
Transcriptome and Metabolome Analysis Provide New Insights into the Process of Tuberization of Sechium edule Roots

 , , ,
, , ,  ,
,
Abstract
:
1. Introduction
2. Results
2.1. Morphological Characteristics of S. edule cv. ‘Tuershao’
2.2. Starch Content and Soil Physical Properties during Tuber Formation
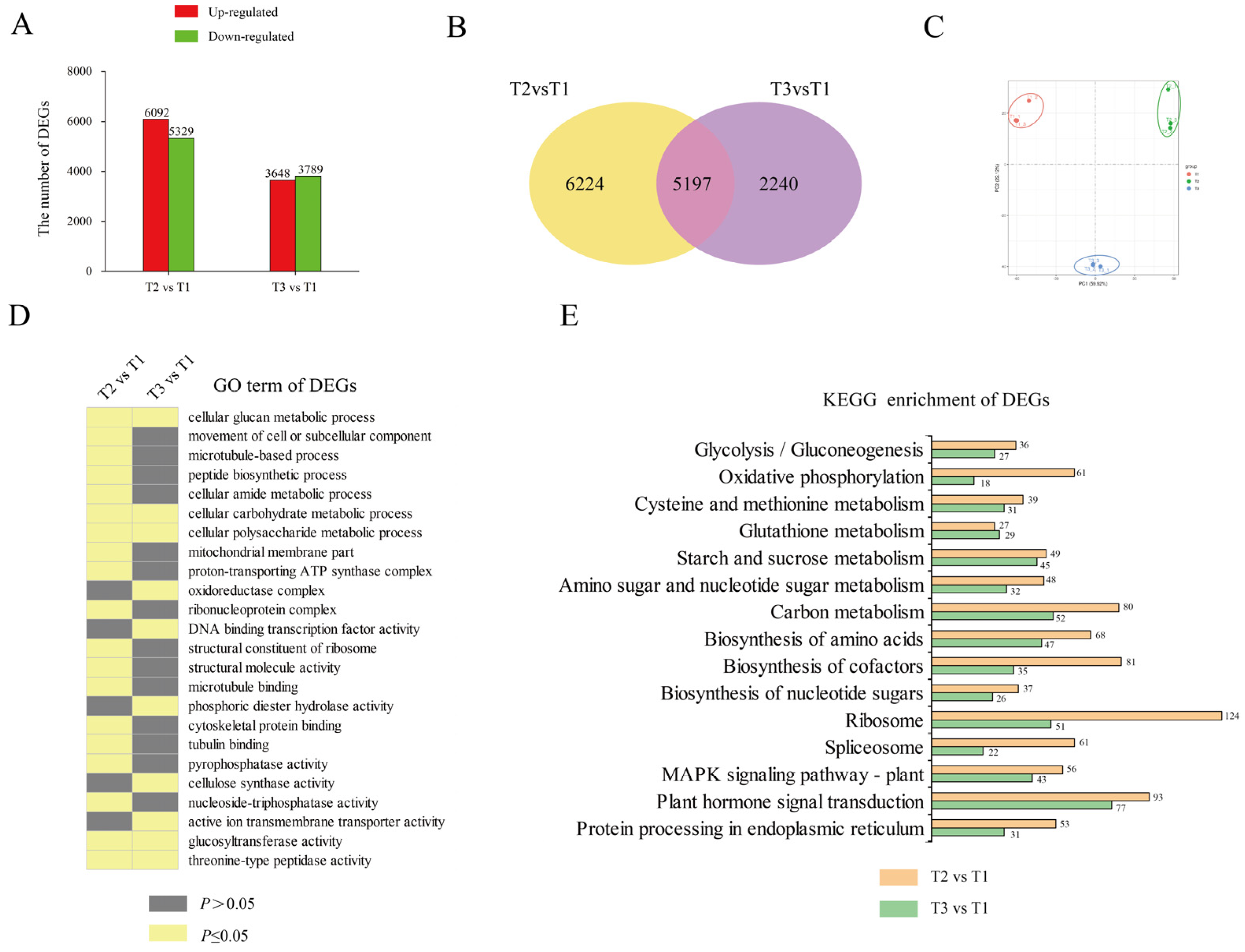
2.3. Transcriptome Characterization during Tuberization
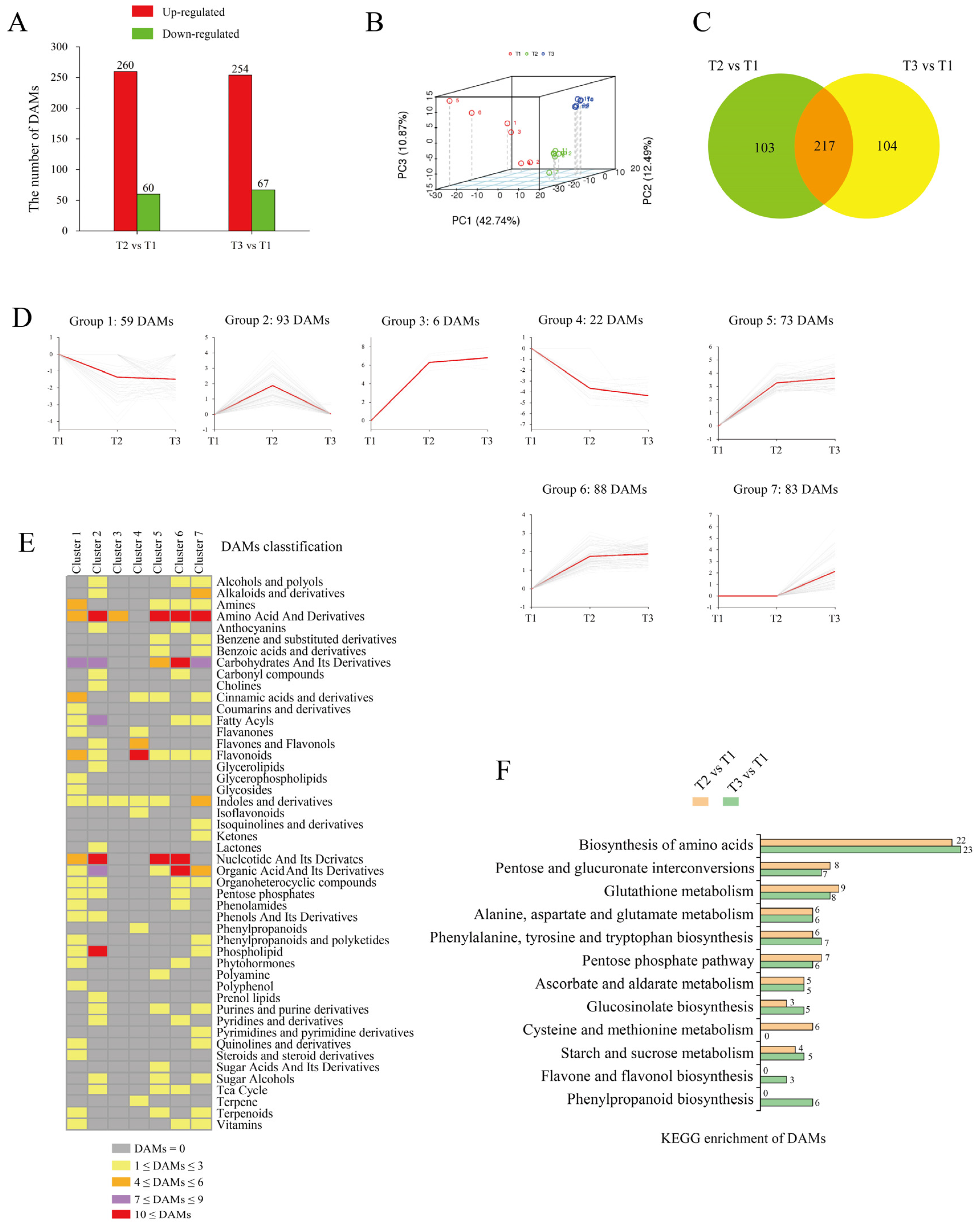
2.4. Characterization of the Metabolome of Tuberization
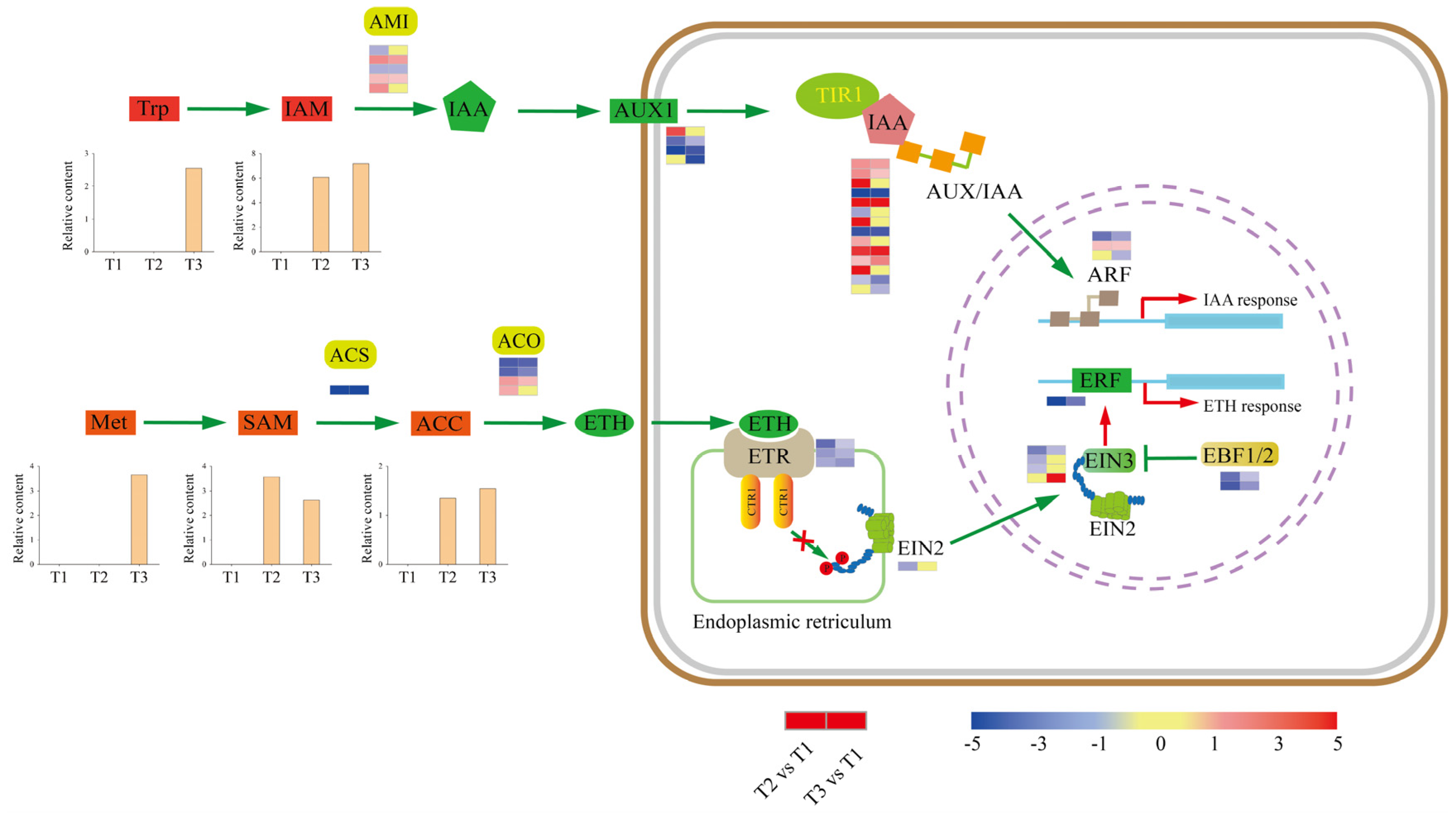
2.5. Plant-Hormone Pathways Response to Tuberization
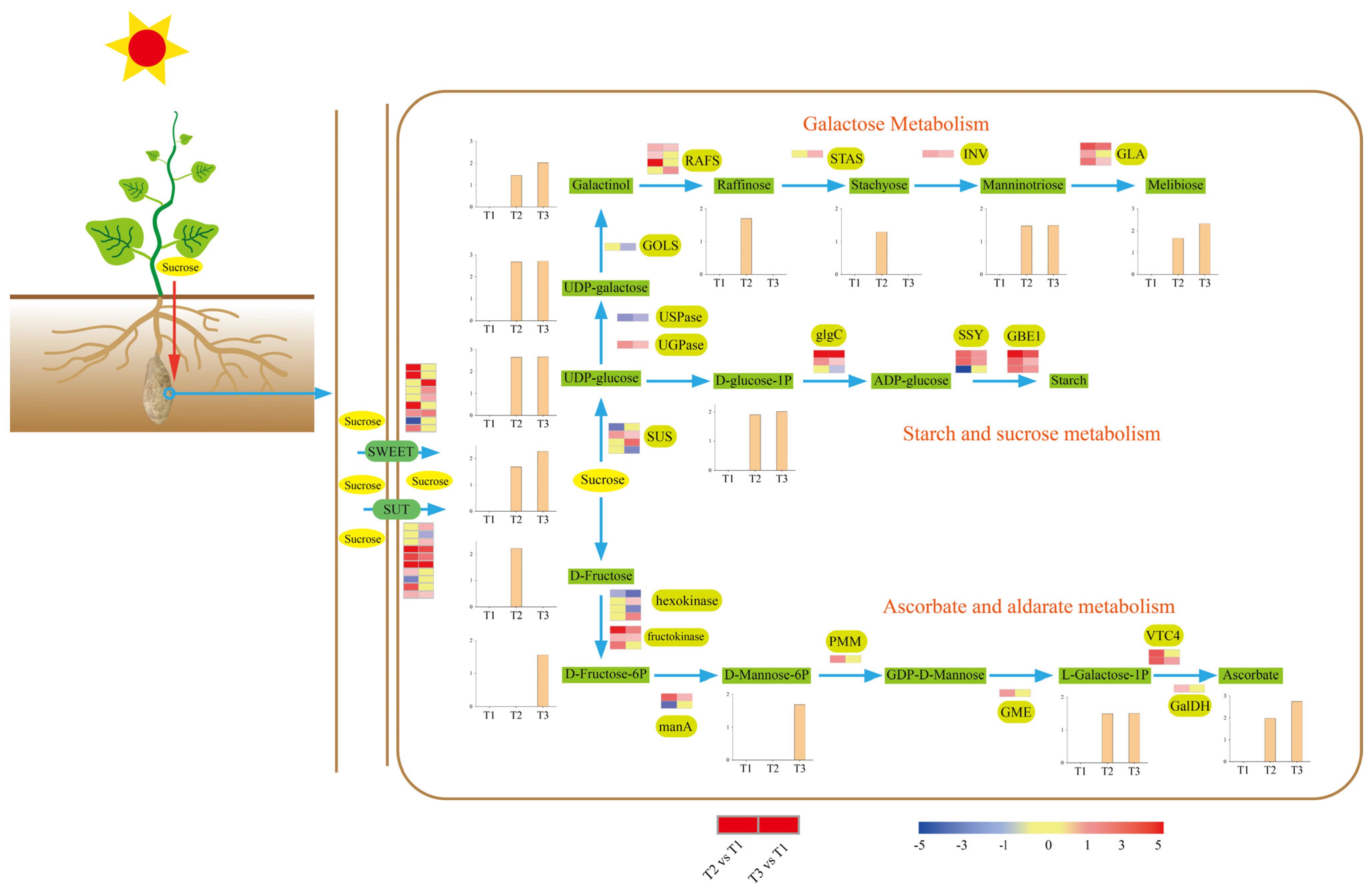
2.6. Sucrose Metabolism during Tuberization
2.7. Identification of Transcription Factors during Tuber Formation
2.8. Signal Network Regulating Tuberization
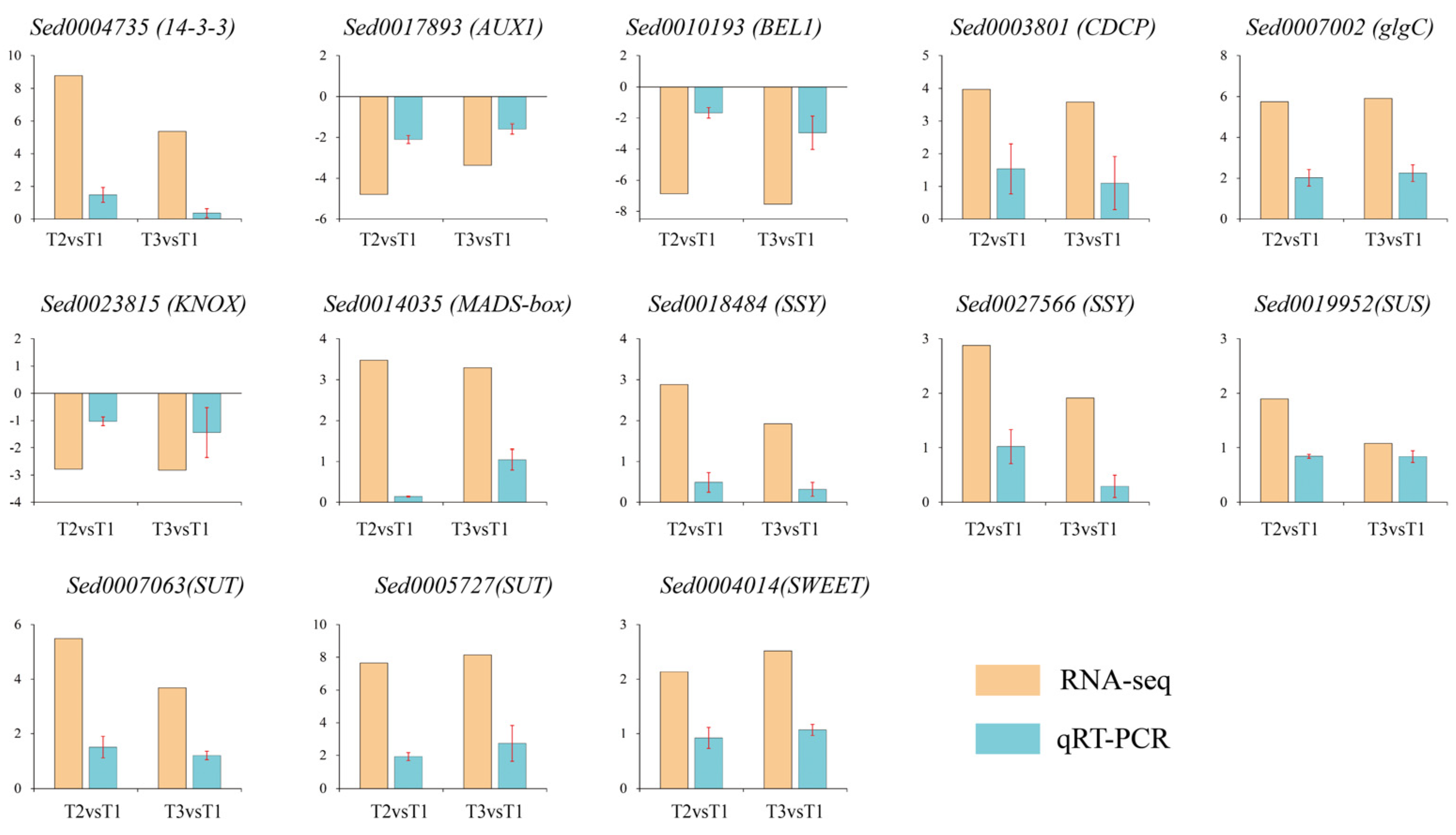
2.9. Validation of DEGs by qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Plant Material, Sampling, and Soil Physical Properties
4.2. Microscopy of Tuber and Root
4.3. Starch Content Detection
4.4. RNA-Seq Analysis
4.4.1. RNA Quantification and Library Preparation
4.4.2. Clustering and Sequencing (Novogene Experimental Department)
4.4.3. Quality Control and Transcriptome Assembly
4.4.4. Gene Functional Annotation and Differential Gene-Expression Analysis
4.5. Metabolome Analysis
4.5.1. Metabolite Extraction
4.5.2. Metabolite Identification and Quantification
4.5.3. Data Analysis
4.6. K-Mean Cluster Analysis
4.7. qRT-PCR Validation
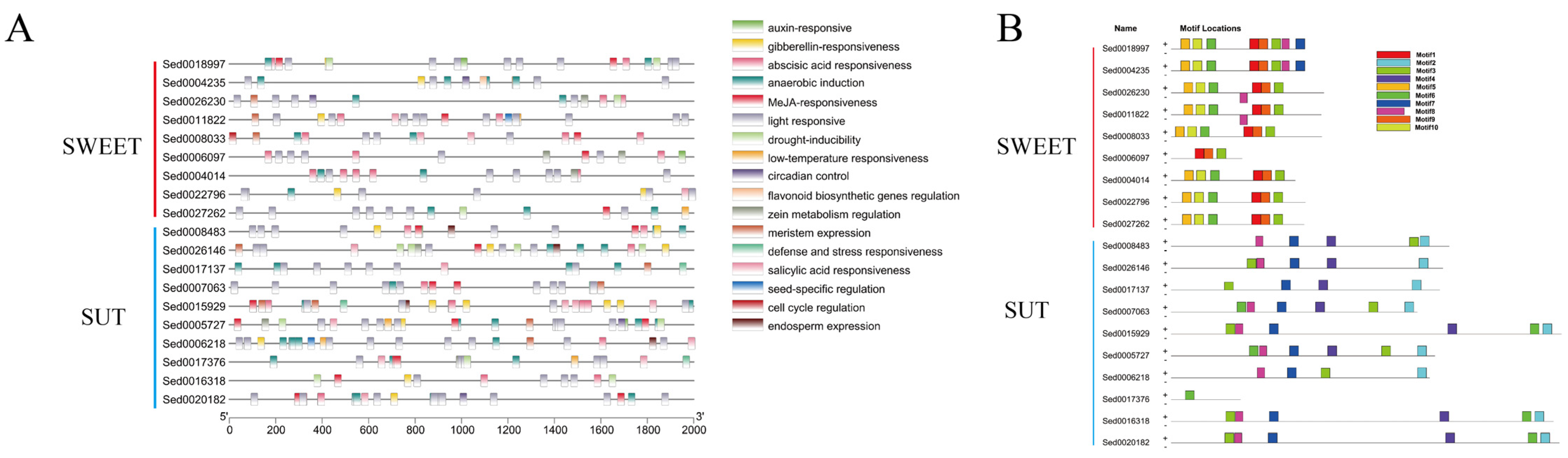
4.8. Cis-Element Analysis and Conserved Motif Analysis of DEGs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdelnour, A.; Rocha, O.J. Genetic characterization of a collection of chayote, Sechium edule (Jacq.) Swartz, in Costa Rica by using isozyme markers. Genet. Resour. Crop Evol. 2008, 55, 163–170. [Google Scholar] [CrossRef]
- Guo, J.; Xu, W.; Hu, Y.; Huang, J.; Zhao, Y.; Zhang, L.; Huang, C.-H.; Ma, H. Phylotranscriptomics in Cucurbitaceae Reveal Multiple Whole-Genome Duplications and Key Morphological and Molecular Innovations. Mol. Plant 2020, 13, 1117–1133. [Google Scholar] [CrossRef] [PubMed]
- Cadena-Iñiguez, J.; Arévalo-Galarza, L.; Avendaño-Arrazate, C.H.; Soto-Hernández, M.; del Mar Ruiz-Posadas, L.; Santiago-Osorio, E.; Acosta-Ramos, M.; Cisneros-Solano, V.M.; Aguirre-Medina, J.F.; Ochoa-Martinez, D. Production, genetics, postharvest management and pharmacological characteristics of Sechium edule (Jacq.) Sw. Fresh Prod. 2007, 1, 41–53. [Google Scholar]
- Siciliano, T.; De Tommasi, N.; Morelli, A.I.; Braca, A. Study of Flavonoids of Sechium edule (Jacq) Swartz (Cucurbitaceae) Different Edible Organs by Liquid Chromatography Photodiode Array Mass Spectrometry. J. Agric. Food Chem. 2004, 52, 6510–6515. [Google Scholar] [CrossRef] [PubMed]
- Shiga, T.M.; Peroni-Okita, F.H.G.; Carpita, N.C.; Lajolo, F.M.; Cordenunsi, B.R. Polysaccharide composition of raw and cooked chayote (Sechium edule Sw.) fruits and tuberous roots. Carbohydr. Polym. 2015, 130, 155–165. [Google Scholar] [CrossRef]
- Fidrianny, I.; Darmawati, A.; Hartati, R. Antioxidant capacities, phenolic, flavonoid and carotenoid content of various polarities extracts from three organs of Sechium edule (Jacq.) Swartz. J. Chem. Pharm. Res. 2015, 7, 914–920. [Google Scholar]
- Sriwichai, W.; Berger, J.; Picq, C.; Avallone, S. Determining Factors of Lipophilic Micronutrient Bioaccessibility in Several Leafy Vegetables. J. Agric. Food Chem. 2016, 64, 1695–1701. [Google Scholar] [CrossRef]
- Ke, J.; Jiang, G.; Shen, G.; Wu, H.; Liu, Y.; Zhang, Z. Optimization, characterization and rheological behavior study of pectin extracted from chayote (Sechium edule) using ultrasound assisted method. Int. J. Biol. Macromol. 2020, 147, 688–698. [Google Scholar] [CrossRef]
- Vieira, E.F.; Pinho, O.; Ferreira, I.M.; Delerue-Matos, C. Chayote (Sechium edule): A review of nutritional composition, bioactivities and potential applications. Food Chem. 2019, 275, 557–568. [Google Scholar] [CrossRef]
- Noumedem, J.A.K.; Mihasan, M.; Lacmata, S.T.; Stefan, M.; Kuiate, J.R.; Kuete, V. Antibacterial activities of the methanol extracts of ten Cameroonian vegetables against Gram-negative multidrug-resistant bacteria. BMC Complement. Altern. Med. 2013, 13, 26. [Google Scholar] [CrossRef] [Green Version]
- Firdous, S.; Sravanthi, K.; Debnath, R.; Neeraja, K. Protective effect of ethanolic extract and its ethylacetate and n-butanol fractions of Sechium edule fruits against carbon tetrachloride induced hepatic injury in rats. Int. J. Pharm. Pharm. Sci. 2012, 4, 354–359. [Google Scholar]
- Wu, C.-H.; Ou, T.-T.; Chang, C.-H.; Chang, X.-Z.; Yang, M.-Y.; Wang, C.-J. The Polyphenol Extract from Sechium edule Shoots Inhibits Lipogenesis and Stimulates Lipolysis via Activation of AMPK Signals in HepG2 Cells. J. Agric. Food Chem. 2014, 62, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-Y.; Chan, K.-C.; Lee, Y.-J.; Chang, X.-Z.; Wu, C.-H.; Wang, C.-J. Sechium edule Shoot Extracts and Active Components Improve Obesity and a Fatty Liver That Involved Reducing Hepatic Lipogenesis and Adipogenesis in High-Fat-Diet-Fed Rats. J. Agric. Food Chem. 2015, 63, 4587–4596. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Uribe, J.P.; García-Suárez, F.J.; Gutiérrez-Meraz, F.; Rodriguez-Ambriz, S.L.; Bello-Perez, L.A. By-products derived of the starch isolation from tubers: Physicochemical and functional properties. J. Food. Agric. Environ. 2014, 12, 43–46. [Google Scholar]
- Jiménez-Hernández, J.; Salazar-Montoya, J.A.; Ramos-Ramírez, E. Physical, chemical and microscopic characterization of a new starch from chayote (Sechium edule) tuber and its comparison with potato and maize starches. Carbohydr. Polym. 2007, 68, 679–686. [Google Scholar] [CrossRef]
- Fu, A.; Wang, Q.; Mu, J.; Ma, L.; Wen, C.; Zhao, X.; Gao, L.; Li, J.; Shi, K.; Wang, Y.; et al. Combined genomic, transcriptomic, and metabolomic analyses provide insights into chayote (Sechium edule) evolution and fruit development. Hortic. Res. 2021, 8, 35. [Google Scholar] [CrossRef]
- Cui, H.; Zhu, Z.; Lu, Z.; Ding, Z.; Zhang, C.; Luan, F. The complete chloroplast genome sequence of the Sechium edule (Jacq.) Swartz. (Cucurbitaceae). Mitochondrial DNA Part B 2021, 6, 97–98. [Google Scholar] [CrossRef]
- Zierer, W.; Rüscher, D.; Sonnewald, U.; Sonnewald, S. Tuber and Tuberous Root Development. Annu. Rev. Plant Biol. 2021, 72, 551–580. [Google Scholar] [CrossRef]
- Navarro, C.; Abelenda, J.A.; Cruz-Oró, E.; Cuéllar, C.A.; Tamaki, S.; Silva, J.; Shimamoto, K.; Prat, S. Control of flowering and storage organ formation in potato by FLOWERING LOCUS T. Nature 2011, 478, 119–122. [Google Scholar] [CrossRef]
- Teo, C.-J.; Takahashi, K.; Shimizu, K.; Shimamoto, K.; Taoka, K.-I. Potato Tuber Induction is Regulated by Interactions Between Components of a Tuberigen Complex. Plant Cell Physiol. 2017, 58, 365–374. [Google Scholar] [CrossRef]
- Lin, T.; Sharma, P.; Gonzalez, D.H.; Viola, I.L.; Hannapel, D.J. The Impact of the Long-Distance Transport of aBEL1-Like Messenger RNA on Development. Plant Physiol. 2013, 161, 760–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.K.; Chatterjee, M.; Yu, Y.; Suh, S.-G.; Miller, W.A.; Hannapel, D.J. Dynamics of a Mobile RNA of Potato Involved in a Long-Distance Signaling Pathway. Plant Cell 2006, 18, 3443–3457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi, V.; Naskar, S.K.; Makeshkumar, T.; Babu, B.; Prakash Krishnan, B.S. Molecular Physiology of Storage Root Formation and Development in Sweet Potato (Ipomoea batatas (L.) Lam.). J. Root Tuber Crops 2009, 35, 1–27. [Google Scholar]
- Eserman, L.A.; Jarret, R.L.; Leebens-Mack, J.H. Parallel evolution of storage roots in morning glories (Convolvulaceae). BMC Plant Biol. 2018, 18, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Yang, C.; Zhu, J.; Zhang, L.; Bai, Y.; Li, E.; Gilbert, R.G. Competition between Granule Bound Starch Synthase and Starch Branching Enzyme in Starch Biosynthesis. Rice 2019, 12, 96. [Google Scholar] [CrossRef] [Green Version]
- Prathap, V.; Tyagi, A. Correlation between expression and activity of ADP glucose pyrophosphorylase and starch synthase and their role in starch accumulation during grain filling under drought stress in rice. Plant Physiol. Biochem. 2020, 157, 239–243. [Google Scholar] [CrossRef]
- Patrick, J.W.; Botha, F.C.; Birch, R.G. Metabolic engineering of sugars and simple sugar derivatives in plants. Plant Biotechnol. J. 2013, 11, 142–156. [Google Scholar] [CrossRef]
- Zhang, K.; Wu, Z.; Tang, D.; Luo, K.; Lu, H.; Liu, Y.; Dong, J.; Wang, X.; Lv, C.; Wang, J.; et al. Comparative Transcriptome Analysis Reveals Critical Function of Sucrose Metabolism Related-Enzymes in Starch Accumulation in the Storage Root of Sweet Potato. Front. Plant Sci. 2017, 8, 914. [Google Scholar] [CrossRef]
- El-Ganiny, A.M.; Sheoran, I.; Sanders, D.A.; Kaminskyj, S.G. Aspergillus nidulans UDP-glucose-4-epimerase UgeA has multiple roles in wall architecture, hyphal morphogenesis, and asexual development. Fungal Genet. Biol. 2010, 47, 629–635. [Google Scholar] [CrossRef]
- Li, D.; Flores-Sandoval, E.; Ahtesham, U.; Coleman, A.; Clay, J.M.; Bowman, J.L.; Chang, C. Ethylene-independent functions of the ethylene precursor ACC in Marchantia polymorpha. Nat. Plants 2020, 6, 1335–1344. [Google Scholar] [CrossRef]
- Dos Santos, T.B.; Budzinski, I.G.; Marur, C.J.; Petkowicz, C.L.; Pereira, L.F.P.; Vieira, L.G. Expression of three galactinol synthase isoforms in Coffea arabica L. and accumulation of raffinose and stachyose in response to abiotic stresses. Plant Physiol. Biochem. 2011, 49, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Megías-Pérez, R.; Hahn, C.; Ruiz-Matute, A.I.; Behrends, B.; Albach, D.C.; Kuhnert, N. Changes in low molecular weight carbohydrates in kale during development and acclimation to cold temperatures determined by chromatographic techniques coupled to mass spectrometry. Food Res. Int. 2020, 127, 108727. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Song, X.; Bartels, D. Sugar metabolism in the desiccation tolerant grass Oropetium thomaeum in response to environmental stresses. Plant Sci. 2018, 270, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Eom, J.-S.; Chen, L.-Q.; Sosso, D.; Julius, B.T.; Lin, I.; Qu, X.-Q.; Braun, D.M.; Frommer, W.B. SWEETs, transporters for intracellular and intercellular sugar translocation. Curr. Opin. Plant Biol. 2015, 25, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-Q.; Qu, X.-Q.; Hou, B.-H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose Efflux Mediated by SWEET Proteins as a Key Step for Phloem Transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef]
- Sosso, D.; Luo, D.; Li, Q.-B.; Sasse, J.; Yang, J.; Gendrot, G.; Suzuki, M.; E Koch, K.; Mccarty, D.R.; Chourey, P.S.; et al. Seed filling in domesticated maize and rice depends on SWEET-mediated hexose transport. Nat. Genet. 2015, 47, 1489–1493. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, D.; Miao, Q.; Yang, J.; Xuan, Y.; Hu, Y. Essential Role of Sugar Transporter OsSWEET11 During the Early Stage of Rice Grain Filling. Plant Cell Physiol. 2017, 58, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Fei, H.; Yang, Z.; Lu, Q.; Wen, X.; Zhang, Y.; Zhang, A.; Lu, C. OsSWEET14 cooperates with OsSWEET11 to contribute to grain filling in rice. Plant Sci. 2021, 306, 110851. [Google Scholar] [CrossRef]
- Yang, J.; Luo, D.; Yang, B.; Frommer, W.B.; Eom, J. SWEET 11 and 15 as key players in seed filling in rice. New Phytol. 2018, 218, 604–615. [Google Scholar] [CrossRef] [Green Version]
- Kühn, C.; Hajirezaei, M.-R.; Fernie, A.R.; Roessner-Tunali, U.; Czechowski, T.; Hirner, B.; Frommer, W.B. The Sucrose Transporter StSUT1 Localizes to Sieve Elements in Potato Tuber Phloem and Influences Tuber Physiology and Development. Plant Physiol. 2003, 131, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Chincinska, I.A.; Liesche, J.; Krügel, U.; Michalska, J.; Geigenberger, P.; Grimm, B.; Kuhn, C. Sucrose Transporter StSUT4 from Potato Affects Flowering, Tuberization, and Shade Avoidance Response. Plant Physiol. 2007, 146, 323–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roumeliotis, E.; Visser, R.G.F.; Bachem, C.W. A crosstalk of auxin and GA during tuber development. Plant Signal. Behav. 2012, 7, 1360–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondhare, K.R.; Patil, A.B.; Giri, A.P. Auxin: An emerging regulator of tuber and storage root development. Plant Sci. 2021, 306, 110854. [Google Scholar] [CrossRef] [PubMed]
- Roumeliotis, E.; Kloosterman, B.; Oortwijn, M.; Kohlen, W.; Bouwmeester, H.J.; Visser, R.G.; Bachem, C.W. The effects of auxin and strigolactones on tuber initiation and stolon architecture in potato. J. Exp. Bot. 2012, 63, 4539–4547. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Zhu, M.; Yu, J.; Han, R.; Tang, C.; Xu, T.; Liu, J.; Li, Z. RNA-Seq and iTRAQ reveal multiple pathways involved in storage root formation and development in sweet potato (Ipomoea batatas L.). BMC Plant Biol. 2019, 19, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakehal, A.; Chaabouni, S.; Cavel, E.; Le Hir, R.; Ranjan, A.; Raneshan, Z.; Novák, O.; Păcurar, D.I.; Perrone, I.; Jobert, F.; et al. A Molecular Framework for the Control of Adventitious Rooting by TIR1/AFB2-Aux/IAA-Dependent Auxin Signaling in Arabidopsis. Mol. Plant 2019, 12, 1499–1514. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, Y.; Liu, Y.; Li, X.; Hao, G.; Han, Q.; Dirk, L.M.A.; Downie, A.B.; Ruan, Y.-L.; Wang, J.; et al. Raffinose synthase enhances drought tolerance through raffinose synthesis or galactinol hydrolysis in maize and Arabidopsis plants. J. Biol. Chem. 2020, 295, 8064–8077. [Google Scholar] [CrossRef]
- Vanderstraeten, L.; Depaepe, T.; Bertrand, S.; Van Der Straeten, D. The Ethylene Precursor ACC Affects Early Vegetative Development Independently of Ethylene Signaling. Front. Plant Sci. 2019, 10, 1591. [Google Scholar] [CrossRef] [Green Version]
- Kendrick, M.D.; Chang, C. Ethylene signaling: New levels of complexity and regulation. Curr. Opin. Plant Biol. 2008, 11, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Y.-L.; Patrick, J.W.; Bouzayen, M.; Osorio, S.; Fernie, A.R. Molecular regulation of seed and fruit set. Trends Plant Sci. 2012, 17, 656–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Muhammad, N.; Yin, K.; Liu, Z.; Wang, L.; Luo, Z.; Wang, L.; Liu, M. Molecular regulation of fruit size in horticultural plants: A review. Sci. Hortic. 2021, 288, 110353. [Google Scholar] [CrossRef]
- Ku, A.T.; Huang, Y.-S.; Wang, Y.-S.; Ma, D.F.; Yeh, K.-W. IbMADS1 (Ipomoea batatas MADS-box 1 gene) is Involved in Tuberous Root Initiation in Sweet Potato (Ipomoea batatas). Ann. Bot. 2008, 102, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-H.; Mizuno, K.; Fujimura, T. Isolation of MADS-box genes from sweet potato (Ipomoea batatas (L.) Lam.) expressed specifically in vegetative tissues. Plant Cell Physiol. 2002, 43, 314–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, S.A.; Lee, H.-S.; Huh, E.J.; Huh, G.H.; Paek, K.-H.; Shin, J.S.; Bae, J.M. SRD1 is involved in the auxin-mediated initial thickening growth of storage root by enhancing proliferation of metaxylem and cambium cells in sweetpotato (Ipomoea batatas). J. Exp. Bot. 2010, 61, 1337–1349. [Google Scholar] [CrossRef]
- Kim, J.-S.; Jang, M.; Nie, H.; Lee, J.; Hong, E.; Kim, S.-J.; Kim, S.H. Differential expression pattern of novel MADS-box genes in early root formation and differentiation of sweet potato. Gene Expr. Patterns 2021, 43, 119216. [Google Scholar] [CrossRef]
- Xiao, Q.; Wang, Y.; Du, J.; Li, H.; Wei, B.; Wang, Y.; Li, Y.; Yu, G.; Liu, H.; Zhang, J.; et al. ZmMYB14 is an important transcription factor involved in the regulation of the activity of the ZmBT1 promoter in starch biosynthesis in maize. FEBS J. 2017, 284, 3079–3099. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Q.; Wang, Y.; Li, H.; Zhang, C.; Wei, B.; Wang, Y.; Huang, H.; Li, Y.; Yu, G.; Liu, H.; et al. Transcription factor ZmNAC126 plays an important role in transcriptional regulation of maize starch synthesis-related genes. Crop J. 2020, 9, 192–203. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Yi, Q.; Hu, Y.; Liu, H.; Liu, Y.; Huang, Y. Novel role of ZmaNAC36 in co-expression of starch synthetic genes in maize endosperm. Plant Mol. Biol. 2013, 84, 359–369. [Google Scholar] [CrossRef]
- Chen, T.-K.; Yang, H.-T.; Fang, S.-C.; Lien, Y.-C.; Yang, T.-T.; Ko, S.-S. Hybrid-Cut: An Improved Sectioning Method for Recalcitrant Plant Tissue Samples. J. Vis. Exp. 2016, e54754. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, Z.-Y.; Zhao, Q.; Lu, C.; Zhang, Q.; Li, L.; Liu, S.; Wang, S.-Q.; Wang, Z.-Z.; Niu, J.-F. Genome-Wide Analysis and the Expression Pattern of the MADS-Box Gene Family in Bletilla striata. Plants 2021, 10, 2184. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Sequence | Function Describe | Gene ID | Sequence | Function Describe |
|---|---|---|---|---|---|
| Sed0004735 | GGCGTATCATGTCCTCGATT | 14-3-3 | Sed0005727 | TTAGGTTGCTTTCTAGCAGGAG | SUT |
| GGGATGAGGTGCTTGTCAAT | AGTTGAAGGTGTAGGAAACAGC | ||||
| Sed0023815 | AGATTGCGGATTTGTTGGAC | KNOX | Sed0018997 | AAGACGAAGAGCGTAGAGTTCA | SWEET |
| AGGCCTTTTGAGATCCGATT | GCGATCTCGGTAGATGAAGTAG | ||||
| Sed0014035 | CAAGCTGGAGAAGGATCTGC | MADS-box | Sed0018484 | AGCTCTACAAAGGAAGGGAAAC | SSY |
| CAAAAGGTGACTTGGCGATT | CTCCACTTCAATCTCTCGTAGC | ||||
| Sed0000607 | TGCTGAACTCTGCTGTTGCT | WRKY | Sed0027566 | TGTAGAAACGATGGAGAGAGGT | SSY |
| GGCTGAGGTTGTTCATGGTT | GAAGACGACTCGAGCTTTAGAA | ||||
| Sed0017893 | GGTGAAGAACAGCTGCAACA | AUX1 | Sed0019952 | CTAGGCCAGGACAATATGAAAG | SUS |
| CTTCCCATCAGCCCATAGAA | CCTATCTGCAAGGAAACCTATG | ||||
| Sed0001824 | CTCAATCCTAAAGCCAACAGAG | Actin | Sed0007002 | AAGACGCCTTTCTACACATCTC | glgC |
| AGTGTGACTAACACCATCACCA | TCAGTTTGGTAACTGTCTGCAC | ||||
| Sed0004014 | GACCTTCCTCTTGAGCCTATCT | SWEET | Sed0010193 | GTCCTACAACAACCACTCAGGT | BEL1 |
| ATTGTACTATCCCCAACACAGC | ATGCTGTTGTAACCCTAACGTC | ||||
| Sed0007063 | GAGGATATACTTGCGCTCAGTC | SUT | Sed0003801 | AGTGAGAAGCGCCTATATCTTG | CDCP |
| ACCAATGCCTGTTACTATGACC | GTGCGACGGTCTATGAGTAGAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, L.; Cheng, S.; Liu, Y.; Xie, Y.; He, Z.; Jia, M.; Zhou, X.; Zhang, R.; Li, C. Transcriptome and Metabolome Analysis Provide New Insights into the Process of Tuberization of Sechium edule Roots. Int. J. Mol. Sci. 2022, 23, 6390. https://doi.org/10.3390/ijms23126390
Su L, Cheng S, Liu Y, Xie Y, He Z, Jia M, Zhou X, Zhang R, Li C. Transcriptome and Metabolome Analysis Provide New Insights into the Process of Tuberization of Sechium edule Roots. International Journal of Molecular Sciences. 2022; 23(12):6390. https://doi.org/10.3390/ijms23126390
Chicago/Turabian StyleSu, Lihong, Shaobo Cheng, Yuhang Liu, Yongdong Xie, Zhongqun He, Mingyue Jia, Xiaoting Zhou, Ruijie Zhang, and Chunyan Li. 2022. "Transcriptome and Metabolome Analysis Provide New Insights into the Process of Tuberization of Sechium edule Roots" International Journal of Molecular Sciences 23, no. 12: 6390. https://doi.org/10.3390/ijms23126390
APA StyleSu, L., Cheng, S., Liu, Y., Xie, Y., He, Z., Jia, M., Zhou, X., Zhang, R., & Li, C. (2022). Transcriptome and Metabolome Analysis Provide New Insights into the Process of Tuberization of Sechium edule Roots. International Journal of Molecular Sciences, 23(12), 6390. https://doi.org/10.3390/ijms23126390