
Prophylactic Evidence of MSCs-Derived Exosomes in Doxorubicin/Trastuzumab-Induced Cardiotoxicity: Beyond Mechanistic Target of NRG-1/Erb Signaling Pathway

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
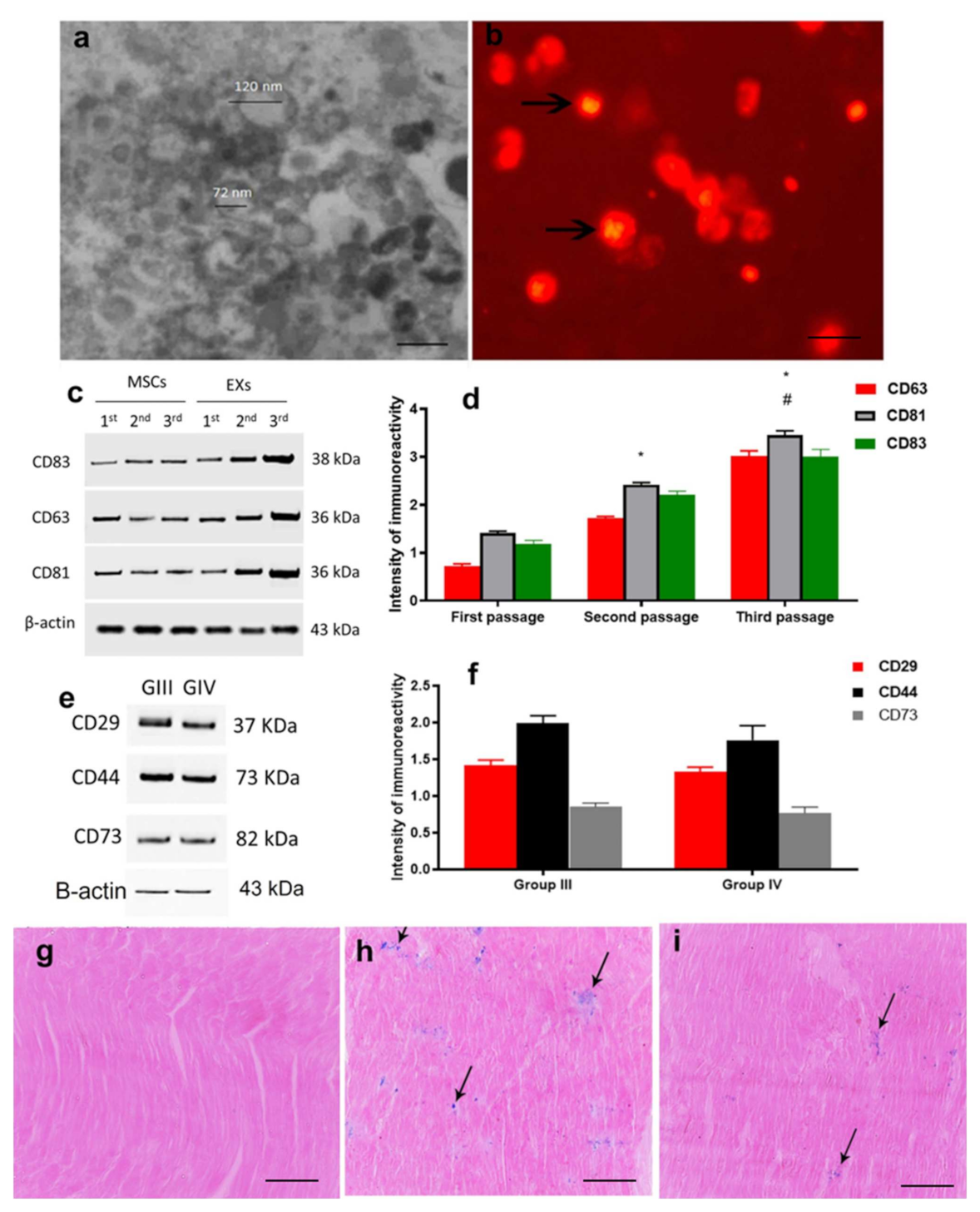
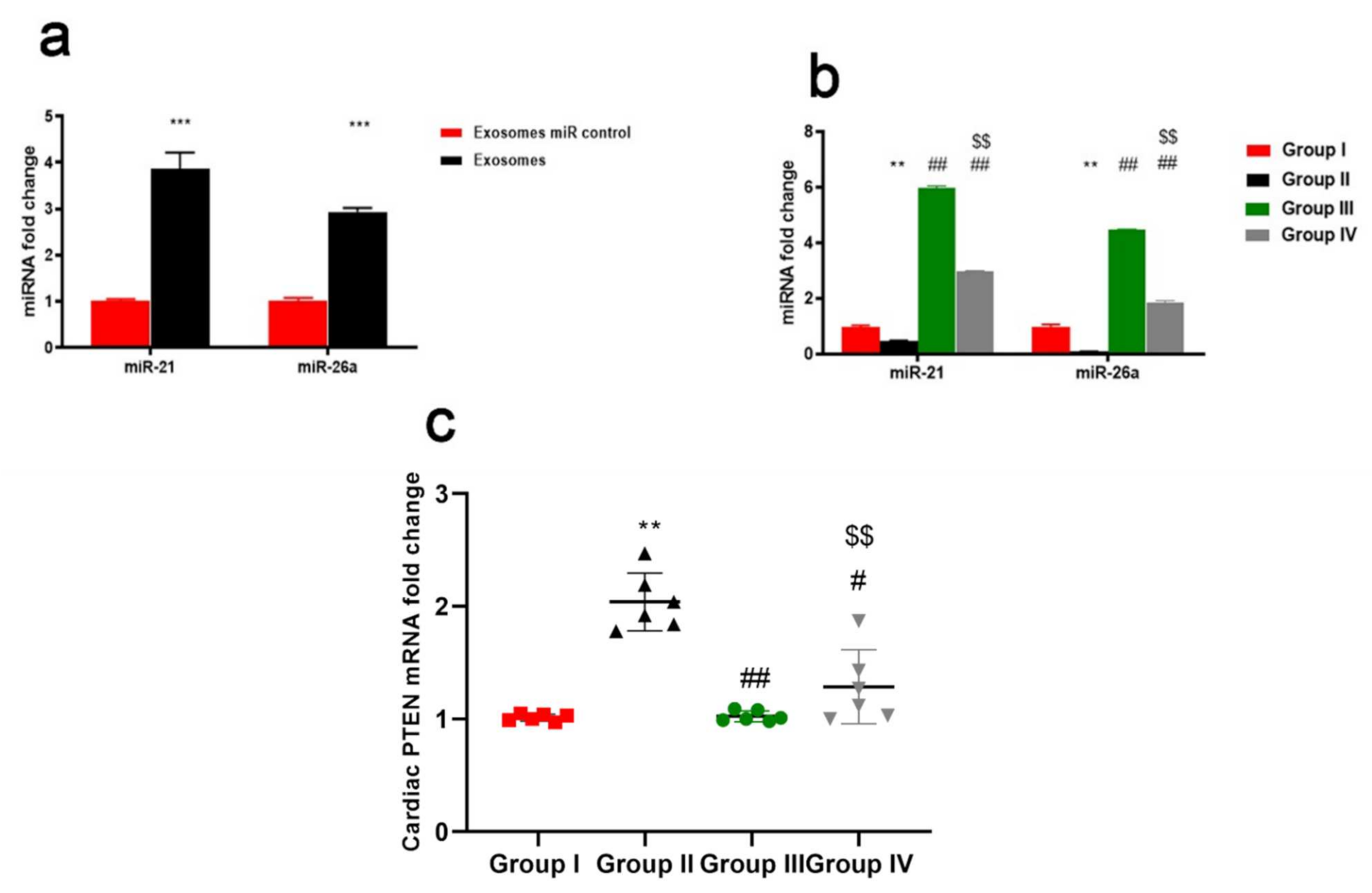
2.1. Characterization and Identification of Exosomes
2.2. ADMSCs-Derived Exosomes Restored DOX/Trz-Induced Alteration in Cardiac Function and Serum Cardiotoxicity Indices
2.3. ADMSCs-Derived Exosomes Ameliorated DOX/Trz-Induced Cardiac Tissue Injury
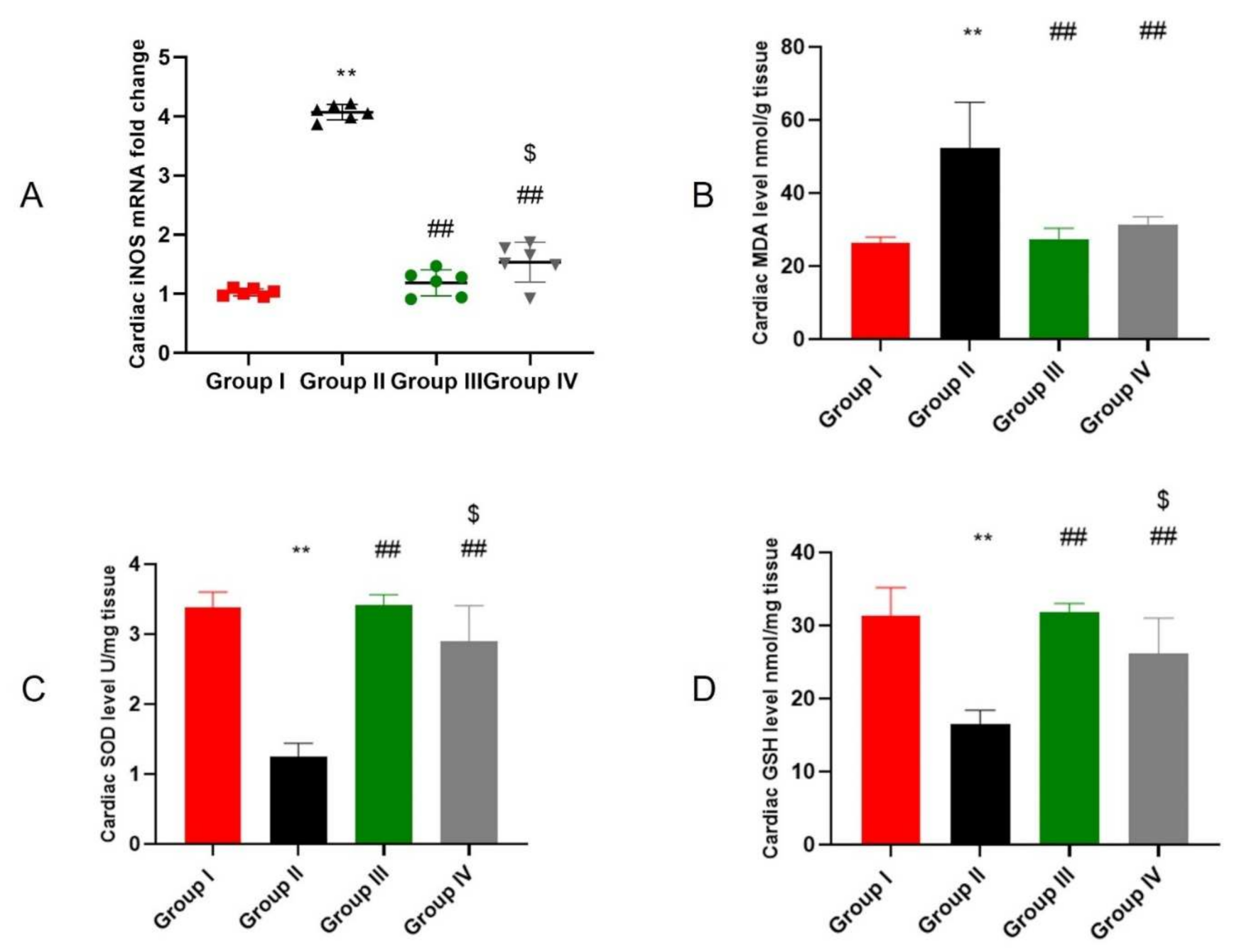
2.4. ADMSCs-Derived Exosomes Attenuated DOX/Trz-Induced Cardiac Oxidative Stress
2.5. ADMSCs-Derived Exosomes Ameliorated DOX/Trz-Induced DNA Damage and Apoptosis in Cardiac Tissues
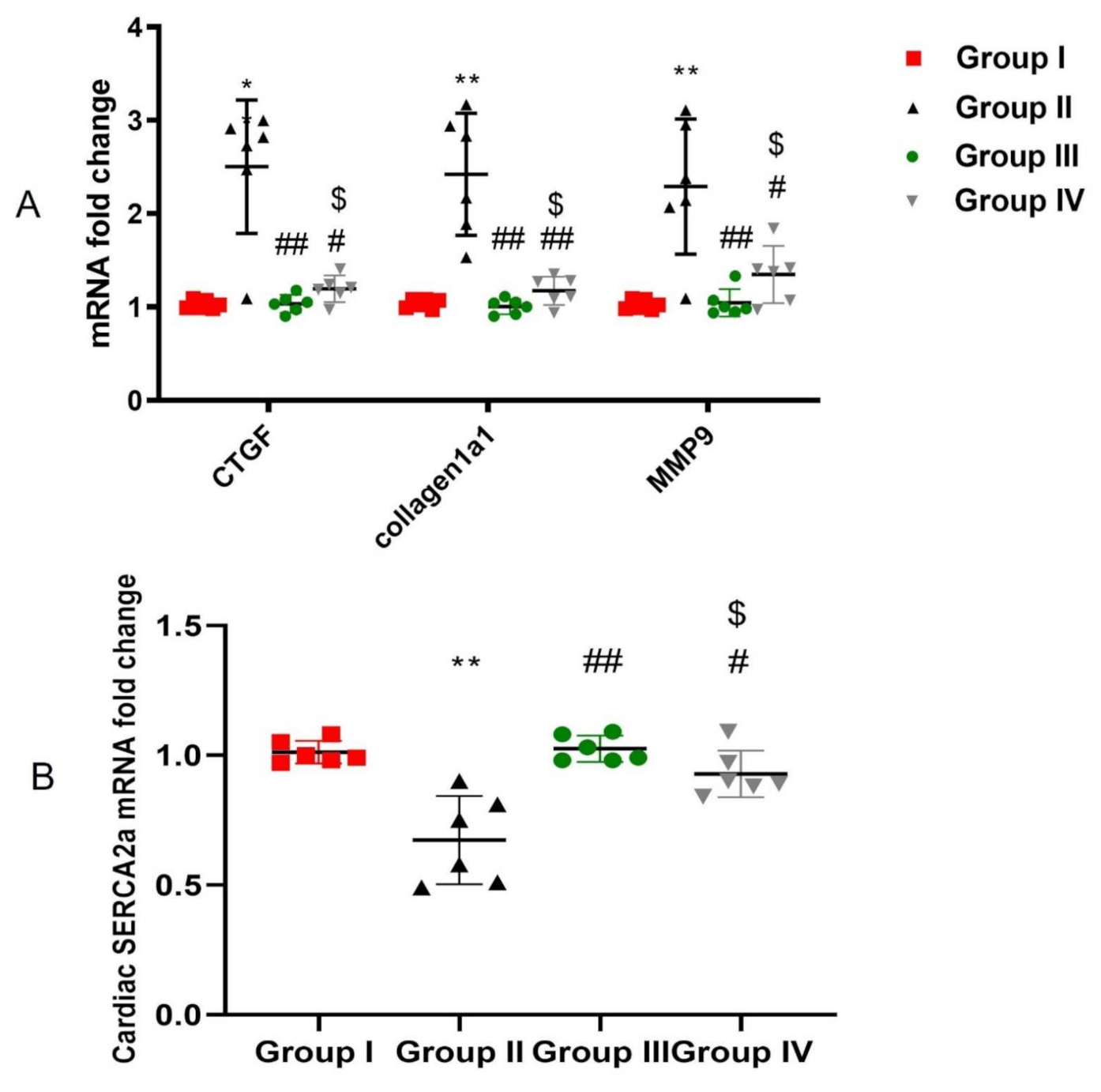
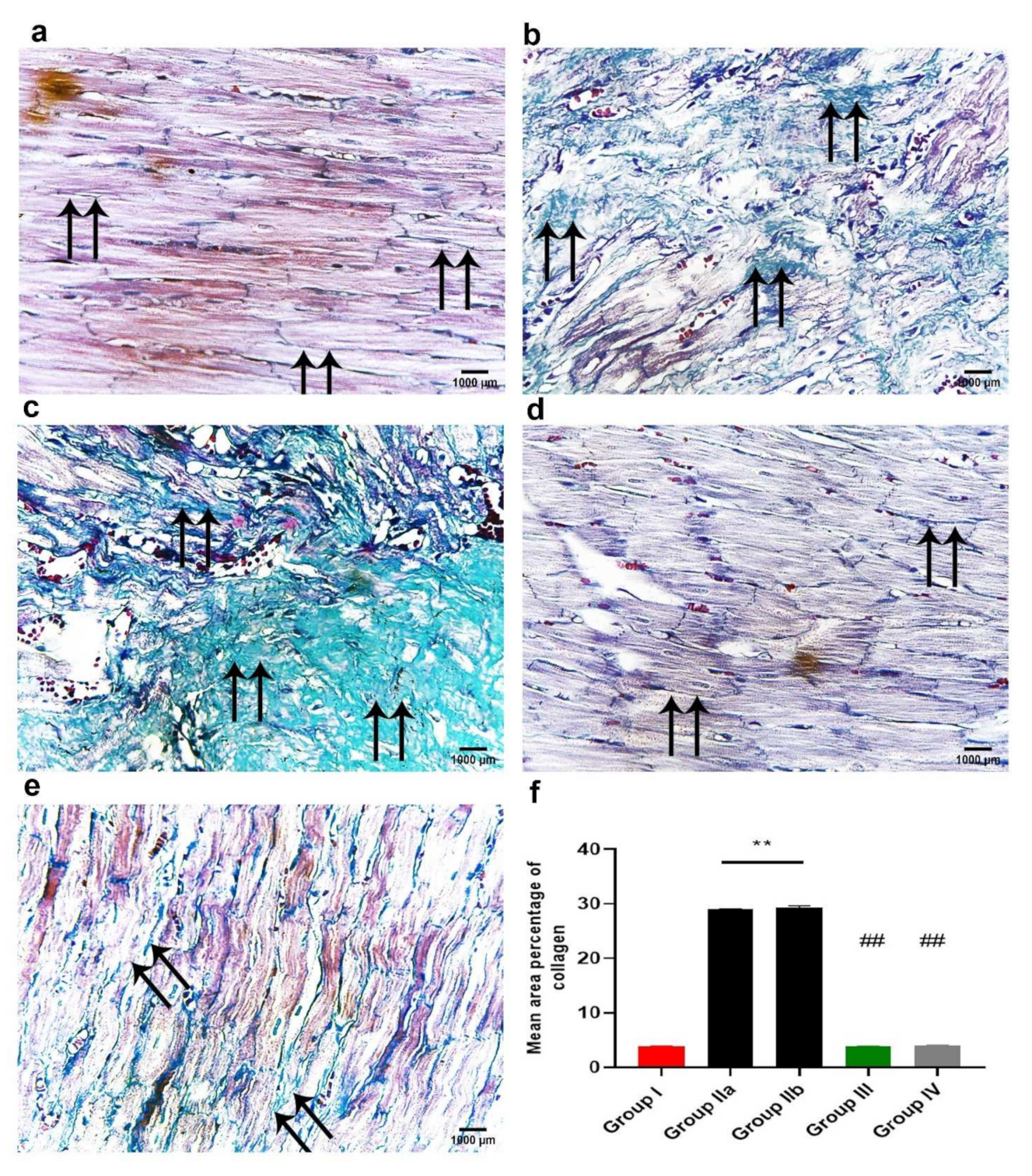
2.6. ADMSCs-Derived Exosomes Attenuated DOX/Trz-Induced Fibrosis in Cardiac Tissues
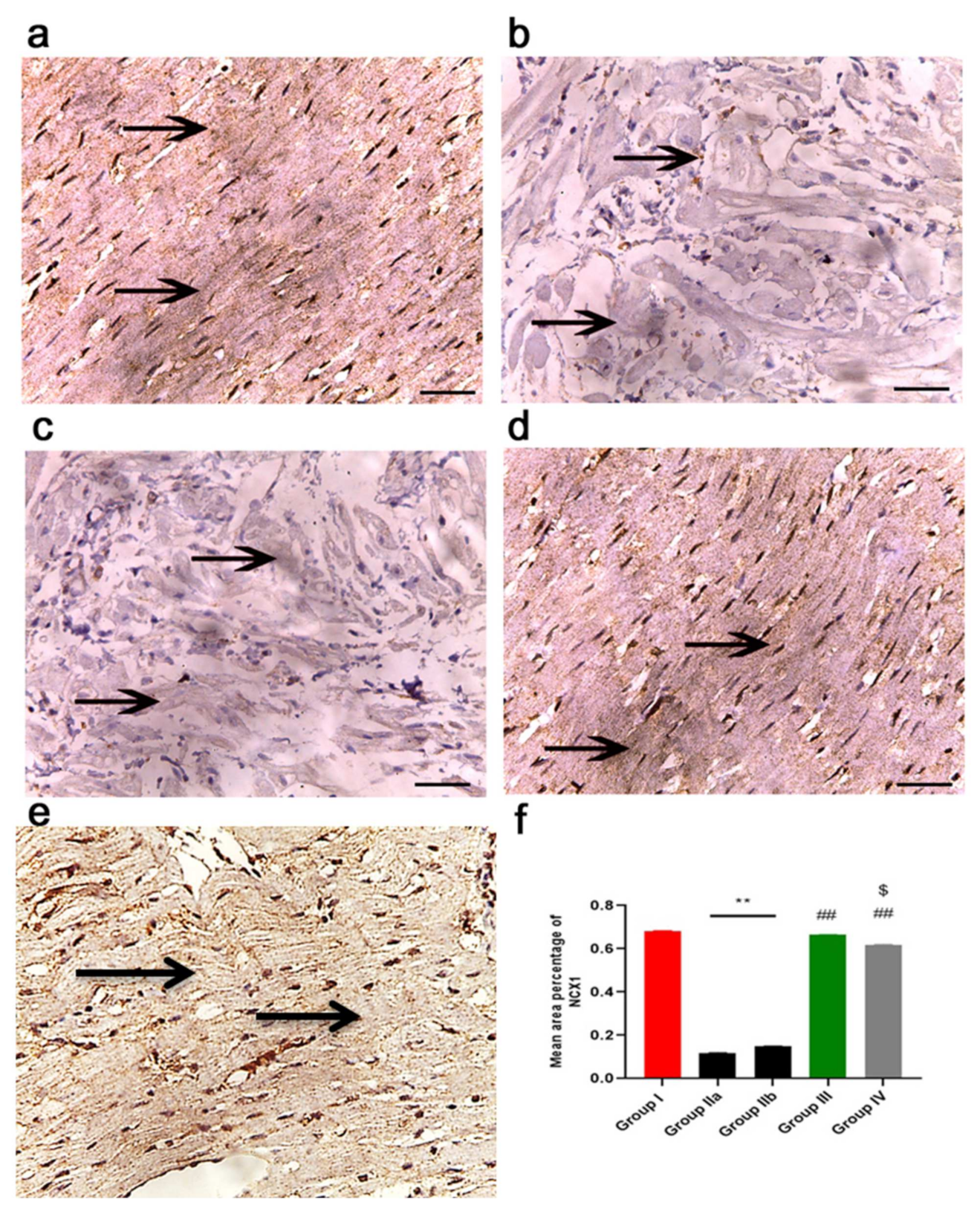
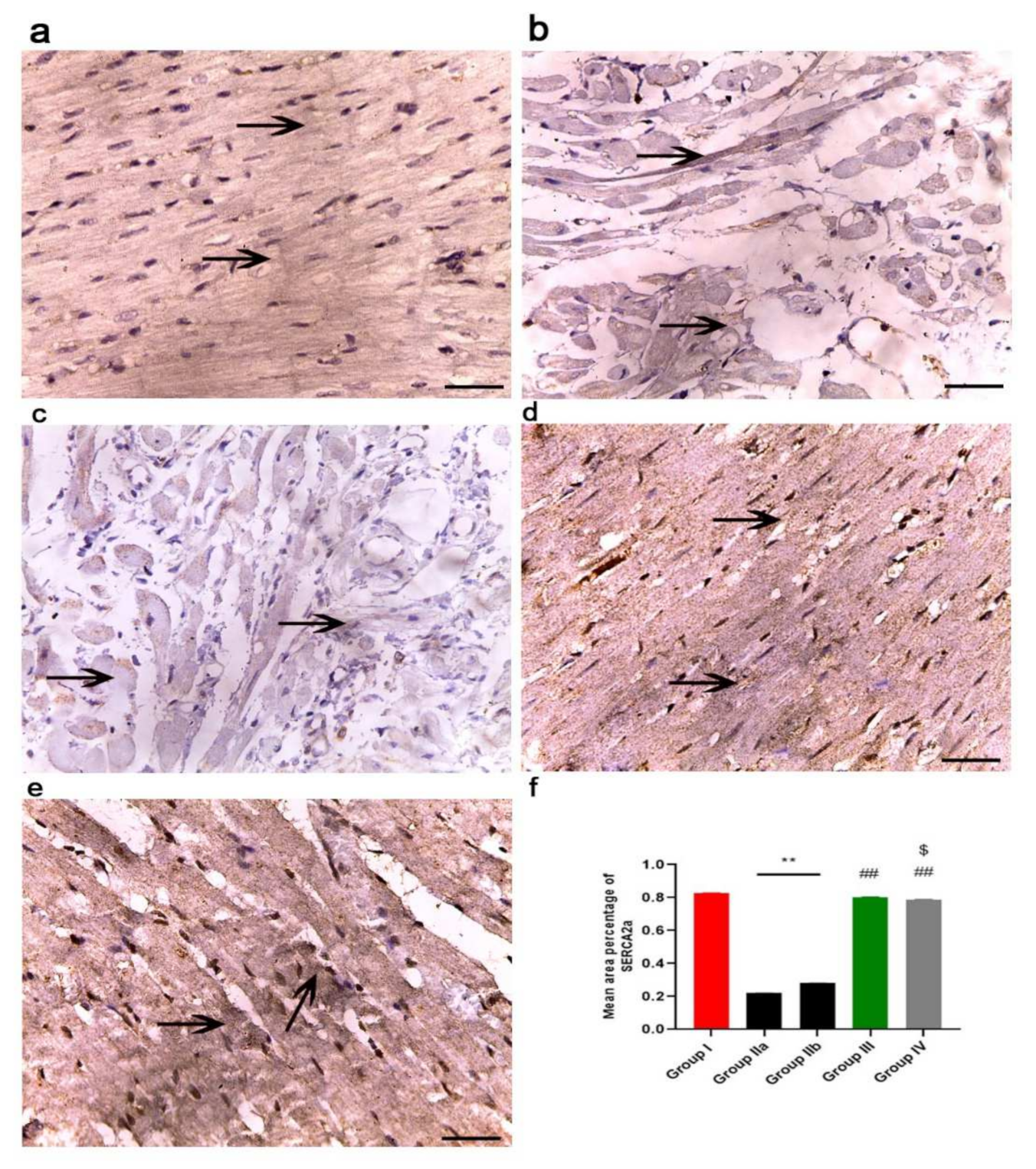
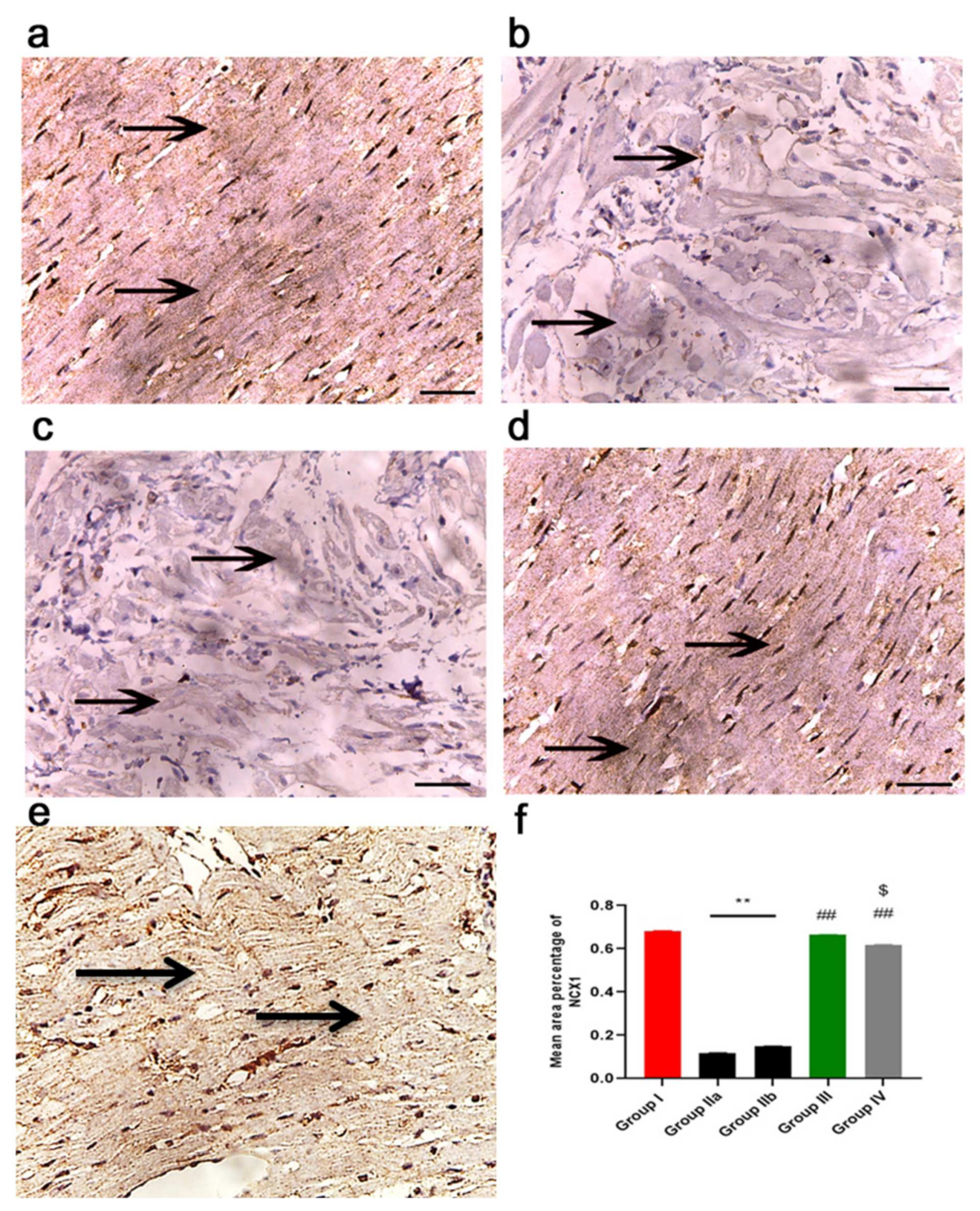
2.7. ADMSCs-Derived Exosomes Improved Cardiac Ca2+ Homeostasis in DOX/Trz-Treated Rats
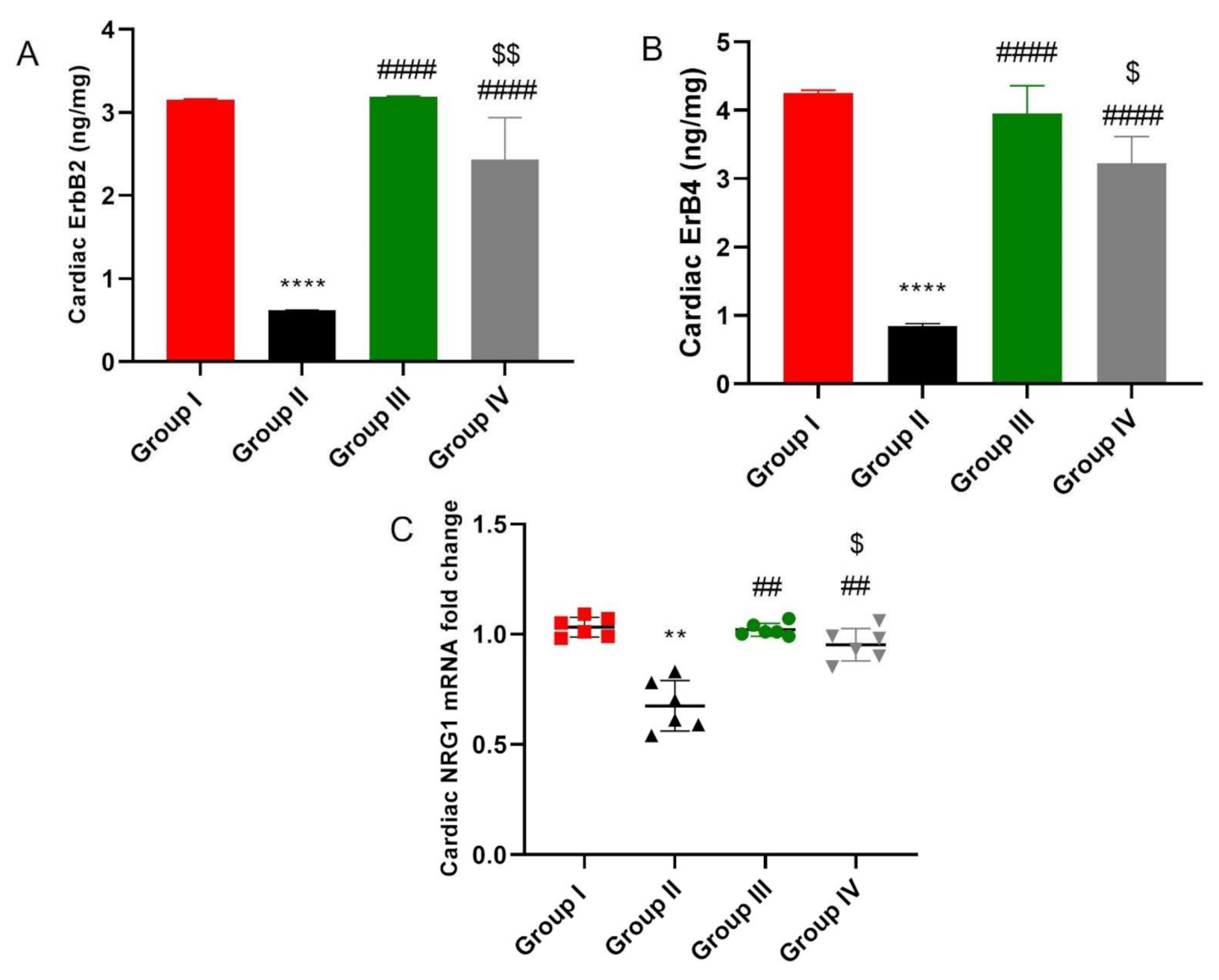
2.8. ADMSCs-Derived Exosomes Restored NRG-1/HER2 Signaling in Cardiac Tissue of DOX/Trz-Treated Rats
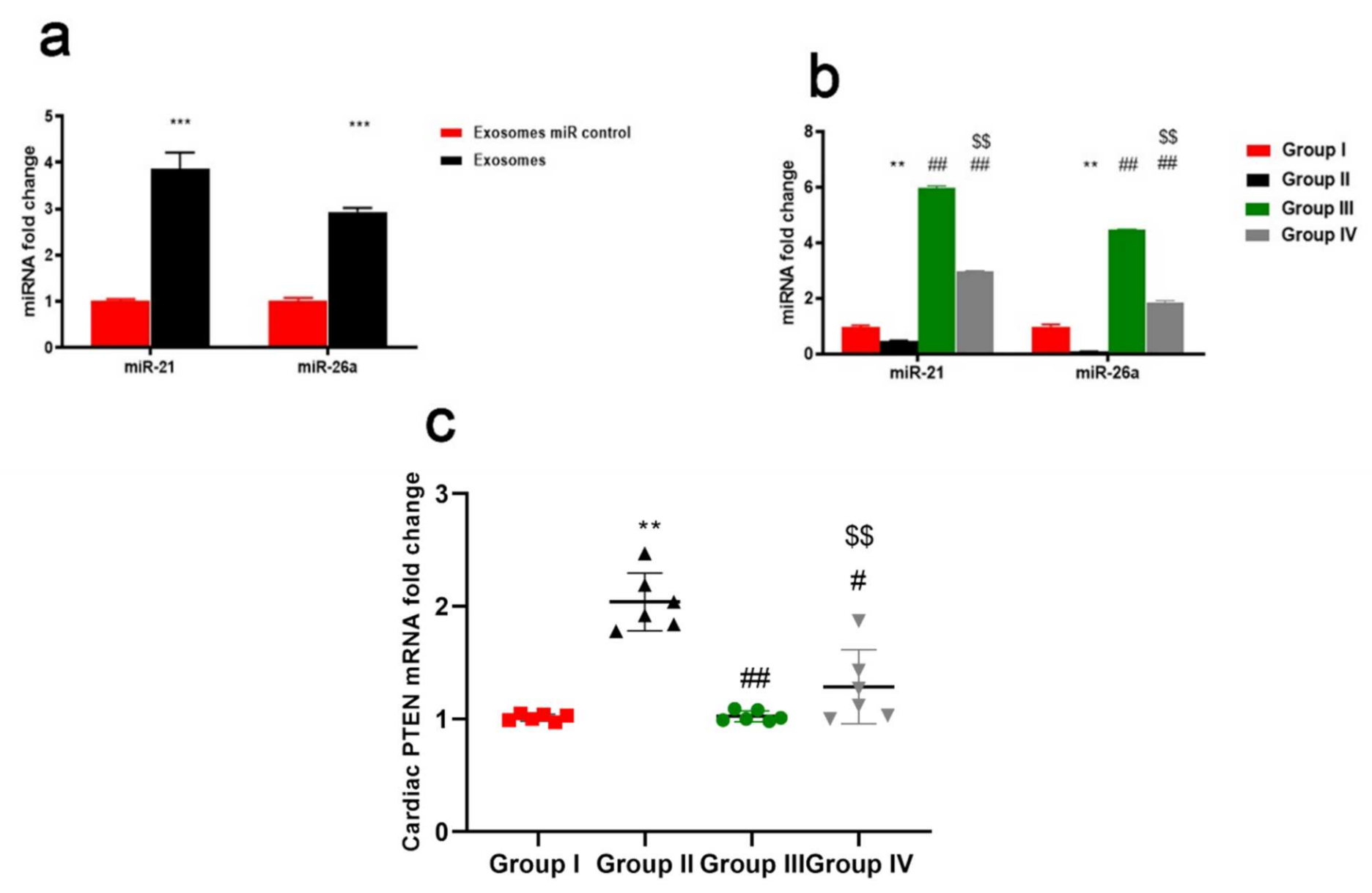
2.9. ADMSCs-Derived Exosomes Increased Expression of miR-21 and miR-26a Restraining PTEN in Cardiac Tissue of DOX/Trz-Treated Rats
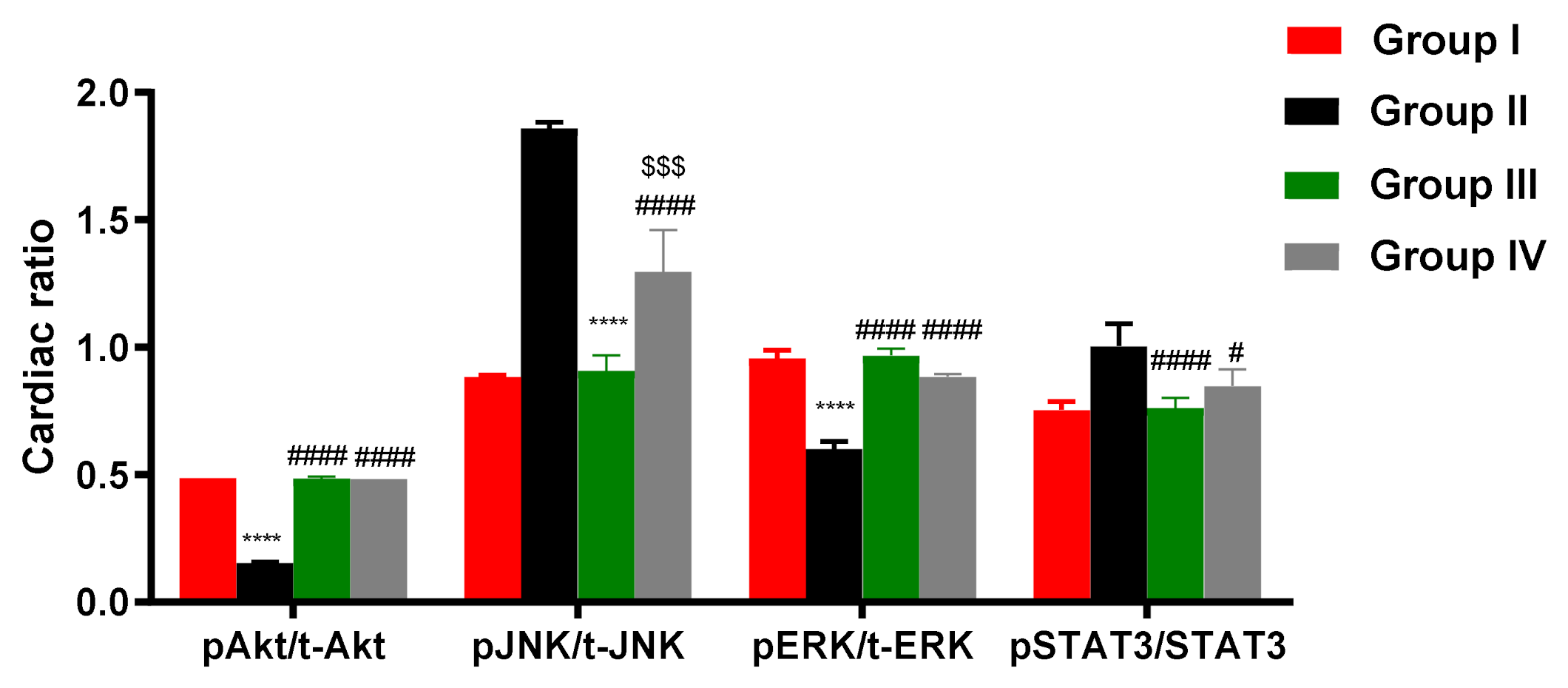
2.10. ADMSCs-Derived Exosomes Modulated pAKT/AKT, pERK/ERK, pJNK/JNK, and PSTAT/STAT in Cardiac Tissue of DOX/Trz-Treated Rats
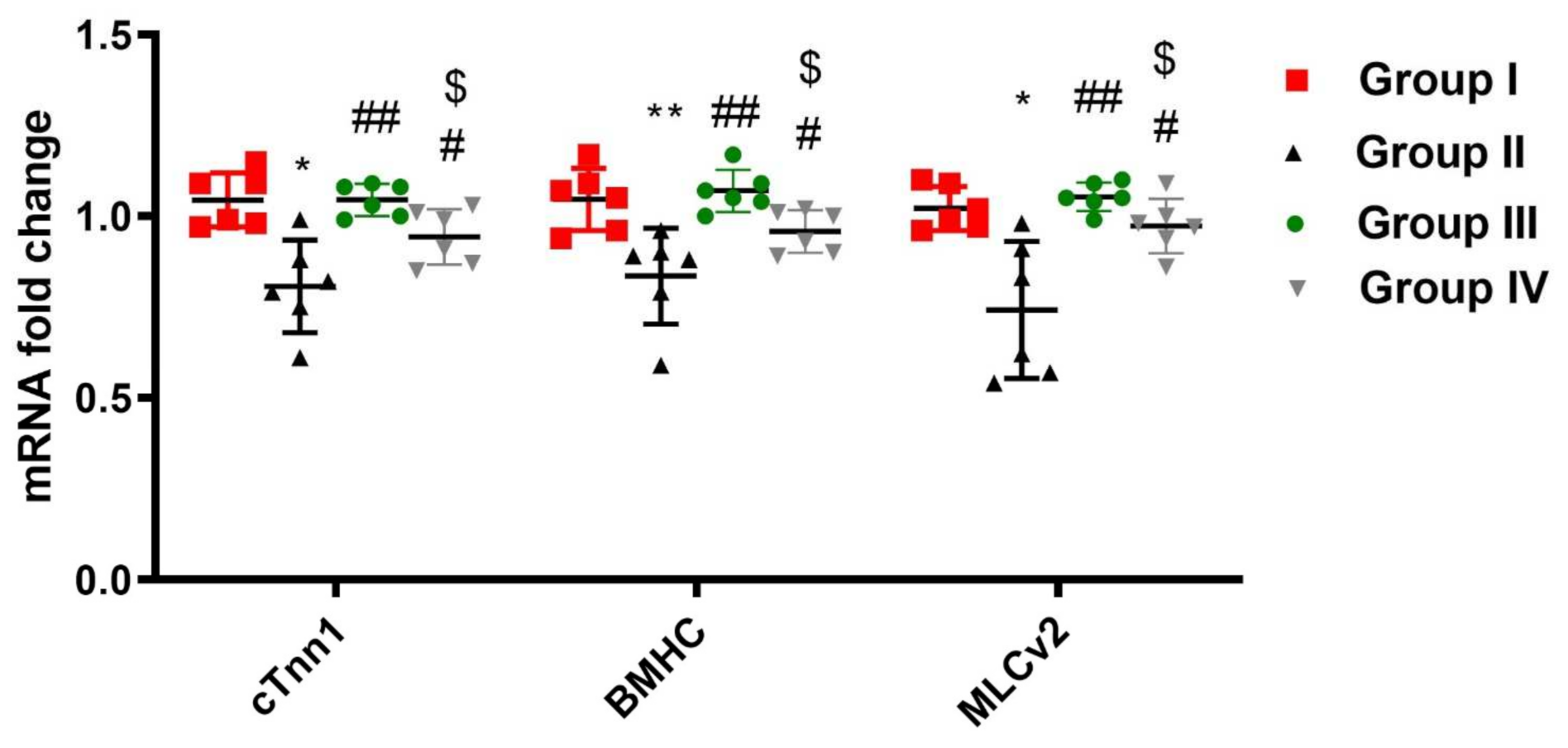
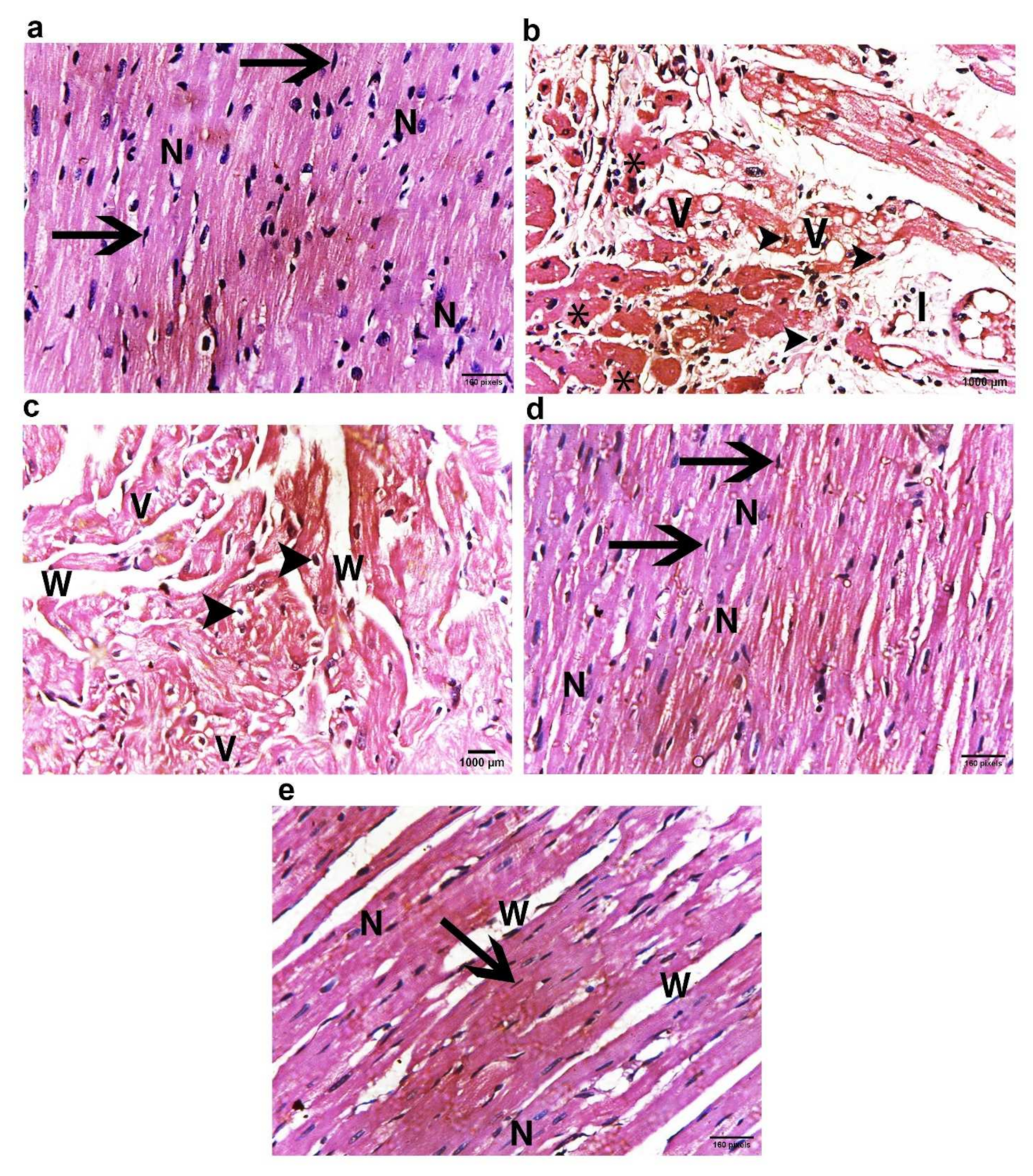
2.11. ADMSCs-Derived Exosomes Restored DOX/Trz-Induced Alteration in Cardiac Muscle Histopathology
2.11.1. Light Microscope Examination
H&E Staining Results
2.11.2. Masson’s Trichrome Stain for Collagen Fibers
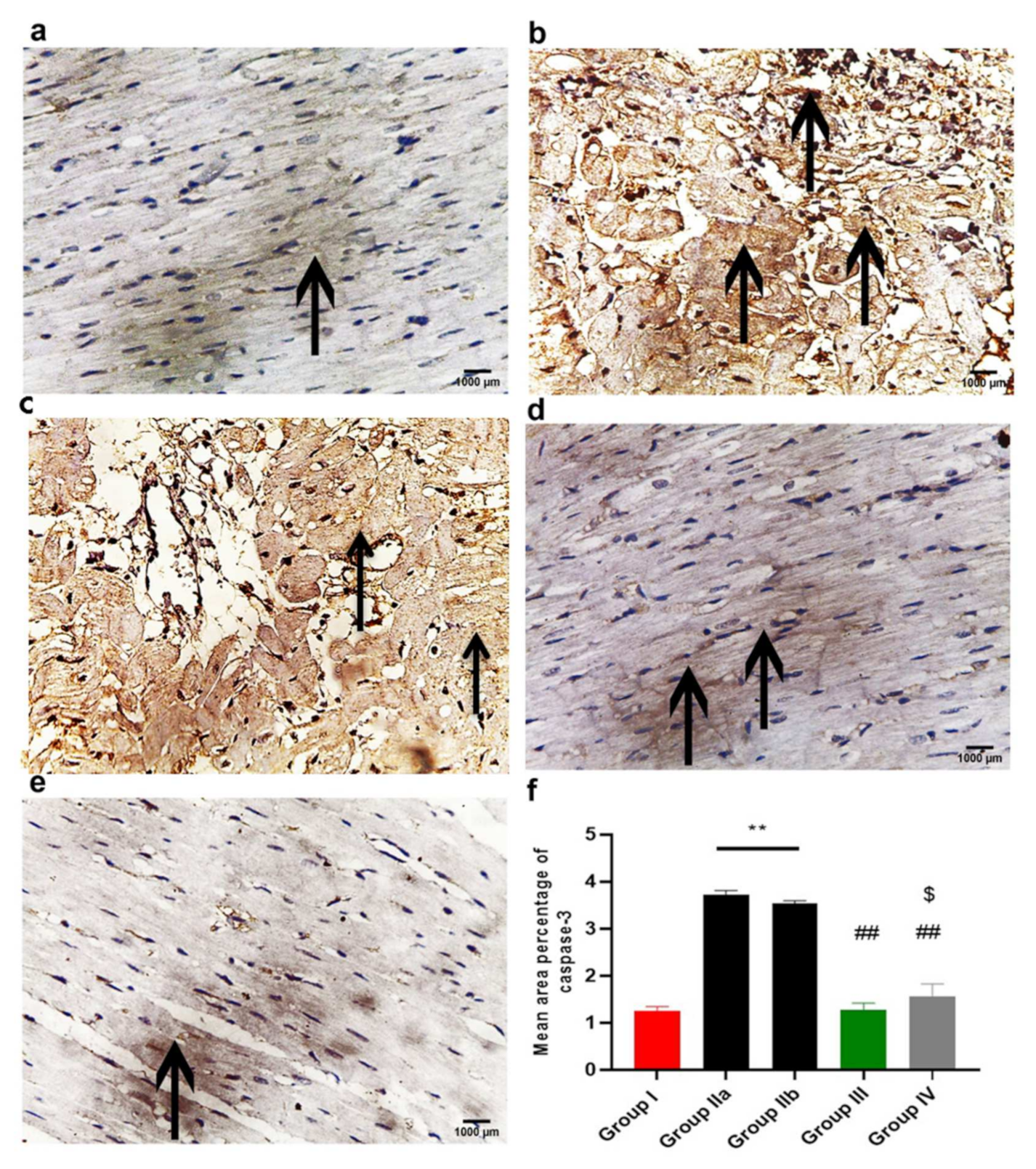
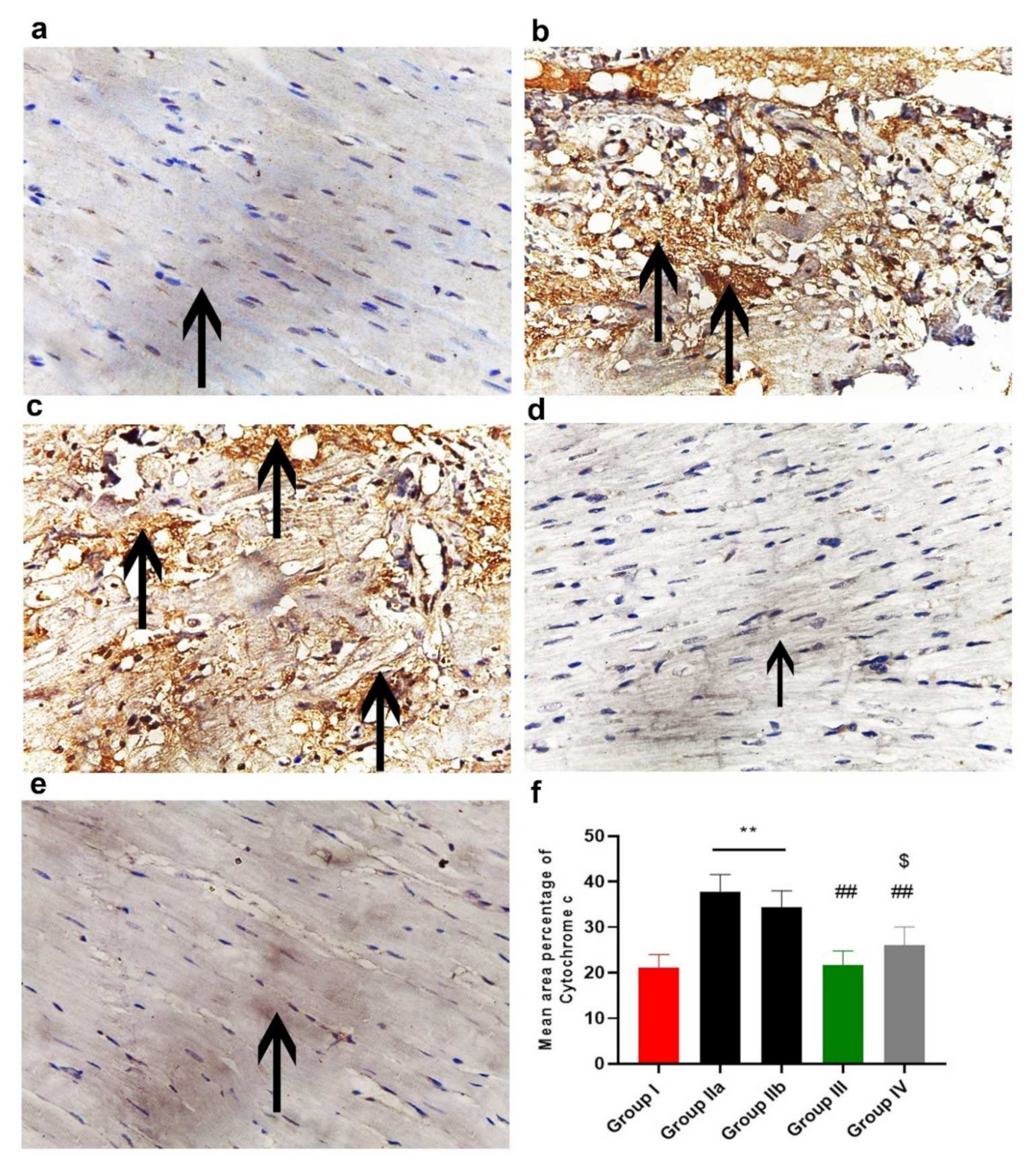
2.11.3. Immunohistochemistry Results
2.11.4. Morphometric Analysis
2.11.5. Transmission Electron Microscopy Study
3. Discussion
4. Materials and Methods
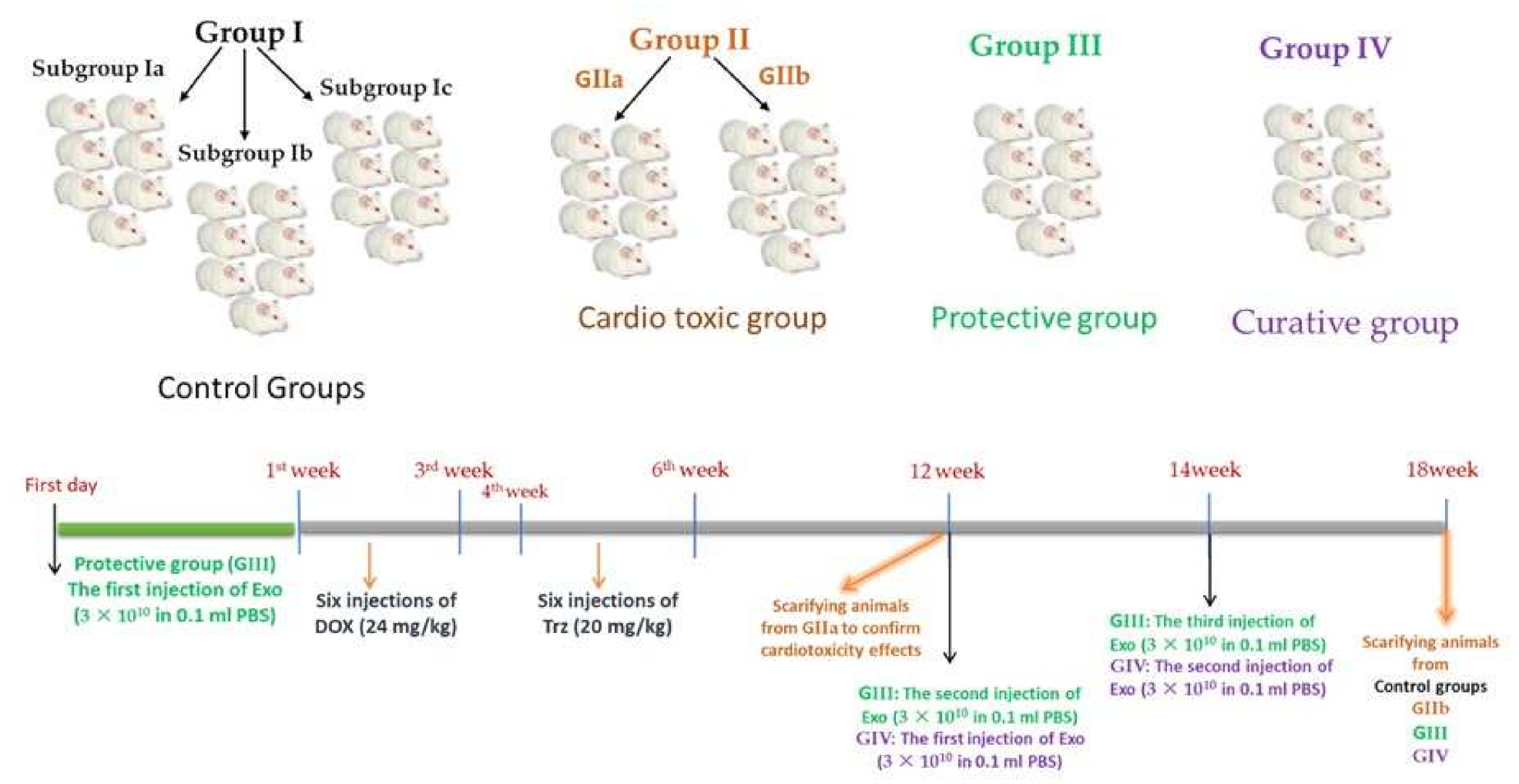
4.1. Experimental Animals
4.2. Preparation of MSCs-Derived Exosomes
4.3. Characterization of MSC-Derived Exosomes
4.4. Western Blot for Characterization and Localization of Exosomes
4.5. Direct Labeling of MSCs with Iron Oxide
4.6. Prussian Blue Staining
4.7. Experimental Chemicals
- − Doxorubicin hydrochloride (DOX), as a powder, was purchased from Genentech, Inc., South San Francisco, CA, USA [Product code; D1515- ID 24893465].
- − Dosage of DOX: Doxorubicin hydrochloride (DOX) powder was dissolved in sterile distilled water before administration. Each 80 mg of DOX powder was dissolved in 10 mL of sterile distilled water, yielding a multi-dose solution containing 8 mg/mL. Each rat of the tested groups was injected intraperitoneally with 0.13 mL of solution (1 mg/dose), according to Nicol et al. [20].
- − Trastuzumab (Trz), as HERCEPTIN® lyophilized powder (440 mg), was purchased from Genentech, Inc., South San Francisco, CA, USA [Product code; SAP-10086840].
- − Dosage of Trz: Trastuzumab (Herceptin) lyophilized powder was dissolved in sterile distilled water before administration. Each vial (440 mg) of Herceptin was dissolved in 60 mL of sterile distilled water to yield a multi-dose solution containing 7.3 mg/mL. Each rat of the tested groups was injected intraperitoneally with 0.12 mL of solution (0.87 mg/dose), according to Nicol et al. [20].
- − Sterile distilled water was purchased from ATECO Pharma (Kaliobeya, Egypt).
- − All drugs and chemicals were of the highest purity and analytical grade and were freshly prepared before each experiment.
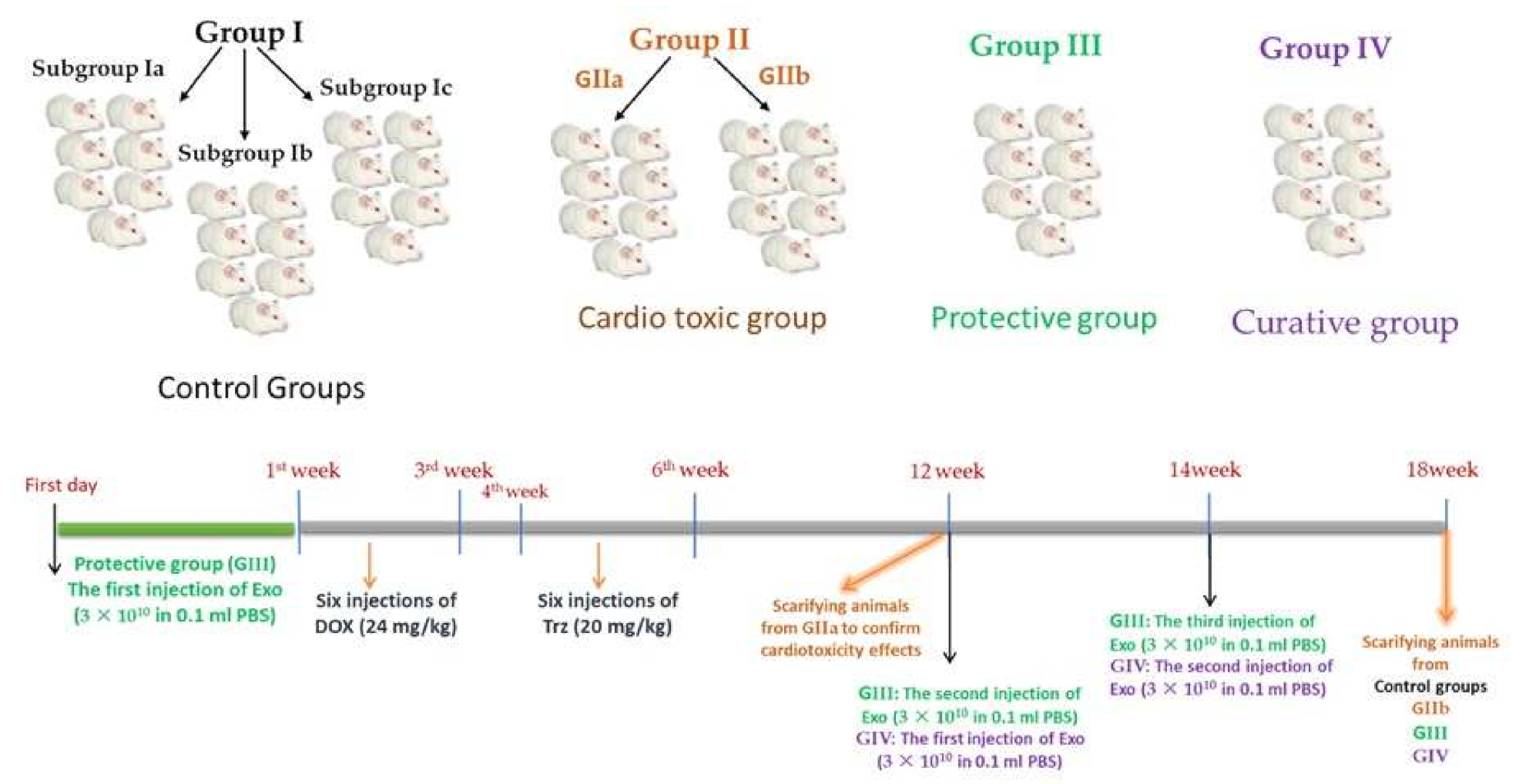
4.8. Experimental Design
4.9. Sampling
4.10. Determination of Serum Cardiac Enzymes and Cardiac Tissue Oxidative Stress Indices
4.11. Echocardiography
4.12. Gene Expression Profile
4.13. Enzyme-Linked Immunosorbent Assay (ELISA)
4.14. Histopathological Analysis
4.15. Immunohistochemical Analysis
4.16. Morphometric Study
4.17. Transmission Electron Microscopy Study
4.18. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Iqbal, N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol. Biol. Int. 2014, 2014, 852748. [Google Scholar] [CrossRef]
- Gianni, L.; Eiermann, W.; Semiglazov, V.; Lluch, A.; Tjulandin, S.; Zambetti, M.; Moliterni, A.; Vazquez, F.; Byakhov, M.J.; Lichinitser, M.; et al. Neoadjuvant and adjuvant trastuzumab in patients with HER2-positive locally advanced breast cancer (NOAH): Follow-up of a randomised controlled superiority trial with a parallel HER2-negative cohort. Lancet Oncol. 2014, 15, 640–647. [Google Scholar] [CrossRef]
- You, Y.; Xu, Z.; Chen, Y. Doxorubicin conjugated with a trastuzumab epitope and an MMP-2 sensitive peptide linker for the treatment of HER2-positive breast cancer. Drug Deliv. 2018, 25, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Zeglinski, M.; Ludke, A.; Jassal, D.S.; Singal, P.K. Trastuzumab-induced cardiac dysfunction: A ‘dual-hit’. Exp. Clin. Cardiol. 2011, 16, 70–74. [Google Scholar] [PubMed]
- Cortazar, P.; Zhang, L.; Untch, M.; Mehta, K.; Costantino, J.P.; Wolmark, N.; Bonnefoi, H.; Cameron, D.; Gianni, L.; Valagussa, P.; et al. Pathological complete response and long-term clinical benefit in breast cancer: The CTNeoBC pooled analysis. Lancet 2014, 384, 164–172. [Google Scholar] [CrossRef]
- Seidman, A.; Hudis, C.; Pierri, M.K.; Shak, S.; Paton, V.; Ashby, M.; Murphy, M.; Stewart, S.J.; Keefe, D. Cardiac dysfunction in the trastuzumab clinical trials experience. J. Clin. Oncol. 2002, 20, 1215–1221. [Google Scholar] [CrossRef]
- Guo, Y.; Yu, Y.; Hu, S.; Chen, Y.; Shen, Z. The therapeutic potential of mesenchymal stem cells for cardiovascular diseases. Cell Death Dis. 2020, 11, 349. [Google Scholar] [CrossRef]
- Yin, K.; Wang, S.; Zhao, R.C. Exosomes from mesenchymal stem/stromal cells: A new therapeutic paradigm. Biomark. Res. 2019, 7, 8. [Google Scholar] [CrossRef]
- Sun, X.; Shan, A.; Wei, Z.; Xu, B. Intravenous mesenchymal stem cell-derived exosomes ameliorate myocardial inflammation in the dilated cardiomyopathy. Biochem. Biophys. Res. Commun. 2018, 503, 2611–2618. [Google Scholar] [CrossRef]
- Goyal, V.; Bews, H.; Cheung, D.; Premecz, S.; Mandal, S.; Shaikh, B.; Best, R.; Bhindi, R.; Chaudhary, R.; Ravandi, A.; et al. The Cardioprotective Role of N-Acetyl Cysteine Amide in the Prevention of Doxorubicin and Trastuzumab-Mediated Cardiac Dysfunction. Can. J. Cardiol. 2016, 32, 1513–1519. [Google Scholar] [PubMed]
- Rawat, P.S.; Jaiswal, A.; Khurana, A.; Bhatti, J.S.; Navik, U. Doxorubicin-induced cardiotoxicity: An update on the molecular mechanism and novel therapeutic strategies for effective management. Biomed. Pharmacother. 2021, 139, 111708. [Google Scholar] [PubMed]
- Vermeulen, Z.; Hervent, A.S.; Dugaucquier, L.; Vandekerckhove, L.; Rombouts, M.; Beyens, M.; Schrijvers, D.M.; De Meyer, G.R.Y.; Maudsley, S.; De Keulenaer, G.W.; et al. Inhibitory actions of the NRG-1/ErbB4 pathway in macrophages during tissue fibrosis in the heart, skin, and lung. Am. J. Physiol Heart Circ. Physiol. 2017, 313, H934–H945. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, F.; Das, S.; Sysa-Shah, P.; Sivakumaran, V.; Stanley, B.; Guo, X.; Paolocci, N.; Aon, M.A.; Nagane, M.; Kuppusamy, P.; et al. ErbB2 overexpression upregulates antioxidant enzymes, reduces basal levels of reactive oxygen species, and protects against doxorubicin cardiotoxicity. Am. J. Physiol Heart Circ. Physiol. 2015, 309, H1271–H1280. [Google Scholar] [CrossRef] [PubMed]
- Ozcelik, C.; Erdmann, B.; Pilz, B.; Wettschureck, N.; Britsch, S.; Hübner, N.; Chien, K.R.; Birchmeier, C.; Garratt, A.N. Conditional mutation of the ErbB2 (HER2) receptor in cardiomyocytes leads to dilated cardiomyopathy. Proc. Natl. Acad. Sci. USA 2002, 99, 8880–8885. [Google Scholar] [CrossRef]
- Rochette, L.; Guenancia, C.; Gudjoncik, A.; Hachet, O.; Zeller, M.; Cottin, Y.; Vergely, C. Anthracyclines/trastuzumab: New aspects of cardiotoxicity and molecular mechanisms. Trends. Pharmacol. Sci. 2015, 36, 326–348. [Google Scholar] [CrossRef]
- Asselin, C.Y.; Lam, A.; Cheung, D.Y.C.; Eekhoudt, C.R.; Zhu, A.; Mittal, I.; Mayba, A.; Solati, Z.; Edel, A.; Austria, J.A.; et al. The Cardioprotective Role of Flaxseed in the Prevention of Doxorubicin- and Trastuzumab-Mediated Cardiotoxicity in C57BL/6 Mice. J. Nutr. 2020, 150, 2353–2363. [Google Scholar]
- Grazette, L.P.; Boecker, W.; Matsui, T.; Semigran, M.; Force, T.L.; Hajjar, R.J.; Rosenzweig, A. Inhibition of ErbB2 causes mitochondrial dysfunction in cardiomyocytes: Implications for herceptin-induced cardiomyopathy. J. Am. Coll. Cardiol. 2004, 44, 2231–2238. [Google Scholar] [CrossRef]
- Walker, J.R.; Sharma, A.; Lytwyn, M.; Bohonis, S.; Thliveris, J.; Singal, P.K.; Jassal, D.S. The cardioprotective role of probucol against anthracycline and trastuzumab-mediated cardiotoxicity. J. Am. Soc Echocardiogr. 2011, 24, 699–705. [Google Scholar]
- Nicol, M.; Sadoune, M.; Polidano, E.; Launay, J.M.; Samuel, J.L.; Azibani, F.; Cohen-Solal, A. Doxorubicin-induced and trastuzumab-induced cardiotoxicity in mice is not prevented by metoprolol. ESC Heart Fail. 2021, 8, 928–937. [Google Scholar] [CrossRef]
- Milano, G.; Biemmi, V.; Lazzarini, E.; Balbi, C.; Ciullo, A.; Bolis, S.; Ameri, P.; Di Silvestre, D.; Mauri, P.; Barile, L.; et al. Intravenous administration of cardiac progenitor cell-derived exosomes protects against doxorubicin/trastuzumab-induced cardiac toxicity. Cardiovasc. Res. 2020, 116, 383–392. [Google Scholar] [CrossRef]
- Xia, P.; Liu, Y.; Cheng, Z. Signaling Pathways in Cardiac Myocyte Apoptosis. Biomed. Res. Int. 2016, 2016, 9583268. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, W.A.; Ahad, A.; Ahsan, H. The mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update. Arch. Toxicol. 2015, 89, 289–317. [Google Scholar] [CrossRef] [PubMed]
- Yee, K.S.; Wilkinson, S.; James, J.; Ryan, K.M.; Vousden, K.H. PUMA- and Bax-induced autophagy contributes to apoptosis. Cell Death Differ. 2009, 16, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.L.; Kerrigan, J.E.; Lin, C.P.; Azarova, A.M.; Tsai, Y.C.; Ban, Y.; Liu, L.F. Topoisomerase IIbeta mediated DNA double-strand breaks: Implications in doxorubicin cardiotoxicity and prevention by dexrazoxane. Cancer Res. 2007, 67, 8839–8846. [Google Scholar] [CrossRef] [PubMed]
- Mohan, N.; Jiang, J.; Dokmanovic, M.; Wu, W.J. Trastuzumab-mediated cardiotoxicity: Current understanding, challenges, and frontiers. Antib. Ther. 2018, 1, 13–17. [Google Scholar] [CrossRef]
- Pentassuglia, L.; Sawyer, D.B. The role of Neuregulin-1beta/ErbB signaling in the heart. Exp. Cell Res. 2009, 315, 627–637. [Google Scholar] [CrossRef]
- Kuramochi, Y.; Guo, X.; Sawyer, D.B. Neuregulin activates erbB2-dependent src/FAK signaling and cytoskeletal remodeling in isolated adult rat cardiac myocytes. J. Mol. Cell. Cardiol. 2006, 41, 228–235. [Google Scholar] [CrossRef]
- Gordon, L.I.; Burke, M.A.; Singh, A.T.K.; Prachand, S.; Lieberman, E.D.; Sun, L.; Naik, T.J.; Prasad, S.V.; Ardehali, H. Blockade of the erbB2 receptor induces cardiomyocyte death through mitochondrial and reactive oxygen species-dependent pathways. J. Biol. Chem. 2009, 284, 2080–2087. [Google Scholar] [CrossRef]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef]
- Ghosh, J.; Das, J.; Manna, P.; Sil, P.C. The protective role of arjunolic acid against doxorubicin induced intracellular ROS dependent JNK-p38 and p53-mediated cardiac apoptosis. Biomaterials 2011, 32, 4857–4866. [Google Scholar] [CrossRef]
- Wang, B.; Shravah, J.; Luo, H.; Raedschelders, K.; Chen, D.D.Y.; Ansley, D.M. Propofol protects against hydrogen peroxide-induced injury in cardiac H9c2 cells via Akt activation and Bcl-2 up-regulation. Biochem Biophys Res. Commun. 2009, 389, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, N.; Baker, S.J. PTEN and the PI3-kinase pathway in cancer. Annu. Rev. Pathol. 2009, 4, 127–150. [Google Scholar] [CrossRef]
- Gottlieb, T.M.; Leal, J.F.M.; Seger, R.; Taya, Y.; Oren, M. Cross-talk between Akt, p53 and Mdm2: Possible implications for the regulation of apoptosis. Oncogene 2002, 21, 1299–1303. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Stephen, C.W.; Luciani, M.G.; Fåhraeus, R. p53 Stability and activity is regulated by Mdm2-mediated induction of alternative p53 translation products. Nat. Cell Biol. 2002, 4, 462–467. [Google Scholar] [CrossRef]
- Mayo, L.D.; Donner, D.B. The PTEN, Mdm2, p53 tumor suppressor-oncoprotein network. Trends Biochem. Sci. 2002, 27, 462–467. [Google Scholar] [CrossRef]
- ElZarrad, M.K.; Mukhopadhyay, P.; Mohan, N.; Hao, E.; Dokmanovic, M.; Hirsch, D.S.; Shen, Y.; Pacher, P.; Wu, W.J. Trastuzumab alters the expression of genes essential for cardiac function and induces ultrastructural changes of cardiomyocytes in mice. PLoS ONE 2013, 8, e79543. [Google Scholar] [CrossRef]
- Bian, Y.; Sun, M.; Silver, M.; Ho, K.K.L.; Marchionni, M.A.; Caggiano, A.O.; Stone, J.R.; Amende, I.; Hampton, T.G.; Morgan, J.P.; et al. Neuregulin-1 attenuated doxorubicin-induced decrease in cardiac troponins. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1974–H1983. [Google Scholar] [CrossRef]
- Yan, X.; Goukassian, D.; Morg, J. Neuregulin1-ErbB Signaling in Doxorubicin-Induced Cardiotoxicity. In Cardiotoxicity of Oncologic Treatments; Fiuza, M., Ed.; InTech: Rijeka, Croatia, 2012; Volume 3, pp. 565–573. [Google Scholar]
- Wehrens, X.H.T.; Marks, A.R. Novel therapeutic approaches for heart failure by normalizing calcium cycling. Nat. Rev. Drug Discov. 2004, 3, 565–573. [Google Scholar] [CrossRef]
- Schmidt, U.; Hajjar, R.J.; Kim, C.S.; Lebeche, D.; Doye, A.A.; Gwathmey, J.K. Human heart failure: cAMP stimulation of SR Ca(2+)-ATPase activity and phosphorylation level of phospholamban. Am. J. Physiol. 1999, 277, H474–H480. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.S.; Pillai, K.K.; Hassan, M.Q.; Dhyani, N.; Ismail, M.V.; Najmi, A.K. Levosimendan reduces myocardial damage and improves cardiodynamics in streptozotocin induced diabetic cardiomyopathy via SERCA2a/NCX1 pathway. Life Sci. 2016, 153, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Olson, R.D.; Gambliel, H.A.; Vestal, R.E.; Shadle, S.E.; Charlier, H.A.; Cusack, B.J. Doxorubicin cardiac dysfunction: Effects on calcium regulatory proteins, sarcoplasmic reticulum, and triiodothyronine. Cardiovasc. Toxicol. 2005, 5, 269–283. [Google Scholar] [CrossRef]
- Berdichevski, A.; Meiry, G.; Milman, F.; Reiter, I.; Sedan, O.; Eliyahu, S.; Duffy, H.S.; Youdim, M.B.; Binah, O. TVP1022 protects neonatal rat ventricular myocytes against doxorubicin-induced functional derangements. J. Pharmacol. Exp. Ther. 2010, 332, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Ujihara, Y.; Iwasaki, K.; Takatsu, S.; Hashimoto, K.; Naruse, K.; Mohri, S.; Katanosaka, Y. Induced NCX1 overexpression attenuates pressure overload-induced pathological cardiac remodelling. Cardiovasc. Res. 2016, 111, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, J.A.; Mergaye, M.; Kumar, N.; Chen, C.A.; Angelos, M.G.; Khan, M. Potential Role of Exosomes in Mending a Broken Heart: Nanoshuttles Propelling Future Clinical Therapeutics Forward. Stem Cells Int. 2017, 2017, 5785436. [Google Scholar] [CrossRef]
- Kim, H.S.; Choi, D.Y.; Yun, S.J.; Choi, S.M.; Kang, J.W.; Jung, J.W.; Hwang, D.; Kim, K.P.; Kim, D.W. Proteomic analysis of microvesicles derived from human mesenchymal stem cells. J. Proteome Res. 2012, 11, 839–849. [Google Scholar] [CrossRef]
- Xu, Y.; Li, X.; Liu, X.; Zhou, M. Neuregulin-1/ErbB signaling and chronic heart failure. Adv. Pharmacol. 2010, 59, 31–51. [Google Scholar]
- Wadugu, B.; Kühn, B. The role of neuregulin/ErbB2/ErbB4 signaling in the heart with special focus on effects on cardiomyocyte proliferation. Am. J. Physiol Heart Circ. Physiol. 2012, 302, H2139–H2147. [Google Scholar] [CrossRef]
- Aries, A.; Paradis, P.; Lefebvre, C.; Schwartz, R.J.; Nemer, M. Essential role of GATA-4 in cell survival and drug-induced cardiotoxicity. Proc. Natl. Acad. Sci. USA 2004, 101, 6975–6980. [Google Scholar] [CrossRef]
- Sun, M.; Yan, X.; Bian, Y.; Caggiano, A.O.; Morgan, J.P. Improving murine embryonic stem cell differentiation into cardiomyocytes with neuregulin-1: Differential expression of microRNA. Am. J. Physiol. Cell Physiol. 2011, 301, C21–C30. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, R.; Miller, T.A.; Kuramochi, Y.; Frantz, S.; Kim, Y.D.; Marchionni, M.A.; Kelly, R.A.; Sawyer, D.B. Neuregulin-1 protects ventricular myocytes from anthracycline-induced apoptosis via erbB4-dependent activation of PI3-kinase/Akt. J. Mol. Cell. Cardiol. 2003, 35, 1473–1479. [Google Scholar] [CrossRef]
- Rohrbach, S.; Muller-Werdan, U.; Werdan, K.; Koch, S.; Gellerich, N.F.; Holtz, J. Apoptosis-modulating interaction of the neuregulin/erbB pathway with anthracyclines in regulating Bcl-xS and Bcl-xL in cardiomyocytes. J. Mol. Cell. Cardiol. 2005, 38, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Figliolini, F.; Ranghino, A.; Grange, C.; Cedrino, M.; Tapparo, M.; Cavallari, C.; Rossi, A.; Togliatto, G.; Femminò, S.; Gugliuzza, M.V.; et al. Extracellular Vesicles From Adipose Stem Cells Prevent Muscle Damage and Inflammation in a Mouse Model of Hind Limb Ischemia: Role of Neuregulin-1. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Mai, Z.; Zhu, X.; Wu, T.; Chen, Y.; Geng, D.; Wang, J. Mesenchymal stem cell-derived exosomes ameliorate cardiomyocyte apoptosis in hypoxic conditions through microRNA144 by targeting the PTEN/AKT pathway. Stem Cell Res. Ther. 2020, 11, 36. [Google Scholar] [CrossRef]
- Sun, X.H.; Wang, X.; Zhang, Y.; Hui, J. Exosomes of bone-marrow stromal cells inhibit cardiomyocyte apoptosis under ischemic and hypoxic conditions via miR-486-5p targeting the PTEN/PI3K/AKT signaling pathway. Thromb. Res. 2019, 177, 23–32. [Google Scholar] [CrossRef]
- Yu, B.; Kim, H.W.; Gong, M.; Wang, J.; Millard, R.W.; Wang, Y.; Ashraf, M.; Xu, M. Exosomes secreted from GATA-4 overexpressing mesenchymal stem cells serve as a reservoir of anti-apoptotic microRNAs for cardioprotection. Int. J. Cardiol. 2015, 182, 349–360. [Google Scholar] [CrossRef]
- Lai, R.C.; Yeo, R.W.Y.; Tan, S.S.; Zhang, B.; Yin, Y.; Sze, N.S.K.; Choo, A.; Lim, S.K. Mesenchymal Stem Cell Exosomes: The Future MSC-Based Therapy. In Mesenchymal Stem Cell Therapy; Chase, L.G., Vemuri, M.C., Eds.; Humana Press: Totowa, NJ, USA, 2013. [Google Scholar]
- Janockova, J.; Slovinska, L.; Harvanova, D.; Spakova, T.; Rosocha, J. New therapeutic approaches of mesenchymal stem cells-derived exosomes. J. Biomed. Sci. 2021, 28, 39. [Google Scholar] [CrossRef]
- Deng, W.; Wang, Y.; Long, X.; Zhao, R.; Wang, Z.; Liu, Z.; Cao, S.; Shi, B. miR-21 Reduces Hydrogen Peroxide-Induced Apoptosis in c-kit+ Cardiac Stem Cells In Vitro through PTEN/PI3K/Akt Signaling. Oxid. Med. Cell Longev. 2016, 2016, 5389181. [Google Scholar] [CrossRef]
- Tu, Y.; Wan, L.; Fan, Y.; Wang, K.; Bu, L.; Huang, T.; Cheng, Z.; Shen, B. Ischemic postconditioning-mediated miRNA-21 protects against cardiac ischemia/reperfusion injury via PTEN/Akt pathway. PLoS ONE 2013, 8, e75872. [Google Scholar] [CrossRef]
- Wang, J.; Feng, Q.; Liang, D.; Shi, J. MiRNA-26a inhibits myocardial infarction-induced apoptosis by targeting PTEN via JAK/STAT pathways. Cells Dev. 2021, 165, 203661. [Google Scholar] [CrossRef] [PubMed]
- Timolati, F.; Ott, D.; Pentassuglia, L.; Giraud, M.N.; Perriard, J.C.; Suter, T.M.; Zuppinger, C. Neuregulin-1 beta attenuates doxorubicin-induced alterations of excitation-contraction coupling and reduces oxidative stress in adult rat cardiomyocytes. J. Mol. Cell. Cardiol. 2006, 41, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Bruno, S.; Grange, C.; Collino, F.; Deregibus, M.C.; Cantaluppi, V.; Biancone, L.; Tetta, C.; Camussi, G. Microvesicles derived from mesenchymal stem cells enhance survival in a lethal model of acute kidney injury. PLoS ONE 2012, 7, e33115. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, N.; Ahmed, I.A.; Hussien, N.I.; Dessouky, A.A.; Farid, A.S.; Elshazly, A.M.; Mostafa, O.; Gazzar, W.B.E.; Sorour, S.M.; Seleem, Y.; et al. Mesenchymal Stem Cell-Derived Exosomes Ameliorated Diabetic Nephropathy by Autophagy Induction through the mTOR Signaling Pathway. Cells 2018, 7, 226. [Google Scholar] [CrossRef] [PubMed]
- Barile, L.; Lionetti, V.; Cervio, E.; Matteucci, M.; Gherghiceanu, M.; Popescu, L.M.; Torre, T.; Siclari, F.; Moccetti, T.; Vassalli, G. Extracellular vesicles from human cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function after myocardial infarction. Cardiovasc. Res. 2014, 103, 530–541. [Google Scholar] [CrossRef]
- Gatti, S.; Bruno, S.; Deregibus, M.C.; Sordi, A.; Cantaluppi, V.; Tetta, C.; Camussi, G. Microvesicles derived from human adult mesenchymal stem cells protect against ischaemia-reperfusion-induced acute and chronic kidney injury. Nephrol. Dial. Transplant. 2011, 26, 1474–1483. [Google Scholar] [CrossRef]
- Lange-Consiglio, A.; Perrini, C.; Albini, G.; Modina, S.; Lodde, V.; Orsini, E.; Esposti, P.; Cremonesi, F. Oviductal microvesicles and their effect on in vitro maturation of canine oocytes. Reproduction 2017, 154, 167–180. [Google Scholar] [CrossRef]
- Riegler, J.; Liew, A.; Hynes, S.O.; Ortega, D.; O’Brien, T.; Day, R.M.; Richards, T.; Sharif, F.; Pankhurst, Q.A.; Lythgoe, M.F. Superparamagnetic iron oxide nanoparticle targeting of MSCs in vascular injury. Biomaterials 2013, 34, 1987–1994. [Google Scholar] [CrossRef]
- Altanerova, U.; Babincova, M.; Babinec, P.; Benejova, K.; Jakubechova, J.; Altanerova, V.; Zduriencikova, M.; Repiska, V.; Altaner, C. Human mesenchymal stem cell-derived iron oxide exosomes allow targeted ablation of tumor cells via magnetic hyperthermia. Int. J. Nanomed. 2017, 12, 7923–7936. [Google Scholar] [CrossRef]
- Bancroft’s Theory and Practice of Histological Techniques; Elsevier: Amsterdam, The Netherlands, 2019; Volume 27, pp. 568–579. Available online: https://linkinghub.elsevier.com/retrieve/pii/C20150001435 (accessed on 16 April 2022).
- Milano, G.; Raucci, A.; Scopece, A.; Daniele, R.; Guerrini, U.; Sironi, L.; Cardinale, D.; Capogrossi, M.C.; Pompilio, G. Doxorubicin and trastuzumab regimen induces biventricular failure in mice. J. Am. Soc. Echocardiogr. 2014, 27, 568–579. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Wang, F.; Yang, J.; Sun, J.; Dong, Y.; Zhao, H.; Shi, H.; Fu, L. Testosterone replacement attenuates mitochondrial damage in a rat model of myocardial infarction. J. Endocrinol. 2015, 225, 101–111. [Google Scholar] [CrossRef]
- Hussien, N.I.; El-Wakeel, H.S.; Souror, S.M.; Ahmed, I.A. Alleviation of cardiac mitochondrial dysfunction and oxidative stress underlies the protective effect of vitamin D in chronic stress-induced cardiac dysfunction in rats. Gen. Physiol. Biophys. 2019, 38, 51–61. [Google Scholar] [CrossRef] [PubMed]
- El-Attrouny, M.M.; Iraqi, M.M.; Sabike, I.I.; Abdelatty, A.M.; Moustafa, M.M.; Badr, O.A. Comparative evaluation of growth performance, carcass characteristics and timed series gene expression profile of GH and IGF-1 in two Egyptian indigenous chicken breeds versus Rhode Island Red. J. Anim. Breed. Genet. 2021, 138, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Buch, L.; Lipi, B.; Langhnoja, J.; Jaldeep, L.; Pillai, P.P.; Prakash, P. Role of astrocytic MeCP2 in regulation of CNS myelination by affecting oligodendrocyte and neuronal physiology and axo-glial interactions. Exp. Brain Res. 2018, 236, 3015–3027. [Google Scholar] [CrossRef]
- Hausburg, F.; Jung, J.J.; Hoch, M.; Wolfien, M.; Yavari, A.; Rimmbach, C.; David, R. (Re-)programming of subtype specific cardiomyocytes. Adv. Drug Deliv. Rev. 2017, 120, 142–167. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.H.; Ramasamy, R.; Yong, Y.K.; Ngalim, S.H.; Lim, V.; Shaharuddin, B.; Tan, J.J. Extracellular matrix from decellularized mesenchymal stem cells improves cardiac gene expressions and oxidative resistance in cardiac C-kit cells. Regen. Ther. 2019, 11, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Garcia, O.; Baumgartner, S.; Macri-Pellizzeri, L.; Rodriguez-Madoz, J.R.; Abizanda, G.; Guruceaga, E.; Albiasu, E.; Corbacho, D.; Benavides-Vallve, C.; Soriano-Navarro, M.; et al. Neuregulin-1β induces mature ventricular cardiac differentiation from induced pluripotent stem cells contributing to cardiac tissue repair. Stem Cells Dev. 2015, 24, 484–496. [Google Scholar] [CrossRef]
- Mantawy, E.M.; Esmat, A.; El-Bakly, W.M.; Salah ElDin, R.A.; El-Demerdash, E. Mechanistic clues to the protective effect of chrysin against doxorubicin-induced cardiomyopathy: Plausible roles of p53, MAPK and AKT pathways. Sci. Rep. 2017, 7, 4795. [Google Scholar] [CrossRef]
- Brown, J.E.P.; Dunmore, S.J. Leptin decreases apoptosis and alters BCL-2 : Bax ratio in clonal rodent pancreatic beta-cells. Diabetes Metab. Res. Rev. 2007, 23, 497–502. [Google Scholar] [CrossRef]
- Manzano-Moreno, F.J.; Illescas-Montes, R.; Melguizo-Rodriguez, L.; Costela-Ruiz, V.J.; García-Martínez, O.; Ruiz, C.; Ramos-Torrecillas, J. Impact of bisphosphonates on the proliferation and gene expression of human fibroblasts. Int. J. Med. Sci. 2019, 16, 1534–1540. [Google Scholar] [CrossRef]
- Khani, F.; Thaler, R.; Paradise, C.R.; Deyle, D.R.; Kruijthof-de Julio, M.; Galindo, M.; Gordon, J.A.; Stein, G.S.; Dudakovic, A.; van Wijnen, A.J. Histone H4 Methyltransferase Suv420h2 Maintains Fidelity of Osteoblast Differentiation. J. Cell. Biochem. 2017, 118, 1262–1272. [Google Scholar] [CrossRef]
- Lucas, S.; Omata, Y.; Hofmann, J.; Böttcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Krönke, G.; et al. Short-chain fatty acids regulate systemic bone mass and protect from pathological bone loss. Nat. Commun. 2018, 9, 55. [Google Scholar] [CrossRef]
- Lipskaia, L.; Keuylian, Z.; Blirando, K.; Mougenot, N.; Jacquet, A.; Rouxel, C.; Sghairi, H.; Elaib, Z.; Blaise, R.; Adnot, S.; et al. Expression of sarco (endo) plasmic reticulum calcium ATPase (SERCA) system in normal mouse cardiovascular tissues, heart failure and atherosclerosis. Biochim. Biophys. Acta 2014, 1843, 2705–2718. [Google Scholar] [CrossRef]
- Rahal, O.M.; Simmen, R.C.M. PTEN and p53 cross-regulation induced by soy isoflavone genistein promotes mammary epithelial cell cycle arrest and lobuloalveolar differentiation. Carcinogenesis 2010, 31, 1491–1500. [Google Scholar] [CrossRef]
- Dong, Y.; Han, L.L.; Xu, Z.X. Suppressed microRNA-96 inhibits iNOS expression and dopaminergic neuron apoptosis through inactivating the MAPK signaling pathway by targeting CACNG5 in mice with Parkinson’s disease. Mol. Med. 2018, 24, 61. [Google Scholar] [CrossRef]
- Badr, O.A.; El-Shawaf, I.I.S.; El-Garhy, H.S.; Moustafa, M.M.A.; Ahmed-Farid, O.A. The potent therapeutic effect of novel cyanobacterial isolates against oxidative stress damage in redox rats. J. Appl. Microbiol. 2019, 126, 1278–1289. [Google Scholar] [CrossRef]
- Chen, Y.; Tian, Z.; He, L.; Liu, C.; Wang, N.; Rong, L.; Liu, B. Exosomes derived from miR-26a-modified MSCs promote axonal regeneration via the PTEN/AKT/mTOR pathway following spinal cord injury. Stem Cell Res. Ther. 2021, 12, 224. [Google Scholar] [CrossRef]
- Zhou, X.H.; Chai, H.X.; Bai, M.; Zhang, Z. LncRNA-GAS5 regulates PDCD4 expression and mediates myocardial infarction-induced cardiomyocytes apoptosis via targeting MiR-21. Cell Cycle 2020, 19, 1363–1377. [Google Scholar] [CrossRef]
- Ebrahim, N.; Al Saihati, H.A.; Shaman, A.; Dessouky, A.A.; Farid, A.S.; Hussien, N.I.; Mostafa, O.; Seleem, Y.; Sabry, D.; Saad, A.S.; et al. Bone marrow-derived mesenchymal stem cells combined with gonadotropin therapy restore postnatal oogenesis of chemo-ablated ovaries in rats via enhancing very small embryonic-like stem cells. Stem Cell Res. Ther. 2021, 12, 517. [Google Scholar] [CrossRef]
- Ebrahim, N.; Badr, O.A.; Yousef, M.M.; Hassouna, A.; Sabry, D.; Farid, A.S.; Mostafa, O.; Saihati, H.A.A.; Seleem, Y.; Abd El Aziz, E.; et al. Functional Recellularization of Acellular Rat Liver Scaffold by Induced Pluripotent Stem Cells: Molecular Evidence for Wnt/B-Catenin Upregulation. Cells 2021, 10, 2819. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques E-Book; Elsevier: Philadelphia, PA, USA, 2018; Available online: https://public.ebookcentral.proquest.com/choice/publicfullrecord.aspx?p=5321317 (accessed on 16 April 2022).
- Suvarna, S.K.; Layton CBancroft John, D. Bancroft’s theory and practice of histological techniques. 2019. Available online: https://search.ebscohost.com/login.aspx?direct=true&scope=site&db=nlebk&db=nlabk&AN=1730520 (accessed on 16 April 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | LVDD (cm) | LVSD (cm) | EF% | FS% |
|---|---|---|---|---|
| I | 7.2 | 3.78 | 85.77 | 47.77 |
| II | 7.7 a | 5.43 a | 65.92 a | 30.33 a |
| III | 7.2 b | 3.8 b,c | 85.28 b,c | 47.27 b,c |
| IV | 7.3 b | 4.23 a,b | 80.83 a,b | 42.18 a,b |
| Target Genes | Primer Sequence 5’ to 3’ | References |
|---|---|---|
| Nrg-1 | 5′-AGCAGACACCAGCTTCAGAC-3′ 5′-CAAGAAGGCAGGGGACCAAA-3′ | [79] |
| cTnnc1 | 5′-AGCCTGTCCTGTGAGCTGTC-3′ 5′-GGCAGCCTTGAACTCATTCT-3′ | [80] |
| BMHC | 5′-GCCAACACCAACCTGTCCAAGTTC-3′ 5′-TGCAAAGGCTCCAGGTCTGAGGGC-3′ | [59] |
| Mlc2v | 5′-ACTATGTCCGGGAGATGCTG-3′ 5′-TGGGTAATGATGTGGACCAA-3′ | [81] |
| Bax | 5′-GATCAGCTCGGGCACTTTA-3 5′-TGTTTGCTGATGGCAA CTTC-3′ | [82] |
| Bcl-2 | 5′-AGGAT TGTGG CCTTC TTTGA GT-3′ 5′-GCCG GTTC AGG TACT CAGT CAT-3′ | [83] |
| Puma | 5′- GTG TGG AGG AGGAGG AGT GG-3′ 5′-TCG GTG TCG ATG TTG CTC TT-3′ | [81] |
| CTGF | 5’-CCTGGTCCAGACCACAGAGT-3′ 5’-TTTTCCTCCAGGTCAGCTTC-3′ | [84] |
| Col1a1 | 5′-CCTCAGGGTATTGCTGGACAAC-3′ 5′-CAGAAGGACCTTGTTTGCCAGG-3′ | [85] |
| Mmp9 | 5′-GCTGACTACGATAAGGACGGCA-3′ 5′-TAGTGGTGCAGGCAGAGTAGGA-3′ | [86] |
| SERCA2a | 5′-CTCCATCTGCTTGTCCAT-3′ 5′-GCGGTTACTCCAGTATTG-3′ | [87] |
| PTEN | 5′-CAATGTTCAGTGGCGGAACTT-3′ 5′-GGCAATGGCTGAGGGAACT-3′ | [88] |
| iNOS | 5′-ATGGACCAGTATAAGGCAAGC-3′ 5′- GCTCTGGATGAGCCTATATTG-3′ | [89] |
| GAPDH | 5′-AGT TCA ACG GCA CAG TCA A -3′ 5′-TAC TCA GCA CCA GCA TCA CC -3′ | [90] |
| miR-26a | 5′-UUCAAGUAAUCCAGGAUAGGCU-3′ 5′-AGCCUAUCCUGGAUUACUUGAA-3′ | [91] |
| miR-21 | 5′-CGCGCTAGCTTATCAGACTGA-3′ 5′-CGGCCCAGTGTTCAGACTAC-3′ | [92] |
| U6 | 5′-CTCGCTTCGGCAGCACA-3′ 5′-AACGCTTCACGAATTTGCGT-3′ | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebrahim, N.; Al Saihati, H.A.; Mostafa, O.; Hassouna, A.; Abdulsamea, S.; Abd El Aziz M. El Gebaly, E.; Abo-Rayah, N.H.; Sabry, D.; El-Sherbiny, M.; Madboly, A.G.; et al. Prophylactic Evidence of MSCs-Derived Exosomes in Doxorubicin/Trastuzumab-Induced Cardiotoxicity: Beyond Mechanistic Target of NRG-1/Erb Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 5967. https://doi.org/10.3390/ijms23115967
Ebrahim N, Al Saihati HA, Mostafa O, Hassouna A, Abdulsamea S, Abd El Aziz M. El Gebaly E, Abo-Rayah NH, Sabry D, El-Sherbiny M, Madboly AG, et al. Prophylactic Evidence of MSCs-Derived Exosomes in Doxorubicin/Trastuzumab-Induced Cardiotoxicity: Beyond Mechanistic Target of NRG-1/Erb Signaling Pathway. International Journal of Molecular Sciences. 2022; 23(11):5967. https://doi.org/10.3390/ijms23115967
Chicago/Turabian StyleEbrahim, Nesrine, Hajir A. Al Saihati, Ola Mostafa, Amira Hassouna, Sameh Abdulsamea, Eman Abd El Aziz M. El Gebaly, Nashwa Hassan Abo-Rayah, Dina Sabry, Mohamed El-Sherbiny, Abdelmonem G. Madboly, and et al. 2022. "Prophylactic Evidence of MSCs-Derived Exosomes in Doxorubicin/Trastuzumab-Induced Cardiotoxicity: Beyond Mechanistic Target of NRG-1/Erb Signaling Pathway" International Journal of Molecular Sciences 23, no. 11: 5967. https://doi.org/10.3390/ijms23115967
APA StyleEbrahim, N., Al Saihati, H. A., Mostafa, O., Hassouna, A., Abdulsamea, S., Abd El Aziz M. El Gebaly, E., Abo-Rayah, N. H., Sabry, D., El-Sherbiny, M., Madboly, A. G., Hussien, N. I., Saadani, R. E. H., Ebrahim, H. A., Badr, O. A. M., Elsherbiny, N. M., & Salim, R. F. (2022). Prophylactic Evidence of MSCs-Derived Exosomes in Doxorubicin/Trastuzumab-Induced Cardiotoxicity: Beyond Mechanistic Target of NRG-1/Erb Signaling Pathway. International Journal of Molecular Sciences, 23(11), 5967. https://doi.org/10.3390/ijms23115967