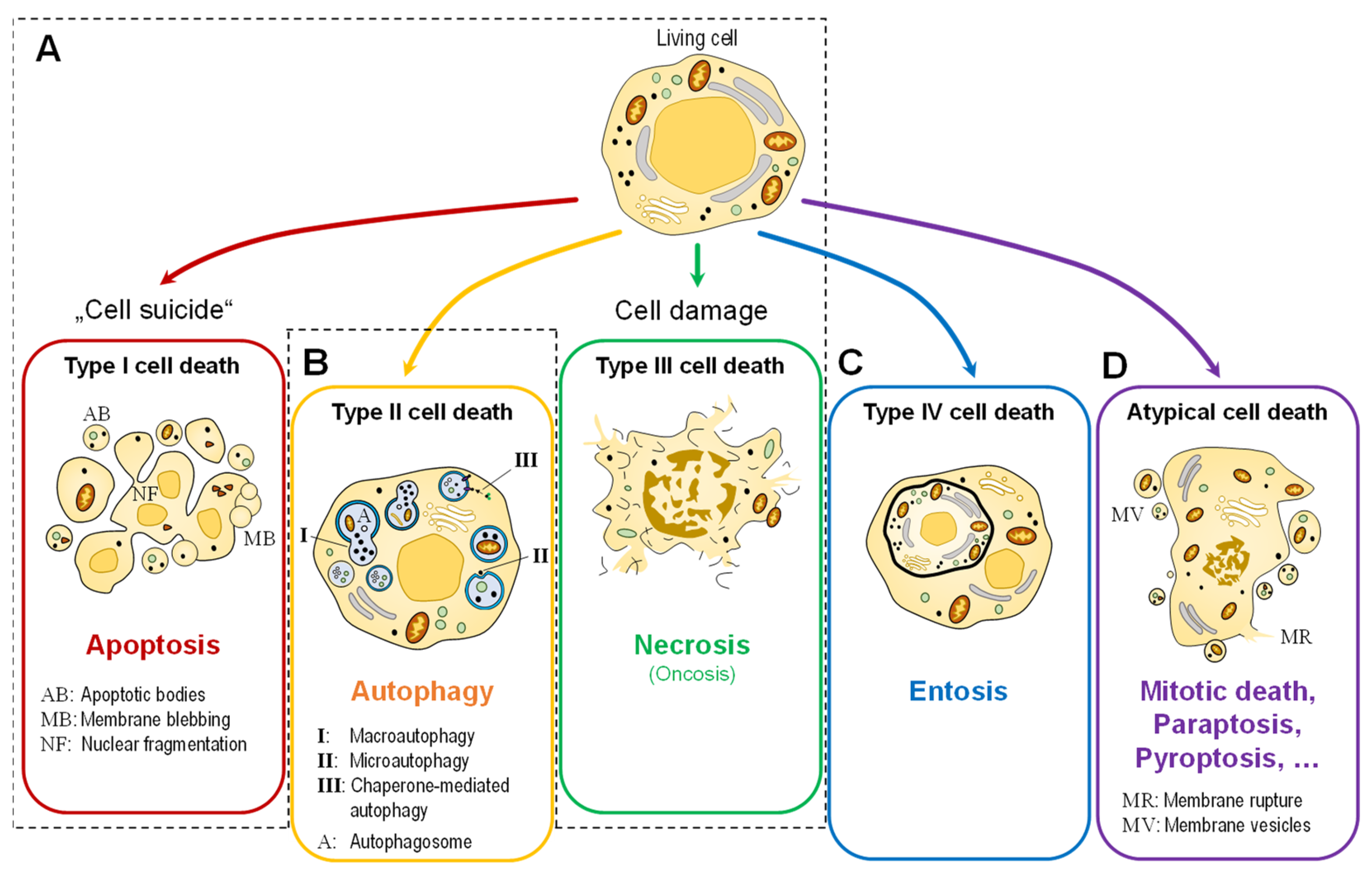
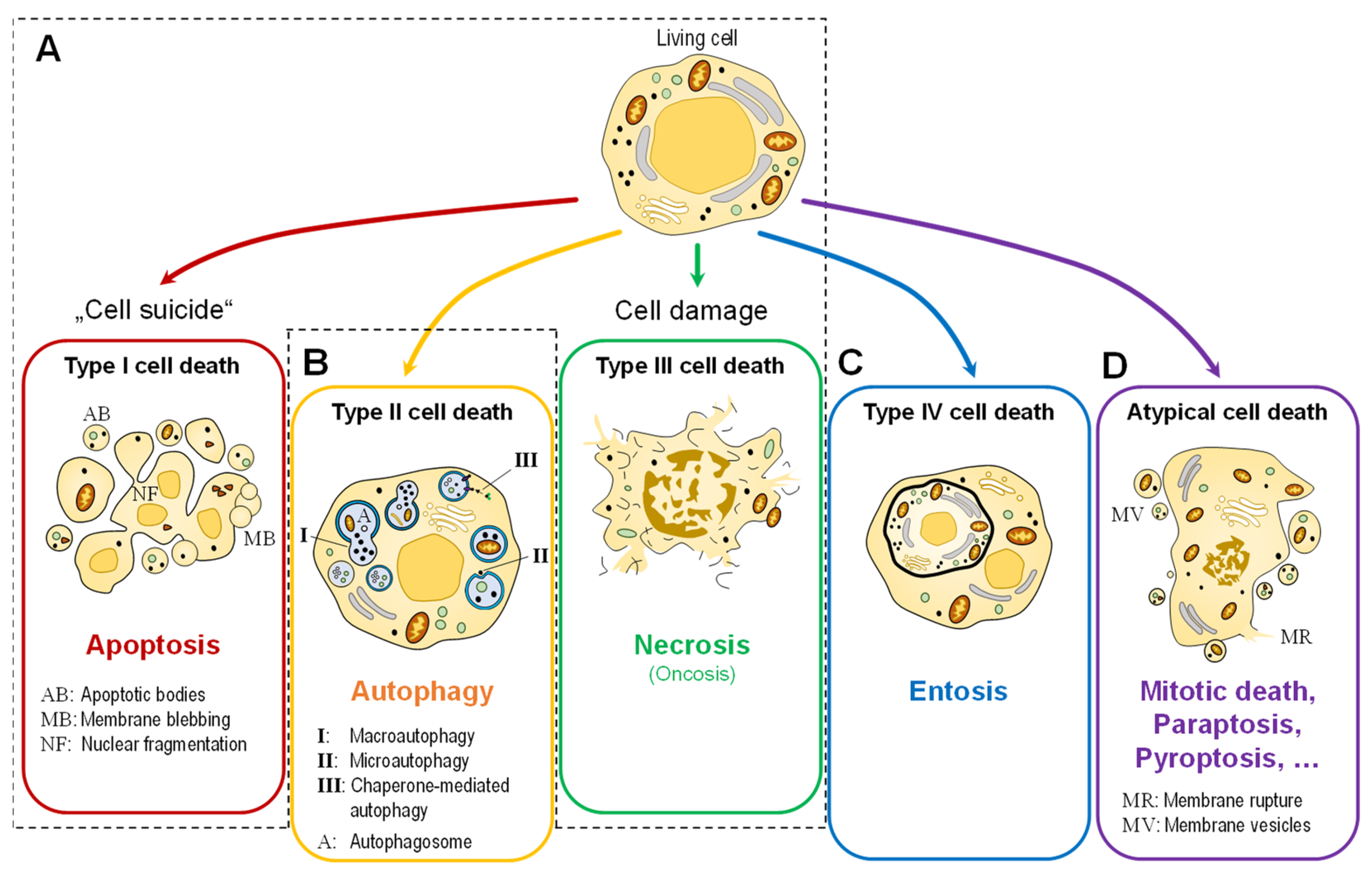
To Die or Not to Die: Cell Death in Biology and Disease



{kind=link}
Author Contributions
Funding
Conflicts of Interest
References
- Vogt, K.C. Untersuchungen über die Entwicklungsgeschichte der Geburtshelferkröte (Alytes obstetricans); Jent & Gassmann: Solothurn, Niedersachsen, Germany, 1842. [Google Scholar]
- Lockshin, R.A.; Williams, C.M. Programmed cell death—II. Endocrine potentiation of the breakdown of the intersegmental muscles of silkmoths. J. Insect Physiol. 1964, 10, 643–649. [Google Scholar] [CrossRef]
- Tata, J.R. Requirement for RNA and protein synthesis for induced regression of the tadpole tail in organ culture. Dev. Biol. 1966, 13, 77–94. [Google Scholar] [CrossRef]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A Basic Biological Phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, M.E. Apoptosis meets necrosis. Nature 2011, 471, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, E.; Cui, Y.-H.; He, Y.-Y. Context-dependent roles of RNA modifications in stress responses and diseases. Int. J. Mol. Sci. 2021, 22, 1949. [Google Scholar] [CrossRef] [PubMed]
- Overholtzer, M.; Mailleux, A.A.; Mouneimne, G.; Normand, G.; Schnitt, S.J.; King, R.W.; Cibas, E.S.; Brugge, J.S. A nonapoptotic cell death process, entosis, that occurs by cell-in-cell invasion. Cell 2007, 131, 966–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kianfar, M.; Balcerak, A.; Chmielarczyk, M.; Tarnowski, L.; Grzybowska, E.A. Cell death by entosis: Triggers, molecular mechanisms and clinical significance. Int. J. Mol. Sci. 2022, 23, 4985. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of Bcl-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Knuth, A.-K.; Huard, A.; Naeem, Z.; Rappl, P.; Bauer, R.; Mota, A.C.; Schmid, T.; Fleming, I.; Brüne, B.; Fulda, S.; et al. Apoptotic cells induce proliferation of peritoneal macrophages. Int. J. Mol. Sci. 2021, 22, 2230. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A target for anticancer therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, E.; Forika, G.; Mohacsi, R.; Nemeth, Z.; Krenacs, T.; Dank, M. Methyl-Donors can induce apoptosis and attenuate both the Akt and the Erk1/2 mediated proliferation pathways in breast and lung cancer cell lines. Int. J. Mol. Sci. 2021, 22, 3598. [Google Scholar] [CrossRef] [PubMed]
- Jozkowiak, M.; Dyszkiewicz-Konwinska, M.; Ramlau, P.; Kranc, W.; Spaczynska, J.; Wierzchowski, M.; Kaczmarek, M.; Jodynis-Liebert, J.; Piotrowska-Kempisty, H. Individual and combined treatments with methylated resveratrol analogue dMU-214 and gefitinib inhibit tongue cancer cells growth via apoptosis induction and egfr inhibition. Int. J. Mol. Sci. 2021, 22, 6180. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, S.; Deloch, L.; Hader, M.; Derer, A.; Grottker, F.; Weissmann, T.; Hecht, M.; Gostian, A.-O.; Fietkau, R.; Frey, B.; et al. Hypofractionated radiotherapy upregulates several immune checkpoint molecules in head and neck squamous cell carcinoma cells independently of the HPV status while ICOS-L is upregulated only on HPV-positive cells. Int. J. Mol. Sci. 2021, 22, 9114. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krüger, M.; Richter, P. To Die or Not to Die: Cell Death in Biology and Disease. Int. J. Mol. Sci. 2022, 23, 6734. https://doi.org/10.3390/ijms23126734
Krüger M, Richter P. To Die or Not to Die: Cell Death in Biology and Disease. International Journal of Molecular Sciences. 2022; 23(12):6734. https://doi.org/10.3390/ijms23126734
Chicago/Turabian StyleKrüger, Marcus, and Peter Richter. 2022. "To Die or Not to Die: Cell Death in Biology and Disease" International Journal of Molecular Sciences 23, no. 12: 6734. https://doi.org/10.3390/ijms23126734
APA StyleKrüger, M., & Richter, P. (2022). To Die or Not to Die: Cell Death in Biology and Disease. International Journal of Molecular Sciences, 23(12), 6734. https://doi.org/10.3390/ijms23126734