
Comparison of Biological Features of Wild European Rabbit Mesenchymal Stem Cells Derived from Different Tissues



 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
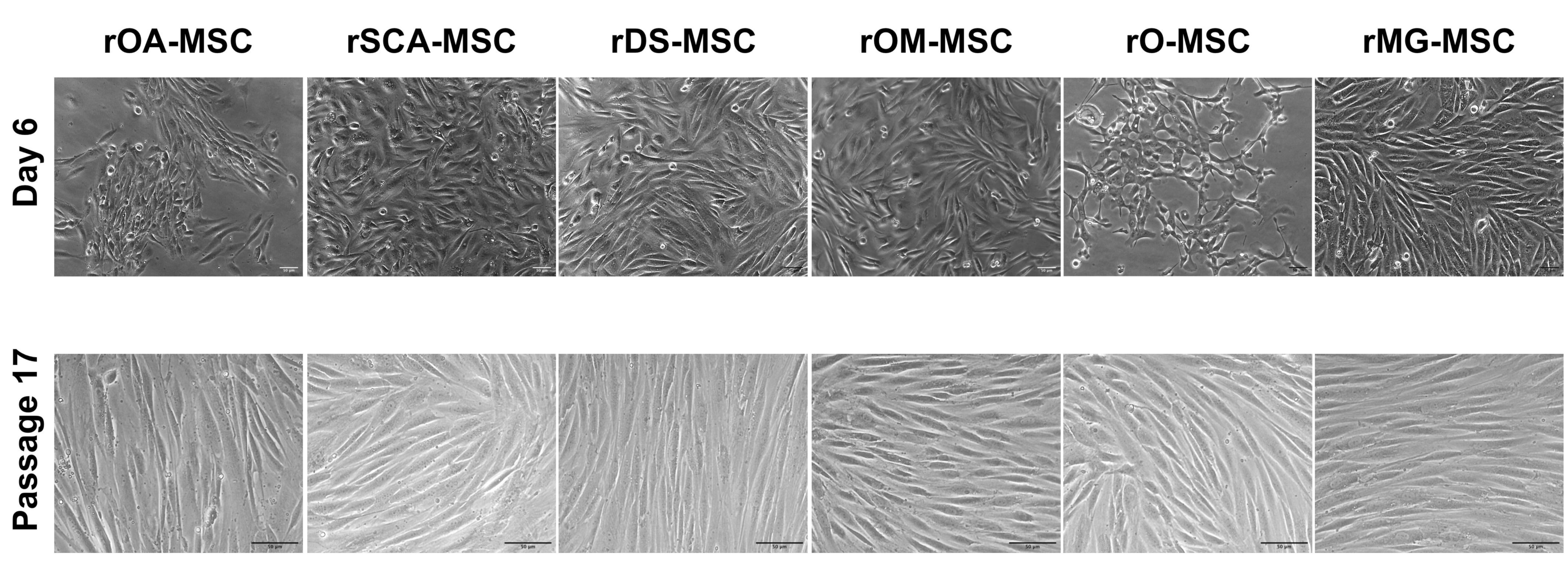
2.1. Morphological Features of Different rMSC
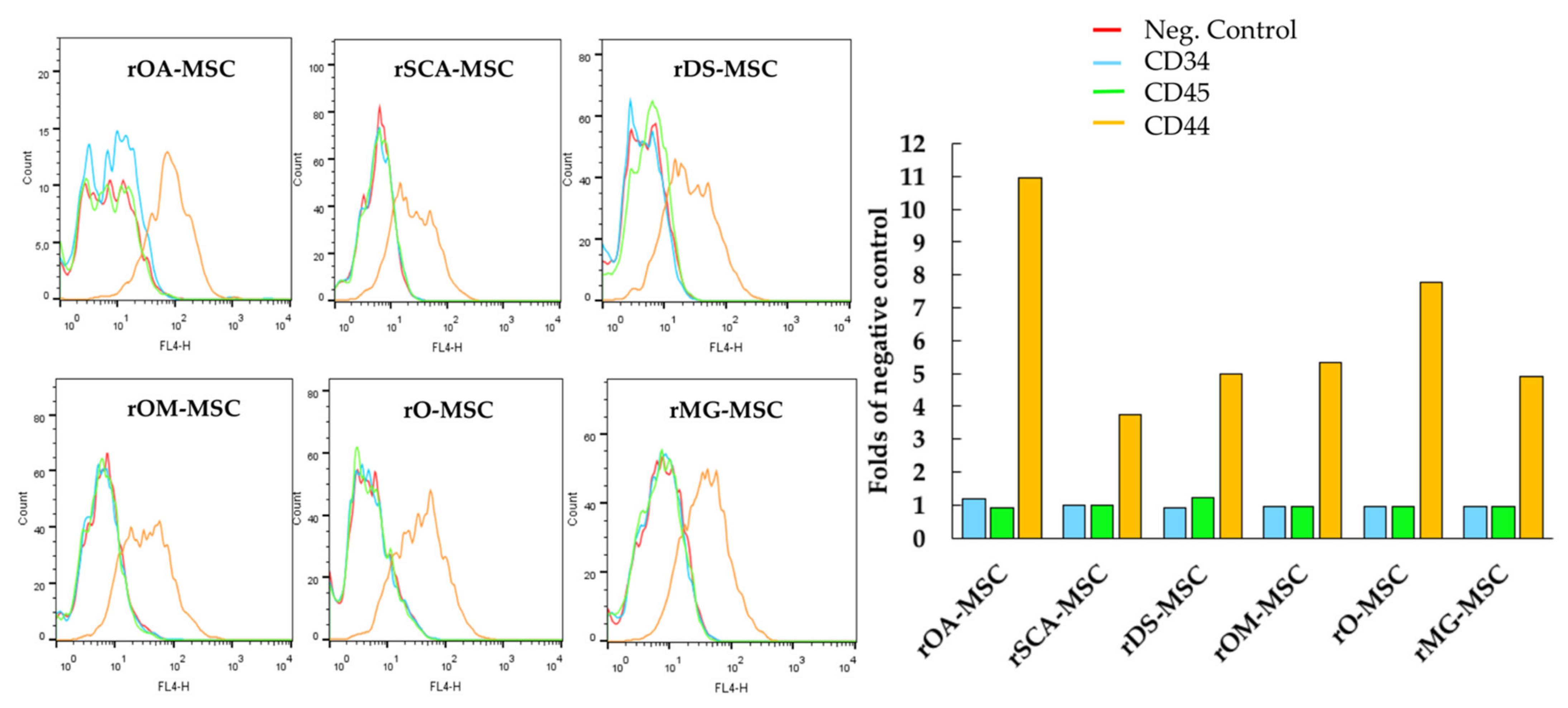
2.2. Immunophenotypic Characterization by Flow Cytometry
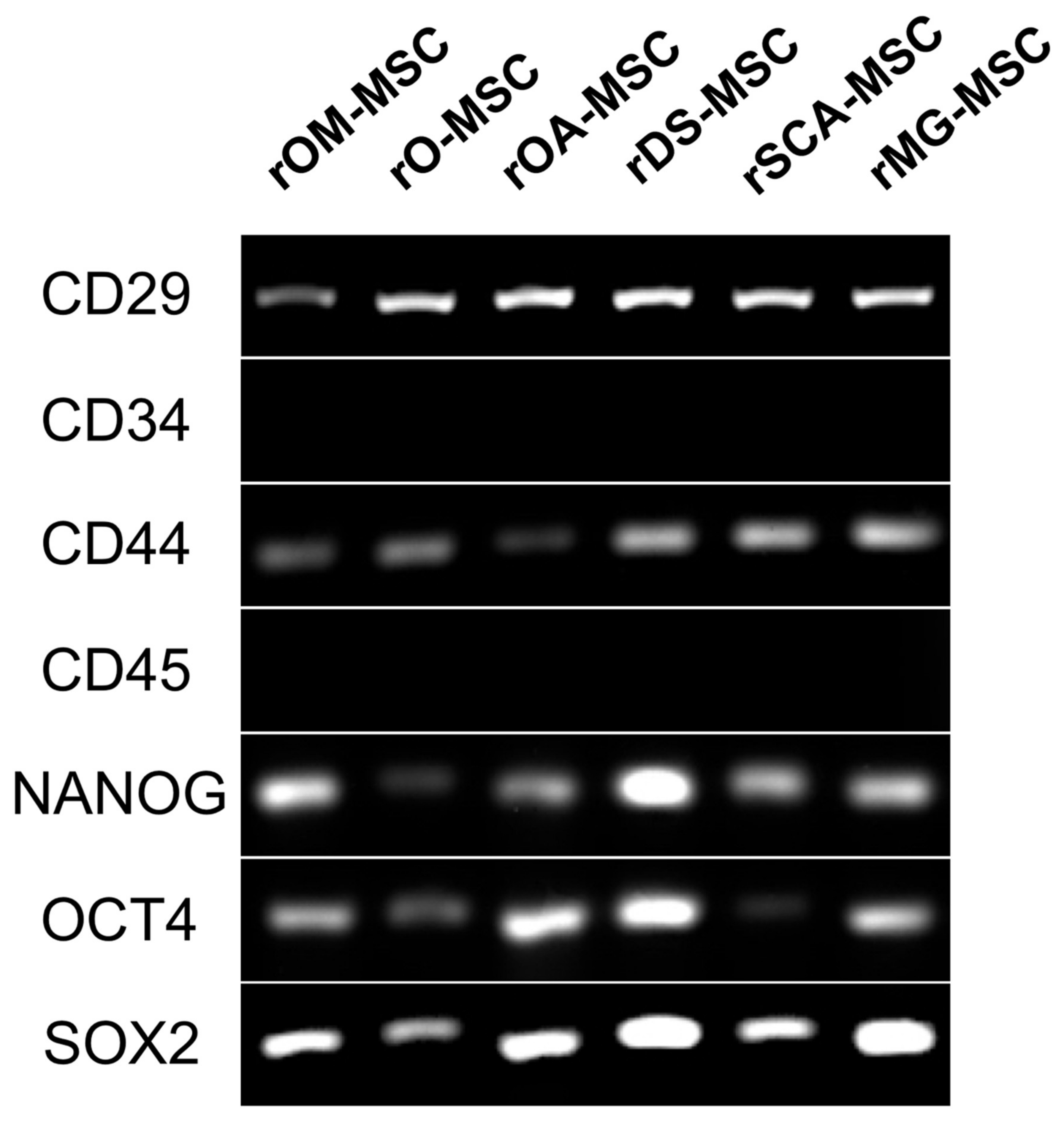
2.3. Gene Expression of Surface and Pluripotency Markers Detected by RT-PCR
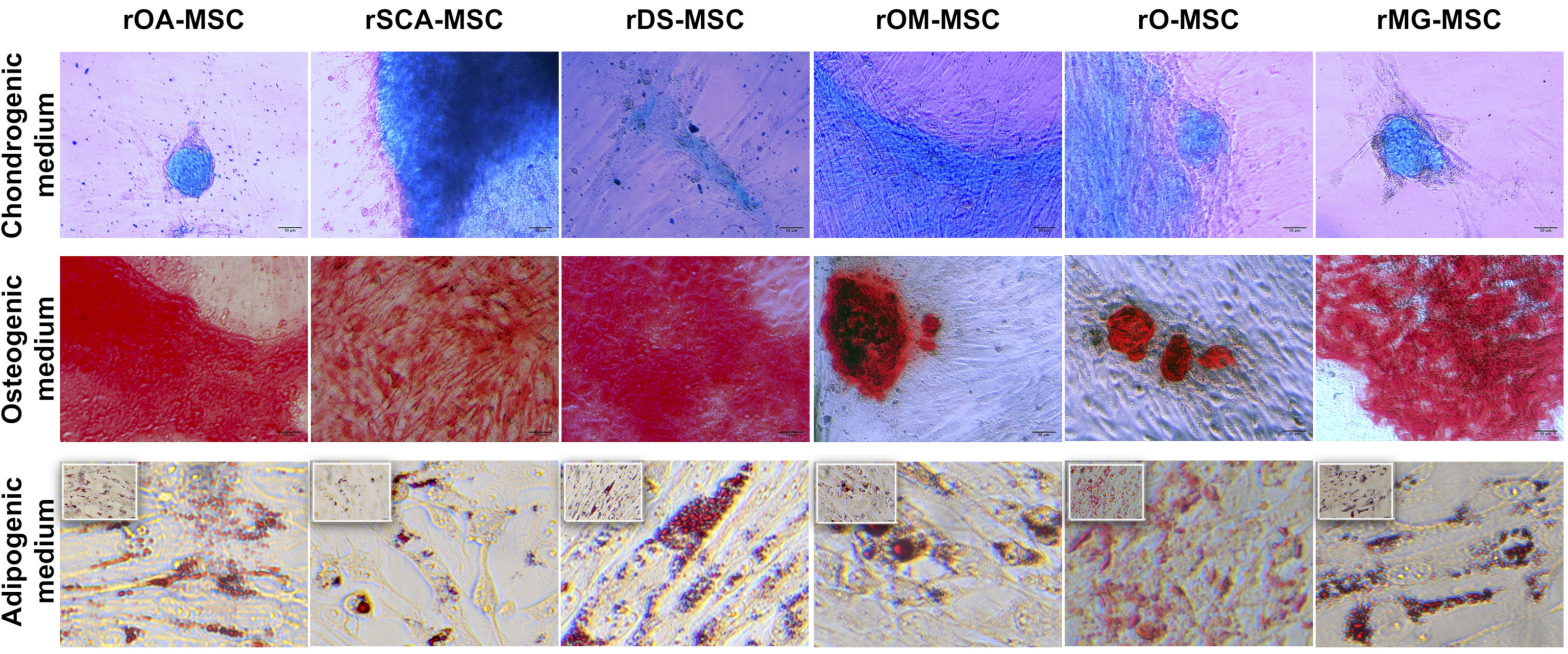
2.4. In Vitro Differentiation of rMSCs
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of rMSCs
4.2. Immunocytochemical Analysis by Flow Cytometry
4.3. RT-PCR Analysis of Characteristic Mesenchymal mRNA Expression in rMSC
4.4. In Vitro Differentiation Potential Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferreira, P.C. European rabbit research in the Iberian Peninsula: State of the art and future perspectives. Eur. J. Wildl. Res. 2012, 58, 885–895. [Google Scholar] [CrossRef]
- Carneiro, M.; Afonso, S.; Geraldes, A.; Garreau, H.; Bolet, G.; Boucher, S.; Tircazes, A.; Queney, G.; Nachman, M.W.; Ferrand, N. The genetic structure of domestic rabbits. Mol. Biol. Evol. 2011, 28, 1801–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, J.M.; Carneiro, M.; Afonso, S.; Lopes, S.; Garreau, H.; Boucher, S.; Allain, D.; Queney, G.; Esteves, P.J.; Bolet, G.; et al. Levels and Patterns of Genetic Diversity and Population Structure in Domestic Rabbits. PLoS ONE 2015, 10, e0144687. [Google Scholar] [CrossRef] [Green Version]
- Abrantes, J.; van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertagnoli, S.; Marchandeau, S. Myxomatosis. Rev. Sci. Tech. 2015, 34, 539–547, 549–556. [Google Scholar] [PubMed]
- Rouco, C.; Abrantes, J.; Serronha, A.; Lopes, A.M.; Maio, E.; Magalhães, M.J.; Blanco, E.; Bárcena, J.; Esteves, P.J.; Santos, N.; et al. Epidemiology of RHDV2 (Lagovirus europaeus/GI.2) in free-living wild European rabbits in Portugal. Transbound. Emerg. Dis. 2018, 65, e373–e382. [Google Scholar] [CrossRef] [PubMed]
- Villafuerte, R.; Castro, F.; Ramírez, E.; Cotilla, I.; Parra, F.; Delibes-Mateos, M.; Recuerda, P.; Rouco, C. Large-scale assessment of myxomatosis prevalence in European wild rabbits (Oryctolagus cuniculus) 60years after first outbreak in Spain. Res. Vet. Sci. 2017, 114, 281–286. [Google Scholar] [CrossRef]
- Effler, P. Australia’s War against Rabbits: The Story of Rabbit Haemorrhagic Disease. Emerg. Infect. Dis. J. 2015, 21, 735. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, L. Frank Fenner and Bernardino Fantini, Biological control of vertebrate pests: The history of myxomatosis, an experiment in evolution, Wallingford, CABI Publishing, 1999, pp. xii, 339, illus., £60.00 (0-85199-323-0). Med. Hist. 2001, 45, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Kerr, P.J.; Hall, R.N.; Strive, T. Viruses for Landscape-Scale Therapy: Biological Control of Rabbits in Australia. Methods Mol Biol. 2021, 2225, 1–23. [Google Scholar] [CrossRef]
- Monterroso, P.; Garrote, G.; Serronha, A.; Santos, E.; Delibes-Mateos, M.; Abrantes, J.; de Ayala, R.P.; Silvestre, F.; Carvalho, J.; Vasco, I.; et al. Disease-mediated bottom-up regulation: An emergent virus affects a keystone prey, and alters the dynamics of trophic webs. Sci. Rep. 2016, 6, 36072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkholder, T.; Linton, G.; Hoyt, R.; Young, R. The Rabbit as an Experimental Model. In The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Suckow, M.A., Stevens, K.A., Wilson, R.P., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 529–560. [Google Scholar] [CrossRef]
- Trohatou, O.; Roubelakis, M.G. Mesenchymal Stem/Stromal Cells in Regenerative Medicine: Past, Present, and Future. Cell Reprogram 2017, 19, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Cagliani, J.; Grande, D.; Molmenti, E.P.; Miller, E.J.; Rilo, H.L.R. Immunomodulation by Mesenchymal Stromal Cells and Their Clinical Applications. J. Stem Cell Regen. Biol. 2017, 3, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Avanzi, S.; Leoni, V.; Rotola, A.; Alviano, F.; Solimando, L.; Lanzoni, G.; Bonsi, L.; di Luca, D.; Marchionni, C.; Alvisi, G.; et al. Susceptibility of human placenta derived mesenchymal stromal/stem cells to human herpesviruses infection. PLoS ONE 2013, 8, e71412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arzi, B.; Kol, A.; Murphy, B.; Walker, N.J.; Wood, J.A.; Clark, K.; Verstraete, F.J.; Borjesson, D.L. Feline foamy virus adversely affects feline mesenchymal stem cell culture and expansion: Implications for animal model development. Stem Cells Dev. 2015, 24, 814–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, M.B.; Sampayo-Escobar, V.; Green, R.; Moore, M.L.; Mohapatra, S.; Mohapatra, S.S. Respiratory Syncytial Virus-Infected Mesenchymal Stem Cells Regulate Immunity via Interferon Beta and Indoleamine-2,3-Dioxygenase. PLoS ONE 2016, 11, e0163709. [Google Scholar] [CrossRef] [Green Version]
- Claessen, C.; Favoreel, H.; Ma, G.; Osterrieder, N.; De Schauwer, C.; Piepers, S.; Van de Walle, G.R. Equid herpesvirus 1 (EHV1) infection of equine mesenchymal stem cells induces a pUL56-dependent downregulation of select cell surface markers. Vet. Microbiol. 2015, 176, 32–39. [Google Scholar] [CrossRef]
- Qian, X.; Xu, C.; Fang, S.; Zhao, P.; Wang, Y.; Liu, H.; Yuan, W.; Qi, Z. Exosomal MicroRNAs Derived From Umbilical Mesenchymal Stem Cells Inhibit Hepatitis C Virus Infection. Stem Cells Transl. Med. 2016, 5, 1190–1203. [Google Scholar] [CrossRef] [Green Version]
- Taechangam, N.; Kol, A.; Arzi, B.; Borjesson, D.L. Multipotent Stromal Cells and Viral Interaction: Current Implications for Therapy. Stem Cell Rev. Rep. 2021, 18, 214–277. [Google Scholar] [CrossRef]
- Tirpáková, M.; Vašíček, J.; Svoradová, A.; Baláži, A.; Tomka, M.; Bauer, M.; Makarevich, A.; Chrenek, P. Phenotypical Characterization and Neurogenic Differentiation of Rabbit Adipose Tissue-Derived Mesenchymal Stem Cells. Genes 2021, 12, 431. [Google Scholar] [CrossRef]
- Nugraha, A.P.; Rantam, F.A.; Narmada, I.B.; Ernawati, D.S.; Ihsan, I.S. Gingival-Derived Mesenchymal Stem Cell from Rabbit (Oryctolagus cuniculus): Isolation, Culture, and Characterization. Eur. J. Dent. 2021, 15, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dong, J.; He, Y.; Mao, W.; Tang, H.; Dong, Y.; Lyu, F. Comparative analysis of mesenchymal stromal cells derived from rabbit bone marrow and Wharton’s jelly for adipose tissue engineering. Connect. Tissue Res. 2020, 61, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Kovac, M.; Vasicek, J.; Kulikova, B.; Bauer, M.; Curlej, J.; Balazi, A.; Chrenek, P. Different RNA and protein expression of surface markers in rabbit amniotic fluid-derived mesenchymal stem cells. Biotechnol. Prog. 2017, 33, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Wang, F.; Rong, S.; Ren, J.; Wan, J.S.; Shi, L.X.; Wu, Z.; Liu, T.; Li, Q. Enhancement of osteogenesis of rabbit bone marrow derived mesenchymal stem cells by transfection of human BMP-2 and EGFP recombinant adenovirus via Wnt signaling pathway. Exp. Ther. Med. 2018, 16, 4030–4036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakhtina, A.; Tohfafarosh, M.; Lichtler, A.; Arinzeh, T.L. Characterization and differentiation potential of rabbit mesenchymal stem cells for translational regenerative medicine. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Huang, Y.H.; Chang, N.K.; Lin, W.C.; Chien, P.W.; Su, T.M.; Hsieh, D.J.; Lee, T.C. Characterization and spinal fusion effect of rabbit mesenchymal stem cells. BMC Res. Notes 2013, 6, 528. [Google Scholar] [CrossRef] [Green Version]
- Dennis, J.E.; Haynesworth, S.E.; Young, R.G.; Caplan, A.I. Osteogenesis in marrow-derived mesenchymal cell porous ceramic composites transplanted subcutaneously: Effect of fibronectin and laminin on cell retention and rate of osteogenic expression. Cell Transpl. 1992, 1, 23–32. [Google Scholar] [CrossRef]
- Alessio, N.; Stellavato, A.; Aprile, D.; Cimini, D.; Vassallo, V.; Di Bernardo, G.; Galderisi, U.; Schiraldi, C. Timely Supplementation of Hydrogels Containing Sulfated or Unsulfated Chondroitin and Hyaluronic Acid Affects Mesenchymal Stromal Cells Commitment Toward Chondrogenic Differentiation. Front. Cell Dev. Biol. 2021, 9, 641529. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Calle, A.; Barrajón-Masa, C.; Gómez-Fidalgo, E.; Martín-Lluch, M.; Cruz-Vigo, P.; Sánchez-Sánchez, R.; Ramírez, M.Á. Iberian pig mesenchymal stem/stromal cells from dermal skin, abdominal and subcutaneous adipose tissues, and peripheral blood: In vitro characterization and migratory properties in inflammation. Stem Cell Res. Ther. 2018, 9, 178. [Google Scholar] [CrossRef] [Green Version]
- Calle, A.; López-Martín, S.; Monguió-Tortajada, M.; Borràs, F.E.; Yáñez-Mó, M.; Ramírez, M.Á. Bovine endometrial MSC: Mesenchymal to epithelial transition during luteolysis and tropism to implantation niche for immunomodulation. Stem Cell Res. Ther. 2019, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Calle, A.; Gutiérrez-Reinoso, M.Á.; Re, M.; Blanco, J.; De la Fuente, J.; Monguió-Tortajada, M.; Borràs, F.E.; Yáñez-Mó, M.; Ramírez, M.Á. Bovine peripheral blood MSCs chemotax towards inflammation and embryo implantation stimuli. J. Cell Physiol. 2021, 236, 1054–1067. [Google Scholar] [CrossRef] [PubMed]
- Zomer, H.D.; Roballo, K.C.; Lessa, T.B.; Bressan, F.F.; Gonçalves, N.N.; Meirelles, F.V.; Trentin, A.G.; Ambrósio, C.E. Distinct features of rabbit and human adipose-derived mesenchymal stem cells: Implications for biotechnology and translational research. Stem Cells Cloning 2018, 11, 43–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Lorenzo, M.J.; Royo-Cañas, M.; Alegre-Aguarón, E.; Desportes, P.; Castiella, T.; García-Alvarez, F.; Larrad, L. Phenotype and chondrogenic differentiation of mesenchymal cells from adipose tissue of different species. J. Orthop. Res. 2009, 27, 1499–1507. [Google Scholar] [CrossRef]
- Lee, T.C.; Lee, T.H.; Huang, Y.H.; Chang, N.K.; Lin, Y.J.; Chien, P.W.; Yang, W.H.; Lin, M.H. Comparison of surface markers between human and rabbit mesenchymal stem cells. PLoS ONE 2014, 9, e111390. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Shen, M.; Chen, W.; Li, X.; Luo, D.; Cai, J.; Yang, Y. Isolation and purification of rabbit mesenchymal stem cells using an optimized protocol. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 1102–1108. [Google Scholar] [CrossRef]
- Yoon, D.S.; Kim, Y.H.; Jung, H.S.; Paik, S.; Lee, J.W. Importance of Sox2 in maintenance of cell proliferation and multipotency of mesenchymal stem cells in low-density culture. Cell Prolif. 2011, 44, 428–440. [Google Scholar] [CrossRef]
- Liu, T.M.; Wu, Y.N.; Guo, X.M.; Hui, J.H.; Lee, E.H.; Lim, B. Effects of ectopic Nanog and Oct4 overexpression on mesenchymal stem cells. Stem Cells Dev. 2009, 18, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Han, S.M.; Han, S.H.; Coh, Y.R.; Jang, G.; Chan Ra, J.; Kang, S.K.; Lee, H.W.; Youn, H.Y. Enhanced proliferation and differentiation of Oct4- and Sox2-overexpressing human adipose tissue mesenchymal stem cells. Exp. Mol. Med. 2014, 46, e101. [Google Scholar] [CrossRef]
- Tsai, C.C.; Su, P.F.; Huang, Y.F.; Yew, T.L.; Hung, S.C. Oct4 and Nanog directly regulate Dnmt1 to maintain self-renewal and undifferentiated state in mesenchymal stem cells. Mol. Cell. 2012, 47, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Ferrer-Lorente, R.; Bejar, M.T.; Badimon, L. Notch signaling pathway activation in normal and hyperglycemic rats differs in the stem cells of visceral and subcutaneous adipose tissue. Stem Cells Dev. 2014, 23, 3034–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchkonia, T.; Giorgadze, N.; Pirtskhalava, T.; Thomou, T.; DePonte, M.; Koo, A.; Forse, R.A.; Chinnappan, D.; Martin-Ruiz, C.; von Zglinicki, T.; et al. Fat depot-specific characteristics are retained in strains derived from single human preadipocytes. Diabetes 2006, 55, 2571–2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.T.; Kahn, C.R. Transplantation of adipose tissue and stem cells: Role in metabolism and disease. Nat. Rev. Endocrinol. 2010, 6, 195–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macotela, Y.; Emanuelli, B.; Mori, M.A.; Gesta, S.; Schulz, T.J.; Tseng, Y.H.; Kahn, C.R. Intrinsic differences in adipocyte precursor cells from different white fat depots. Diabetes 2012, 61, 1691–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gesta, S.; Blüher, M.; Yamamoto, Y.; Norris, A.W.; Berndt, J.; Kralisch, S.; Boucher, J.; Lewis, C.; Kahn, C.R. Evidence for a role of developmental genes in the origin of obesity and body fat distribution. Proc. Natl. Acad. Sci. USA 2006, 103, 6676–6681. [Google Scholar] [CrossRef] [Green Version]
- Tchkonia, T.; Lenburg, M.; Thomou, T.; Giorgadze, N.; Frampton, G.; Pirtskhalava, T.; Cartwright, A.; Cartwright, M.; Flanagan, J.; Karagiannides, I.; et al. Identification of depot-specific human fat cell progenitors through distinct expression profiles and developmental gene patterns. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E298–E307. [Google Scholar] [CrossRef] [Green Version]
- Fischer, B.; Chavatte-Palmer, P.; Viebahn, C.; Navarrete Santos, A.; Duranthon, V. Rabbit as a reproductive model for human health. Reproduction 2012, 144, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Jazedje, T.; Perin, P.M.; Czeresnia, C.E.; Maluf, M.; Halpern, S.; Secco, M.; Bueno, D.F.; Vieira, N.M.; Zucconi, E.; Zatz, M. Human fallopian tube: A new source of multipotent adult mesenchymal stem cells discarded in surgical procedures. J. Transl. Med. 2009, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, Y.; Chang, C.; Wu, S.; Gao, J.; Zhang, Y.; Chen, Y.; Zhong, F.; Deng, G. Human fallopian tube proteome shows high coverage of mesenchymal stem cells associated proteins. Biosci. Rep. 2016, 36, e00297. [Google Scholar] [CrossRef] [Green Version]
- Rahimzadeh, A.; Mirakabad, F.S.; Movassaghpour, A.; Shamsasenjan, K.; Kariminekoo, S.; Talebi, M.; Shekari, A.; Zeighamian, V.; Ghalhar, M.G.; Akbarzadeh, A. Biotechnological and biomedical applications of mesenchymal stem cells as a therapeutic system. Artif. Cells Nanomed. Biotechnol. 2016, 44, 559–570. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, Y.; Wu, X.; Yin, S.; Chuai, Y.; Wang, A. The utility of human fallopian tube mucosa as a novel source of multipotent stem cells for the treatment of autologous reproductive tract injury. Stem Cell Res. Ther. 2015, 6, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jazedje, T.; Bueno, D.F.; Almada, B.V.P.; Caetano, H.; Czeresnia, C.E.; Perin, P.M.; Halpern, S.; Maluf, M.; Evangelista, L.P.; Nisenbaum, M.G.; et al. Human fallopian tube mesenchymal stromal cells enhance bone regeneration in a xenotransplanted model. Stem Cell Rev. Rep. 2012, 8, 355–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shackleton, M.; Vaillant, F.; Simpson, K.J.; Stingl, J.; Smyth, G.K.; Asselin-Labat, M.L.; Wu, L.; Lindeman, G.J.; Visvader, J.E. Generation of a functional mammary gland from a single stem cell. Nature 2006, 439, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Dalerba, P.; Dylla, S.J.; Park, I.-K.; Liu, R.; Wang, X.; Cho, R.W.; Hoey, T.; Gurney, A.; Huang, E.H.; Simeone, D.M.; et al. Phenotypic characterization of human colorectal cancer stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 10158–10163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Heidt, D.G.; Dalerba, P.; Burant, C.F.; Zhang, L.; Adsay, V.; Wicha, M.; Clarke, M.F.; Simeone, D.M. Identification of pancreatic cancer stem cells. Cancer Res. 2007, 67, 1030–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, C.A.; Visvader, J.E. The Cellular Organization of the Mammary Gland: Insights From Microscopy. J. Mammary Gland Biol. Neoplasia. 2021, 26, 71–85. [Google Scholar] [CrossRef]
- Song, J.; Ding, W.; Liu, Y.; Lin, L.; Jia, M.; Liu, S.; Xue, K. The mesenchymal property of mouse mammary anlagen repopulating cell population is associated with its stemness. Gene Expr. Patterns 2020, 38, 119151. [Google Scholar] [CrossRef]
- Joshi, P.A.; Waterhouse, P.D.; Kasaian, K.; Fang, H.; Gulyaeva, O.; Sul, H.S.; Boutros, P.C.; Khokha, R. PDGFRα+ stromal adipocyte progenitors transition into epithelial cells during lobulo-alveologenesis in the murine mammary gland. Nat. Commun. 2019, 10, 1760. [Google Scholar] [CrossRef] [Green Version]
- Guest, I.; Ilic, Z.; Ma, J. Preparation of epithelial and mesenchymal stem cells from murine mammary gland. Curr. Protoc. Toxicol. 2011. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calle, A.; Zamora-Ceballos, M.; Bárcena, J.; Blanco, E.; Ramírez, M.Á. Comparison of Biological Features of Wild European Rabbit Mesenchymal Stem Cells Derived from Different Tissues. Int. J. Mol. Sci. 2022, 23, 6420. https://doi.org/10.3390/ijms23126420
Calle A, Zamora-Ceballos M, Bárcena J, Blanco E, Ramírez MÁ. Comparison of Biological Features of Wild European Rabbit Mesenchymal Stem Cells Derived from Different Tissues. International Journal of Molecular Sciences. 2022; 23(12):6420. https://doi.org/10.3390/ijms23126420
Chicago/Turabian StyleCalle, Alexandra, María Zamora-Ceballos, Juan Bárcena, Esther Blanco, and Miguel Ángel Ramírez. 2022. "Comparison of Biological Features of Wild European Rabbit Mesenchymal Stem Cells Derived from Different Tissues" International Journal of Molecular Sciences 23, no. 12: 6420. https://doi.org/10.3390/ijms23126420
APA StyleCalle, A., Zamora-Ceballos, M., Bárcena, J., Blanco, E., & Ramírez, M. Á. (2022). Comparison of Biological Features of Wild European Rabbit Mesenchymal Stem Cells Derived from Different Tissues. International Journal of Molecular Sciences, 23(12), 6420. https://doi.org/10.3390/ijms23126420