Terazosin Stimulates Pgk1 to Remedy Gastrointestinal Disorders


Abstract
:1. Introduction
2. Results
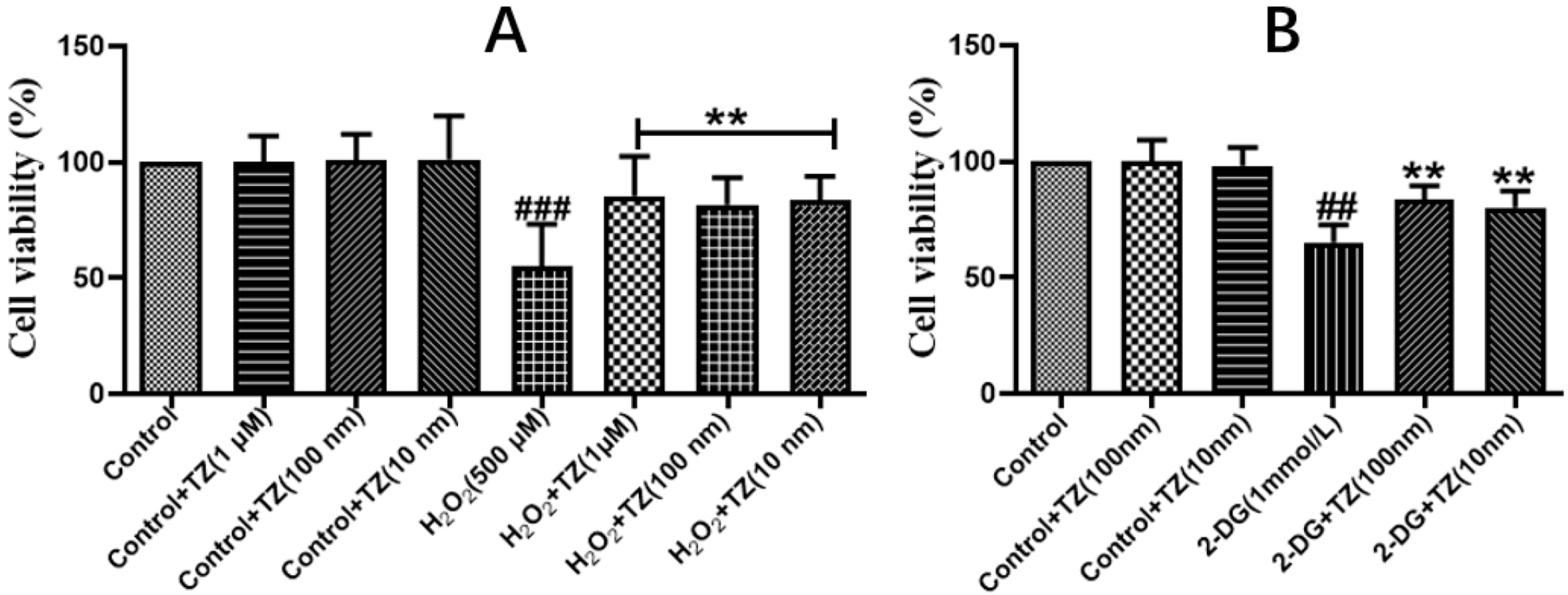
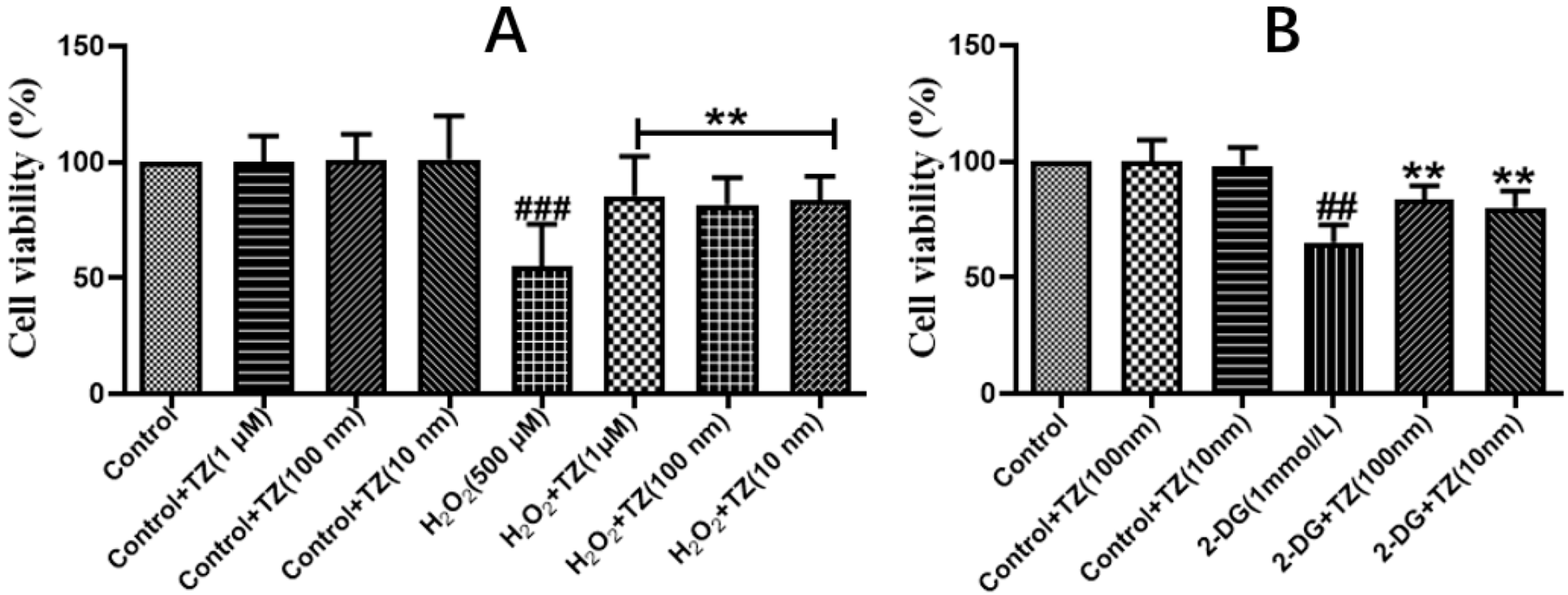
2.1. Effect of Terazosin on Cell Viability under H2O2- and 2-DG-Induced Stresses
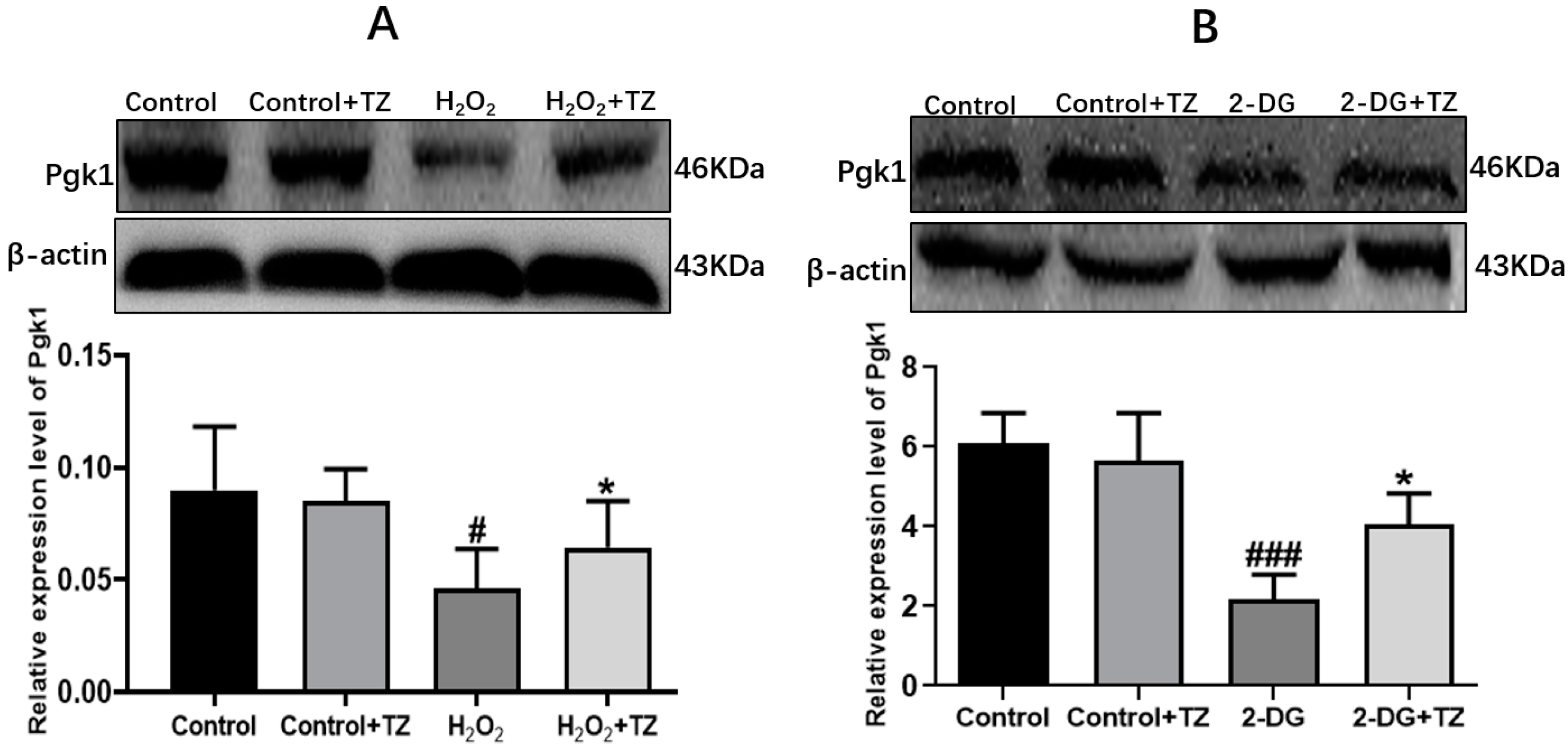
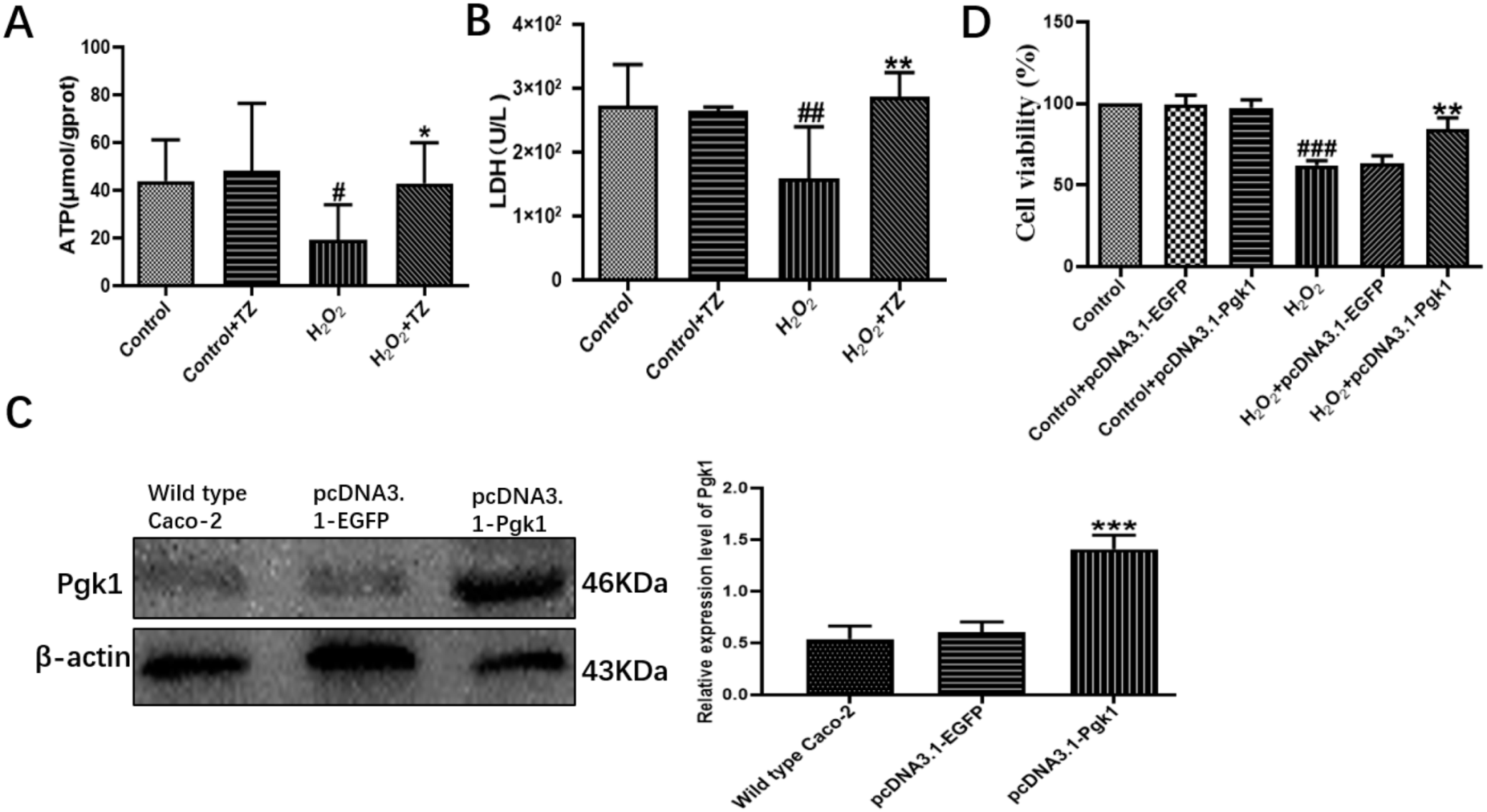
2.2. Terazosin Activates Glucose Metabolism
2.3. Overexpression of Pgk1 in Caco-2 Cells
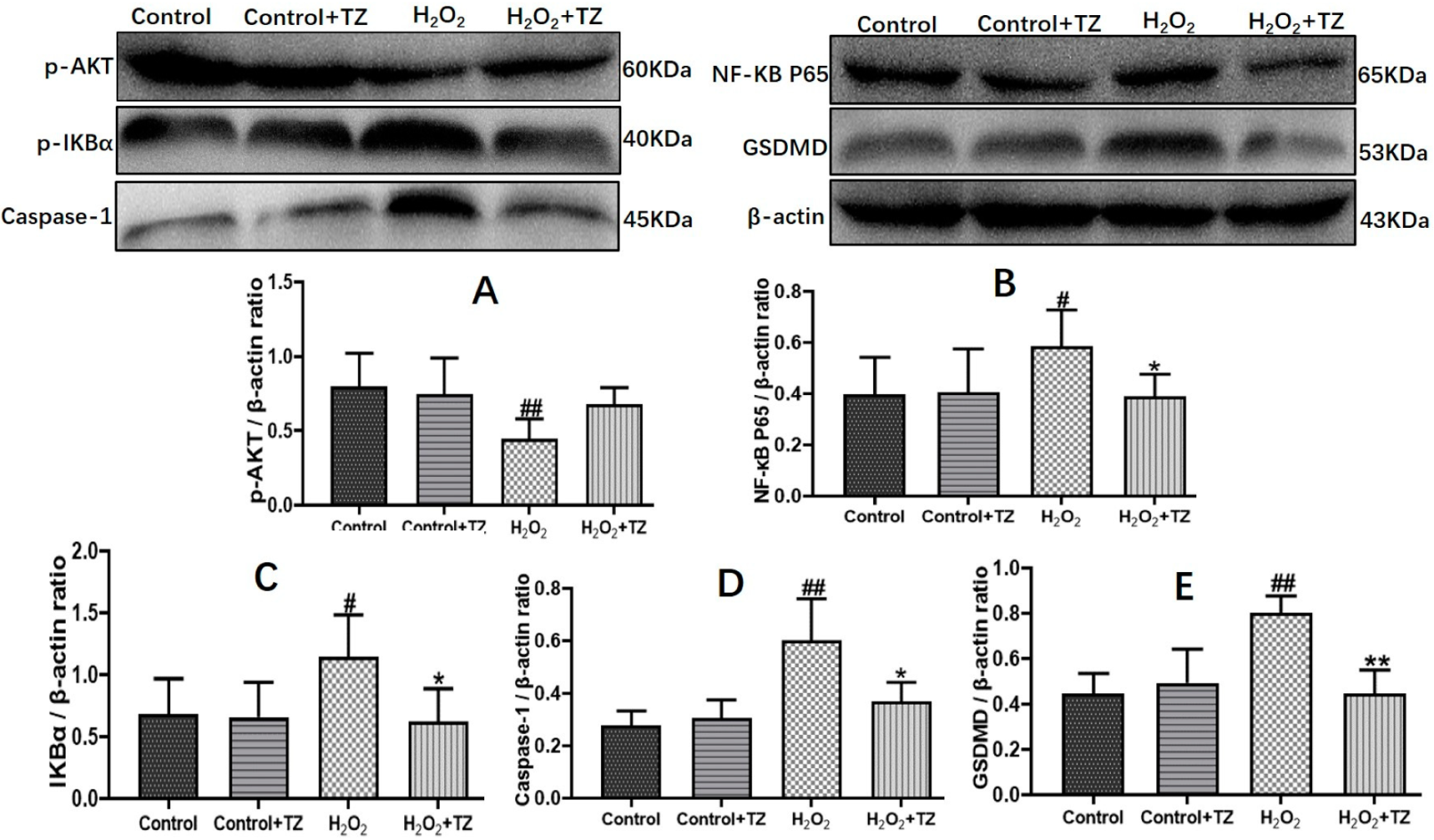
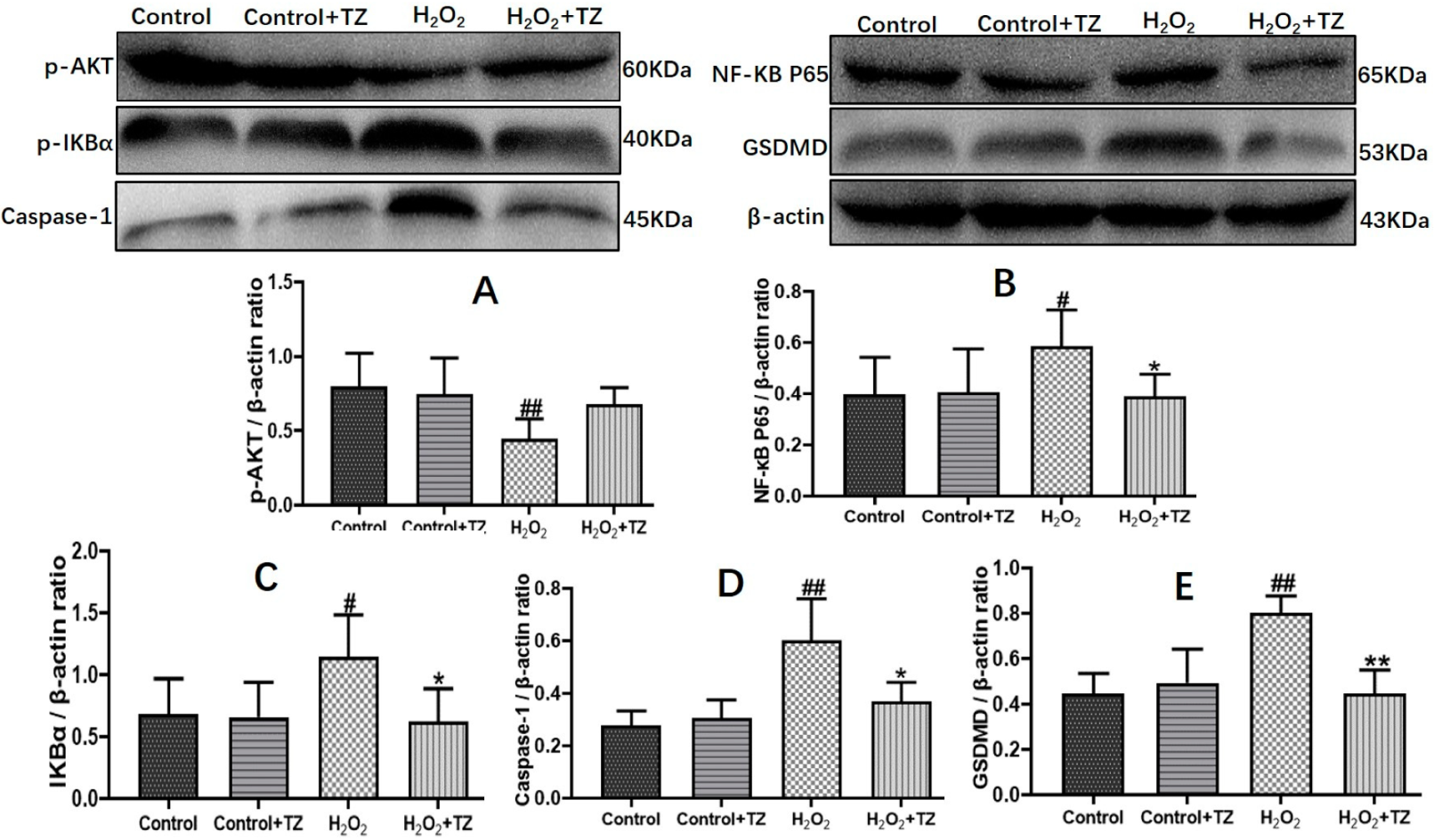
2.4. Terazosin Blocks H2O2-Induced Cell Pyroptosis
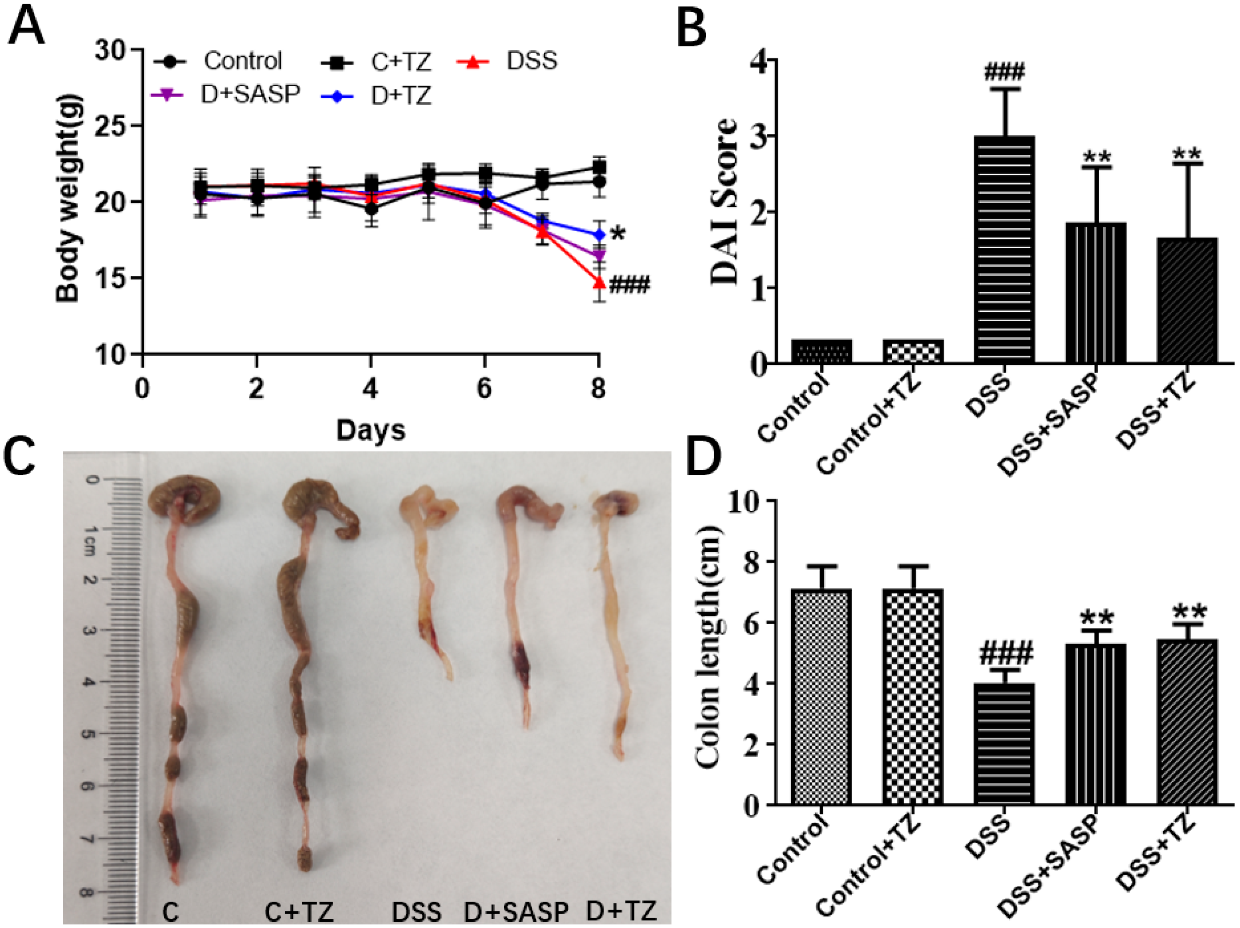
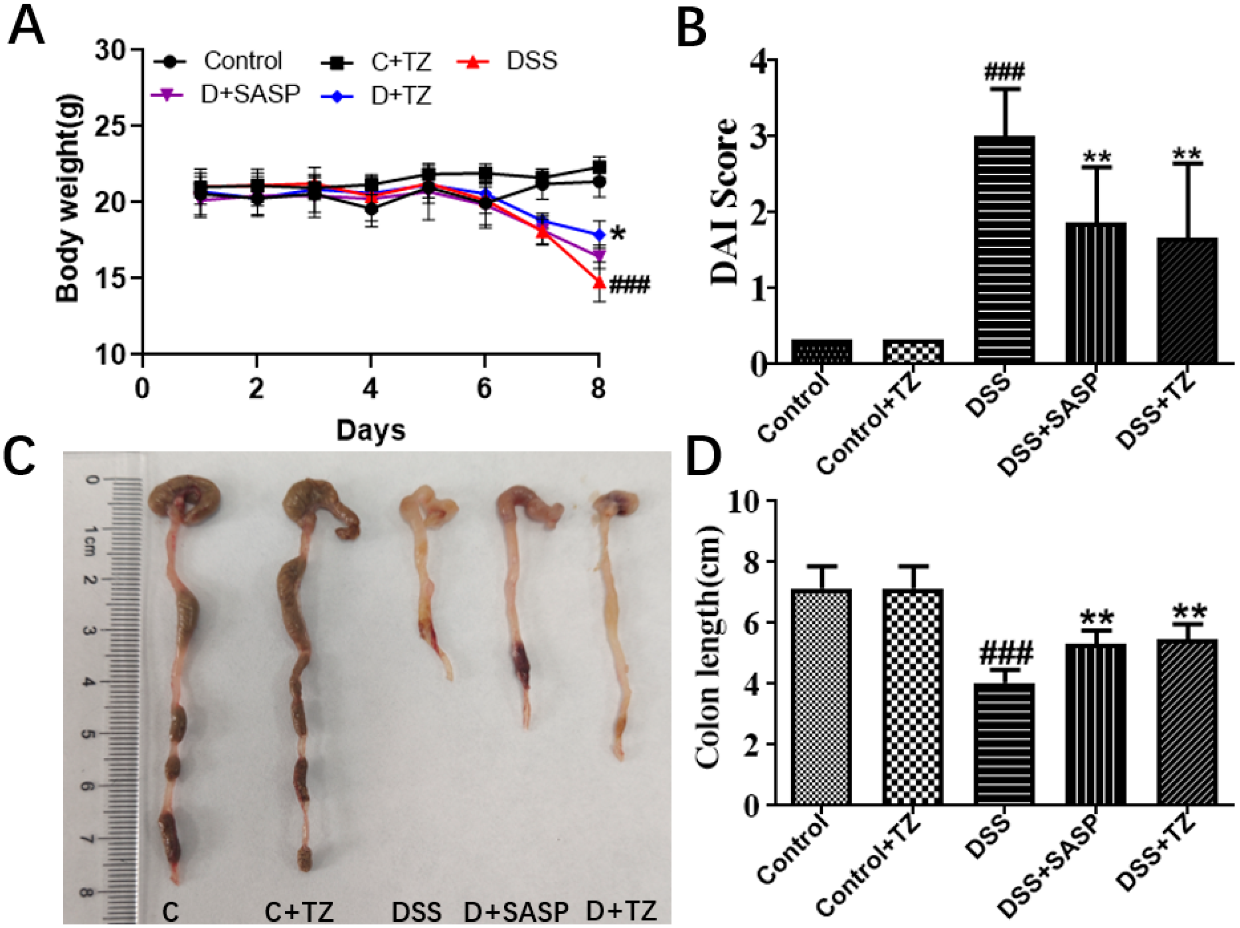
2.5. Terazosin Treatment Attenuates Development of Colitis in Mice
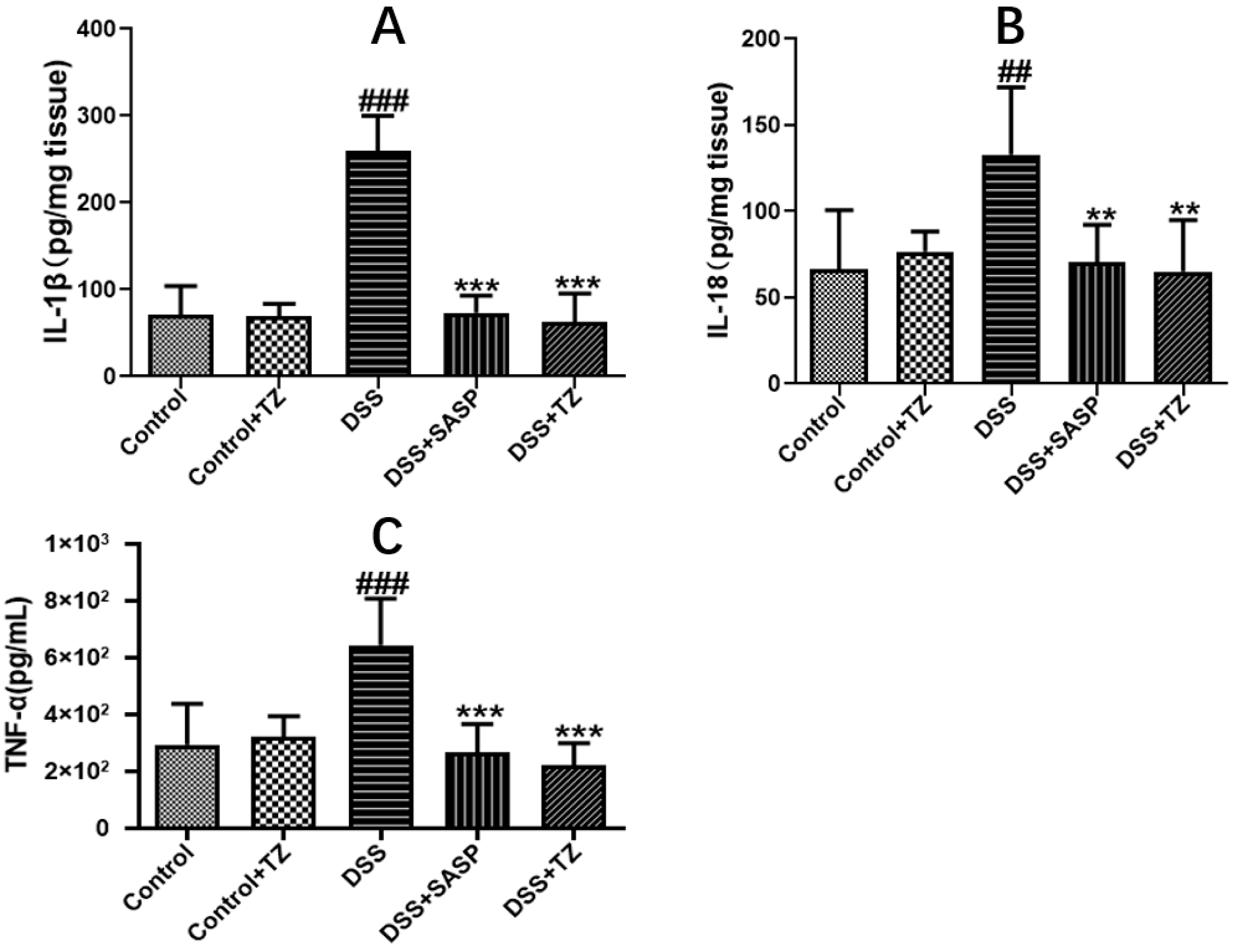
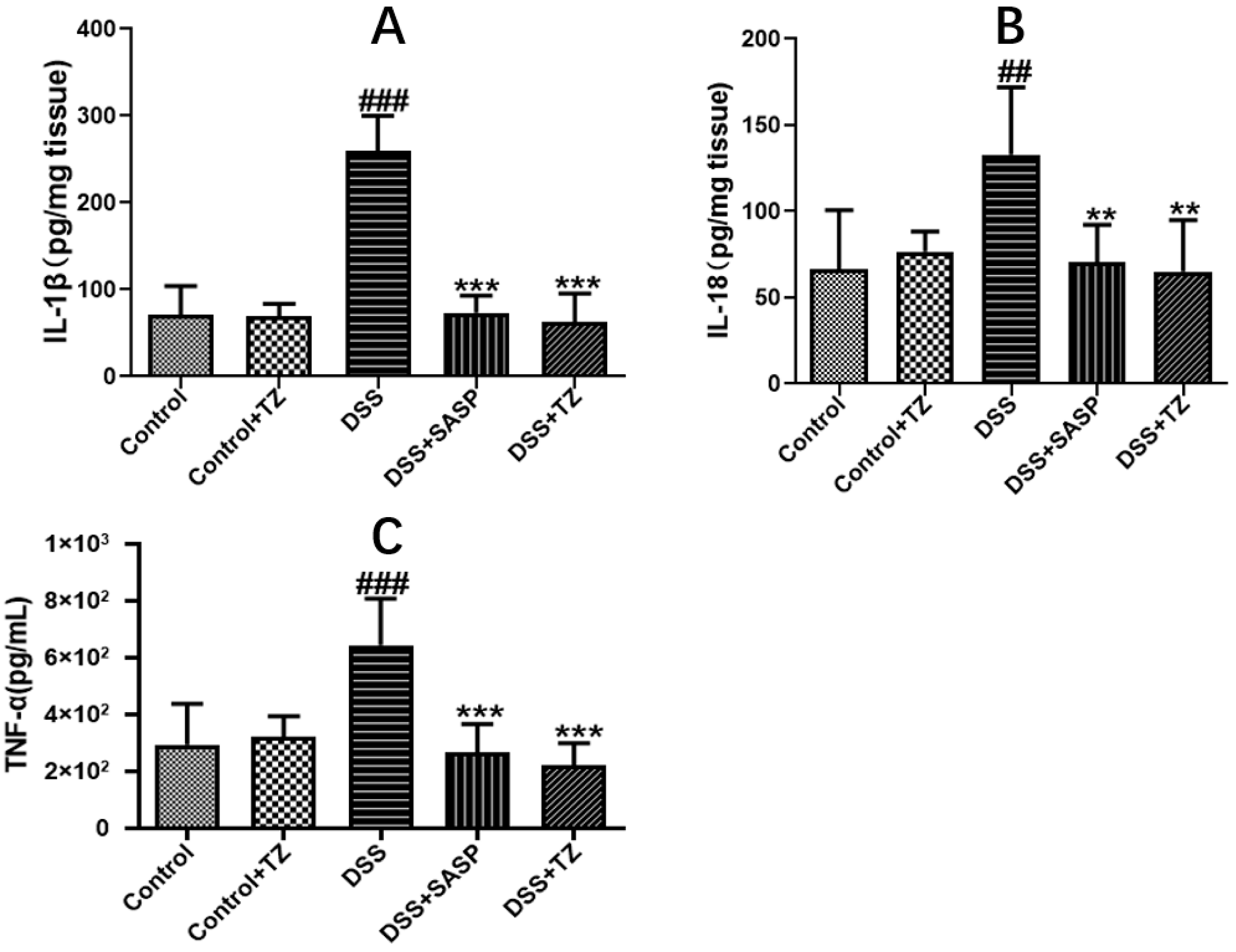
2.6. Terazosin Suppresses Pro-Inflammatory Cytokines in DSS-Induced Colitis in Mice
2.7. Effects of Terazosin on SOD, MDA, Lactic Acid Amounts, and MPO Activity
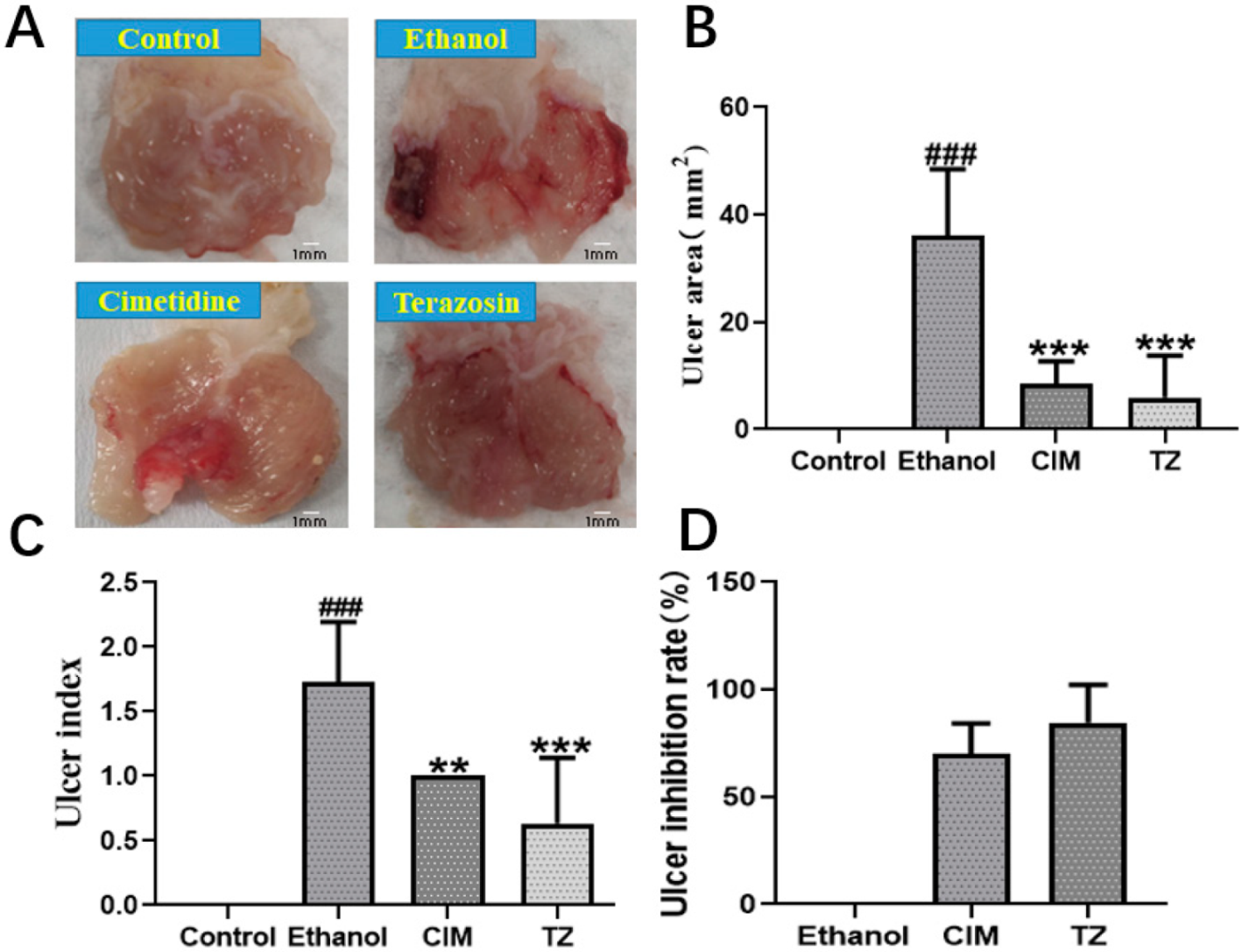
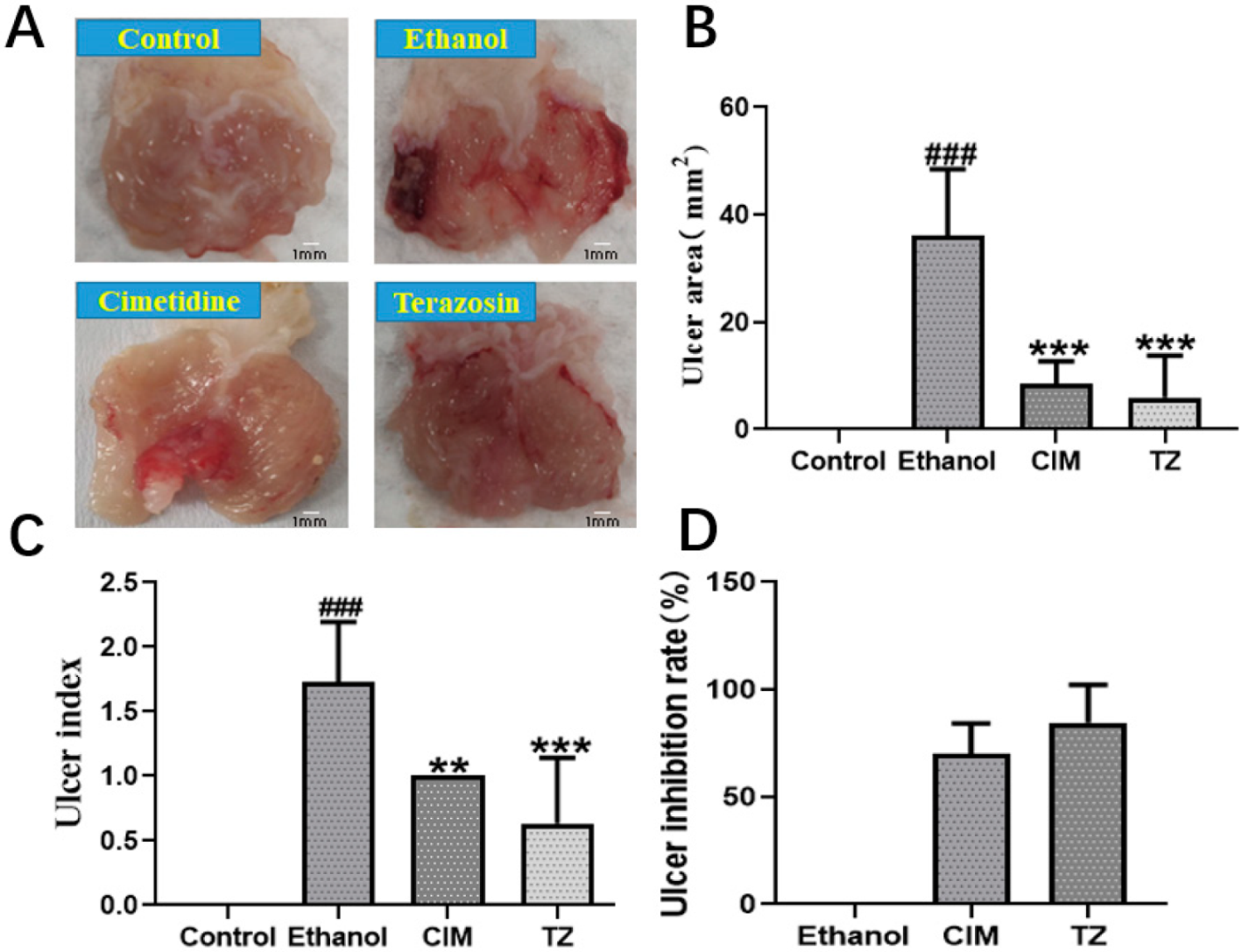
2.8. Effect of Terazosin on Gastric Ulcer in Mice
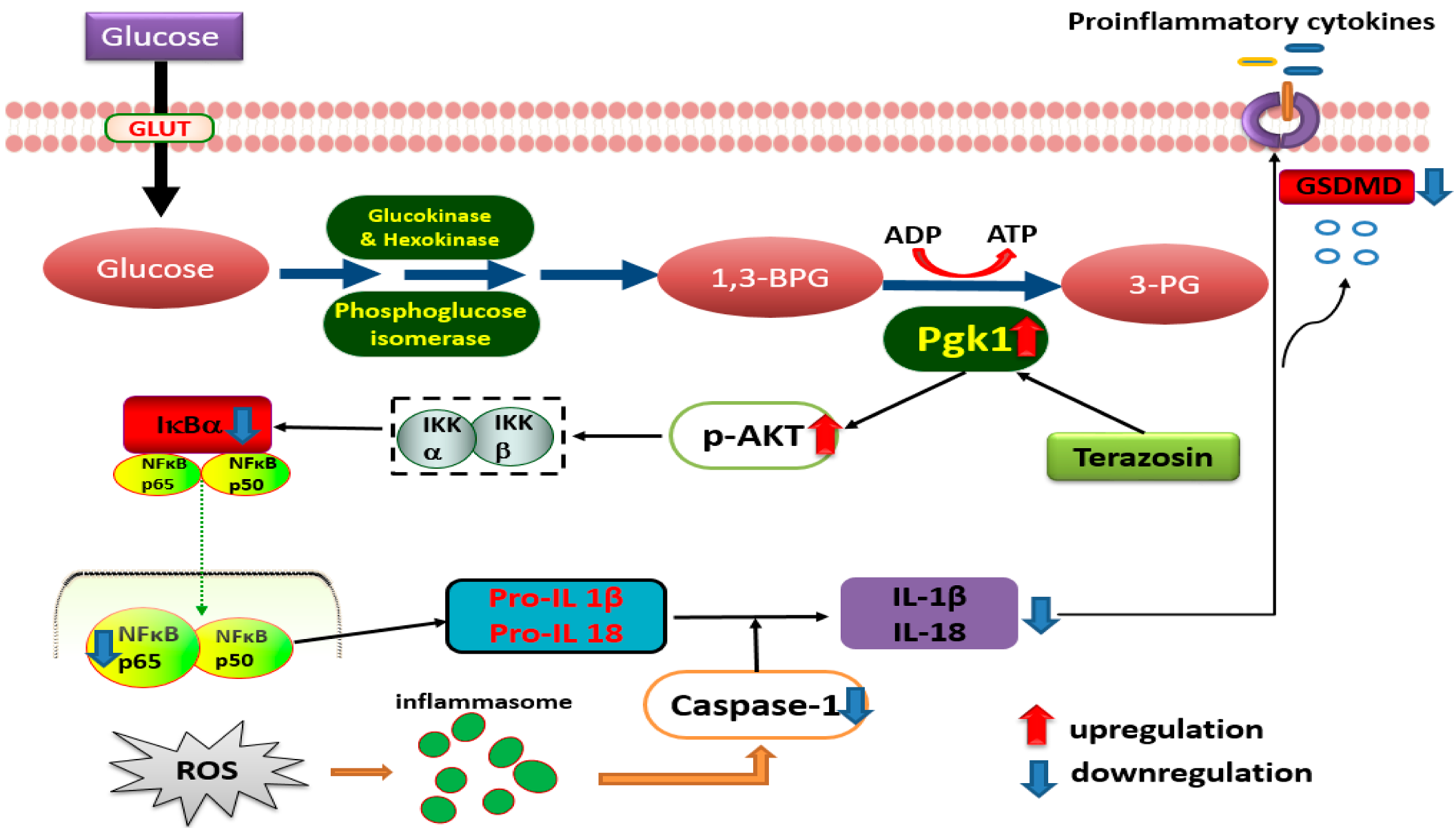
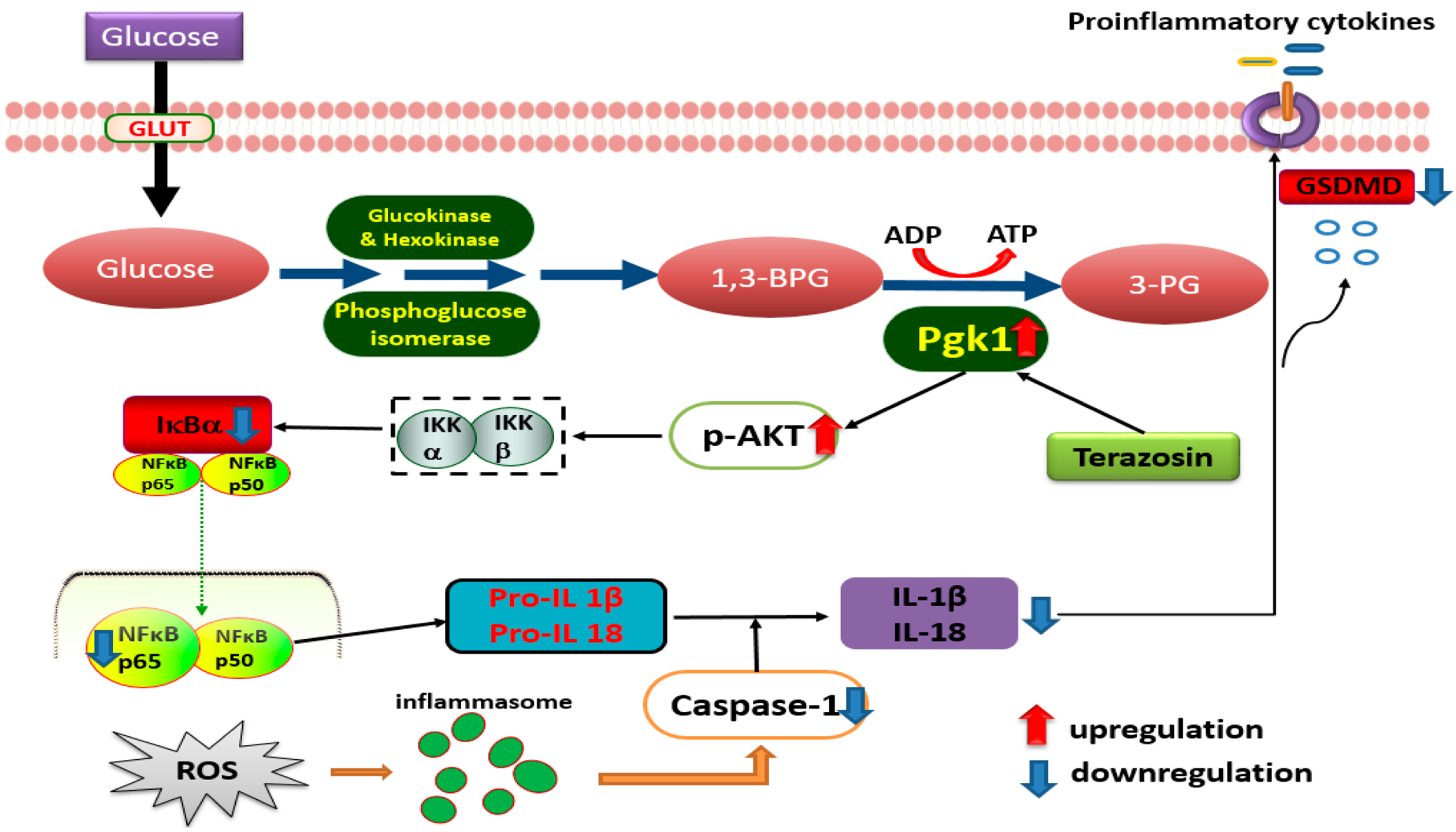
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Cell Lines and Cell Culture
4.3. Cytotoxicity and Viability Assay
4.4. Quantification of Released LDH and Intracellular ATP in Caco-2 Cells
4.5. Transfection of Plasmids
4.6. Western Blot
4.7. Experimental Animals
4.8. Experimental Design
4.9. Assessment of Disease Activity Index
4.10. Measurement of Oxidative Stress Indexes in Serum and Colon Tissue
4.11. Myeloperoxidase Activity Assay in Serum and Colon Tissue
4.12. Measurement of L-LA in Serum and Colon Tissue
4.13. Determination of Cytokines in Colon Tissue
4.14. Ethanol-Induced Gastric Ulcer
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zeng, L.; Wang, Y.; Tang, C. Gastrointestinal: Abdominal actinomycosis: A disease mimicking Crohn’s disease. J. Gastroen. Hepatol. 2021, 36, 300. [Google Scholar] [CrossRef]
- M’Koma, A.E. Inflammatory Bowel Disease: An Expanding Global Health Problem. Clin. Med. Insights Gastroenterol. 2013, 6, 33–47. [Google Scholar] [CrossRef]
- Sattler, S.; Shapiro, L.; Magro, C. Review of Gastrointestinal Manifestations of Kohlmeier-Degos Disease. Am. J. Gastroenterol. 2020, 115, S652–S653. [Google Scholar] [CrossRef]
- Boeing, T.; de Souza, P.; Bonomini, T.J.; Mariano, L.N.B.; Somensi, L.B.; Lucinda, R.M.; Malheiros, A.; da Silva, L.M.; de Andrade, S.F. Antioxidant and anti-inflammatory effect of plumieride in dextran sulfate sodium-induced colitis in mice. Biomed. Pharmacother. 2018, 99, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Gao, X.; Lin, Y.; Yue, C.; Wang, Y.; Quan, F.; Zhang, Z.; Liu, X.; Lu, Y.; Zhan, Y.; et al. Thymopentin ameliorates dextran sulfate sodium-induced colitis by triggering the production of IL-22 in both innate and adaptive lymphocytes. Theranostics 2019, 9, 7490–7505. [Google Scholar] [CrossRef]
- Rapa, S.; Di Paola, R.; Cordaro, M.; Siracusa, R.; D’Amico, R.; Fusco, R.; Autore, G.; Cuzzocrea, S.; Stuppner, H.; Marzocco, S. Plumericin Protects against Experimental Inflammatory Bowel Disease by Restoring Intestinal Barrier Function and Reducing Apoptosis. Biomedicines 2021, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, J. Anti-inflammatory effects of iridoid glycosides fraction of Folium syringae leaves on TNBS-induced colitis in rats. J. Ethnopharmacol. 2011, 133, 780–787. [Google Scholar] [CrossRef]
- Arafa, H.M.M.; Hemeida, R.A.; El-Bahrawy, A.I.M.; Hamada, F.M.A. Prophylactic role of curcumin in dextran sulfate sodium (DSS)-induced ulcerative colitis murine model. Food Chem Toxicol. 2009, 47, 1311–1317. [Google Scholar] [CrossRef]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef]
- Loftus, E.V.; Silverstein, M.D.; Sandborn, W.J.; Tremaine, W.J.; Harmsen, W.S.; Zinsmeister, A.R. Ulcerative colitis in Olmsted County, Minnesota, 1940–1993: Incidence, prevalence, and survival. Gut 2000, 46, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Walker-Smith, J.A.; Lebenthal, E.; Branski, D. Pediatric and Inflammatory Bowel Disease: Perspective and Consequences; Karger: Basel, Switzerland, 2009; p. 204. [Google Scholar]
- Zhuang, H.; Lv, Q.; Zhong, C.; Cui, Y.; He, L.; Zhang, C.; Yu, J. Tiliroside Ameliorates Ulcerative Colitis by Restoring the M1/M2 Macrophage Balance via the HIF-1a/glycolysis Pathway. Front. Immunol. 2021, 12, 951. [Google Scholar] [CrossRef] [PubMed]
- Algieri, F.; Rodriguez-Nogales, A.; Garrido-Mesa, J.; Camuesco, D.; Vezza, T.; Utrilla, P.; Rodriguez-Cabezas, M.E.; Pischel, I.; Galvez, J. Intestinal anti-inflammatory activity of calcium pyruvate in the TNBS model of rat colitis: Comparison with ethyl pyruvate. Biochem. Pharmacol. 2016, 103, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.R.; Gonçalves, P.; Magro, F.; Martel, F. Microbiota-derived butyrate regulates intestinal inflammation: Focus on inflammatory bowel disease. Pharmacol. Res. 2020, 159, 104947. [Google Scholar] [CrossRef]
- Suh, J.H.; Degagne, E.; Gleghorn, E.E.; Setty, M.; Rodriguez, A.; Park, K.T.; Verstraete, S.G.; Heyman, M.B.; Patel, A.S.; Irek, M.; et al. Sphingosine-1-Phosphate Signaling and Metabolism Gene Signature in Pediatric Inflammatory Bowel Disease: A Matched-case Control Pilot Study. Inflamm. Bowel Dis. 2018, 24, 1321–1334. [Google Scholar] [CrossRef]
- Sun, X.; Yao, L.; Liang, H.; Wang, D.; He, Y.; Wei, Y.; Ye, L.; Wang, K.; Li, L.; Chen, J.; et al. Intestinal epithelial PKM2 serves as a safeguard against experimental colitis via activating β-catenin signaling. Mucosal Immunol. 2019, 12, 1280–1290. [Google Scholar] [CrossRef]
- Iatsenko, I.; Boquete, J.-P.; Lemaitre, B. Microbiota-Derived Lactate Activates Production of Reactive Oxygen Species by the Intestinal NADPH Oxidase Nox and Shortens Drosophila Lifespan. Immunity 2018, 49, 929–942.e5. [Google Scholar] [CrossRef] [Green Version]
- Tian, N.; Hu, L.; Lu, Y.; Tong, L.; Feng, M.; Liu, Q.; Li, Y.; Zhu, Y.; Wu, L.; Ji, Y.; et al. TKT maintains intestinal ATP production and inhibits apoptosis-induced colitis. Cell Death Dis. 2021, 12, 853. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, S.; Jeon, R.; Vuckovic, I.; Jiang, X.; Lerman, A.; Folmes, C.; Dzeja, P.D.; Herrmann, J. Interferon Gamma Induces Reversible Metabolic Reprogramming of M1 Macrophages to Sustain Cell Viability and Pro-Inflammatory Activity. EBioMedicine 2018, 30, 303–316. [Google Scholar] [CrossRef] [Green Version]
- Ward, S.K.; Stevens, C.A.; Keates-Baleeiro, J.; Bhakta, M. Therapeutic Benefit of Blood Transfusion in a Patient with Novel PGK1 Mutation (c.461T>C [p.L154P]). J. Pediatr. Hematol. 2020, 42, e228–e230. [Google Scholar] [CrossRef]
- Chiarelli, L.R.; Morera, S.M.; Bianchi, P.; Fermo, E.; Zanella, A.; Galizzi, A.; Valentini, G. Molecular Insights on Pathogenic Effects of Mutations Causing Phosphoglycerate Kinase Deficiency. PLoS ONE 2012, 7, e32065. [Google Scholar] [CrossRef] [Green Version]
- Pey, A.L.; Mesa-Torres, N.; Chiarelli, L.R.; Valentini, G. Structural and Energetic Basis of Protein Kinetic Destabilization in Human Phosphoglycerate Kinase 1 Deficiency. Biochemistry 2013, 52, 1160–1170. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, C.; Li, X.; Wang, T.; Li, Y.; Cao, C.; Ding, Y.; Dong, M.; Finci, L.; Wang, J.-H.; et al. Terazosin activates Pgk1 and Hsp90 to promote stress resistance. Nat. Chem. Biol. 2015, 11, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Yu, Z. Phosphoglycerate kinase 1 (PGK1) in cancer: A promising target for diagnosis and therapy. Life Sci. 2020, 256, 117863. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Ni, J.; Wu, X.; Wei, H.; Peng, J. GPA peptide inhibits NLRP3 inflammasome activation to ameliorate colitis through AMPK pathway. Aging-Us 2020, 12, 18522–18544. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Yang, H.; Tao, Y.; Wei, S.; Li, L.; Liu, M.; Li, J. Artesunate ameliorates DSS-induced ulcerative colitis by protecting intestinal barrier and inhibiting inflammatory response. Inflammation 2020, 43, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Vernia, P.; Gnaedinger, A.; Hauck, W.; Breuer, R.I. Organic anions and the diarrhea of inflammatory bowel disease. Am. J. Dig. Dis. 1988, 33, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-M.; Li, B.-C.; Li, P.; Liu, J.-Z.; Cui, J.-L.; Mei, Z.-Q. Effects of Na-FA on gastrointestinal movement and gastric ulcer in mice. J. Chin. Med. Mater. 2011, 34, 1565–1569. [Google Scholar]
- Al-Wajeeh, N.S.; Hajerezaie, M.; Noor, S.M.; Halabi, M.F.; Al-Henhena, N.; Azizan, A.H.S.; Kamran, S.; Hassandarvish, P.; Shwter, A.N.; Karimian, H.; et al. The gastro protective effects of Cibotium barometz hair on ethanol-induced gastric ulcer in Sprague-Dawley rats. BMC Veter. Res. 2016, 13, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolgazi, M.; Ozdemir-Kumral, Z.N.; Cantali-Ozturk, C.; Demirci, E.K.; Yuksel, M.; Sirvanci, S.; Yegen, B.C. Anti-inflammatory effects of nesfatin-1 on acetic acid-induced gastric ulcer in rats: Involvement of cyclo-oxygenase pathway. J. Physiol. Pharmacol. 2017, 68, 765–777. [Google Scholar]
- Chen, H.; Liao, H.; Liu, Y.; Zheng, Y.; Wu, X.; Su, Z.; Zhang, X.; Lai, Z.; Lai, X.; Lin, Z.X.; et al. Protective effects of po-gostone from Pogostemonis Herba against ethanol-induced gastric ulcer in rats. Fitoterapia 2015, 100, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Lin, Q.; Li, X.; Nie, Y.; Wang, L.; Shi, L.; Xu, W.; Hu, T.; Guo, T.; Luo, F. Octacosanol Attenuates Inflammation in Both RAW264.7 Macrophages and a Mouse Model of Colitis. J. Agric. Food Chem. 2017, 65, 3647–3658. [Google Scholar] [CrossRef]
- Liu, F.; Yao, Y.; Lu, Z.; Zhang, Q.; Liu, C.; Zhu, C.; Lin, C. 5-Hydroxy-4-methoxycanthin-6-one alleviates dextran sodium sulfate-induced colitis in rats via regulation of metabolic profiling and suppression of NF-κB/p65 signaling pathway. Phytomedicine 2020, 82, 153438. [Google Scholar] [CrossRef] [PubMed]
- Berends, S.E.; Strik, A.S.; Löwenberg, M.; D’Haens, G.R.; Mathôt, R.A.A. Clinical Pharmacokinetic and Pharmacodynamic Considerations in the Treatment of Ulcerative Colitis. Clin. Pharmacokinet. 2019, 58, 15–37. [Google Scholar] [CrossRef] [Green Version]
- Stevceva, L.; Pavli, P.; Matthei, K.; Young, I.; Doe, W.F. Dextran Sulfate Sodium (Dss)-Induced Colitis Develops in Il-5 Knockout Mice. Gastroenterology 1995, 108, A922. [Google Scholar]
- Dieleman, L.A.; Ridwan, B.U.; Tennyson, G.S.; Beagley, K.W.; Elson, C.O. Dextran Sodium-Sulfate (Dss)-Induced Colitis Occurs in Severe Combined Immunodeficient (Scid) Mice. Gastroenterology 1993, 104, A692. [Google Scholar]
- Swarnakar, S.; Mishra, A.; Ganguly, K.; Sharma, A.V. Matrix metalloproteinase-9 activity and expression is reduced by mel-atonin during prevention of ethanol-induced gastric ulcer in mice. J. Pineal Res. 2007, 43, 56–64. [Google Scholar] [CrossRef]
- Pan, L.R.; Tang, Q.; Fu, Q.; Hu, B.R.; Xiang, J.Z.; Qian, J.Q. Roles of nitric oxide in protective effect of berberine in ethanol-induced gastric ulcer mice. Acta Pharmacol. Sin. 2005, 26, 1334–1338. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Lin, Q.; Yang, T.; Zeng, L.; Shi, L.; Chen, Y.; Luo, F. Oat β-glucan ameliorates dextran sulfate sodium (DSS)-induced ulcerative colitis in mice. Food Funct. 2015, 6, 3454–3463. [Google Scholar] [CrossRef]
- George, M.Y.; Esmat, A.; Tadros, M.G.; El-Demerdash, E. In vivo cellular and molecular gastroprotective mechanisms of chrysin; Emphasis on oxidative stress, inflammation and angiogenesis. Eur. J. Pharmacol. 2018, 818, 486–498. [Google Scholar] [CrossRef]
- Sahu, B.D.; Kumar, J.M.; Sistla, R. Fisetin, a dietary flavonoid, ameliorates experimental colitis in mice: Relevance of NF-κB signaling. J. Nutr. Biochem. 2016, 28, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Seril, D.N.; Liao, J.; Yang, G.-Y.; Yang, C.S. Oxidative stress and ulcerative colitis-associated carcinogenesis: Studies in humans and animal models. Carcinogenesis 2003, 24, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Shen, Y.; Shi, H.; Ma, X.; Lin, B.; Xiao, T.; Wu, F.; Zhu, J.; Li, Z.; Xiao, J.; et al. Gastroprotective effects of Kangfuxin-against ethanol-induced gastric ulcer via attenuating oxidative stress and ER stress in mice. Chem. Interact. 2016, 260, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Wang, J.Y.; Liu, L.; Li, Y.X.; Xun, A.Y.; Zeng, W.S.; Jia, C.H.; Wei, X.X.; Feng, J.L.; Zhao, L.; et al. Antioxidant Effects of Resveratrol on Mice with DSS-induced Ulcerative Colitis. Arch. Med. Res. 2010, 41, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Yuan, J.-Q.; Lv, Y.-B.; Gao, X.; Yin, Z.-X.; Kraus, V.B.; Luo, J.-S.; Chei, C.-L.; Matchar, D.B.; Zeng, Y.; et al. Associations between superoxide dismutase, malondialdehyde and all-cause mortality in older adults: A community-based cohort study. BMC Geriatr. 2019, 19, 104. [Google Scholar] [CrossRef] [PubMed]
- Lacey, C.A.; Mitchell, W.J.; Dadelahi, A.S.; Skyberg, J.A. Caspase-1 and Caspase-11 Mediate Pyroptosis, Inflammation, and Control of Brucella Joint Infection. Infect. Immun. 2018, 86, 9. [Google Scholar] [CrossRef] [Green Version]
- Alavala, S.; Sangaraju, R.; Nalban, N.; Sahu, B.D.; Jerald, M.K.; Kilari, E.K.; Sistla, R. Stevioside, a diterpenoid glycoside, shows anti-inflammatory property against Dextran Sulphate Sodium-induced ulcerative colitis in mice. Eur. J. Pharmacol. 2019, 855, 192–201. [Google Scholar] [CrossRef]
- Man, S.M.; Kanneganti, T.-D. Gasdermin D: The long-awaited executioner of pyroptosis. Cell Res. 2015, 25, 1183–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Q.; Yi, T.; Chen, C. NF-kB-Gasdermin D (GSDMD) Axis Couples Oxidative Stress and NACHT, LRR and PYD Do-mains-Containing Protein 3 (NLRP3) Inflammasome-Mediated Cardiomyocyte Pyroptosis Following Myocardial Infarction. Med. Sci. Monit. 2018, 24, 6044–6052. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gan, L.; Xu, Y.; Luo, D.; Ren, Q.; Wu, S.; Sun, C. Melatonin alleviates inflammasome-induced pyroptosis through inhibiting NF-κB/ GSDMD signal in mice adipose tissue. J. Pineal Res. 2017, 63, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Viatour, P.; Merville, M.P.; Bours, V.; Chariot, A. Phosphorylation of NF-kB and IkB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Chung, K.S.; Jin, B.R.; Cheon, S.Y.; Nugroho, A.; Roh, S.S.; An, H.J. Anti-inflammatory effects of an ethanol extract of Aster glehni via inhibition of NF-κB activation in mice with DSS-induced colitis. Food Funct. 2017, 8, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Wan, Z.P.; Han, L.; Liu, H.Y.; Li, H.S. Effect of Huangqin Decoction on pyroptosis pathway of NLRP3/caspase-1 in mice with ulcerative colitis. China J. Chin. Mater. Med. 2021, 46, 1191–1196. [Google Scholar]
- Kondo, Y.; Rose, I.; Young, G.P.; Whitehead, R.H. Growth of Characteristic Epithelial-Cells from Rat Esophagus, Fundus of Stomach, Antrum of Stomach, Small-Intestine, and Colon in Monolayer-Culture. Gastroenterology 1983, 84, 1212. [Google Scholar]
- Medicherla, K.; Ketkar, A.; Sahu, B.D.; Sudhakar, G.; Sistla, R. Rosmarinus officinalis L. extract ameliorates intestinal inflammation through MAPKs/NF-κB signaling in a murine model of acute experimental colitis. Food Funct. 2016, 7, 3233–3243. [Google Scholar] [CrossRef] [PubMed]
- Ho, E.; Galougahi, K.K.; Liu, C.C.; Bhindi, R.; Figtree, G.A. Biological markers of oxidative stress: Applications to cardiovascular research and practice. Redox Biol. 2013, 1, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Park, C.-H.; Nam, D.-Y.; Son, H.-U.; Lee, S.-R.; Lee, H.-J.; Heo, J.-C.; Cha, T.-Y.; Baek, J.-H.; Lee, S.-H. Polymer fraction of Aloe vera exhibits a protective activity on ethanol-induced gastric lesions. Int. J. Mol. Med. 2011, 27, 511–518. [Google Scholar]
- Indran, M.; Mahmood, A.A.; Kuppusamy, U.R. Protective effect of Carica papaya L leaf extract against alcohol induced acute gastric damage and blood oxidative stress in rats. West Indian Med. J. 2008, 57, 323–326. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Colon Tissue | Serum | |||||||
|---|---|---|---|---|---|---|---|---|
| Groups | Dose | Lactic Acid (mmol/gprot) | MPO(U/g) | SOD (U/mgprot) | MDA (nmol/mgprot) | MPO(U/L) | SOD(U/mL) | MDA (nmol/mL) |
| Control | - | 0.04 ± 0.004 | 58.45 ± 8.51 | 83.57 ± 2.38 | 58.45 ± 8.51 | 56.89 ± 8.84 | 83.57 ± 2.38 | 3.07 ± 0.68 |
| Control + TZ | 4 mg/kg/day | 0.04 ± 0.004 | 52.72 ± 10.42 | 81.95 ± 3.92 | 52.72 ± 10.42 | 48.45 ± 6.57 | 81.95 ± 3.92 | 0.52 ± 0.33 |
| DSS | - | 0.13 ± 0.006 ### | 99.75 ± 11.74 ### | 69.44 ± 5.24 ### | 99.75 ± 11.74 ## | 134.29 ± 13.24 ### | 69.44±5.24 # | 39.26 ± 4.47 ### |
| DSS + SASP | 80 mg/kg/day | 0.09 ± 0.009 *** | 18.26 ± 4.45 *** | 83.29 ± 3.52 *** | 18.26 ± 4.46 *** | 61.06 ± 6.37 *** | 83.29 ± 3.52 * | 6.86 ± 1.09 *** |
| DSS + TZ | 4 mg/kg/day | 0.08 ± 0.006 *** | 34.87 ± 13.42 *** | 80.67 ± 3.81 *** | 34.88 ± 13.42 *** | 76.11 ± 1.02 ** | 80.67 ± 3.81 | 2.76 ± 1.34 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhao, W.; Li, C.; Wu, T.; Han, L.; Hu, Z.; Li, X.; Zhou, J.; Chen, X. Terazosin Stimulates Pgk1 to Remedy Gastrointestinal Disorders. Int. J. Mol. Sci. 2022, 23, 416. https://doi.org/10.3390/ijms23010416
Liu J, Zhao W, Li C, Wu T, Han L, Hu Z, Li X, Zhou J, Chen X. Terazosin Stimulates Pgk1 to Remedy Gastrointestinal Disorders. International Journal of Molecular Sciences. 2022; 23(1):416. https://doi.org/10.3390/ijms23010416
Chicago/Turabian StyleLiu, Jingjing, Wenyang Zhao, Chun Li, Tongyu Wu, Liang Han, Zhuozhou Hu, Xiangxiang Li, Jing Zhou, and Xinping Chen. 2022. "Terazosin Stimulates Pgk1 to Remedy Gastrointestinal Disorders" International Journal of Molecular Sciences 23, no. 1: 416. https://doi.org/10.3390/ijms23010416
APA StyleLiu, J., Zhao, W., Li, C., Wu, T., Han, L., Hu, Z., Li, X., Zhou, J., & Chen, X. (2022). Terazosin Stimulates Pgk1 to Remedy Gastrointestinal Disorders. International Journal of Molecular Sciences, 23(1), 416. https://doi.org/10.3390/ijms23010416