
Seizures in PPT1 Knock-In Mice Are Associated with Inflammatory Activation of Microglia

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Seizures in PPT1 KI Mice
2.2. Epileptiform Activity in Brain Slices of PPT1 KI Mice
2.3. Microglial Activation in Hippocampus Correlates with the Occurrence of Seizures
2.4. Astrocyte Is Activated at the Early Stage of PPT1 KI Mice
2.5. Age-Dependent Changes in Synaptic Proteins GluN2B and GABAARα1 in PPT1 KI Mice
2.6. Iba-1 Immunoreactivity and Morphological Changes of Microglia in Hippocampus of PPT1 KI Mice
2.7. Age-Related Neuronal Loss in PPT1 KI Mice

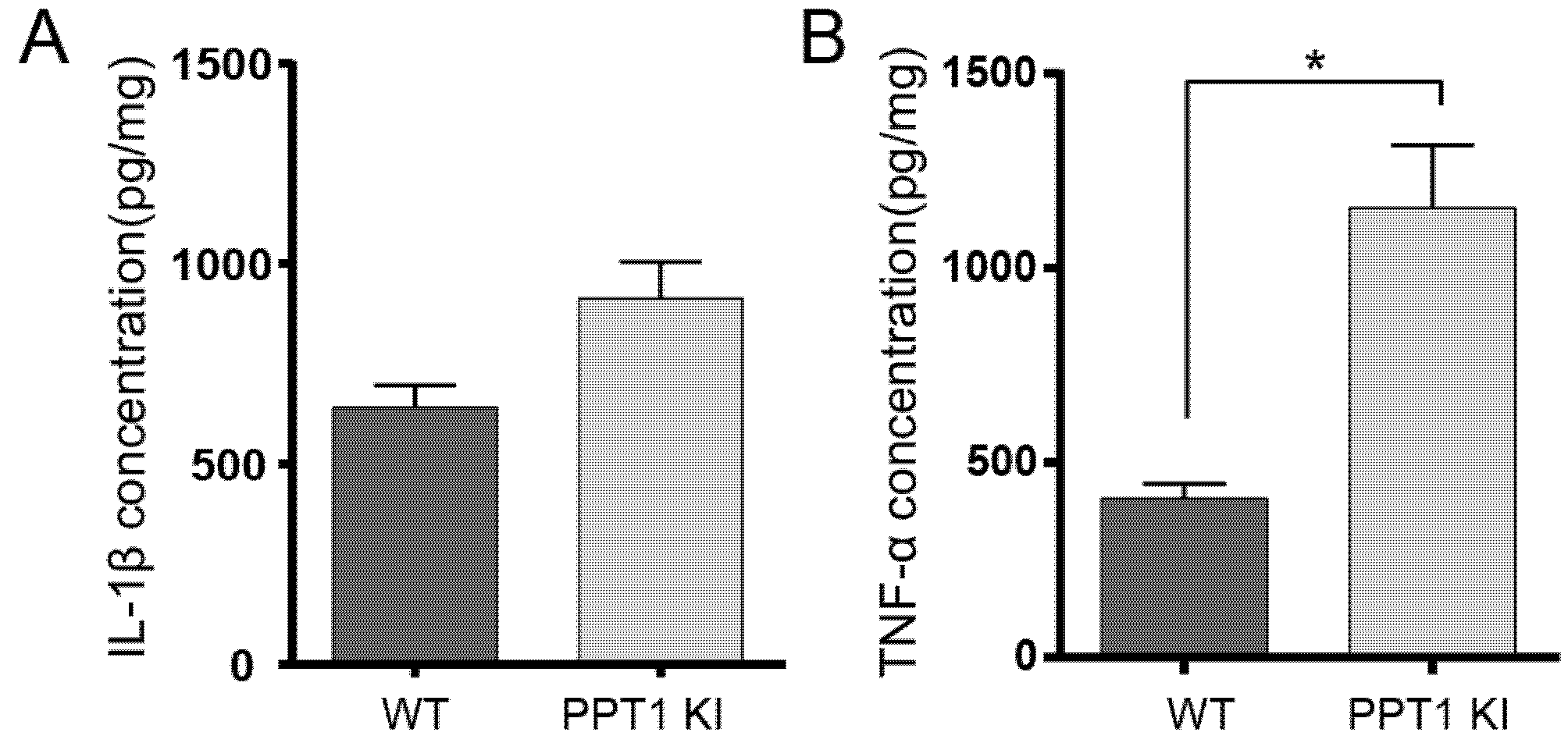
2.8. Cytokines IL-1β and TNF-α Changes in PPT1 KI Mice
2.9. Inhibition of ATP-Sensitive P2X7R Repressed Seizure in PPT1 KI Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Behavioral Scaling in Seizures
4.3. Slice Preparation and Extracellular Field Recordings
4.4. Western Blotting
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. ATP Assays
4.7. Drug Administration
4.8. IHC and Confocal Imaging
4.9. DAB-Staining IHC
4.10. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cooper, J.D. Progress towards understanding the neurobiology of Batten disease or neuronal ceroid lipofuscinosis. Curr. Opin. Neurol. 2003, 16, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, R.M. Clinical features and molecular genetic basis of the neuronal ceroid lipofuscinoses. Adv. Neurol. 2002, 89, 211–215. [Google Scholar] [PubMed]
- Hofmann, S.L.; Atashband, A.; Cho, S.K.; Das, A.K.; Gupta, P.; Lu, J.Y. Neuronal ceroid lipofuscinoses caused by defects in soluble lysosomal enzymes (CLN1 and CLN2). Curr. Mol. Med. 2002, 2, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, H.M.; Mole, S.E. Neurodegenerative disease: The neuronal ceroid lipofuscinoses (Batten disease). Curr. Opin. Neurol. 2001, 14, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, K.E.; Zhong, N.; Philippart, M. Pheno/genotypic correlations of neuronal ceroid lipofuscinoses. Neurology 2001, 57, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Haltia, M.; Rapola, J.; Santavuori, P.; Keranen, A. Infantile type of so-called neuronal ceroid-lipofuscinosis. 2. Morphological and biochemical studies. J. Neurol. Sci. 1973, 18, 269–285. [Google Scholar] [CrossRef]
- Kielar, C.; Maddox, L.; Bible, E.; Pontikis, C.C.; Macauley, S.L.; Griffey, M.A.; Wong, M.; Sands, M.S.; Cooper, J.D. Successive neuron loss in the thalamus and cortex in a mouse model of infantile neuronal ceroid lipofuscinosis. Neurobiol. Dis. 2007, 25, 150–162. [Google Scholar] [CrossRef]
- Fukata, Y.; Fukata, M. Protein palmitoylation in neuronal development and synaptic plasticity. Nat. Rev. Neurosci. 2010, 11, 161–175. [Google Scholar] [CrossRef]
- Groh, J.; Kuhl, T.G.; Ip, C.W.; Nelvagal, H.R.; Sri, S.; Duckett, S.; Mirza, M.; Langmann, T.; Cooper, J.D.; Martini, R. Immune cells perturb axons and impair neuronal survival in a mouse model of infantile neuronal ceroid lipofuscinosis. Brain 2013, 136, 1083–1101. [Google Scholar] [CrossRef][Green Version]
- Bond, M.; Holthaus, S.M.; Tammen, I.; Tear, G.; Russell, C. Use of model organisms for the study of neuronal ceroid lipofuscinosis. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 1842–1865. [Google Scholar] [CrossRef]
- Faller, K.M.; Gutierrez-Quintana, R.; Mohammed, A.; Rahim, A.A.; Tuxworth, R.I.; Wager, K.; Bond, M. The neuronal ceroid lipofuscinoses: Opportunities from model systems. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2015, 1852, 2267–2278. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.N.; Kovacs, A.D.; Pearce, D.A. The novel Cln1(R151X) mouse model of infantile neuronal ceroid lipofuscinosis (INCL) for testing nonsense suppression therapy. Hum. Mol. Genet. 2015, 24, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Bouchelion, A.; Zhang, Z.; Li, Y.; Qian, H.; Mukherjee, A.B. Mice homozygous for c.451C>T mutation in Cln1 gene recapitulate INCL phenotype. Ann. Clin. Transl. Neurol. 2014, 1, 1006–1023. [Google Scholar] [CrossRef] [PubMed]
- Fries, P.; Nikolic, D.; Singer, W. The gamma cycle. Trends Neurosci. 2007, 30, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Hajos, N.; Paulsen, O. Network mechanisms of gamma oscillations in the CA3 region of the hippocampus. Neural Netw. 2009, 22, 1113–1119. [Google Scholar] [CrossRef]
- Lu, C.B.; Jefferys, J.G.; Toescu, E.C.; Vreugdenhil, M. In vitro hippocampal gamma oscillation power as an index of in vivo CA3 gamma oscillation strength and spatial reference memory. Neurobiol. Learn. Mem. 2011, 95, 221–230. [Google Scholar] [CrossRef]
- Pasquetti, M.V.; Meier, L.; Marafiga, J.R.; Barbieri Caus, L.; Tort, A.B.L.; Calcagnotto, M.E. Hippocampal CA1 and cortical interictal oscillations in the pilocarpine model of epilepsy. Brain Res. 2019, 1722, 146351. [Google Scholar] [CrossRef]
- Benedek, K.; Berenyi, A.; Gombkoto, P.; Piilgaard, H.; Lauritzen, M. Neocortical gamma oscillations in idiopathic generalized epilepsy. Epilepsia 2016, 57, 796–804. [Google Scholar] [CrossRef]
- Fisher, R.S.; van Emde Boas, W.; Blume, W.; Elger, C.; Genton, P.; Lee, P.; Engel, J., Jr. Epileptic seizures and epilepsy: Definitions proposed by the International League Against Epilepsy (ILAE) and the International Bureau for Epilepsy (IBE). Epilepsia 2005, 46, 470–472. [Google Scholar] [CrossRef]
- Dalby, N.O.; Mody, I. The process of epileptogenesis: A pathophysiological approach. Curr. Opin. Neurol. 2001, 14, 187–192. [Google Scholar] [CrossRef]
- Sharma, A.K.; Reams, R.Y.; Jordan, W.H.; Miller, M.A.; Thacker, H.L.; Snyder, P.W. Mesial temporal lobe epilepsy: Pathogenesis, induced rodent models and lesions. Toxicol. Pathol. 2007, 35, 984–999. [Google Scholar] [CrossRef] [PubMed]
- Gale, K. Role of GABA in the genesis of chemoconvulsant seizures. Toxicol. Lett. 1992, 64, 417–428. [Google Scholar] [CrossRef]
- Patterson, S.L. Immune dysregulation and cognitive vulnerability in the aging brain: Interactions of microglia, IL-1beta, BDNF and synaptic plasticity. Neuropharmacology 2015, 96, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Facci, L.; Barbierato, M.; Skaper, S.D. Astrocyte/Microglia cocultures as a model to study neuroinflammation. In Neurotrophic Factors; Humana Press: New York, NY, USA, 2018; Volume 1727, pp. 127–137. [Google Scholar]
- Bhaskar, K.; Maphis, N.; Xu, G.; Varvel, N.H.; Kokiko-Cochran, O.N.; Weick, J.P.; Staugaitis, S.M.; Cardona, A.; Ransohoff, R.M.; Herrup, K.; et al. Microglial derived tumor necrosis factor-alpha drives Alzheimer′s disease-related neuronal cell cycle events. Neurobiol. Dis. 2014, 62, 273–285. [Google Scholar] [CrossRef]
- Kia, A.; McAvoy, K.; Krishnamurthy, K.; Trotti, D.; Pasinelli, P. Astrocytes expressing ALS-linked mutant FUS induce motor neuron death through release of tumor necrosis factor-alpha. Glia 2018, 66, 1016–1033. [Google Scholar] [CrossRef]
- Gupta, P.; Soyombo, A.A.; Atashband, A.; Wisniewski, K.E.; Shelton, J.M.; Richardson, J.A.; Hammer, R.E.; Hofmann, S.L. Disruption of PPT1 or PPT2 causes neuronal ceroid lipofuscinosis in knockout mice. Proc. Natl. Acad. Sci. USA 2001, 98, 13566–13571. [Google Scholar] [CrossRef]
- West, P.K.; Viengkhou, B.; Campbell, I.L.; Hofer, M.J. Microglia responses to interleukin-6 and type I interferons in neuroinflammatory disease. Glia 2019, 67, 1821–1841. [Google Scholar] [CrossRef]
- Hansen, D.V.; Hanson, J.E.; Sheng, M. Microglia in Alzheimer′s disease. J. Cell Biol. 2018, 217, 459–472. [Google Scholar] [CrossRef]
- Sellgren, C.M.; Gracias, J.; Watmuff, B.; Biag, J.D.; Thanos, J.M.; Whittredge, P.B.; Fu, T.; Worringer, K.; Brown, H.E.; Wang, J.; et al. Increased synapse elimination by microglia in schizophrenia patient-derived models of synaptic pruning. Nat. Neurosci. 2019, 22, 374–385. [Google Scholar] [CrossRef]
- Illes, P. P2X7 Receptors Amplify CNS Damage in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 5996. [Google Scholar] [CrossRef]
- Szopa, A.; Socala, K.; Serefko, A.; Doboszewska, U.; Wrobel, A.; Poleszak, E.; Wlaz, P. Purinergic transmission in depressive disorders. Pharmacol. Ther. 2021, 224, 107821. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Pacheco, A.; Diaz-Hernandez, M.; Arribas-Blazquez, M.; Sanz-Rodriguez, A.; Olivos-Ore, L.A.; Artalejo, A.R.; Alves, M.; Letavic, M.; Miras-Portugal, M.T.; Conroy, R.M.; et al. Transient P2X7 Receptor Antagonism Produces Lasting Reductions in Spontaneous Seizures and Gliosis in Experimental Temporal Lobe Epilepsy. J. Neurosci. 2016, 36, 5920–5932. [Google Scholar] [CrossRef] [PubMed]
- Beamer, E.; Fischer, W.; Engel, T. The ATP-Gated P2X7 Receptor As a Target for the Treatment of Drug-Resistant Epilepsy. Front. Neurosci. 2017, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Illes, P.; Verkhratsky, A.; Tang, Y. Surveilling microglia dampens neuronal activity: Operation of a purinergically mediated negative feedback mechanism. Signal Transduct. Target. Ther. 2021, 6, 160. [Google Scholar] [CrossRef] [PubMed]
- Monif, M.; Reid, C.A.; Powell, K.L.; Smart, M.L.; Williams, D.A. The P2X7 receptor drives microglial activation and proliferation: A trophic role for P2X7R pore. J. Neurosci. 2009, 29, 3781–3791. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Wang, Q.; Ao, H.; Shoblock, J.R.; Lord, B.; Aluisio, L.; Fraser, I.; Nepomuceno, D.; Neff, R.A.; Welty, N.; et al. Pharmacological characterization of a novel centrally permeable P2X7 receptor antagonist: JNJ-47965567. Br. J. Pharmacol. 2013, 170, 624–640. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Dal Ben, D.; Sarti, A.C.; Giuliani, A.L.; Falzoni, S. The P2X7 Receptor in Infection and Inflammation. Immunity 2017, 47, 15–31. [Google Scholar] [CrossRef]
- Song, P.; Hu, J.; Liu, X.; Deng, X. Increased expression of the P2X7 receptor in temporal lobe epilepsy: Animal models and clinical evidence. Mol. Med. Rep. 2019, 19, 5433–5439. [Google Scholar] [CrossRef]
- Liu, Z.; Ren, F.; Yang, B.; Zhang, H.; Guo, F.; Zhang, X.; Lu, C.; Zhang, R. Region-dependent regulation of acute ethanol on gamma oscillation in the rat hippocampal slices. Psychopharmacology 2020, 237, 2959–2966. [Google Scholar] [CrossRef]
- Ghit, A.; Assal, D.; Al-Shami, A.S.; Hussein, D.E.E. GABAA receptors: Structure, function, pharmacology, and related disorders. J. Genet. Eng. Biotechnol. 2021, 19, 123. [Google Scholar] [CrossRef]
- Koster, K.P.; Francesconi, W.; Berton, F.; Alahmadi, S.; Srinivas, R.; Yoshii, A. Developmental NMDA receptor dysregulation in the infantile neuronal ceroid lipofuscinosis mouse model. Elife 2019, 8, e40316. [Google Scholar] [CrossRef] [PubMed]
- Osorio, I.; Frei, M.G.; Wilkinson, S.B. Real-time automated detection and quantitative analysis of seizures and short-term prediction of clinical onset. Epilepsia 1998, 39, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Setkowicz, Z.; Kosonowska, E.; Janeczko, K. Inflammation in the developing rat modulates astroglial reactivity to seizures in the mature brain. J. Anat. 2017, 231, 366–379. [Google Scholar] [CrossRef] [PubMed]
- Lange, J.; Haslett, L.J.; Lloyd-Evans, E.; Pocock, J.M.; Sands, M.S.; Williams, B.P.; Cooper, J.D. Compromised astrocyte function and survival negatively impact neurons in infantile neuronal ceroid lipofuscinosis. Acta Neuropathol. Commun. 2018, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Lu, L.; Rao, M.; Huang, Y.; Liu, C.E.; Liu, S.; Zhao, Y.; Liu, H.; Zhu, J.; Chao, T.; et al. GFAP hyperpalmitoylation exacerbates astrogliosis and neurodegenerative pathology in PPT1-deficient mice. Proc. Natl. Acad. Sci. USA 2021, 118, e2022261118. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, C.; Chandra, G.; Peng, S.; Zhang, Z.; Liu, A.; Mukherjee, A.B. Neuroprotection and lifespan extension in Ppt1(-/-) mice by NtBuHA. therapeutic implications for INCL. Nat. Neurosci. 2013, 16, 1608–1617. [Google Scholar] [CrossRef]
- Finn, R.; Kovacs, A.D.; Pearce, D.A. Altered glutamate receptor function in the cerebellum of the Ppt1-/- mouse, a murine model of infantile neuronal ceroid lipofuscinosis. J. Neurosci. Res. 2012, 90, 367–375. [Google Scholar] [CrossRef]
- Boer, K.; Spliet, W.G.; van Rijen, P.C.; Redeker, S.; Troost, D.; Aronica, E. Evidence of activated microglia in focal cortical dysplasia. J. Neuroimmunol. 2006, 173, 188–195. [Google Scholar] [CrossRef]
- Avignone, E.; Lepleux, M.; Angibaud, J.; Nagerl, U.V. Altered morphological dynamics of activated microglia after induction of status epilepticus. J. Neuroinflammation 2015, 12, 202. [Google Scholar] [CrossRef]
- Bechade, C.; Cantaut-Belarif, Y.; Bessis, A. Microglial control of neuronal activity. Front. Cell. Neurosci. 2013, 7, 32. [Google Scholar] [CrossRef]
- Temkin, N.R. Preventing and treating posttraumatic seizures: The human experience. Epilepsia 2009, 50, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Gaitatzis, A.; Johnson, A.L.; Chadwick, D.W.; Shorvon, S.D.; Sander, J.W. Life expectancy in people with newly diagnosed epilepsy. Brain 2004, 127, 2427–2432. [Google Scholar] [CrossRef] [PubMed]
- Pascual, O.; Ben Achour, S.; Rostaing, P.; Triller, A.; Bessis, A. Microglia activation triggers astrocyte-mediated modulation of excitatory neurotransmission. Proc. Natl. Acad. Sci. USA 2012, 109, E197–E205. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, A.; Keno, Y.; Marumiya, A.; Sudo, Y.; Uda, Y.; Matsuda, K.; Morita, Y.; Furuta, T.; Nishida, K.; Nagasawa, K. Expression level of P2X7 receptor is a determinant of ATP-induced death of mouse cultured neurons. Neuroscience 2016, 319, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Berve, K.; West, B.L.; Martini, R.; Groh, J. Sex- and region-biased depletion of microglia/macrophages attenuates CLN1 disease in mice. J. Neuroinflammation 2020, 17, 323. [Google Scholar] [CrossRef] [PubMed]
- Macauley, S.L.; Wong, A.M.; Shyng, C.; Augner, D.P.; Dearborn, J.T.; Pearse, Y.; Roberts, M.S.; Fowler, S.C.; Cooper, J.D.; Watterson, D.M.; et al. An anti-neuroinflammatory that targets dysregulated glia enhances the efficacy of CNS-directed gene therapy in murine infantile neuronal ceroid lipofuscinosis. J. Neurosci. 2014, 34, 13077–13082. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Arcuino, G.; Takano, T.; Lin, J.; Peng, W.G.; Wan, P.; Li, P.; Xu, Q.; Liu, Q.S.; Goldman, S.A.; et al. P2X7 receptor inhibition improves recovery after spinal cord injury. Nat. Med. 2004, 10, 821–827. [Google Scholar] [CrossRef]
- Abbracchio, M.P.; Burnstock, G.; Verkhratsky, A.; Zimmermann, H. Purinergic signalling in the nervous system: An overview. Trends Neurosci. 2009, 32, 19–29. [Google Scholar] [CrossRef]
- Engel, T.; Gomez-Villafuertes, R.; Tanaka, K.; Mesuret, G.; Sanz-Rodriguez, A.; Garcia-Huerta, P.; Miras-Portugal, M.T.; Henshall, D.C.; Diaz-Hernandez, M. Seizure suppression and neuroprotection by targeting the purinergic P2X7 receptor during status epilepticus in mice. FASEB J. 2012, 26, 1616–1628. [Google Scholar] [CrossRef]
- Mesuret, G.; Engel, T.; Hessel, E.V.; Sanz-Rodriguez, A.; Jimenez-Pacheco, A.; Miras-Portugal, M.T.; Diaz-Hernandez, M.; Henshall, D.C. P2X7 receptor inhibition interrupts the progression of seizures in immature rats and reduces hippocampal damage. CNS Neurosci. Ther. 2014, 20, 556–564. [Google Scholar] [CrossRef]
- Racine, R.J. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wang, M.; Feng, B.; Zhang, Q.; Tong, J.; Wang, M.; Lu, C.; Peng, S. Seizures in PPT1 Knock-In Mice Are Associated with Inflammatory Activation of Microglia. Int. J. Mol. Sci. 2022, 23, 5586. https://doi.org/10.3390/ijms23105586
Zhang X, Wang M, Feng B, Zhang Q, Tong J, Wang M, Lu C, Peng S. Seizures in PPT1 Knock-In Mice Are Associated with Inflammatory Activation of Microglia. International Journal of Molecular Sciences. 2022; 23(10):5586. https://doi.org/10.3390/ijms23105586
Chicago/Turabian StyleZhang, Xusheng, Mengting Wang, Bingyan Feng, Qiuyu Zhang, Jia Tong, Mingyong Wang, Chengbiao Lu, and Shiyong Peng. 2022. "Seizures in PPT1 Knock-In Mice Are Associated with Inflammatory Activation of Microglia" International Journal of Molecular Sciences 23, no. 10: 5586. https://doi.org/10.3390/ijms23105586
APA StyleZhang, X., Wang, M., Feng, B., Zhang, Q., Tong, J., Wang, M., Lu, C., & Peng, S. (2022). Seizures in PPT1 Knock-In Mice Are Associated with Inflammatory Activation of Microglia. International Journal of Molecular Sciences, 23(10), 5586. https://doi.org/10.3390/ijms23105586