
Treatment with Mesenchymal Stromal Cells Overexpressing Fas-Ligand Ameliorates Acute Graft-versus-Host Disease in Mice

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
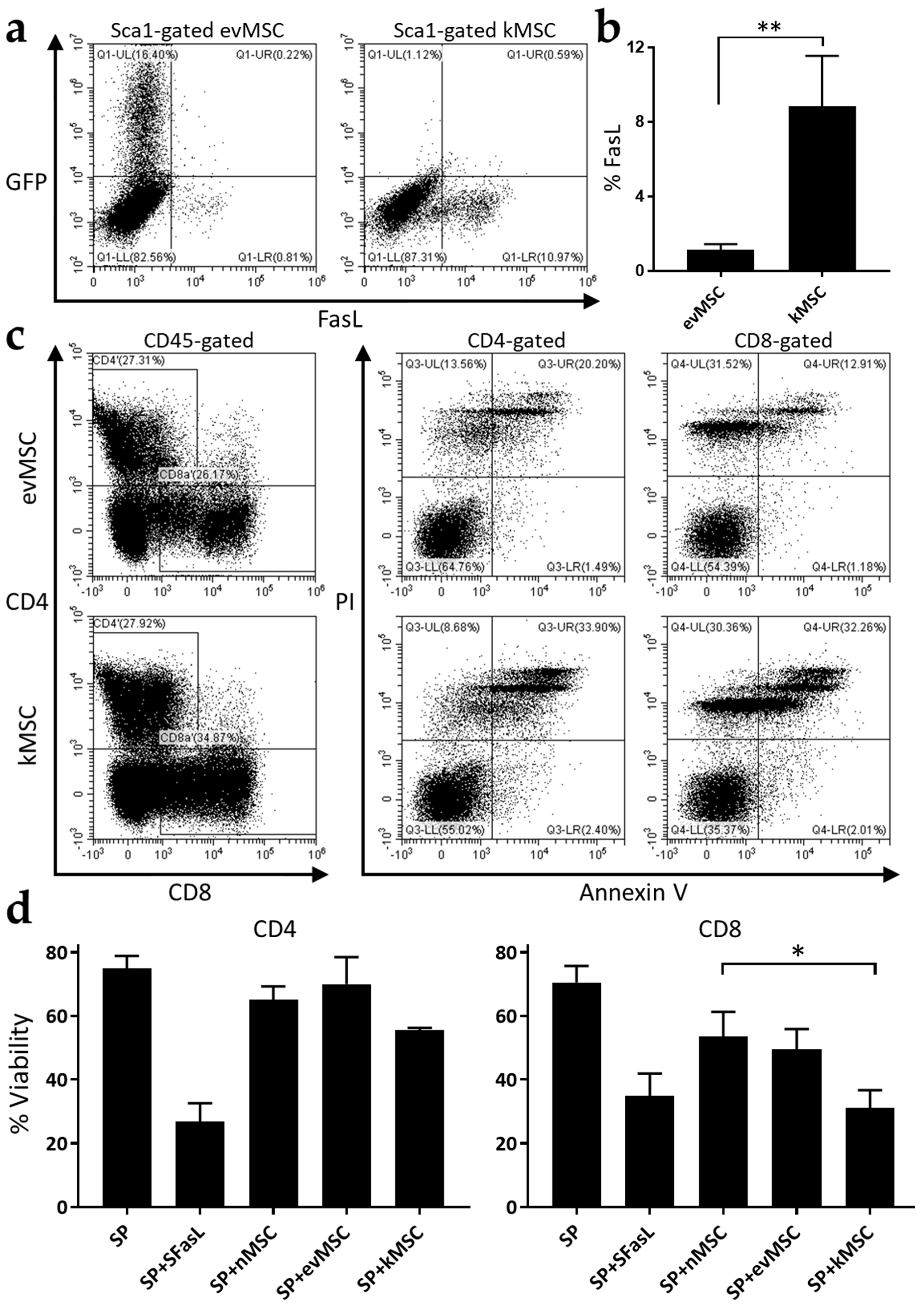
2.1. Generation of Killer MSC
2.2. Treatment with Killer MSC Reduces GvHD Severity and Extends Survival
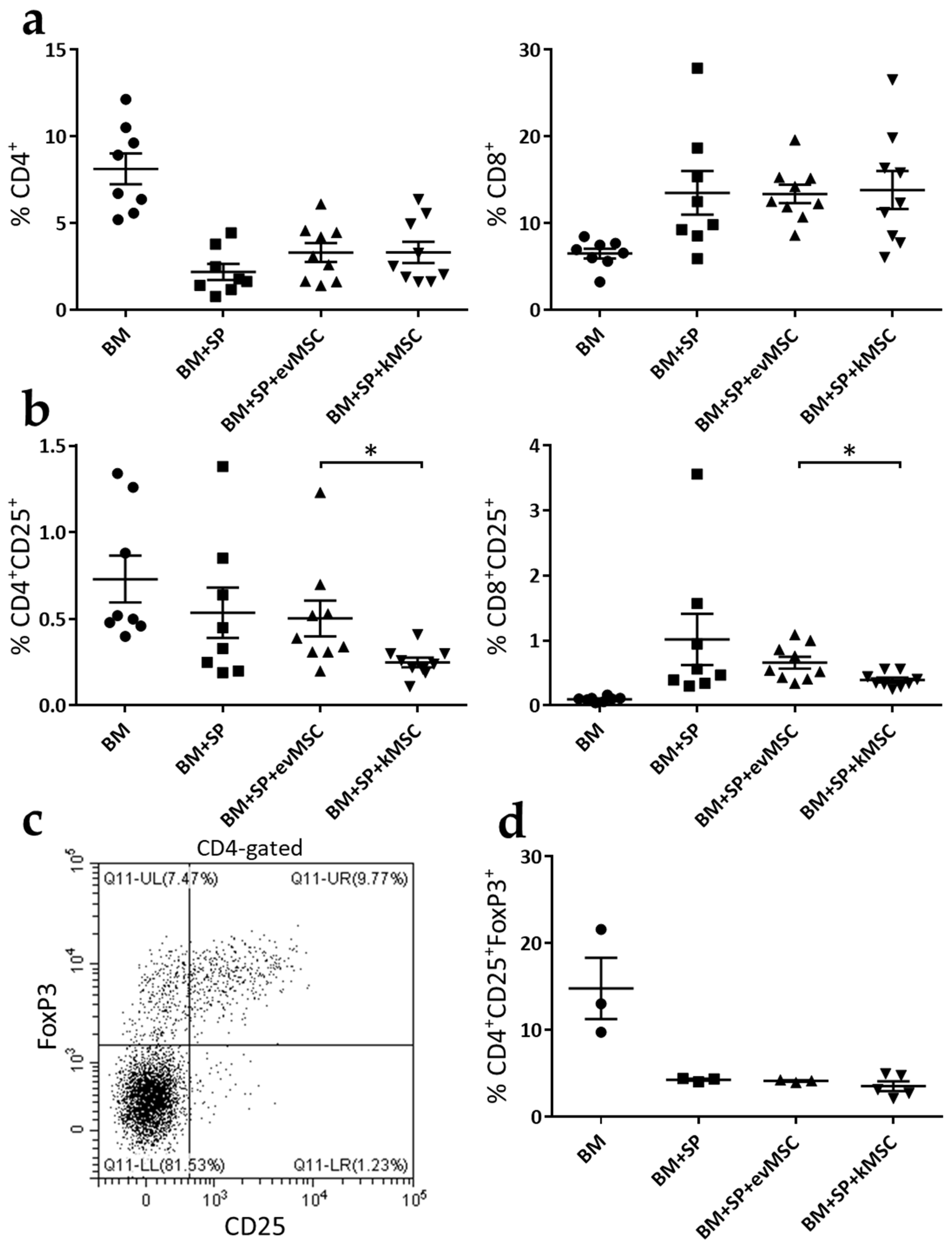
2.3. kMSC Treatment Induces a Decrease in CD25 Expression on Effector T Cells
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. MSC Isolation and Culture
4.3. Osteogenic and Adipogenic Differentiation of MSC
4.4. Viral Transduction of MSC
4.5. Flow Cytometry Analysis
4.6. Immunosuppression Assay
4.7. GvHD Mouse Model and Treatment
4.8. Histology
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Ferrara, J.L.; Levine, J.E.; Reddy, P.; Holler, E. Graft-versus-host disease. Lancet 2009, 373, 1550–1561. [Google Scholar] [CrossRef]
- Zeiser, R.; Blazar, B.R. Acute Graft-versus-Host Disease—Biologic Process, Prevention, and Therapy. N. Engl. J. Med. 2017, 377, 2167–2179. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Peault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.; Jiang, J.; Arp, J.; Liu, W.; Garcia, B.; Wang, H. Regulatory T-cell generation and kidney allograft tolerance induced by mesenchymal stem cells associated with indoleamine 2,3-dioxygenase expression. Transplantation 2010, 90, 1312–1320. [Google Scholar] [CrossRef]
- Weiss, A.R.R.; Dahlke, M.H. Immunomodulation by Mesenchymal Stem Cells (MSCs): Mechanisms of Action of Living, Apoptotic, and Dead MSCs. Front. Immunol. 2019, 10, 1191. [Google Scholar] [CrossRef]
- Morata-Tarifa, C.; Macias-Sanchez, M.D.M.; Gutierrez-Pizarraya, A.; Sanchez-Pernaute, R. Mesenchymal stromal cells for the prophylaxis and treatment of graft-versus-host disease-a meta-analysis. Stem Cell Res. Ther. 2020, 11, 64. [Google Scholar] [CrossRef]
- Nagata, S. Apoptosis by death factor. Cell 1997, 88, 355–365. [Google Scholar] [CrossRef]
- Castro, J.E.; Listman, J.A.; Jacobson, B.A.; Wang, Y.; Lopez, P.A.; Ju, S.; Finn, P.W.; Perkins, D.L. Fas modulation of apoptosis during negative selection of thymocytes. Immunity 1996, 5, 617–627. [Google Scholar] [CrossRef]
- Bouillet, P.; O’Reilly, L.A. CD95, BIM and T cell homeostasis. Nat. Rev. Immunol. 2009, 9, 514–519. [Google Scholar] [CrossRef]
- Volpe, E.; Sambucci, M.; Battistini, L.; Borsellino, G. Fas-Fas Ligand: Checkpoint of T Cell Functions in Multiple Sclerosis. Front. Immunol. 2016, 7, 382. [Google Scholar] [CrossRef] [PubMed]
- Schulte, M.; Reiss, K.; Lettau, M.; Maretzky, T.; Ludwig, A.; Hartmann, D.; de Strooper, B.; Janssen, O.; Saftig, P. ADAM10 regulates FasL cell surface expression and modulates FasL-induced cytotoxicity and activation-induced cell death. Cell Death Differ. 2007, 14, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- LA, O.R.; Tai, L.; Lee, L.; Kruse, E.A.; Grabow, S.; Fairlie, W.D.; Haynes, N.M.; Tarlinton, D.M.; Zhang, J.G.; Belz, G.T.; et al. Membrane-bound Fas ligand only is essential for Fas-induced apoptosis. Nature 2009, 461, 659–663. [Google Scholar] [CrossRef]
- Josefsen, D.; Myklebust, J.H.; Lynch, D.H.; Stokke, T.; Blomhoff, H.K.; Smeland, E.B. Fas ligand promotes cell survival of immature human bone marrow CD34+CD38- hematopoietic progenitor cells by suppressing apoptosis. Exp. Hematol. 1999, 27, 1451–1459. [Google Scholar] [CrossRef]
- Rippo, M.R.; Babini, L.; Prattichizzo, F.; Graciotti, L.; Fulgenzi, G.; Tomassoni Ardori, F.; Olivieri, F.; Borghetti, G.; Cinti, S.; Poloni, A.; et al. Low FasL levels promote proliferation of human bone marrow-derived mesenchymal stem cells, higher levels inhibit their differentiation into adipocytes. Cell Death Dis. 2013, 4, e594. [Google Scholar] [CrossRef] [PubMed]
- Fiorina, P.; Jurewicz, M.; Augello, A.; Vergani, A.; Dada, S.; La Rosa, S.; Selig, M.; Godwin, J.; Law, K.; Placidi, C.; et al. Immunomodulatory function of bone marrow-derived mesenchymal stem cells in experimental autoimmune type 1 diabetes. J. Immunol. 2009, 183, 993–1004. [Google Scholar] [CrossRef]
- Sun, L.; Akiyama, K.; Zhang, H.; Yamaza, T.; Hou, Y.; Zhao, S.; Xu, T.; Le, A.; Shi, S. Mesenchymal stem cell transplantation reverses multiorgan dysfunction in systemic lupus erythematosus mice and humans. Stem Cells 2009, 27, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Mazar, J.; Thomas, M.; Bezrukov, L.; Chanturia, A.; Pekkurnaz, G.; Yin, S.; Kuznetsov, S.A.; Robey, P.G.; Zimmerberg, J. Cytotoxicity mediated by the Fas ligand (FasL)-activated apoptotic pathway in stem cells. J. Biol. Chem. 2009, 284, 22022–22028. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Chen, C.; Wang, D.; Xu, X.; Qu, C.; Yamaza, T.; Cai, T.; Chen, W.; Sun, L.; Shi, S. Mesenchymal-stem-cell-induced immunoregulation involves FAS-ligand-/FAS-mediated T cell apoptosis. Cell Stem Cell 2012, 10, 544–555. [Google Scholar] [CrossRef]
- Vacaru, A.M.; Dumitrescu, M.; Vacaru, A.M.; Fenyo, I.M.; Ionita, R.; Gafencu, A.V.; Simionescu, M. Enhanced Suppression of Immune Cells In Vitro by MSC Overexpressing FasL. Int. J. Mol. Sci. 2020, 22, 348. [Google Scholar] [CrossRef] [PubMed]
- Cooke, K.R.; Kobzik, L.; Martin, T.R.; Brewer, J.; Delmonte, J., Jr.; Crawford, J.M.; Ferrara, J.L. An experimental model of idiopathic pneumonia syndrome after bone marrow transplantation: I. The roles of minor H antigens and endotoxin. Blood 1996, 88, 3230–3239. [Google Scholar] [CrossRef]
- Schroeder, M.A.; Ashami, K.; Staser, K. Modeling Chronic Graft Versus Host Disease in Mice Using Allogeneic Bone Marrow and Splenocyte Transfer. Curr. Protoc. Pharmacol. 2018, 83, e47. [Google Scholar] [CrossRef]
- Ross, S.H.; Cantrell, D.A. Signaling and Function of Interleukin-2 in T Lymphocytes. Annu. Rev. Immunol. 2018, 36, 411–433. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Rasmusson, I.; Sundberg, B.; Gotherstrom, C.; Hassan, M.; Uzunel, M.; Ringden, O. Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet 2004, 363, 1439–1441. [Google Scholar] [CrossRef]
- Cheung, T.S.; Bertolino, G.M.; Giacomini, C.; Bornhauser, M.; Dazzi, F.; Galleu, A. Mesenchymal Stromal Cells for Graft Versus Host Disease: Mechanism-Based Biomarkers. Front. Immunol. 2020, 11, 1338. [Google Scholar] [CrossRef] [PubMed]
- Eggenhofer, E.; Luk, F.; Dahlke, M.H.; Hoogduijn, M.J. The life and fate of mesenchymal stem cells. Front. Immunol. 2014, 5, 148. [Google Scholar] [CrossRef]
- Ullah, M.; Liu, D.D.; Thakor, A.S. Mesenchymal Stromal Cell Homing: Mechanisms and Strategies for Improvement. iScience 2019, 15, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Dumitrescu, M.; Vacaru, A.M.; Trusca, V.G.; Fenyo, I.M.; Ionita, R.; Gafencu, A.V. K2 Transfection System Boosts the Adenoviral Transduction of Murine Mesenchymal Stromal Cells. Int. J. Mol. Sci. 2021, 22, 598. [Google Scholar] [CrossRef]
- Askenasy, N.; Mizrahi, K.; Ash, S.; Askenasy, E.M.; Yaniv, I.; Stein, J. Depletion of naive lymphocytes with fas ligand ex vivo prevents graft-versus-host disease without impairing T cell support of engraftment or graft-versus-tumor activity. Biol. Blood Marrow Transplant. 2013, 19, 185–195. [Google Scholar] [CrossRef]
- Mizrahi, K.; Yaniv, I.; Ash, S.; Stein, J.; Askenasy, N. Apoptotic signaling through Fas and TNF receptors ameliorates GVHD in mobilized peripheral blood grafts. Bone Marrow Transplant. 2014, 49, 640–648. [Google Scholar] [CrossRef][Green Version]
- Yolcu, E.S.; Singh, N.P. Mouse T cells engineered to display on their surface a novel form of FasL protein undergo apoptosis when stimulated with alloantigens: Implications for graft-versus-host disease. Transplant. Proc. 2013, 45, 1850–1852. [Google Scholar] [CrossRef] [PubMed]
- Yolcu, E.S.; Kaminitz, A.; Mizrahi, K.; Ash, S.; Yaniv, I.; Stein, J.; Shirwan, H.; Askenasy, N. Immunomodulation with donor regulatory T cells armed with Fas-ligand alleviates graft-versus-host disease. Exp. Hematol. 2013, 41, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Yarkoni, S.; Kaminitz, A.; Sagiv, Y.; Yaniv, I.; Askenasy, N. Involvement of IL-2 in homeostasis of regulatory T cells: The IL-2 cycle. Bioessays 2008, 30, 875–888. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vacaru, A.M.; Mazilu, A.-M.; Dumitrescu, M.; Fenyo, I.M.; Gafencu, A.V.; Vacaru, A.-M. Treatment with Mesenchymal Stromal Cells Overexpressing Fas-Ligand Ameliorates Acute Graft-versus-Host Disease in Mice. Int. J. Mol. Sci. 2022, 23, 534. https://doi.org/10.3390/ijms23010534
Vacaru AM, Mazilu A-M, Dumitrescu M, Fenyo IM, Gafencu AV, Vacaru A-M. Treatment with Mesenchymal Stromal Cells Overexpressing Fas-Ligand Ameliorates Acute Graft-versus-Host Disease in Mice. International Journal of Molecular Sciences. 2022; 23(1):534. https://doi.org/10.3390/ijms23010534
Chicago/Turabian StyleVacaru, Andrei Mircea, Ana-Maria Mazilu, Madalina Dumitrescu, Ioana Madalina Fenyo, Anca Violeta Gafencu, and Ana-Maria Vacaru. 2022. "Treatment with Mesenchymal Stromal Cells Overexpressing Fas-Ligand Ameliorates Acute Graft-versus-Host Disease in Mice" International Journal of Molecular Sciences 23, no. 1: 534. https://doi.org/10.3390/ijms23010534
APA StyleVacaru, A. M., Mazilu, A.-M., Dumitrescu, M., Fenyo, I. M., Gafencu, A. V., & Vacaru, A.-M. (2022). Treatment with Mesenchymal Stromal Cells Overexpressing Fas-Ligand Ameliorates Acute Graft-versus-Host Disease in Mice. International Journal of Molecular Sciences, 23(1), 534. https://doi.org/10.3390/ijms23010534