
Basic Science with Preclinical Models to Investigate and Develop Liquid Biopsy: What Are the Available Data and Is It a Fruitful Approach?


Abstract
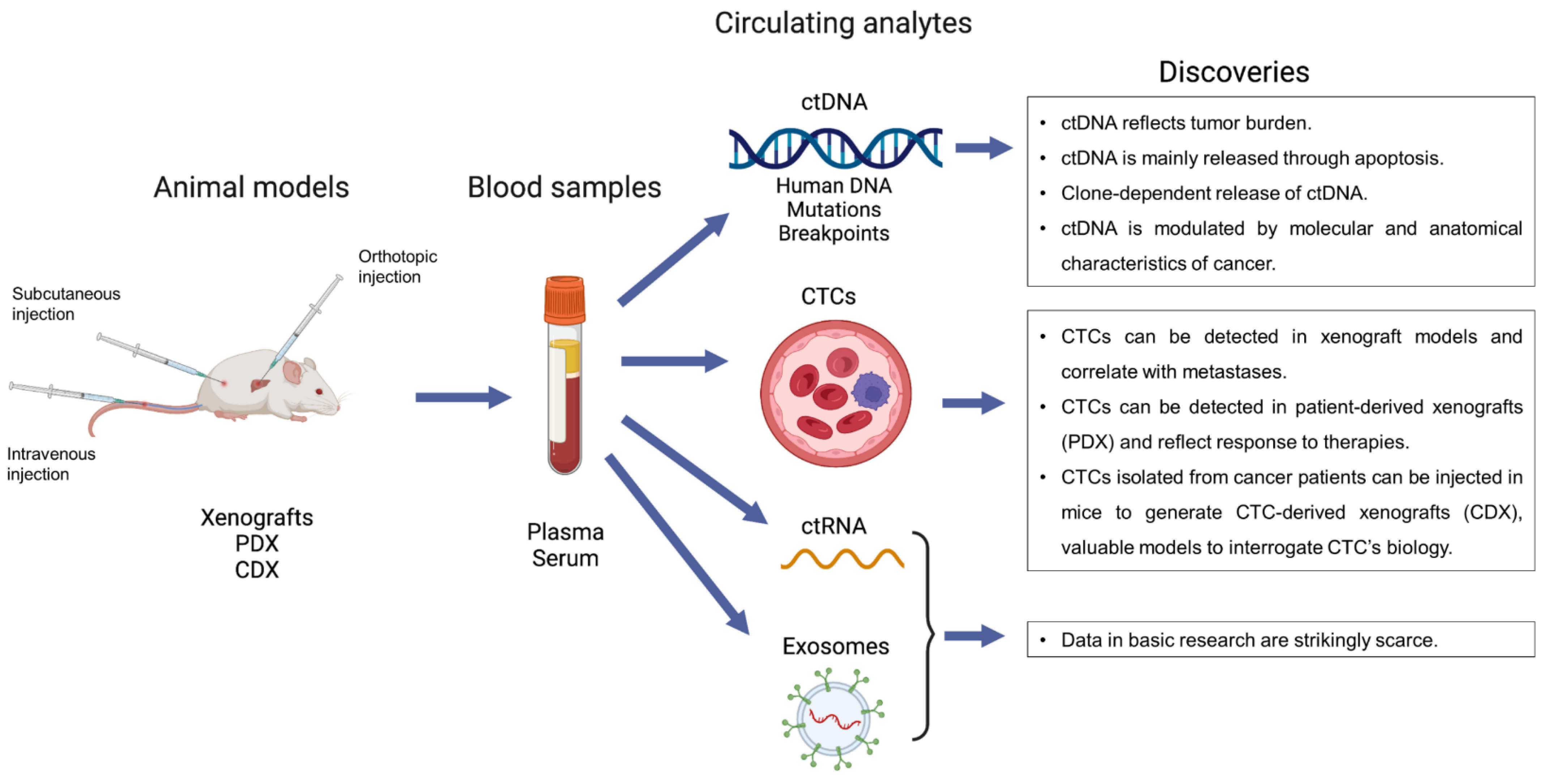
:1. Introduction
2. Circulating Tumor DNA (ctDNA)
2.1. ctDNA in Basic Research at a Glance
2.2. ctDNA Recapitulates Tumor Burden
2.3. ctDNA Is Mainly Released through Apoptosis
2.4. ctDNA Release Is Clone-Dependent
2.5. Exploring the Determinants of ctDNA Release
3. Circulating Tumor Cells (CTCs)
3.1. CTCs in Basic Research at a Glance
3.2. CTCs in Xenografts
3.3. CTCs in PDX
3.4. CTCs in CDX
4. ctRNA and Exosomes
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CTCs | Circulating tumor cells |
| ctDNA | Circulating tumor DNA |
| ctRNA | Circulating tumor RNA |
| CDX | CTCs-derived xenograft |
| PDX | Patient-derived xenograft |
References
- Norton, M.E.; Jacobsson, B.; Swamy, G.K.; Laurent, L.C.; Ranzini, A.C.; Brar, H.; Tomlinson, M.W.; Pereira, L.; Spitz, J.L.; Hollemon, D.; et al. Cell-free DNA Analysis for Noninvasive Examination of Trisomy. N. Engl. J. Med. 2015, 372, 1589–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, D.W.; Chudova, D.; Sehnert, A.J.; Bhatt, S.; Murray, K.; Prosen, T.L.; Garber, J.E.; Wilkins-Haug, L.; Vora, N.; Warsof, S.; et al. Noninvasive Prenatal Testing and Incidental Detection of Occult Maternal Malignancies. JAMA 2015, 314, 162–169. [Google Scholar] [CrossRef] [Green Version]
- MIT Technology Review. 2015. Available online: https://www.technologyreview.com/magazines/10-breakthrough-technologies-2015 (accessed on 30 March 2022).
- Von Felden, J.; Garcia-Lezana, T.; Dogra, N.; Gonzalez-Kozlova, E.; Ahsen, M.E.; Craig, A.; Gifford, S.; Wunsch, B.; Smith, J.T.; Kim, S.; et al. Unannotated small RNA clusters associated with circulating extracellular vesicles detect early stage liver cancer. Gut 2021, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.D.; Li, L.; Wang, Y.; Thoburn, C.; Afsari, B.; Danilova, L.; Douville, C.; Javed, A.A.; Wong, F.; Mattox, A.; et al. Detection and localization of surgically resectable cancers with a multi-analyte blood test. Science 2018, 359, 926–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettegowda, C.; Sausen, M.; Leary, R.J.; Kinde, I.; Wang, Y.; Agrawal, N.; Bartlett, B.R.; Wang, H.; Luber, B.; Alani, R.M.; et al. Detection of circulating tumor DNA in early- and late-stage human malignancies. Sci. Transl. Med. 2014, 6, 224. [Google Scholar] [CrossRef] [Green Version]
- Tie, J.; Wang, Y.; Tomasetti, C.; Li, L.; Springer, S.; Kinde, I.; Silliman, N.; Tacey, M.; Wong, H.-L.; Christie, M.; et al. Circulating tumor DNA analysis detects minimal residual disease and predicts recurrence in patients with stage II colon cancer. Sci. Transl. Med. 2016, 8, 346ra92. [Google Scholar] [CrossRef] [Green Version]
- Tie, J.; Cohen, J.D.; Wang, Y.; Christie, M.; Simons, K.; Lee, M.; Wong, R.; Kosmider, S.; Ananda, S.; McKendrick, J.; et al. Circulating Tumor DNA Analyses as Markers of Recurrence Risk and Benefit of Adjuvant Therapy for Stage III Colon Cancer. JAMA Oncol. 2019, 5, 1710–1717. [Google Scholar] [CrossRef]
- Lee, B.; Lipton, L.; Cohen, J.; Tie, J.; Javed, A.; Li, L.; Goldstein, D.; Burge, M.; Cooray, P.; Nagrial, A.; et al. Circulating tumor DNA as a potential marker of adjuvant chemotherapy benefit following surgery for localized pancreatic cancer. Ann. Oncol. 2019, 30, 1472–1478. [Google Scholar] [CrossRef]
- Von Felden, J.; Craig, A.J.; Garcia-Lezana, T.; Labgaa, I.; Haber, P.K.; D’Avola, D.; Asgharpour, A.; Dieterich, D.; Bonaccorso, A.; Torres-Martin, M.; et al. Mutations in circulating tumor DNA predict primary resistance to systemic therapies in advanced hepa-tocellular carcinoma. Oncogene 2021, 40, 140–151. [Google Scholar] [CrossRef]
- Labgaa, I.; Villanueva, A. Liquid biopsy in liver cancer. Discov. Med. 2015, 19, 263–273. [Google Scholar]
- Labgaa, I.; Villanueva, A.; Dormond, O.; Demartines, N.; Melloul, E. The Role of Liquid Biopsy in Hepatocellular Carcinoma Prognostication. Cancers 2021, 13, 659. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Siravegna, G.; Blaszkowsky, L.S.; Corti, G.; Crisafulli, G.; Ahronian, L.G.; Mussolin, B.; Kwak, E.L.; Buscarino, M.; Lazzari, L.; et al. Tumor Heterogeneity and Lesion-Specific Response to Targeted Therapy in Colorectal Cancer. Cancer Discov. 2015, 6, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, M.; Misale, S.; Wei, G.; Siravegna, G.; Crisafulli, G.; Lazzari, L.; Corti, G.; Rospo, G.; Novara, L.; Mussolin, B.; et al. Acquired Resistance to the TRK Inhibitor Entrectinib in Colorectal Cancer. Cancer Discov. 2015, 6, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siravegna, G.; Mussolin, B.; Buscarino, M.; Corti, G.; Cassingena, A.; Crisafulli, G.; Ponzetti, A.; Cremolini, C.; Amatu, A.; Lauricella, C.; et al. Clonal evolution and resistance to EGFR blockade in the blood of colorectal cancer patients. Nat. Med. 2015, 21, 795–801, Erratium in Nat. Med. 2015, 21, 827. [Google Scholar]
- Parikh, A.R.; Leshchiner, I.; Elagina, L.; Goyal, L.; Levovitz, C.; Siravegna, G.; Livitz, D.; Rhrissorrakrai, K.; Martin, E.E.; Van Seventer, E.E.; et al. Liquid versus tissue biopsy for detecting acquired resistance and tumor heterogeneity in gastrointestinal cancers. Nat. Med. 2019, 25, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Sidaway, P. From the EACR–ESMO liquid biopsy meeting. Nat. Rev. Clin. Oncol. 2019, 16, 466. [Google Scholar] [CrossRef]
- Jiang, P.; Chan, C.W.M.; Chan, K.C.A.; Cheng, S.H.; Wong, J.; Wong, V.W.-S.; Wong, G.L.H.; Chan, S.L.; Mok, T.S.K.; Chan, H.L.Y.; et al. Lengthening and shortening of plasma DNA in hepatocellular carcinoma patients. Proc. Natl. Acad. Sci. USA 2015, 112, E1317–E1325. [Google Scholar] [CrossRef] [Green Version]
- Deveson, I.W.; Gong, B.; Lai, K.; LoCoco, J.S.; Richmond, T.A.; Schageman, J.; Zhang, Z.; Novoradovskaya, N.; Willey, J.C.; Jones, W.; et al. Evaluating the analytical validity of circulating tumor DNA sequencing assays for precision oncology. Nat. Biotechnol. 2021, 39, 1115–1128. [Google Scholar] [CrossRef]
- Labgaa, I.; von Felden, J.; Craig, A.J.; Martins-Filho, S.N.; Villacorta-Martin, C.; Demartines, N.; Dormond, O.; D’Avola, D.; Villanueva, A. Experimental Models of Liquid Biopsy in Hepatocellular Carcinoma Reveal Clone-Dependent Release of Circulating Tumor DNA. Hepatol. Commun. 2021, 5, 1095–1105. [Google Scholar] [CrossRef]
- Grabuschnig, S.; Bronkhorst, A.J.; Holdenrieder, S.; Rodriguez, I.R.; Schliep, K.P.; Schwendenwein, D.; Ungerer, V.; Sensen, C.W. Putative Origins of Cell-Free DNA in Humans: A Review of Active and Passive Nucleic Acid Release Mechanisms. Int. J. Mol. Sci. 2020, 21, 8062. [Google Scholar] [CrossRef]
- Labgaa, I.; Villacorta-Martin, C.; D’Avola, D.; Craig, A.J.; Von Felden, J.; Filho, S.M.; Sia, D.; Stueck, A.; Ward, S.C.; Fiel, M.I.; et al. A pilot study of ultra-deep targeted sequencing of plasma DNA identifies driver mutations in hepatocellular carcinoma. Oncogene 2018, 37, 3740–3752. [Google Scholar] [CrossRef] [PubMed]
- Kamat, A.A.; Bischoff, F.Z.; Dang, D.; Baldwin, M.; Han, L.Y.; Lin, Y.G.; Merritt, W.M.; Landen, C.N.; Lu, C.; Gershenson, D.M.; et al. Circulating cell-free DNA: A novel biomarker for response to therapy in ovarian carcinoma. Cancer Biol. Ther. 2006, 5, 1369–1374. [Google Scholar] [CrossRef] [Green Version]
- Rago, C.; Huso, D.L.; Diehl, F.; Karim, B.; Liu, G.; Papadopoulos, N.; Samuels, Y.; Velculescu, V.; Vogelstein, B.; Kinzler, K.W.; et al. Serial Assessment of Human Tumor Burdens in Mice by the Analysis of Circulating DNA. Cancer Res. 2007, 67, 9364–9370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M.; Chu, D.; Meyer, C.F.; Llosa, N.J.; Bs, G.M.; Morris, C.D.; Levin, A.; Wolinsky, J.-P.; Albert, C.M.; Steppan, D.A.; et al. Highly personalized detection of minimal Ewing sarcoma disease burden from plasma tumor DNA. Cancer 2016, 122, 3015–3023. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Nolley, R.; Chan, A.M.W.; Rankin, E.B.; Peehl, N.M. Cabozantinib inhibits tumor growth and metastasis of a patient-derived xenograft model of papillary renal cell carcinoma with MET mutation. Cancer Biol. Ther. 2016, 18, 863–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouliere, F.; Robert, B.; Peyrotte, E.A.; Del Rio, M.; Ychou, M.; Molina, F.; Gongora, C.; Thierry, A.R. High Fragmentation Characterizes Tumour-Derived Circulating DNA. PLoS ONE 2011, 6, e23418. [Google Scholar] [CrossRef] [PubMed]
- Mouliere, F.; El Messaoudi, S.; Gongora, C.; Guedj, A.-S.; Robert, B.; Del Rio, M.; Molina, F.; Lamy, P.-J.; Lopez-Crapez, E.; Mathonnet, M.; et al. Circulating Cell-Free DNA from Colorectal Cancer Patients May Reveal High KRAS or BRAF Mutation Load. Transl. Oncol. 2013, 6, 319-IN8. [Google Scholar] [CrossRef] [Green Version]
- Marques, J.F.; Neto, S.; Pinheiro, J.; Machado, J.C.; Costa, J.L. Induction of apoptosis increases sensitivity to detect cancer mutations in plasma. Eur. J. Cancer 2020, 127, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thierry, A.R.; Mouliere, F.; Gongora, C.; Ollier, J.; Robert, B.; Ychou, M.; Del Rio, M.; Molina, F. Origin and quantification of circulating DNA in mice with human colorectal cancer xenografts. Nucleic Acids Res. 2010, 38, 6159–6175. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Omura-Minamisawa, M.; Kang, Y.; Hara, T.; Koike, I.; Inoue, T. Quantification of circulating cell-free DNA in the plasma of cancer patients during radiation therapy. Cancer Sci. 2009, 100, 303–309. [Google Scholar] [CrossRef]
- Gorges, T.M.; Schiller, J.; Schmitz, A.; Schuetzmann, D.; Schatz, C.; Zollner, T.M.; Krahn, T.; Von Ahsen, O. Cancer therapy monitoring in xenografts by quantitative analysis of circulating tumor DNA. Biomarkers 2012, 17, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Gasparello, J.; Allegretti, M.; Tremante, E.; Fabbri, E.; Amoreo, C.A.; Romania, P.; Melucci, E.; Messana, K.; Borgatti, M.; Giacomini, P.; et al. Liquid biopsy in mice bearing colorectal carcinoma xenografts: Gateways regulating the levels of circulating tumor DNA (ctDNA) and miRNA (ctmiRNA). J. Exp. Clin. Cancer Res. 2018, 37, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.; Xie, L.; Wang, X.; Ma, H.; Lv, L.; Liu, L.; Song, X. Circulating tumor DNA measurement provides reliable mutation detection in mice with human lung cancer xenografts. Lab. Investig. 2018, 98, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-F.; Wang, X.-J.; Lu, Z.; Liu, S.-R.; Jiang, Y.; Wan, X.-Q.; Cheng, C.-C.; Shi, L.-H.; Wang, L.-H.; Ding, Y. Overexpression of Stat3 increases circulating cfDNA in breast cancer. Breast Cancer Res. Treat. 2021, 187, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Rostami, A.; Lambie, M.; Yu, C.W.; Stambolic, V.; Waldron, J.N.; Bratman, S.V. Senescence, Necrosis, and Apoptosis Govern Circulating Cell-free DNA Release Kinetics. Cell Rep. 2020, 31, 107830. [Google Scholar] [CrossRef]
- Mishra, A.; Zennami, K.; Velarde, E.; Thorek, D.L.J.; Yegnasubramanian, S.; DeWeese, T.L.; Lupold, S.E. Longitudinal measurement of subcutaneous and intratibial human prostate cancer xenograft growth and response to ionizing radiation by plasma Alu and LINE-1 ctDNA: A comparison to standard methods. Prostate 2021, 81, 745–753. [Google Scholar] [CrossRef]
- Noh, J.M.; Kim, Y.J.; Lee, H.Y.; Choi, C.; Ahn, W.-G.; Lee, T.; Pyo, H.; Park, J.H.; Park, D.; Park, W.-Y. Targeted Liquid Biopsy Using Irradiation to Facilitate the Release of Cell-Free DNA from a Spatially Aimed Tumor Tissue. Cancer Res. Treat. 2022, 54, 40–53. [Google Scholar] [CrossRef]
- Losic, B.; Craig, A.J.; Villacorta-Martin, C.; Martins-Filho, S.N.; Akers, N.; Chen, X.; Ahsen, M.E.; Von Felden, J.; Labgaa, I.; D’Avola, D.; et al. Intratumoral heterogeneity and clonal evolution in liver cancer. Nat. Commun. 2020, 11, 291. [Google Scholar] [CrossRef]
- D’Avola, D.; Villacorta-Martin, C.; Martins-Filho, S.N.; Craig, A.; Labgaa, I.; von Felden, J.; Kimaada, A.; Bonaccorso, A.; Tabrizian, P.; Hartmann, B.M.; et al. High-density single cell mRNA sequencing to characterize circulating tumor cells in hepatocellular carci-noma. Sci. Rep. 2018, 8, 11570. [Google Scholar] [CrossRef]
- Sun, Y.F.; Guo, W.; Xu, Y.; Shi, Y.H.; Gong, Z.J.; Ji, Y.; Du, M.; Zhang, X.; Hu, B.; Huang, A.; et al. Circulating Tumor Cells from Different Vascular Sites Exhibit Spatial Heterogeneity in Epithelial and Mesen-chymal Composition and Distinct Clinical Significance in Hepatocellular Carcinoma. Clin. Cancer Res. 2018, 24, 547–559. [Google Scholar] [CrossRef] [Green Version]
- Eliane, J.-P.; Repollet, M.; Luker, K.E.; Brown, M.; Rae, J.M.; Dontu, G.; Schott, A.F.; Wicha, M.; Doyle, G.V.; Hayes, D.F.; et al. Monitoring Serial Changes in Circulating Human Breast Cancer Cells in Murine Xenograft Models. Cancer Res. 2008, 68, 5529–5532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torphy, R.J.; Tignanelli, C.J.; Kamande, J.W.; Moffitt, R.A.; Loeza, S.G.H.; Soper, S.A.; Yeh, J.J. Circulating Tumor Cells as a Biomarker of Response to Treatment in Patient-Derived Xenograft Mouse Models of Pancreatic Adenocarcinoma. PLoS ONE 2014, 9, e89474. [Google Scholar] [CrossRef]
- Pei, H.; Li, L.; Wang, Y.; Sheng, R.; Wang, Y.; Xie, S.; Shui, L.; Si, H.; Tang, B. Single-Cell Phenotypic Profiling of CTCs in Whole Blood Using an Integrated Microfluidic Device. Anal. Chem. 2019, 91, 11078–11084. [Google Scholar] [CrossRef]
- Ramirez, A.B.; Bhat, R.; Sahay, D.; De Angelis, C.; Thangavel, H.; Hedayatpour, S.; Dobrolecki, L.E.; Nardone, A.; Giuliano, M.; Nagi, C.; et al. Circulating tumor cell investigation in breast cancer patient-derived xenograft models by automated immunofluorescence staining, image acquisition, and single cell retrieval and analysis. BMC Cancer 2019, 19, 220. [Google Scholar] [CrossRef]
- Giuliano, M.; Herrera, S.; Christiny, P.; Shaw, C.; Creighton, C.J.; Mitchell, T.; Bhat, R.; Zhang, X.; Mao, S.; Dobrolecki, L.E.; et al. Circulating and disseminated tumor cells from breast cancer patient-derived xenograft-bearing mice as a novel model to study metastasis. Breast Cancer Res. 2015, 17, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baccelli, I.; Schneeweiss, A.; Riethdorf, S.; Stenzinger, A.; Schillert, A.; Vogel, V.; Klein, C.; Saini, M.; Bäuerle, T.; Wallwiener, M.; et al. Identification of a population of blood circulating tumor cells from breast cancer patients that initiates me-tastasis in a xenograft assay. Nat. Biotechnol. 2013, 31, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Bardia, A.; Aceto, N.; Bersani, F.; Madden, M.W.; Donaldson, M.C.; Desai, R.; Zhu, H.; Comaills, V.; Zheng, Z.; et al. Ex vivo culture of circulating breast tumor cells for individualized testing of drug susceptibility. Science 2014, 345, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Vishnoi, M.; Liu, N.H.; Yin, W.; Boral, D.; Scamardo, A.; Hong, D.; Marchetti, D. The identification of a TNBC liver metastasis gene signature by sequential CTC-xenograft modeling. Mol. Oncol. 2019, 13, 1913–1926. [Google Scholar] [CrossRef] [Green Version]
- Pereira-Veiga, T.; Abreu, M.; Robledo, D.; Matias-Guiu, X.; Santacana, M.; Sánchez, L.; Cueva, J.; Palacios, P.; Abdulkader, I.; López-López, R.; et al. CTCs-derived xenograft development in a triple negative breast cancer case. Int. J. Cancer 2018, 144, 2254–2265. [Google Scholar] [CrossRef] [Green Version]
- Hodgkinson, C.L.; Morrow, C.J.; Li, Y.; Metcalf, R.L.; Rothwell, D.G.; Trapani, F.; Polanski, R.; Burt, D.J.; Simpson, K.L.; Morris, K.; et al. Tumorigenicity and genetic profiling of circulating tumor cells in small-cell lung cancer. Nat. Med. 2014, 20, 897–903. [Google Scholar] [CrossRef]
- Drapkin, B.J.; George, J.; Christensen, C.L.; Mino-Kenudson, M.; Dries, R.; Sundaresan, T.; Phat, S.; Myers, D.T.; Zhong, J.; Igo, P.; et al. Genomic and Functional Fidelity of Small Cell Lung Cancer Patient-Derived Xenografts. Cancer Discov. 2018, 8, 600–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lallo, A.; Frese, K.K.; Morrow, C.J.; Sloane, R.; Gulati, S.; Schenk, M.W.; Trapani, F.; Simms, N.; Galvin, M.; Brown, S.; et al. The Combination of the PARP Inhibitor Olaparib and the WEE1 Inhibitor AZD1775 as a New Therapeutic Option for Small Cell Lung Cancer. Clin. Cancer Res. 2018, 24, 5153–5164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, C.J.; Trapani, F.; Metcalf, R.L.; Bertolini, G.; Hodgkinson, C.L.; Khandelwal, G.; Kelly, P.; Galvin, M.; Carter, L.; Simpson, K.L.; et al. Tumourigenic non-small-cell lung cancer mesenchymal circulating tumour cells: A clinical case study. Ann. Oncol. 2016, 27, 1155–1160. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.; Rugge, M.; Facchinetti, A.; Pizzi, M.; Nardo, G.; Barbieri, V.; Manicone, M.; De Faveri, S.; Scaini, M.C.; Basso, U.; et al. Retaining the long-survive capacity of Circulating Tumor Cells (CTCs) followed by xeno-transplantation: Not only from metastatic cancer of the breast but also of prostate cancer patients. Oncoscience 2013, 1, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Girotti, M.R.; Gremel, G.; Lee, R.; Galvani, E.; Rothwell, D.; Viros, A.; Mandal, A.K.; Lim, K.H.J.; Saturno, G.; Furney, S.J.; et al. Application of Sequencing, Liquid Biopsies, and Patient-Derived Xenografts for Personalized Medicine in Melanoma. Cancer Discov. 2015, 6, 286–299. [Google Scholar] [CrossRef] [Green Version]
- Sia, D.; Moeini, A.; Labgaa, I.; Villanueva, A. The future of patient-derived tumor xenografts in cancer treatment. Pharmacogenomics 2015, 16, 1671–1683. [Google Scholar] [CrossRef]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef]

{kind=link}
| Cancer Type(s) (Cell Line(s)) | Animal Model(s) | Technique(s) of Detection | Readout(s) | Threshold (Volume of Blood) | Main Finding(s) | Reference |
|---|---|---|---|---|---|---|
| Human ovarian carcinoma (HeyA8) | Female athymic nude mice (orthotopic xenograft model) | qRT-PCR | Human specific β-actin | 200 µL of plasma | cfDNA levels correlated with tumor burden. After chemotherapy, cfDNA showed a transient increase followed by a drop. The initial increase in cfDNA after treatment correlated with apoptosis. | [23] |
| Human colon cancer | Female athymic nude mice (subcutaneous xenograft model) | qRT-PCR | hLINE-1 | 100 µL of plasma | High-sensitive assay to quantify circulating human DNA (proxy of ctDNA) in small volumes of mouse plasma. CtDNA reliably reflected tumor progression, showing transient spike after cytotoxic or surgical treatments and a decrease after successful interventions. | [24] |
| Human colorectal adenocarcinoma (CL-188) | Female athymic nude mice (intravenous xenograft model) | |||||
| Human osteosarcoma (143B) | Female athymic nude mice (subcutaneous xenograft model) | |||||
| Human Ewing Sarcoma (TC71, EWS1, EWS4) | NSG mice implanted in the pretibial space with either TC71 xenografts or EWS1/4 PDX | ddPCR | EWS-FLI1 tumor specific breakpoint | 50 µL of whole blood | A sensitive assay was developed to detect breakpoint DNA fragments in xenograft and PDX models of Ewing sarcoma. CtDNA was detected and its concentration correlated with tumor burden, showing a significant decrease after surgical resection. | [25] |
| Human papillary renal cell carcinoma (pRCC) with MET mutation | Patient-derived xenografts (PDX) (orthotopic and subcutaneous engraftment) | qRT-PCR | MET | 50 µL of plasma | A PDX model was established with pRCC tissue carrying a MET mutation. Mutant ctDNA was detected in plasma. Placebo group showed higher level of ctDNA compared to the group treated with Cabozantinib. Mice treated with cabozantinib showed decreasing amount of ctDNA upon treatment. | [26] |
| Human colon cancer (SW620, HT29) | Female athymic nude mice (subcutaneous xenograft model) | qRT-PCR | KRAS and ACTB | 200 µL of plasma | CtDNA fragmentation increased with tumor size and tumor ctDNA concentration. CtDNA fragments showed a typical length of 60–100 bp. | [27] |
| Human colorectal cancer (SW620) | Female athymic nude mice (subcutaneous xenograft model) | qRT-PCR | KRAS | 08–1.0 mL of whole blood | The proportion of mutant cfDNA varied but was overall quite high, suggesting that ctDNA account for an important part of cfDNA. In addition, ctDNA was more fragmented than non-tumor cfDNA. | [28] |
| Human small-cell lung carcinoma (H1975) | C57BL/6- Rag2−/− IL2rg−/− mice model (subcutaneous xenograft model) | qRT-PCR | EGFR mutation | 160–600 µL of plasma | Docetaxel was used to induce apoptosis in a xenograft mice model harboring lung cancer. This treatment promoted apoptosis and facilitated the detection of ctDNA. Of note, the sensitivity of detection was maximal 24 h after treatment. | [29] |
| Human colorectal carcinoma (HCT116-s, SW620, HT29) | Female athymic nude mice (subcutaneous xenograft model) | qRT-PCR | KRAS, BRAF, PSAT1, ACTB | 200 µL of plasma | In this xenograft model, non-tumoral circulating DNA remained low and constant whereas circulating tumor DNA (ctDNA) correlated with tumor progression. Circulating DNA features varied during cancer development. | [30] |
| Human lymphoblastoid cell (RPMI 1788) | Female BALB/c nude mice (subcutaneous xenograft model) | qRT-PCR | Mouse/Human β-actin | 200 µL of plasma | Plasma samples from mice bearing human tumors contained human-specific DNA but also showed higher concentrations of mouse-specific DNA than control mice. This finding suggested that cancer development may also be associated with an increased release of healthy DNA fragments. | [31] |
| Human colon adenocarcinoma (KM12C, DLD-1) | ||||||
| Human lung squamous cell carcinoma (SQ5) | ||||||
| Human epidermoid carcinoma (A431) | ||||||
| Human lung small cell carcinoma (SR-OV-3) | ||||||
| Human breast cancer (MDA-MB-231, MDA-MB-468, KPL-4) | Athymic nude mice (subcutaneous and orthotopic xenograft models) | qRT-PCR | Human GAPDH, hLINE-1 and AluJ | 200 µL of plasma | CtDNA was monitored during tumor progression in preclinical models treated with MEK inhibitor. CtDNA profile reflected tumor burden and response to therapy. Tumor size and levels of ctDNA were decreased in mice treated with MEK inhibitor, compared to controls. Importantly, the different cell lines showed varying ctDNA levels at similar tumor size. | [32] |
| Human colon cancer (Colo205) | ||||||
| Human hepatocellular carcinoma (Huh7, HepG2) | Female athymic nude mice (subcutaneous xenograft model) | qRT-PCR and ddPCR | APOB, FGA and hLINE-1 | 0.5–1.0 mL of whole blood | Treatment with Sorafenib impacted mutation detection. Altogether, qRT-PCR and ddPCR results suggested a clone-dependent release of ctDNA into the bloodstream. | [20] |
| Human colorectal carcinoma (HT29, LoVo, LS174T) | Nu/CD1 nude mice (subcutaneous xenograft model) | ddPCR | KRAS (G12D and G13D) | 2 mL of plasma | Cancer cell lines showed different release of ctDNA in vitro compared to in vivo. Altogether, results suggested that ctDNA release is modulated by multiple factors including non-housekeeping control gateways, selectively regulating the release of DNA fragments into the bloodstream. | [33] |
| Human non-small cell lung cancer (NCI-H1975, NCI-H460) | BALB/c nude mice (subcutaneous xenograft model) | ddPCR | hLINE-1 and ACTB, KRAS and EGFR | Not available | The concentration of ctDNA, but not non-tumor DNA, was positively correlated with tumor weight in both models. The fragmentation and detection rates of EGFR and KRAS mutations in plasma cfDNA increased along with ctDNA concentration and tumor weight. Moreover, H1975 and H460 xenografts showed varying ctDNA levels at similar tumor sizes, suggesting that the amounts of ctDNA released during tumor growth may be specific to each cell line. | [34] |
| Mouse breast cancer (4T1) | BALB/c nude mice (orthotopic xenograft modelusing 4T1 cells expressing Stat3 transcription factor at different levels) | qRT-PCR | GAPDH and B1 | 200 µL of plasma | Three breast cancer xenograft models were generated, with different expression of Stat3. Both ctDNA and CTCs were detected and their detection rates correlated with Stat3 expression. | [35] |
| Human head and neck squamous cell carcinoma (HMS-001, Cal33, Vu147T) | NOD-Scid-Gamma and NOD-Rag-Gamma male mice (subcutaneous xenograft model) | qRT-PCR | hLINE-1 | 1 mL of whole blood | Treatment type and the time interval from the treatment exposure are key factors impacting cfDNA release and detection. In addition, senescence was identified as a novel determinant of cfDNA release. | [36] |
| Human non-small cell lung cancer (HCC-827, PC-9) | ||||||
| Human prostate cancer (LNCaP) | Athymic nude mice (subcutaneous and bone xenograft models) | qRT-PCR | hLINE-1 and Alu | 35 µL of whole blood | Standard methods (slide caliper and bioluminescence) were compared to liquid biopsy to assess tumor burden in metastatic prostate cancer xenograft models treated with radiotherapy. Of note, although most animals showed a transient increase in ctDNA after ionizing radiation, this spike was not observed in animals bearing intratibial tumors. | [37] |
| Human non-small cell lung cancer (H1299, H460, H1975, HCC827) | BALB/c nude mice (subcutaneous xenograft model) | qRT-PCR and ddPCR | hLINE-1, NRAS and EGFR | Not available | A lung cancer xenograft model treated with radiotherapy. Both cfDNA and ctDNA were monitored and circulating mutant DNA was detected. While cfDNA was unchanged at 6 Gy, ctDNA was increased after radiotherapy. | [38] |
| Animal Model | Cancer Type | Technique | Threshold (Volume of Whole Blood) | Main Findings | Reference |
|---|---|---|---|---|---|
| Xenografts and PDX | Breast cancer (BC) | Modified CellTracks system | 0.5–1.0 mL | Establishment of a method to quantify serial changes in CTC in human breast cancer xenografts and PDX. | [42] |
| Xenografts | Liver cancer (HCC) | Flow cytometry using DIVA software | 0.5–1.0 mL | CTC detection is a predictive factor of lung metastasis. 4/11 mice developed metastasis. CTCs were only detected in these 4 metastatic mice whereas no CTCs were detected in 7/11 mice without metastasis. The amount of CTCs correlated with tumor volume, but not with the number of nodules or the largest nodule diameter. | [20] |
| BC | MACS technology | 100 µL | Three breast cancer xenograft models were generated, with different expression of Stat3. Both ctDNA and CTCs were detected, and their detection rates correlated with Stat3 expression. | [35] | |
| PDX | Pancreatic cancer (PDAC) | Microfluidic Chip | 180–1000 µL | PDX deriving from PDAC patients were generated and divided in two groups: a group treated with an oral phosphatidylinositol-3-kinase inhibitor (BKM120) and a group treated with vehicle. Median CTCs count decreased in pre- and post-treatment in the tested group but remained unchanged in the control group. | [43] |
| Colorectal cancer (CRC) | Microfluidic IMD Device | Not available | Blood samples were collected from PDX and submitted to CTCs enumeration in two groups of mice: a group treated with paclitaxel and a vehicle group. CTCs count reflected tumor burden in both groups, but the vehicle group showed higher CTCs count compared to the treated group. In addition, CTCs gradually tended towards a mesenchymal phenotype overtime. | [44] | |
| BC | AccuCyte®-CyteFinder® System | 400–600 µL | CTCs were detected in PDX deriving from patients with breast cancer. | [45] | |
| Quantitative immunohistochemisty (IHC) | 500–700 µL | BC-PDX models can provide a continuous and renewable source of human CTCs. There is a significant association between the presence of CTC clusters and lung metastasis potential. There is variability in CTCs number in different mice within the same PDX line, might attributed to the intratumoral heterogeneity | [46] | ||
| CDX | Breast Cancer (BC) | CellSearch System | 10 mL | Analyzes identified a subpopulation of CTCs associated with the development of metastases in a xenograft assay. This subgroup of CTCs was labeled as metastasis-initiating cells (MICs), expressing EPCAM, CD44, CD47 and MET. | [47] |
| CTC-iChip | 20 mL | Proof-of-concept study on the feasibility to generate cell lines deriving from CTCs. A total of 6 CTCs-derived cell lines were generated. Among 5 cell lines injected in mice, three showed tumorigenic properties (BRx-07, BRx-68 and BRx-61).CTCs-derived cell lines allowed testing drug sensitivity. | [48] | ||
| Multiparametric flow cytometry and CellSearch system | 30–35 mL | CTCs from patients with TNBC were isolated and injected in xenografts, generating CDX. This permitted to interrogate transcriptomics, identifying a 597-genes signature specific of liver metastasis. | [49] | ||
| CellSearch System | 15 mL | A CDX model was developed from CTCs isolated from TNBC patient, demonstrating tumorigenicity. Characterization of CDX revealed WNT signaling as an important driver of these tumors. | [50] | ||
| Lung cancer (SCLC) | CellSearch System | 20 mL | CTCs isolated from SCLC were tumorigenic in immune-compromised mice, allowing to generate CDX. CDX outcomes and response to chemotherapy correlated with patients. | [51] | |
| CTC-iChip | 15–20 mL | CDX were generated with CTCs isolated from SCLC patients, displaying a successful engraftment rate of 38%. CDX deriving from 1 individual but generated from CTCs isolated at different time points reliably recapitulated the drug sensitivity course of that patient. | [52] | ||
| CellSearch System | 10 mL | CDX were longitudinally generated before and upon tumor progression to test new therapeutic options in SCLC. The standard cisplatin/etoposide was compared to a new regimen with a PARP inhibitor olaparib alone or in combination with the WEE1 kinase inhibitor AZD1775 in 10 phenotypically distinct CDX. Response to therapy varied but tended to decrease when tested in CDX generated in tumor progression. | [53] | ||
| Lung cancer (NSCLC) | CellSearch System | 30 mL | CTCs were isolated from a NSCLC and injected in immune-compromised mice, allowing to generate CDX. EpCAM-dependent platform did not detect CTCs while size-based CTCs enrichment permitted to detect an abundant population of CTCs of which a majority expressed mesenchymal surface markers. | [54] | |
| Prostate cancer (PC) and breast cancer (BC) | CellSearch System | 7.5 mL | CDX were generated with CTCs isolated from BC and PC patients. CTCs were detected in 8/8 blood samples, 6/8 bone marrow samples. In addition, human cytokeratin was detected in 6/8 harvested spleens, suggesting a persistant migratory capacity of CTCs in CDX. | [55] | |
| Melanoma | CellSearch System | 7.5 mL | Demonstration of CTC tumorigenicity of advanced melanoma and a strategy to develop animal models when tumor material is inaccessible for PDX generation. CDX tumor growth was detected 1 month after implantation and were representative of patient tumor and treatment response. | [56] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cena, B.; Melloul, E.; Demartines, N.; Dormond, O.; Labgaa, I. Basic Science with Preclinical Models to Investigate and Develop Liquid Biopsy: What Are the Available Data and Is It a Fruitful Approach? Int. J. Mol. Sci. 2022, 23, 5343. https://doi.org/10.3390/ijms23105343
Cena B, Melloul E, Demartines N, Dormond O, Labgaa I. Basic Science with Preclinical Models to Investigate and Develop Liquid Biopsy: What Are the Available Data and Is It a Fruitful Approach? International Journal of Molecular Sciences. 2022; 23(10):5343. https://doi.org/10.3390/ijms23105343
Chicago/Turabian StyleCena, Benedetta, Emmanuel Melloul, Nicolas Demartines, Olivier Dormond, and Ismail Labgaa. 2022. "Basic Science with Preclinical Models to Investigate and Develop Liquid Biopsy: What Are the Available Data and Is It a Fruitful Approach?" International Journal of Molecular Sciences 23, no. 10: 5343. https://doi.org/10.3390/ijms23105343
APA StyleCena, B., Melloul, E., Demartines, N., Dormond, O., & Labgaa, I. (2022). Basic Science with Preclinical Models to Investigate and Develop Liquid Biopsy: What Are the Available Data and Is It a Fruitful Approach? International Journal of Molecular Sciences, 23(10), 5343. https://doi.org/10.3390/ijms23105343