-beta receptor. N. Engl. J. Med. 2006, 355, 788–798. [Google Scholar] [CrossRef]
- Kähäri, V.M. Dexamethasone suppresses elastin gene expression in human skin fibroblasts in culture. Biochem. Biophys. Res. Commun. 1994, 201, 1189–1196. [Google Scholar] [CrossRef]
- Mecham, R.P.; Morris, S.L.; Levy, B.D.; Wrenn, D.S. Glucocorticoids stimulate elastin production in differentiated bovine ligament fibroblasts but do not induce elastin synthesis in undifferentiated cells. J. Biol. Chem. 1984, 259, 12414–12418. [Google Scholar] [CrossRef]
- Humphrey, J.D. Possible mechanical roles of glycosaminoglycans in thoracic aortic dissection and associations with dysregulated transforming growth factor-β. Author reply. J. Vasc. Res. 2013, 50, 198–199. [Google Scholar] [CrossRef] [Green Version]
- Roccabianca, S.; Ateshian, G.A.; Humphrey, J.D. Biomechanical roles of medial pooling of glycosaminoglycans in thoracic aortic dissection. Biomech. Model Mechanobiol. 2014, 13, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Roccabianca, S.; Bellini, C.; Humphrey, J.D. Computational modelling suggests good, bad and ugly roles of glycosaminoglycans in arterial wall mechanics and mechanobiology. J. R. Soc. Interface 2014, 11, 20140397. [Google Scholar] [CrossRef]
- Koch, C.D.; Lee, C.M.; Apte, S.S. Aggrecan in Cardiovascular Development and Disease. J. Histochem. Cytochem. 2020, 68, 777–795. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, V.K.; Evans, A.N.; Wansapura, J.P.; Osinska, H.; Maddy, K.E.; Biechler, S.V.; Narmoneva, D.A.; Goodwin, R.L.; Hinton, R.B. Asymmetric cell-matrix and biomechanical abnormalities in elastin insufficiency induced aortopathy. Ann. Biomed. Eng. 2014, 42, 2014–2028. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Zhou, Y.; Chen, Y.; Feng, X.; Liu, C.; Jiang, D.S.; Geng, J.; Li, X.; Jiang, X.; Fang, Z.M. The Expression Patterns and Roles of Lysyl Oxidases in Aortic Dissection. Front. Cardiovasc. Med. 2021, 8, 692856. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, R.H.; Beeman, S.C.; Broekelmann, T.J.; Liu, D.; Tsang, K.M.; Kovacs, A.; Ye, L.; Danback, J.R.; Watson, A.; Wardlaw, A.; et al. Minoxidil improves vascular compliance, restores cerebral blood flow, and alters extracellular matrix gene expression in a model of chronic vascular stiffness. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H18–H32. [Google Scholar] [CrossRef]
- Malide, D. In Vivo Cell Tracking Using Two-Photon Microscopy. Methods Mol. Biol. 2016, 1444, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Bleck, C.K.E.; Kim, Y.; Willingham, T.B.; Glancy, B. Subcellular connectomic analyses of energy networks in striated muscle. Nat. Commun. 2018, 9, 5111. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Zuiderveld, K. Contrast limited adaptive histogram equalization. In Graphics Gems IV; Academic Press: Cambridge, MA, USA, 1994; pp. 474–485. [Google Scholar]
- Rasband, W.S. ImageJ: Image Processing and Analysis in Java. Astrophys. Source Code Libr. 2012, 1, 06013. [Google Scholar]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinform. 2015, 51, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. The Subread aligner: Fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013, 41, e108. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.H. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Gürtler, A.; Kunz, N.; Gomolka, M.; Hornhardt, S.; Friedl, A.A.; McDonald, K.; Kohn, J.E.; Posch, A. Stain-Free technology as a normalization tool in Western blot analysis. Anal. Biochem. 2013, 433, 105–111. [Google Scholar] [CrossRef]
- Gilda, J.E.; Gomes, A.V. Stain-Free total protein staining is a superior loading control to β-actin for Western blots. Anal. Biochem. 2013, 440, 186–188. [Google Scholar] [CrossRef] [Green Version]
- Rivero-Gutiérrez, B.; Anzola, A.; Martínez-Augustin, O.; de Medina, F.S. Stain-free detection as loading control alternative to Ponceau and housekeeping protein immunodetection in Western blotting. Anal. Biochem. 2014, 467, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Wagenseil, J.E.; Nerurkar, N.L.; Knutsen, R.H.; Okamoto, R.J.; Li, D.Y.; Mecham, R.P. Effects of elastin haploinsufficiency on the mechanical behavior of mouse arteries. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H1209–H1217. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsang, K.M.; Knutsen, R.H.; Billington, C.J., Jr.; Lindberg, E.; Steenbock, H.; Fu, Y.-P.; Wardlaw-Pickett, A.; Liu, D.; Malide, D.; Yu, Z.-X.; et al. Copper-Binding Domain Variation in a Novel Murine Lysyl Oxidase Model Produces Structurally Inferior Aortic Elastic Fibers Whose Failure Is Modified by Age, Sex, and Blood Pressure. Int. J. Mol. Sci. 2022, 23, 6749. https://doi.org/10.3390/ijms23126749
Tsang KM, Knutsen RH, Billington CJ Jr., Lindberg E, Steenbock H, Fu Y-P, Wardlaw-Pickett A, Liu D, Malide D, Yu Z-X, et al. Copper-Binding Domain Variation in a Novel Murine Lysyl Oxidase Model Produces Structurally Inferior Aortic Elastic Fibers Whose Failure Is Modified by Age, Sex, and Blood Pressure. International Journal of Molecular Sciences. 2022; 23(12):6749. https://doi.org/10.3390/ijms23126749
Chicago/Turabian StyleTsang, Kit Man, Russell H. Knutsen, Charles J. Billington, Jr., Eric Lindberg, Heiko Steenbock, Yi-Ping Fu, Amanda Wardlaw-Pickett, Delong Liu, Daniela Malide, Zu-Xi Yu, and et al. 2022. "Copper-Binding Domain Variation in a Novel Murine Lysyl Oxidase Model Produces Structurally Inferior Aortic Elastic Fibers Whose Failure Is Modified by Age, Sex, and Blood Pressure" International Journal of Molecular Sciences 23, no. 12: 6749. https://doi.org/10.3390/ijms23126749
APA StyleTsang, K. M., Knutsen, R. H., Billington, C. J., Jr., Lindberg, E., Steenbock, H., Fu, Y.-P., Wardlaw-Pickett, A., Liu, D., Malide, D., Yu, Z.-X., Bleck, C. K. E., Brinckmann, J., & Kozel, B. A. (2022). Copper-Binding Domain Variation in a Novel Murine Lysyl Oxidase Model Produces Structurally Inferior Aortic Elastic Fibers Whose Failure Is Modified by Age, Sex, and Blood Pressure. International Journal of Molecular Sciences, 23(12), 6749. https://doi.org/10.3390/ijms23126749