
Association Mechanism and Conformational Changes in Trypsin on Its Interaction with Atrazine: A Multi- Spectroscopic and Biochemical Study with Computational Approach

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. Instrumentations
2.4. Methods
2.4.1. UV-Vis Spectral Measurements
2.4.2. Steady-State Fluorescence Spectroscopy
2.4.3. Synchronous Fluorescence Spectroscopy (SFS) Experiments
2.4.4. Circular Dichroism (CD) Spectroscopy Measurements
2.4.5. Fourier Transform Infrared Spectroscopic (FTIR) Analysis
2.4.6. Computational Analysis
2.4.7. Measurement of Trypsin Activity
3. Results and Discussion
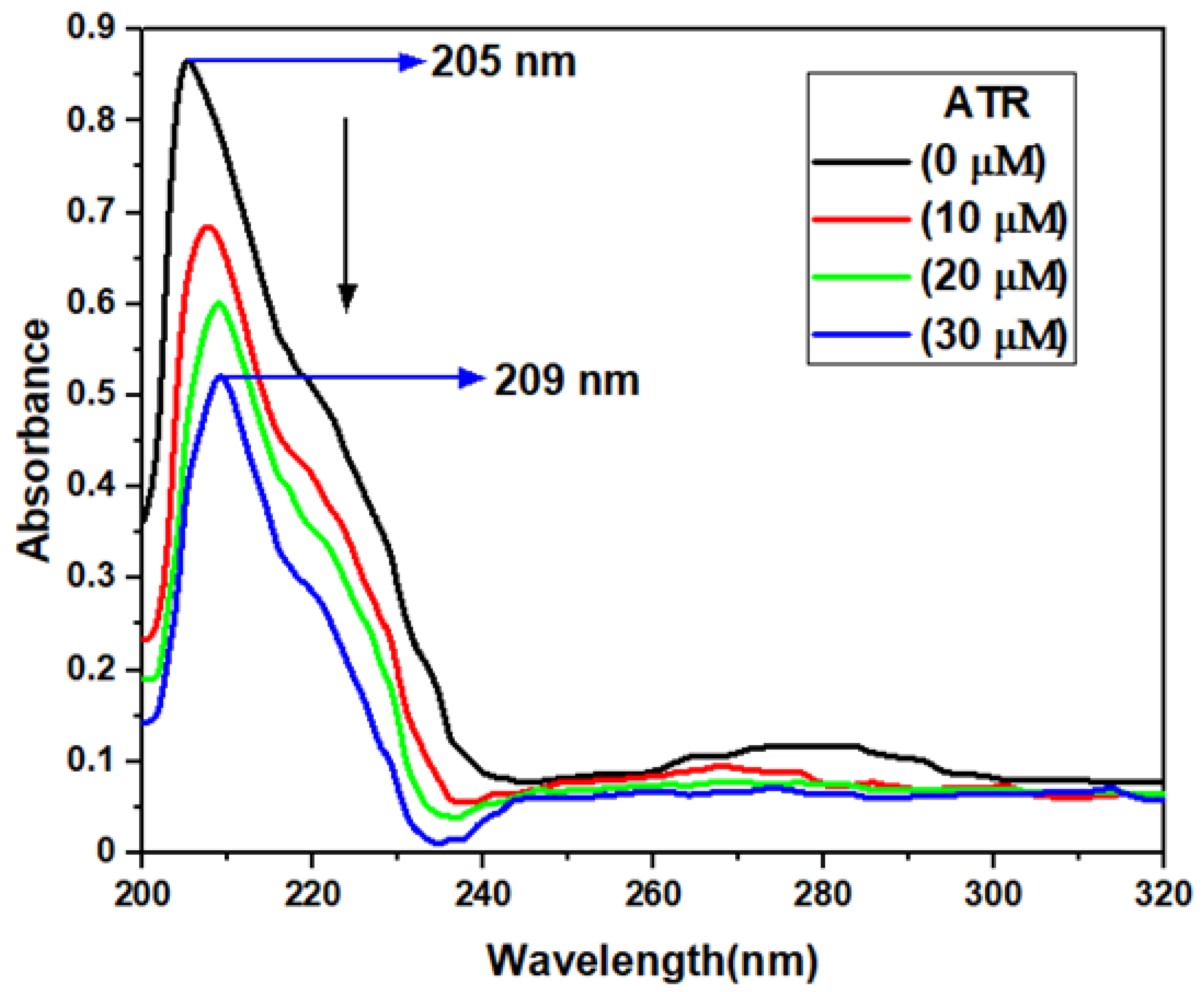
3.1. UV-Vis Absorption Spectroscopy Investigations
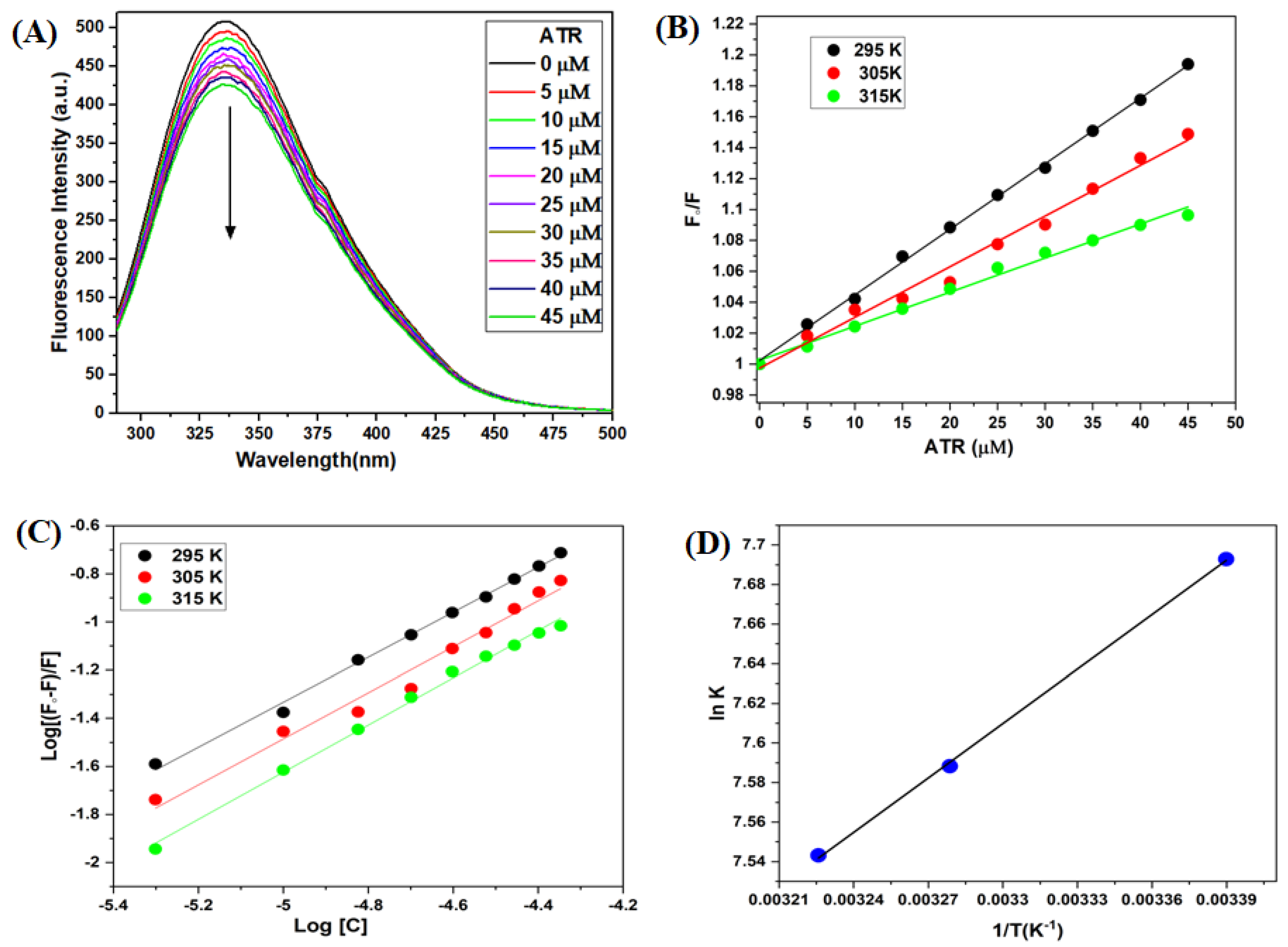
3.2. Fluorescence Emission Spectroscopy of TYP-ATR Complex
3.2.1. Fluorescence Quenching Mechanism
3.2.2. Binding Constant Evaluation
3.2.3. Determination of the Binding Forces between TYP and ATR
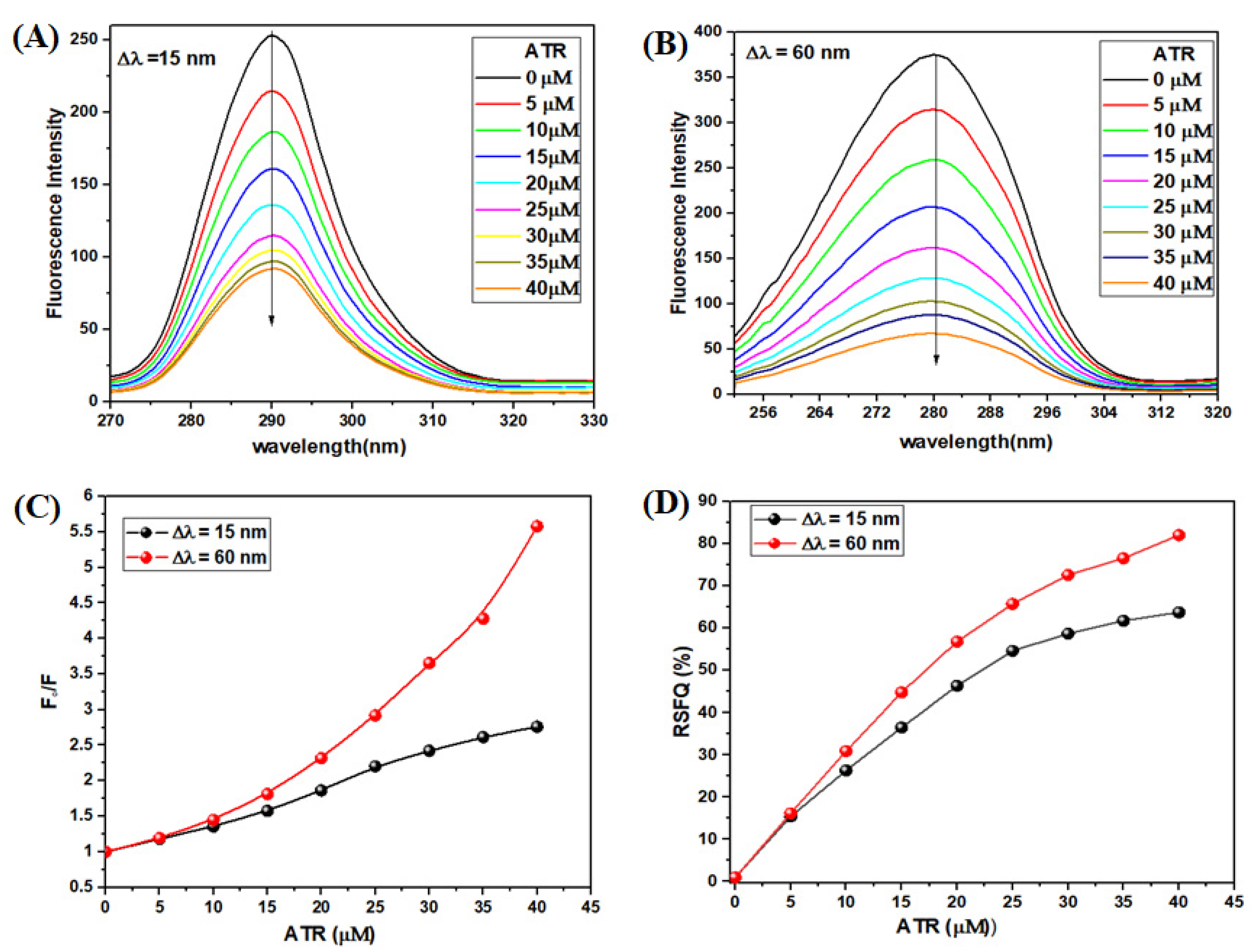
3.2.4. Synchronous Fluorescence Spectroscopy (SFS) Experiment
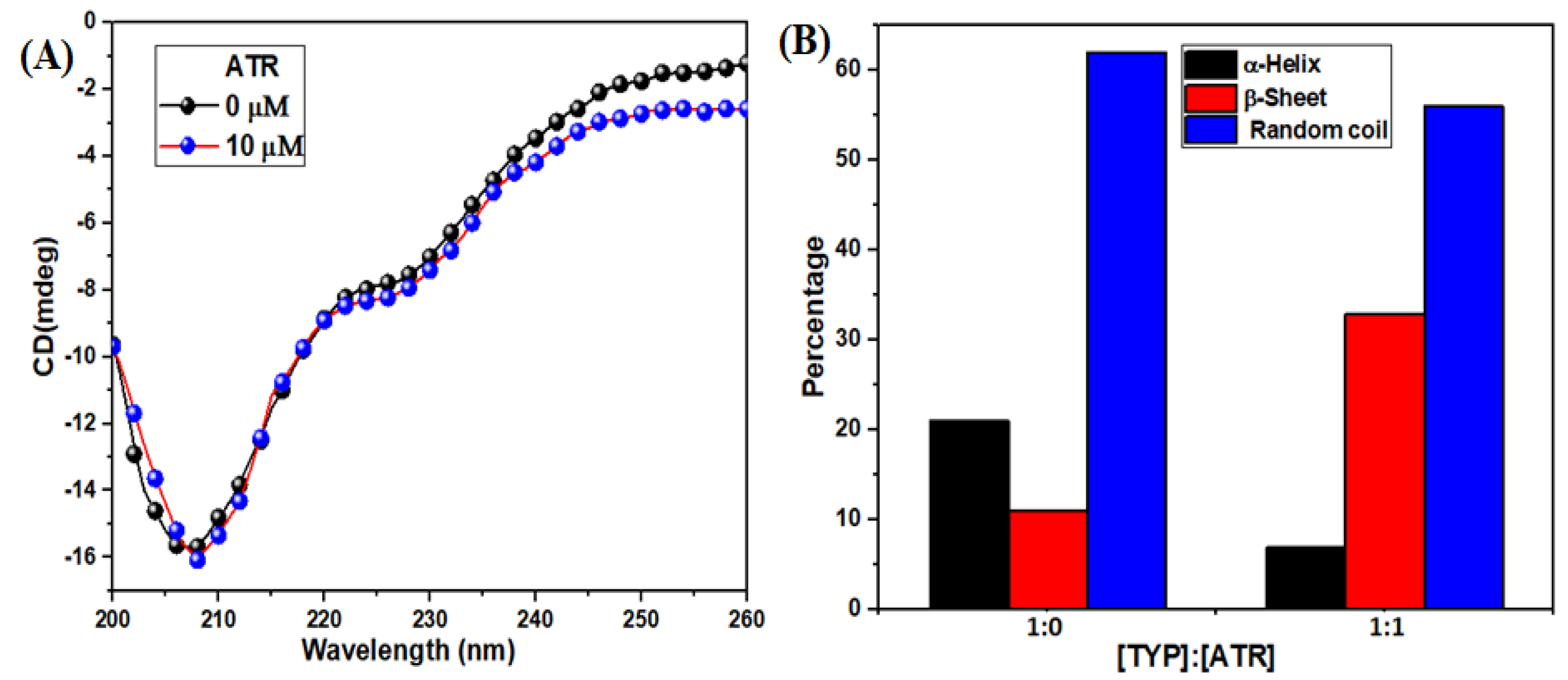
3.2.5. Circular Dichroism (CD) Spectra Changes in TYP upon ATR Binding
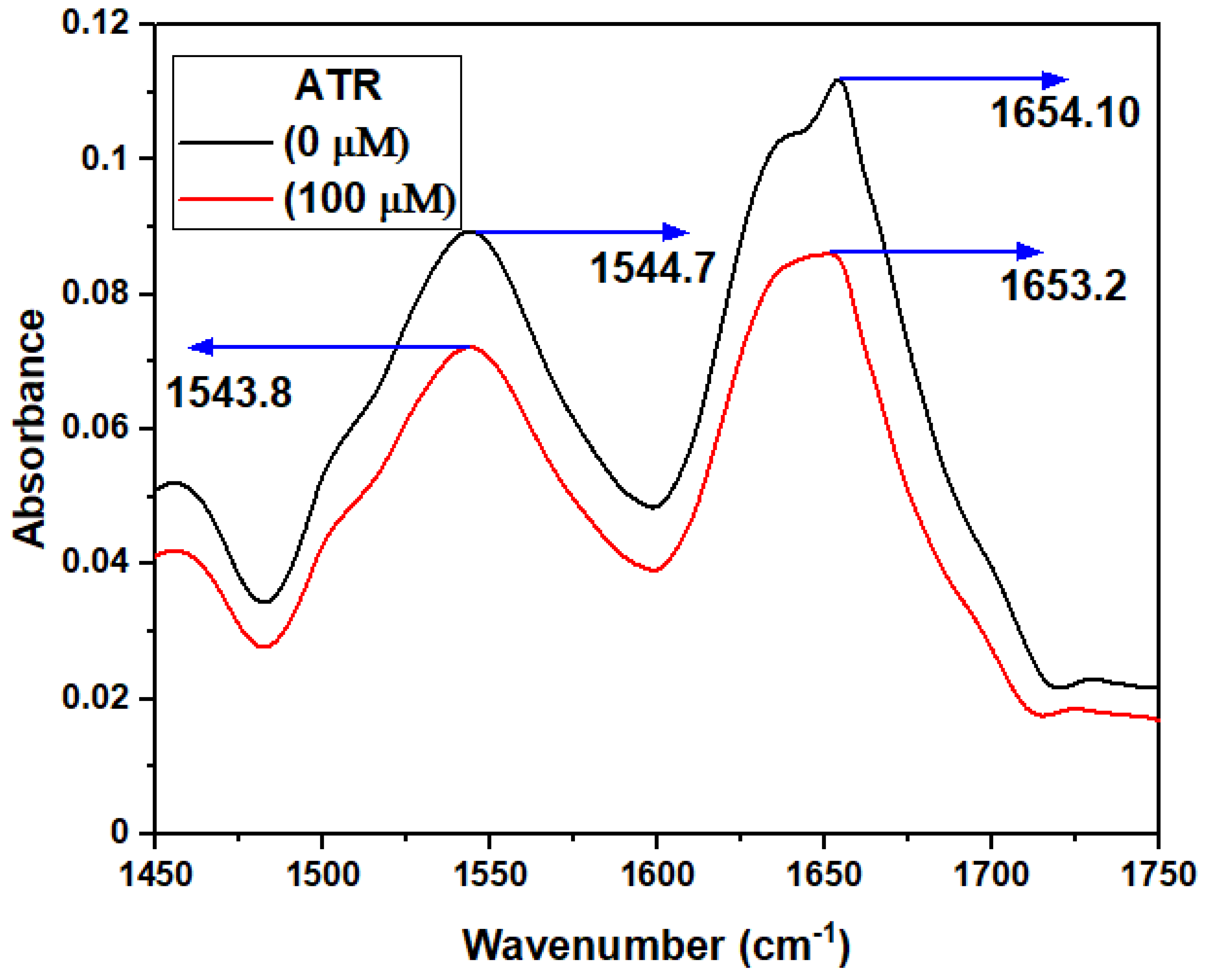
3.2.6. FT-IR Spectroscopy
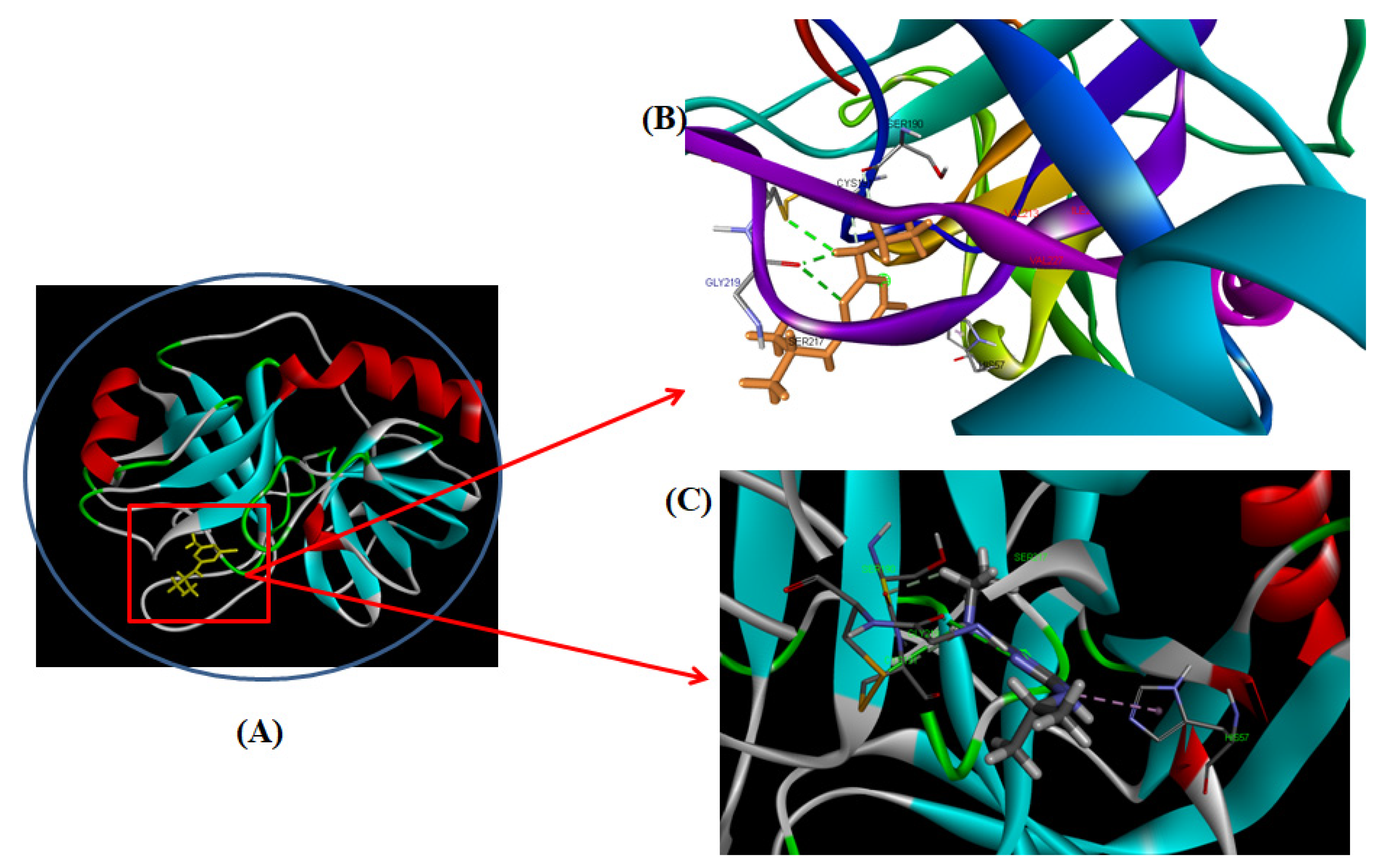
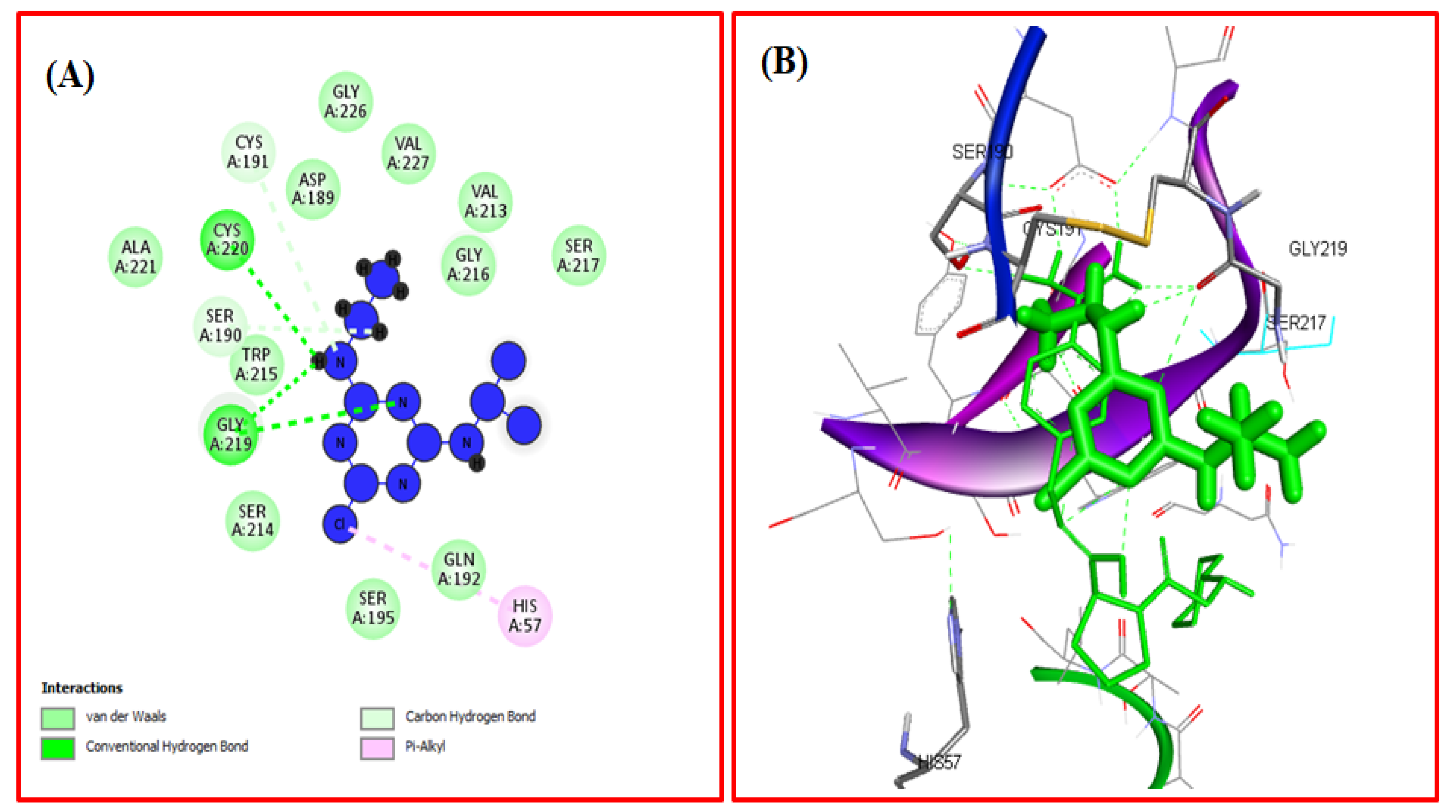
3.2.7. Computational Modeling of the TYP-ATR Complex
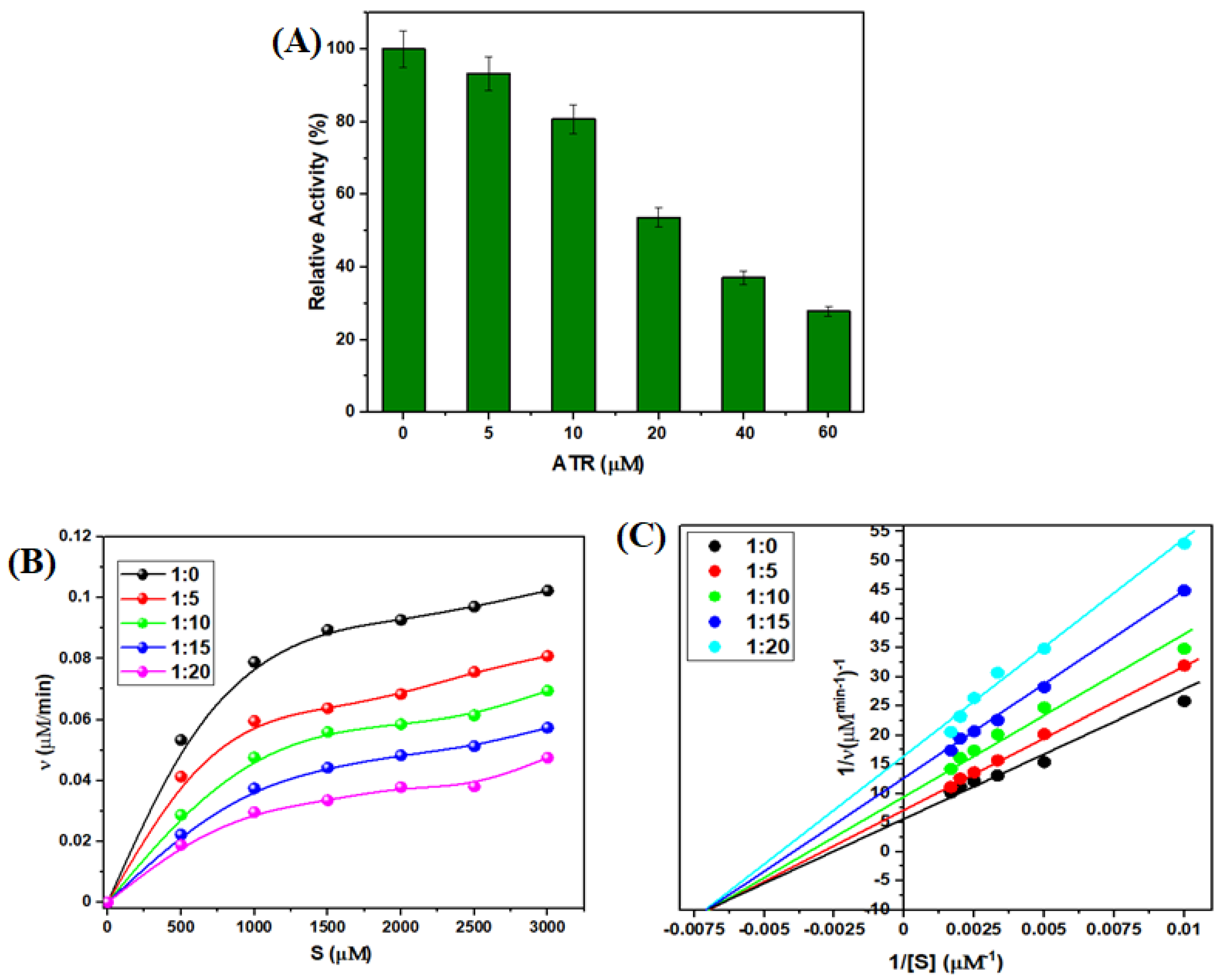
3.2.8. Effect of ATR on TYP Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gianessi, L.P. The increasing importance of herbicides in worldwide crop production. Pest Manag. Sci. 2013, 69, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Kudsk, P.; Streibig, J. Herbicides-a two-edged sword. Weed Res. 2003, 43, 90–102. [Google Scholar] [CrossRef]
- Gupta, P.K. Toxicity of Herbicides, Veterinary Toxicology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 553–567. [Google Scholar]
- Mesnage, R.; Bernay, B.; Séralini, G.-E. Ethoxylated adjuvants of glyphosate-based herbicides are active principles of human cell toxicity. Toxicology 2013, 313, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Marin-Morales, M.A.; Ventura-Camargo, B.D.C.; Hoshina, M.M. Toxicity of herbicides: Impact on aquatic and soil biota and human health. In Herbicides-Current Research and Case Studies in Use; Intechopen: London, UK, 2013; pp. 399–443. [Google Scholar]
- Ahmad, A.; Zafar, A.; Zargar, S.; Bazgaifan, A.; Wani, T.A.; Ahmad, M. Protective effects of apigenin against edifenphos-induced genotoxicity and cytotoxicity in rat hepatocytes. J. Biomol. Struct. Dyn. 2021, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Alavanja, M. Pesticides use and exposure extensive worldwide. Rev. Environ. Health 2009, 24, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.; Boerboom, C.M.; Rabaey, T.L. Significance of Atrazine in Sweet Corn Weed Management Systems. Weed Technol. 2010, 24, 139–142. [Google Scholar] [CrossRef]
- Buchanan, G.A.; Hiltbold, A.E. Performance and Persistence of Atrazine. Weed Sci. 1973, 21, 413–416. [Google Scholar] [CrossRef]
- Comber, S.D.W. Abiotic persistence of atrazine and simazine in water. Pestic. Sci. 1999, 55, 696–702. [Google Scholar] [CrossRef]
- Schwab, A.P.; Splichal, P.A.; Banks, M.K. Persistence of Atrazine and Alachlor in Ground Water Aquifers and Soil. Water Air Soil Pollut. 2006, 171, 203–235. [Google Scholar] [CrossRef]
- Pathak, R.K.; Dikshit, A.K. Atrazine and human health. Int. J. Ecosyst. 2011, 1, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Giusi, G.; Facciolo, R.M.; Canonaco, M.; Alleva, E.; Belloni, V.; Dessi’-Fulgheri, F.; Santucci, D. The endocrine disruptor atrazine accounts for a dimorphic somatostatinergic neuronal expression pattern in mice. Toxicol. Sci. 2006, 89, 257–2644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, T. More feedback on whether atrazine is a potent endocrine disruptor chemical. Environ. Sci. Technol.-Columb. 2009, 43, 6115. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, J.C.; Stevens, J.T.; Breckenridge, C.B. Atrazine interaction with estrogen expression systems. Rev. Environ. Contam. Toxicol. 2008, 196, 147–160. [Google Scholar] [PubMed]
- McMullin, T.S.; Andersen, M.E.; Nagahara, A.; Lund, T.D.; Pak, T.; Handa, R.J.; Hanneman, W.H. Evidence that atrazine and diaminochlorotriazine inhibit the estrogen/progesterone induced surge of luteinizing hormone in female Sprague-Dawley rats without changing estrogen receptor action. Toxicol. Sci. 2004, 79, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Xing, H.; Wang, C.; Wu, H.; Chen, D.; Li, S.; Xu, S. Effects of atrazine and chlorpyrifos on DNA methylation in the brain and gonad of the common carp. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 168, 11–19. [Google Scholar] [CrossRef]
- Wang, F.; Yang, Q.-W.; Zhao, W.-J.; Du, Q.-Y.; Chang, Z.-J. Effects of short-time exposure to atrazine on miRNA expression profiles in the gonad of common carp (Cyprinus carpio). BMC Genom. 2019, 20, 587. [Google Scholar] [CrossRef] [Green Version]
- Karami-Mohajeri, S.; Abdollahi, M. Toxic influence of organophosphate, carbamate, and organochlorine pesticides on cellular metabolism of lipids, proteins, and carbohydrates. Hum. Exp. Toxicol. 2011, 30, 1119–1140. [Google Scholar] [CrossRef]
- Sharma, A.K.; Gaur, K.; Tiwari, R.K.; Gaur, M.S. Computational interaction analysis of organophosphorus pesticides with different metabolic proteins in humans. J. Biomed. Res. 2011, 25, 335–347. [Google Scholar] [CrossRef] [Green Version]
- Wani, T.A.; Alsaif, N.; Bakheit, A.H.; Zargar, S.; Al-Mehizia, A.A.; Khan, A.A. Interaction of an abiraterone with calf thymus DNA: Investigation with spectroscopic technique and modelling studies. Bioorg. Chem. 2020, 100, 103957. [Google Scholar] [CrossRef]
- Shah, D.; Mital, K. The Role of Trypsin:Chymotrypsin in Tissue Repair. Adv. Ther. 2018, 35, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Luo, W.; Reiser, G. Reiser, Trypsin and trypsin-like proteases in the brain: Proteolysis and cellular functions. Cell. Mol. Life Sci. 2008, 65, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Whitcomb, D.C.; Lowe, M.E. Human pancreatic digestive enzymes. Dig. Dis. Sci. 2007, 52, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pillai, J.R.; Wali, A.F.; Menezes, G.A.; Rehman, M.U.; Wani, T.A.; Arafah, A.; Zargar, S.; Mir, T.M. Chemical Composition Analysis, Cytotoxic, Antimicrobial and Antioxidant Activities of Physalis angulata L. A Comparative Study of Leaves and Fruit. Molecules 2022, 27, 1480. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, R.; Qin, P.; Liu, R. Assessing the potential toxic effect of one persistent organic pollutant: Non-covalent interaction of dicofol with the enzyme trypsin. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2012, 89, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huo, M.; Zhao, L.; Cao, Z.; Xu, M.; Wan, J.; Niu, Q.; Huo, C.; Tang, J.; Liu, R. Study of the effects of ultrafine carbon black on the structure and function of trypsin. J. Mol. Recognit. 2020, 34, e2874. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Liang, J.; Luo, H.; Li, H.; Yang, J.; Huang, S. Investigations of conformational structures and activities of trypsin and pepsin affected by food colourant allura red. J. Mol. Liq. 2020, 319, 114359. [Google Scholar] [CrossRef]
- Huang, S.; Li, H.; Liu, Y.; Yang, L.; Wang, D.; Xiao, Q. Investigations of conformational structure and enzymatic activity of trypsin after its binding interaction with graphene oxide. J. Hazard. Mater. 2020, 392, 122285. [Google Scholar] [CrossRef]
- Harish, B.S.; Uppuluri, K.B. Potential Anticoagulant Activity of Trypsin Inhibitor Purified from an Isolated Marine Bacterium Oceanimonas Sp. BPMS22 and its Kinetics. Mar. Biotechnol. 2018, 20, 780–791. [Google Scholar] [CrossRef]
- Schmid, F.X. Biological Macromolecules: UV-Visible Spectrophotometry; e LS: Hoboken, NJ, USA, 2001. [Google Scholar]
- Anthis, N.J.; Clore, G.M. Sequence-specific determination of protein and peptide concentrations by absorbance at 205 nm. Protein Sci. 2013, 22, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-Q.; Zhang, H.-M. Effects of Bisphenol S on the Structures and Activities of Trypsin and Pepsin. J. Agric. Food Chem. 2014, 62, 11303–11311. [Google Scholar] [CrossRef]
- Wani, T.A.; Alsaif, N.A.; Alanazi, M.M.; Bakheit, A.H.; Khan, A.A.; Zargar, S. Binding of colchicine and ascorbic acid (vitamin C) to bovine serum albumin: An in-vitro interaction study using multispectroscopic, molecular docking and molecular dynamics simulation study. J. Mol. Liq. 2021, 342, 117542. [Google Scholar] [CrossRef]
- Zargar, S.; Wani, T.A. Exploring the binding mechanism and adverse toxic effects of persistent organic pollutant (dicofol) to human serum albumin: A biophysical, biochemical and computational approach. Chem.-Bio. Interact. 2021, 350, 109707. [Google Scholar] [CrossRef] [PubMed]
- Wani, T.A.; Bakheit, A.H.; Al-Majed, A.A.; Altwaijry, N.; Baquaysh, A.; Aljuraisy, A.; Zargar, S. Binding and drug displacement study of colchicine and bovine serum albumin in presence of azithromycin using multispectroscopic techniques and molecular dynamic simulation. J. Mol. Liq. 2021, 333, 115934. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Zargar, S.; Wani, T.A.; Alsaif, N.A.; Khayyat, A.I.A. A Comprehensive Investigation of Interactions between Antipsychotic Drug Quetiapine and Human Serum Albumin Using Multi-Spectroscopic, Biochemical, and Molecular Modeling Approaches. Molecules 2022, 27, 2589. [Google Scholar] [CrossRef] [PubMed]
- Wani, T.A.; Alsaif, N.; Alanazi, M.M.; Bakheit, A.H.; Zargar, S.; Bhat, M.A. A potential anticancer dihydropyrimidine derivative and its protein binding mechanism by multispectroscopic, molecular docking and molecular dynamic simulation along with its in-silico toxicity and metabolic profile. Eur. J. Pharm. Sci. 2021, 158, 105686. [Google Scholar] [CrossRef] [PubMed]
- Mokaberi, P.; Babayan-Mashhadi, F.; Zadeh, Z.A.T.; Saberi, M.R.; Chamani, J. Analysis of the interaction behavior between Nano-Curcumin and two human serum proteins: Combining spectroscopy and molecular stimulation to understand protein-protein interaction. J. Biomol. Struct. Dyn. 2021, 39, 3358–3377. [Google Scholar] [CrossRef]
- Alsaif, N.A.; Al-Mehizia, A.A.; Bakheit, A.H.; Zargar, S.; Wani, T.A. A Spectroscopic, Thermodynamic and Molecular Docking Study of the Binding Mechanism of Dapoxetine with Calf Thymus DNA. S. Afr. J. Chem. 2020, 73, 44–50. [Google Scholar] [CrossRef]
- Gerbanowski, A.; Rabiller, C.; Larré, C.; Guéguen, J. Grafting of Aliphatic and Aromatic Probes on Bovine Serum Albumin: Influence on Its Structural and Physicochemical Characteristics. J. Protein Chem. 1999, 18, 325–336. [Google Scholar] [CrossRef]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, G.; Liao, Y.; Wang, Y. Binding characteristics of psoralen with trypsin: Insights from spectroscopic and molecular modeling studies. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 151, 498–505. [Google Scholar] [CrossRef]
- Lloyd, J.B.F. Synchronized Excitation of Fluorescence Emission Spectra. Nat. Phys. Sci. 1971, 231, 64–65. [Google Scholar] [CrossRef]
- Kou, S.-B.; Lin, Z.-Y.; Wang, B.-L.; Shi, J.-H.; Liu, Y.-X. Evaluation of the binding behavior of olmutinib (HM61713) with model transport protein: Insights from spectroscopic and molecular docking studies. J. Mol. Struct. 2021, 1224, 129024. [Google Scholar] [CrossRef]
- Ranjbar, S.; Shokoohinia, Y.; Ghobadi, S.; Bijari, N.; Gholamzadeh, S.; Moradi, N.; Ashrafi-Kooshk, M.R.; Aghaei, A.; Khodarahmi, R. Studies of the Interaction between Isoimperatorin and Human Serum Albumin by Multispectroscopic Method: Identification of Possible Binding Site of the Compound Using Esterase Activity of the Protein. Sci. World, J. 2013, 2013, 305081. [Google Scholar] [CrossRef] [PubMed]
- Wani, T.A.; Bakheit, A.H.; Zargar, S.; Khayyat, A.I.A.; Al-Majed, A.A. Influence of Rutin, Sinapic Acid, and Naringenin on Binding of Tyrosine Kinase Inhibitor Erlotinib to Bovine Serum Albumin Using Analytical Techniques Along with Computational Approach. Appl. Sci. 2022, 12, 3575. [Google Scholar] [CrossRef]
- Johnson, W.C., Jr. Secondary structure of proteins through circular dichroism spectroscopy. Annu. Rev. Biophys. Biophys. Chem. 1988, 17, 145–166. [Google Scholar] [CrossRef]
- Sarkar, D.; Mahata, A.; Das, P.; Girigoswami, A.; Ghosh, D.; Chattopadhyay, N. Deciphering the perturbation of serum albumins by a ketocyanine dye: A spectroscopic approach. J. Photochem. Photobiol. B Biol. 2009, 96, 136–143. [Google Scholar] [CrossRef]
- Wani, T.A.; Bakheit, A.H.; Zargar, S.; Alanazi, Z.S.; Al-Majed, A.A. Influence of antioxidant flavonoids quercetin and rutin on the in-vitro binding of neratinib to human serum albumin. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 246, 118977. [Google Scholar] [CrossRef]
- Darwish, S.M.; Abu Sharkh, S.E.; Abu Teir, M.M.; Makharza, S.; Abu-Hadid, M.M. Spectroscopic investigations of pentobarbital interaction with human serum albumin. J. Mol. Struct. 2010, 963, 122–129. [Google Scholar] [CrossRef]
- Zargar, S.; Wani, T.A. Protective Role of Quercetin in Carbon Tetrachloride Induced Toxicity in Rat Brain: Biochemical, Spectrophotometric Assays and Computational Approach. Molecules 2021, 26, 7526. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Higaki, J.; Gibson, B.; Craik, C. Evolution of Catalysis in the Serine Proteases, Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1987; pp. 615–621. [Google Scholar]
- Wang, Y.; Zhang, H.; Cao, J.; Zhou, Q. Interaction of methotrexate with trypsin analyzed by spectroscopic and molecular modeling methods. J. Mol. Struct. 2013, 1051, 78–85. [Google Scholar] [CrossRef]
- Luo, H.; Li, H.; Liu, Y.; Yang, L.; Xiao, Q.; Huang, S. Investigation on conformational variation and activity of trypsin affected by black phosphorus quantum dots via multi-spectroscopy and molecular modeling. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 256, 119746. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Temp (K) | Ksv (×103 M−1) | kq (×1011 M−1 s−1) | R2 |
|---|---|---|---|---|
| 7.4 | 295 | 4.24 | 4.24 | 0.998 |
| 305 | 3.28 | 3.28 | 0.996 | |
| 315 | 2.49 | 2.89 | 0.990 |
| pH | Temp (K) | Kb (×103 M−1) | n | R2 |
|---|---|---|---|---|
| 7.4 | 295 | 2.192 | 0.93 | 0.995 |
| 305 | 1.974 | 0.95 | 0.980 | |
| 315 | 1.887 | 0.98 | 0.996 |
| Temp (K) | H (KJ mol−1) | S (Jmol−1 k−1) | G (KJ mol−1) | R2 |
|---|---|---|---|---|
| 295 | 5.8 | 44.21 | −7.2 | 0.9954 |
| 305 | −7.6 | 0.9988 | ||
| 315 | −8.1 | 0.9986 |
| TYP/ATR | Vmax (µM s−1) | Km (µM) | Kcat (s−1) | Kcat/Km (µM−1s−1) |
|---|---|---|---|---|
| 1:0 | 5.44 × 10−4 | 3.89 × 10−3 | 3.62 × 10−5 | 9.33 × 10−3 |
| 1:5 | 4.05 × 10−4 | 3.02 × 10−3 | 2.70 × 10−5 | 8.93 × 10−3 |
| 1:10 | 4.14 × 10−4 | 4.71 × 10−3 | 2.76 × 10−5 | 5.84 × 10−3 |
| 1:15 | 3.08 × 10−4 | 3.81 × 10−3 | 2.05 × 10−5 | 5.40 × 10−3 |
| 1:20 | 2.59 × 10−4 | 4.36 × 10−3 | 1.72 × 10−5 | 3.96 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khayyat, A.I.A.; Zargar, S.; Wani, T.A.; Rehman, M.U.; Khan, A.A. Association Mechanism and Conformational Changes in Trypsin on Its Interaction with Atrazine: A Multi- Spectroscopic and Biochemical Study with Computational Approach. Int. J. Mol. Sci. 2022, 23, 5636. https://doi.org/10.3390/ijms23105636
Khayyat AIA, Zargar S, Wani TA, Rehman MU, Khan AA. Association Mechanism and Conformational Changes in Trypsin on Its Interaction with Atrazine: A Multi- Spectroscopic and Biochemical Study with Computational Approach. International Journal of Molecular Sciences. 2022; 23(10):5636. https://doi.org/10.3390/ijms23105636
Chicago/Turabian StyleKhayyat, Arwa Ishaq A., Seema Zargar, Tanveer A. Wani, Muneeb U. Rehman, and Azmat Ali Khan. 2022. "Association Mechanism and Conformational Changes in Trypsin on Its Interaction with Atrazine: A Multi- Spectroscopic and Biochemical Study with Computational Approach" International Journal of Molecular Sciences 23, no. 10: 5636. https://doi.org/10.3390/ijms23105636
APA StyleKhayyat, A. I. A., Zargar, S., Wani, T. A., Rehman, M. U., & Khan, A. A. (2022). Association Mechanism and Conformational Changes in Trypsin on Its Interaction with Atrazine: A Multi- Spectroscopic and Biochemical Study with Computational Approach. International Journal of Molecular Sciences, 23(10), 5636. https://doi.org/10.3390/ijms23105636