
Neuroprotective Effect of Valproic Acid on Salicylate-Induced Tinnitus

 , , ,
, , ,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
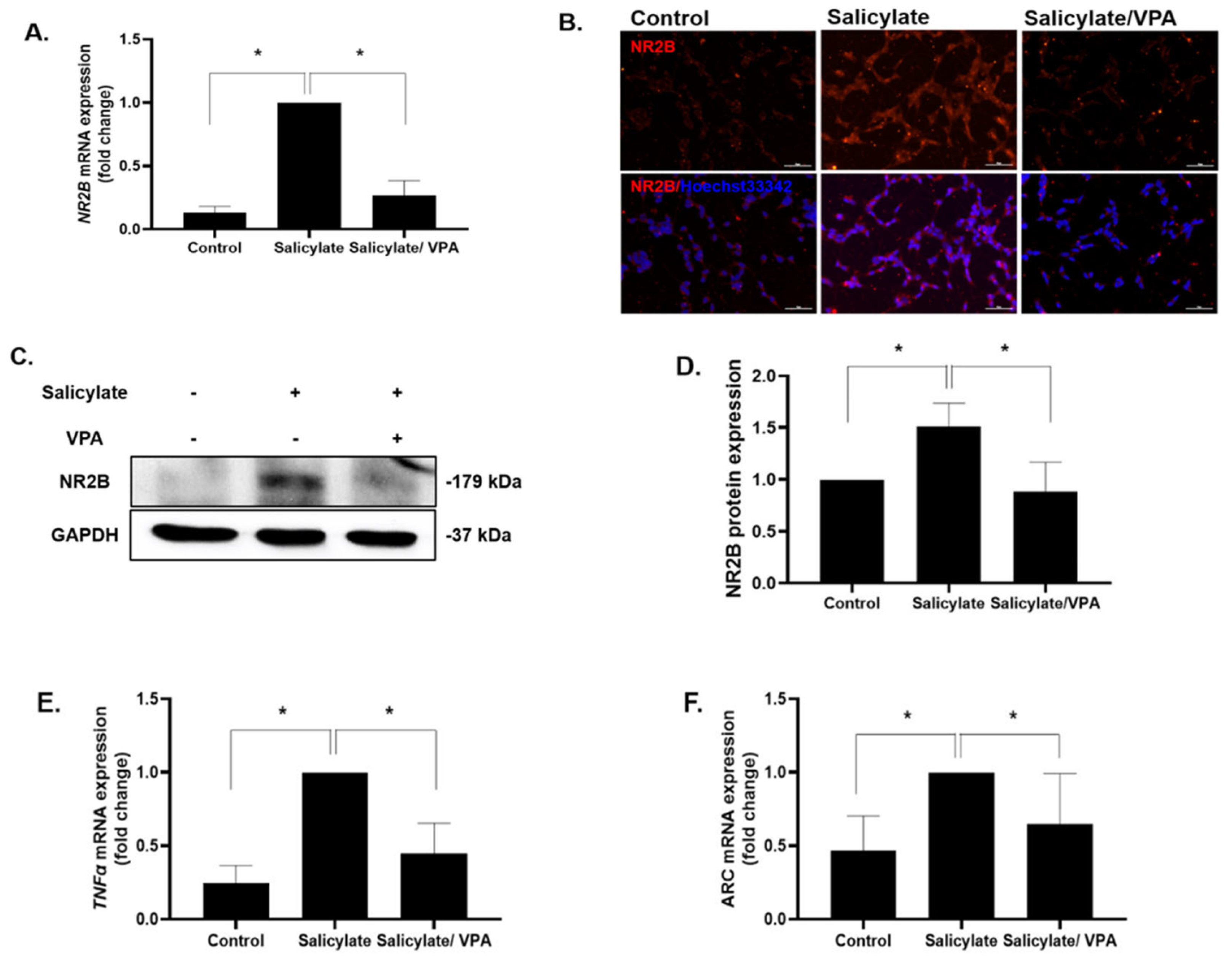
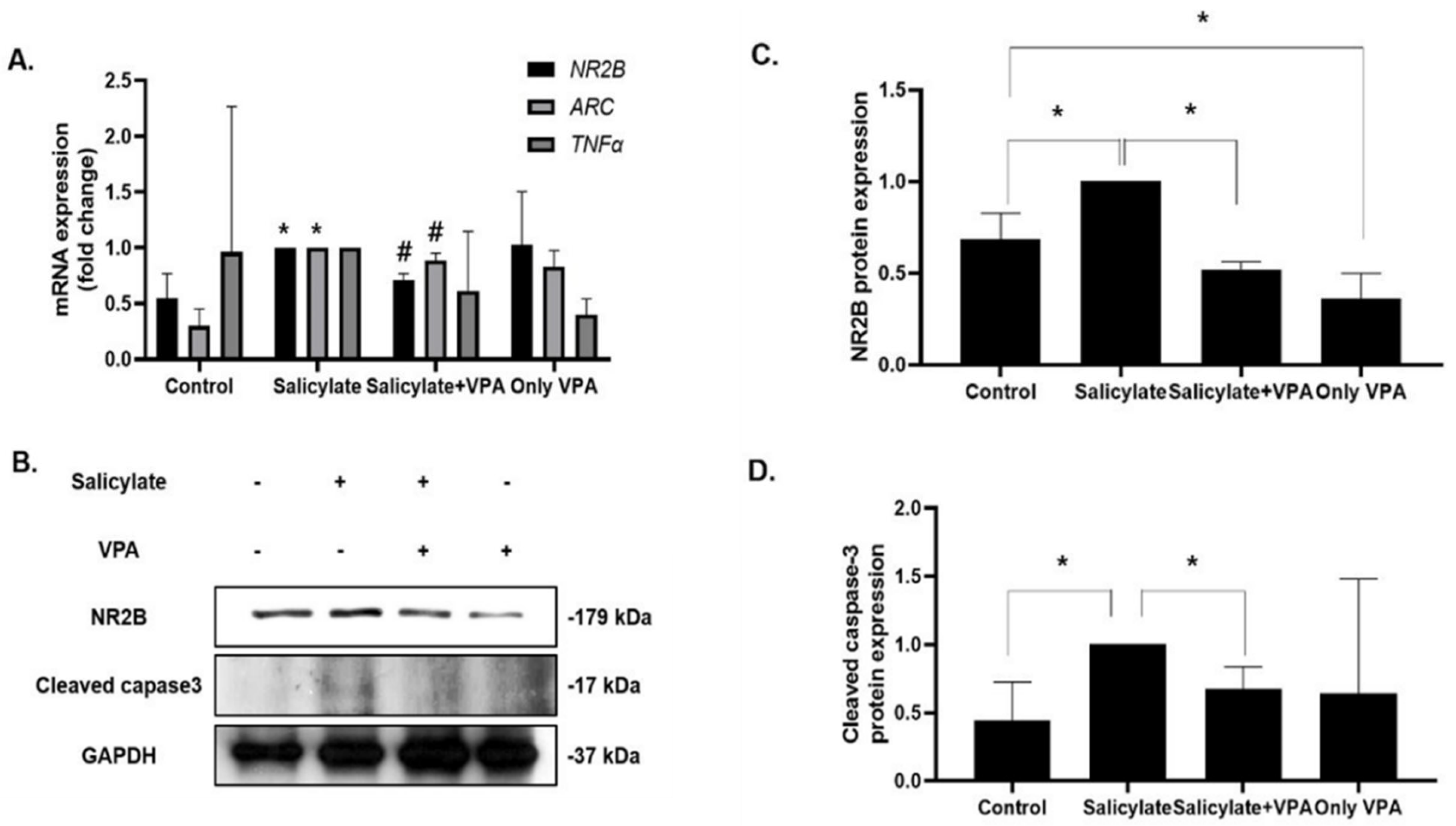
2.1. Gene Expression Levels of ARC, TNFα, and NR2B
2.2. Expression of p-CREB
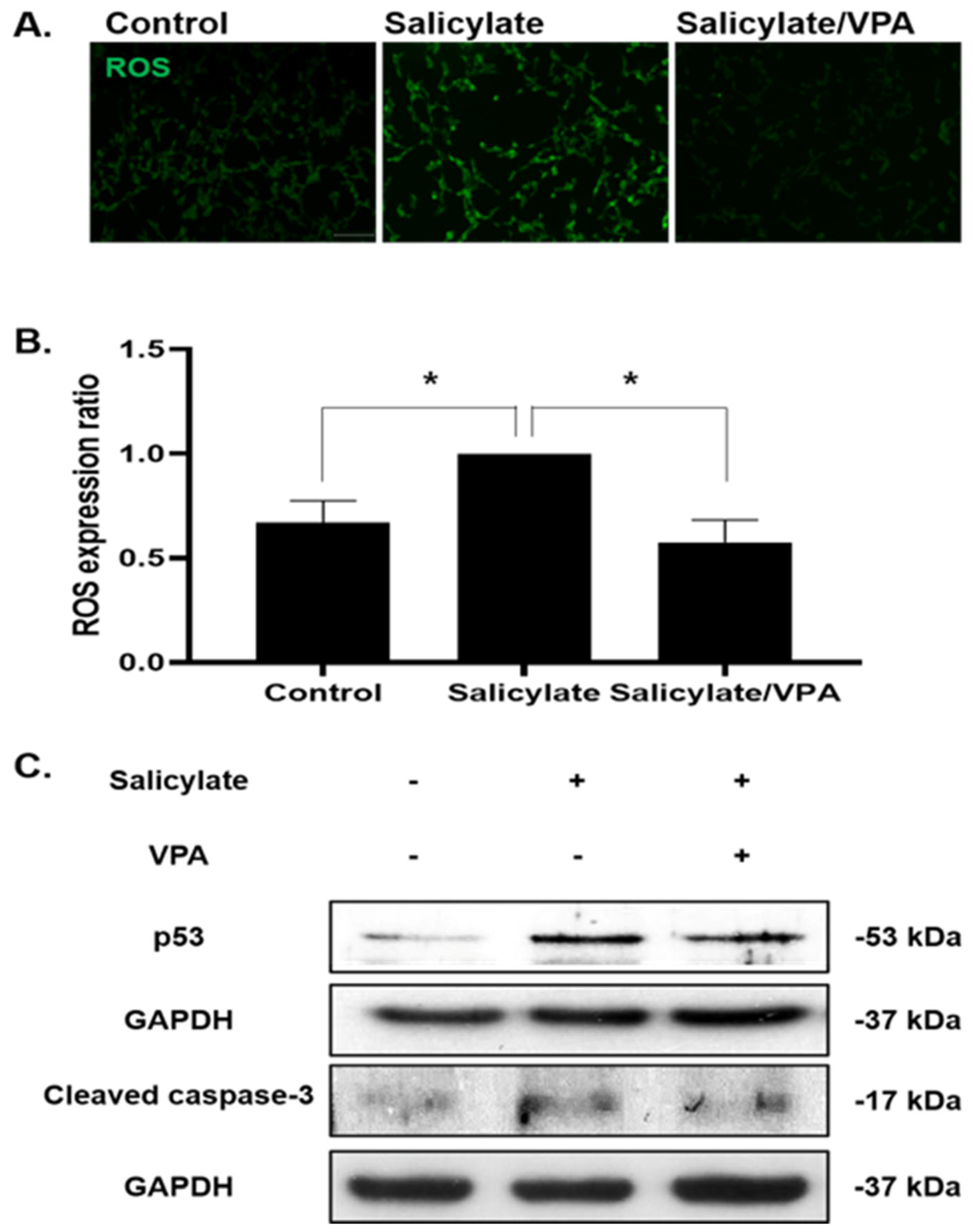
2.3. Neuroprotective Effect via Modulation of ROS Production
2.4. Effects of VPA in Rat Cortical Neurons
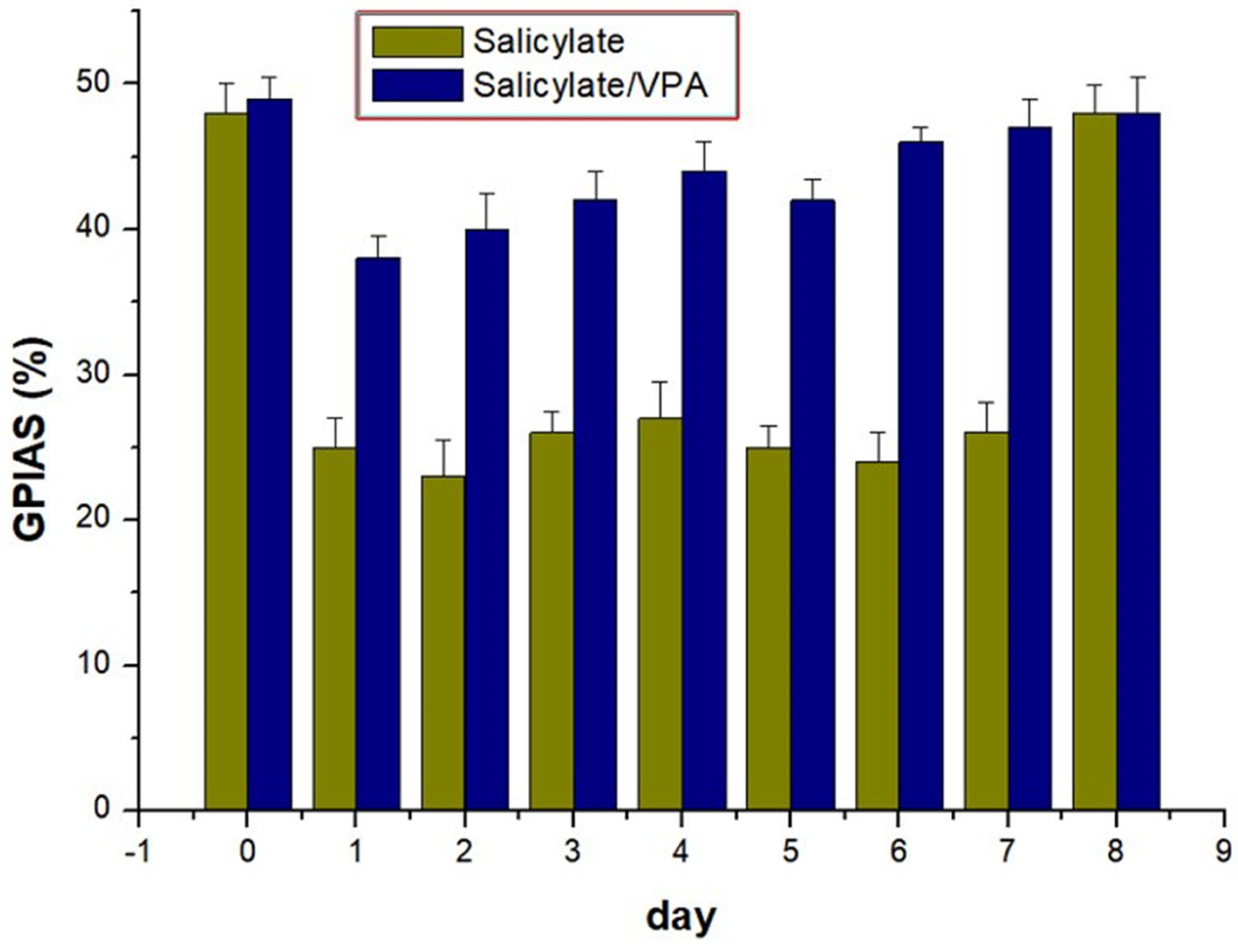
2.5. GPIAS
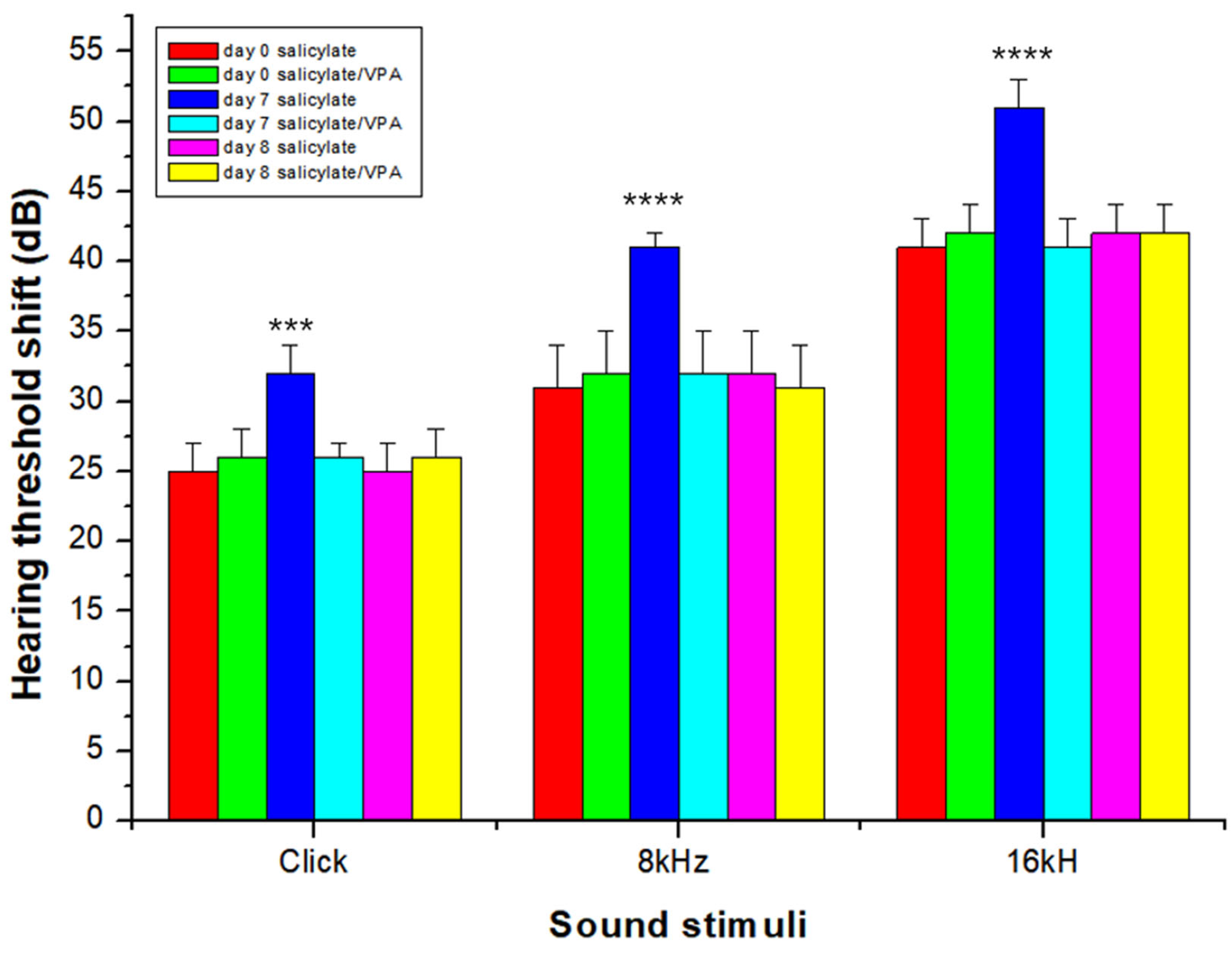
2.6. Auditory Brainstem Response
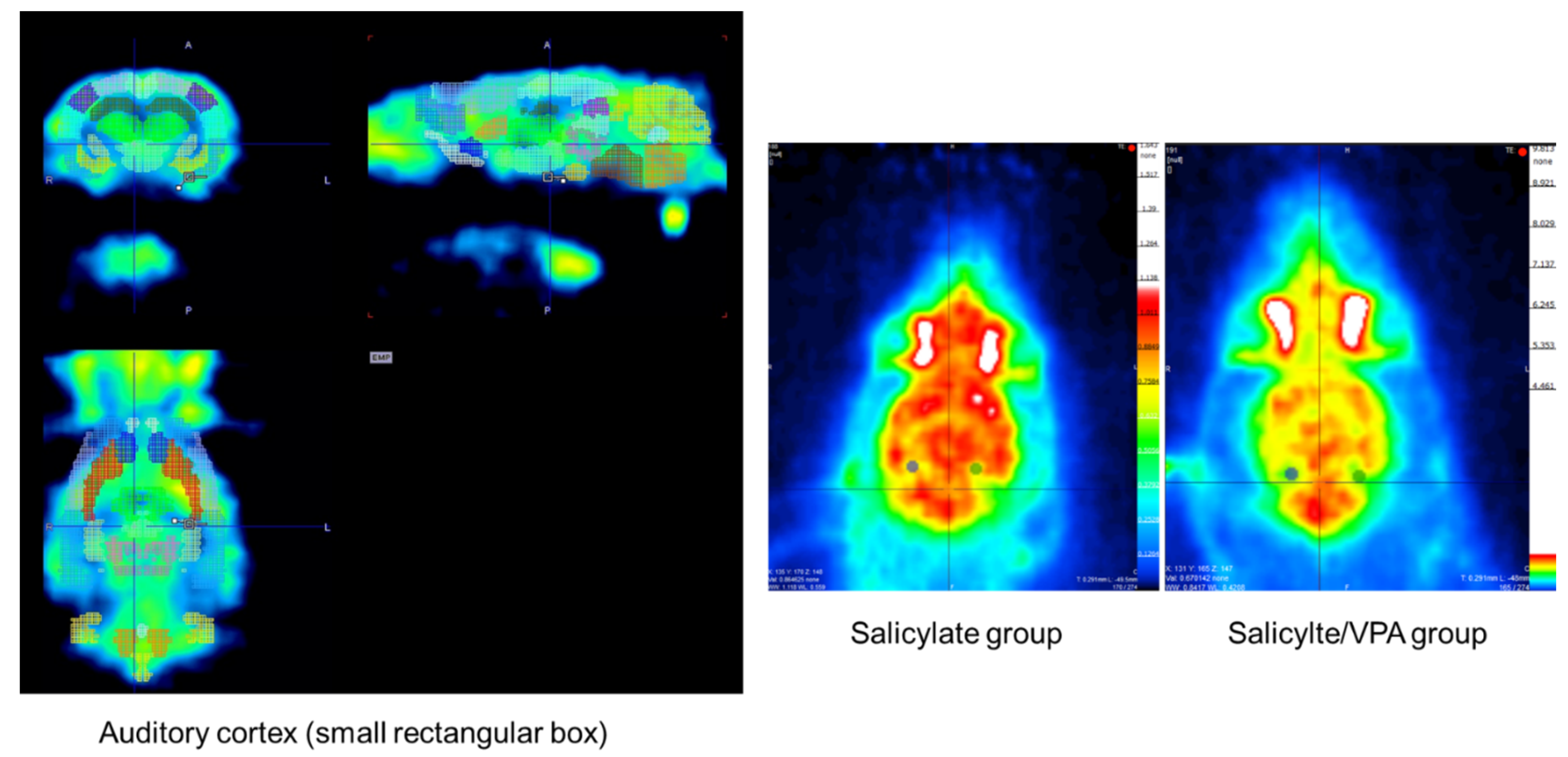
2.7. MicroPET CT Findings
2.8. Spatial Expression of NR2B in the Auditory Cortex after Salicylate and VPA Treatments
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Quantitative Polymerase Chain Reaction (qPCR)
4.3. Immunoblot Analysis
4.4. Immunocytochemical Staining
4.5. Detection of Intracellular ROS
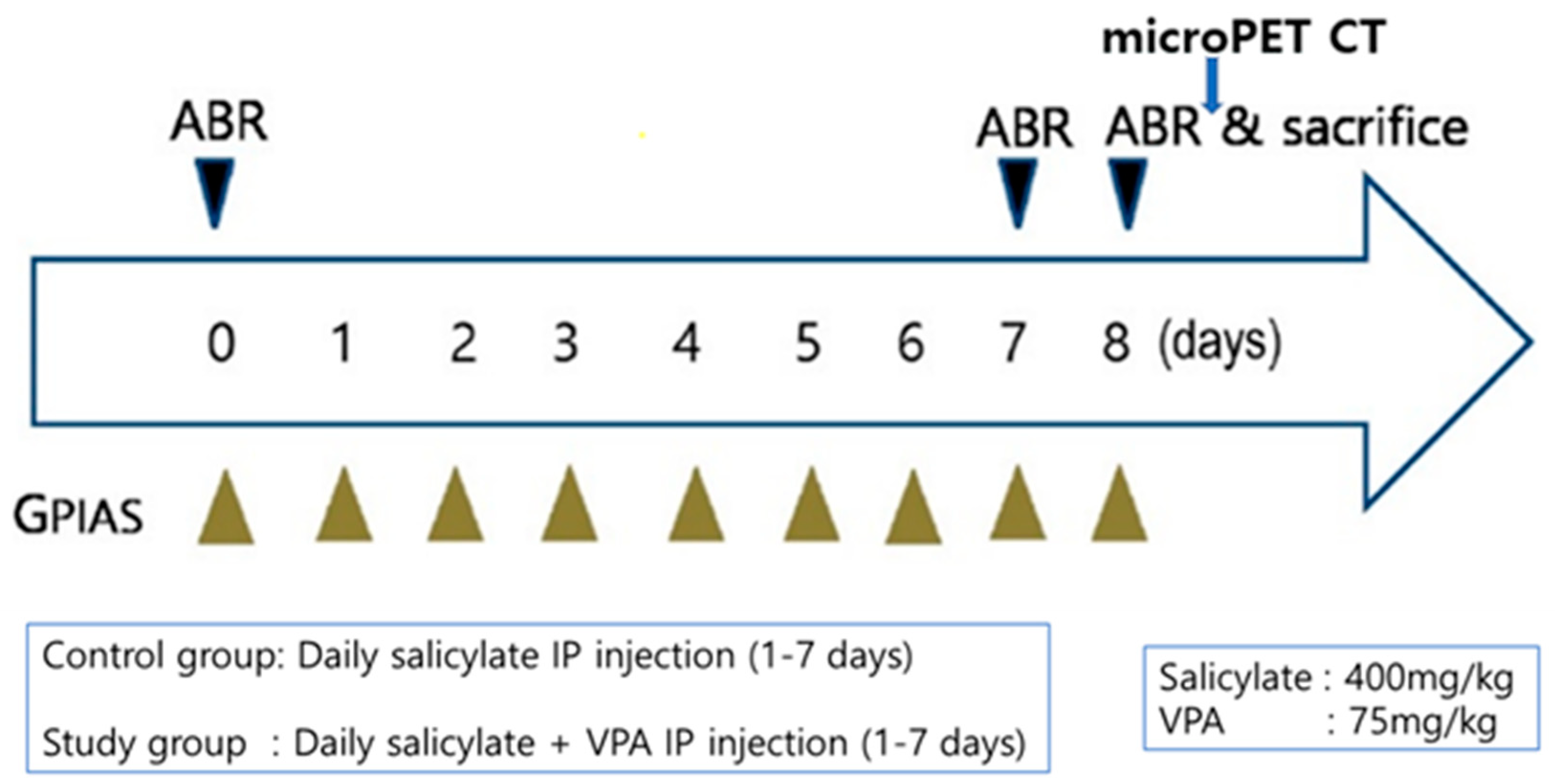
4.6. In Vivo Experiments
4.6.1. Animals
4.6.2. Gap and Noise Burst Prepulse Inhibition of Acoustic Startle
4.6.3. Measurement of Auditory Brainstem Response
4.6.4. 18F-FDG PET Imaging
4.6.5. Preparation of Free-Floating Sections
4.6.6. Immunohistochemistry of Free-Floating Sections
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nondahl, D.M.; Cruickshanks, K.J.; Huang, G.H.; Klein, B.E.; Klein, R.; Nieto, F.J.; Tweed, T.S. Tinnitus and its risk factors in the Beaver Dam offspring study. Int. J. Audiol. 2011, 50, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Baguley, D.; McFerran, D.; Hall, D. Tinnitus. Lancet 2013, 382, 1600–1607. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.; Seidman, M. Tinnitus in the older adult: Epidemiology, pathophysiology and treatment options. Drugs Aging 2004, 21, 297–305. [Google Scholar] [CrossRef]
- Ettinger, A.B.; Argoff, C.E. Use of antiepileptic drugs for nonepileptic conditions: Psychiatric disorders and chronic pain. Neurother. J. Am. Soc. Exp. NeuroTher. 2007, 4, 75–83. [Google Scholar] [CrossRef]
- Levine, R.A. Typewriter tinnitus: A carbamazepine-responsive syndrome related to auditory nerve vascular compression. ORL J. Otorhinolaryngol. Relat. Spec. 2006, 68, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Mardini, M.K. Ear-clicking “tinnitus” responding to carbamazepine. N. Engl. J. Med. 1987, 317, 1542. [Google Scholar] [CrossRef]
- Sunwoo, W.; Jeon, Y.J.; Bae, Y.J.; Jang, J.H.; Koo, J.W.; Song, J.J. Typewriter tinnitus revisited: The typical symptoms and the initial response to carbamazepine are the most reliable diagnostic clues. Sci. Rep. 2017, 7, 10615. [Google Scholar] [CrossRef] [Green Version]
- Menkes, D.B.; Larson, P.M. Sodium valproate for tinnitus. J. Neurol. Neurosurg. Psychiatry 1998, 65, 803. [Google Scholar] [CrossRef] [Green Version]
- Jang, C.H.; Lee, S.; Park, I.Y.; Song, A.; Moon, C.; Cho, G.W. Memantine Attenuates Salicylate-induced Tinnitus Possibly by Reducing NR2B Expression in Auditory Cortex of Rat. Exp. Neurobiol. 2019, 28, 495–503. [Google Scholar] [CrossRef]
- Jastreboff, P.J.; Sasaki, C.T. An animal model of tinnitus: A decade of development. Am. J. Otol. 1994, 15, 19–27. [Google Scholar] [PubMed]
- Zhao, J.; Wang, B.; Wang, X.; Shang, X. Up-regulation of Ca(2+)/CaMKII/CREB signaling in salicylate-induced tinnitus in rats. Mol. Cell. Biochem. 2018, 448, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Yi, B.; Wu, C.; Shi, R.; Han, K.; Sheng, H.; Li, B.; Mei, L.; Wang, X.; Huang, Z.; Wu, H. Long-term Administration of Salicylate-induced Changes in BDNF Expression and CREB Phosphorylation in the Auditory Cortex of Rats. Otol. Neurotol. 2018, 39, e173–e180. [Google Scholar] [CrossRef] [Green Version]
- Song, R.B.; Lou, W.H. Monosialotetrahexosylganglioside Inhibits the Expression of p-CREB and NR2B in the Auditory Cortex in Rats with Salicylate-Induced Tinnitus. Clin. Lab. 2015, 61, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Bosetti, F.; Weerasinghe, G.R.; Rosenberger, T.A.; Rapoport, S.I. Valproic acid down-regulates the conversion of arachidonic acid to eicosanoids via cyclooxygenase-1 and -2 in rat brain. J. Neurochem. 2003, 85, 690–696. [Google Scholar] [CrossRef]
- Iacobucci, G.J.; Popescu, G.K. Spatial Coupling Tunes NMDA Receptor Responses via Ca(2+) Diffusion. J. Neurosci. 2019, 39, 8831–8844. [Google Scholar] [CrossRef] [PubMed]
- Hernández, D.E.; Salvadores, N.A.; Moya-Alvarado, G.; Catalán, R.J.; Bronfman, F.C.; Court, F.A. Axonal degeneration induced by glutamate excitotoxicity is mediated by necroptosis. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Ding, D.; Su, J.; Manohar, S.; Salvi, R. Salicylate selectively kills cochlear spiral ganglion neurons by paradoxically up-regulating superoxide. Neurotox Res. 2013, 24, 307–319. [Google Scholar] [CrossRef]
- Chen, J.C.; Zhang, Y.; Jie, X.M.; She, J.; Dongye, G.Z.; Zhong, Y.; Deng, Y.Y.; Wang, J.; Guo, B.Y.; Chen, L.M. Ruthenium(II) salicylate complexes inducing ROS-mediated apoptosis by targeting thioredoxin reductase. J. Inorg. Biochem. 2019, 193, 112–123. [Google Scholar] [CrossRef]
- Feng, H.; Yin, S.H.; Tang, A.Z. Blocking caspase-3-dependent pathway preserves hair cells from salicylate-induced apoptosis in the guinea pig cochlea. Mol. Cell. Biochem. 2011, 353, 291–303. [Google Scholar] [CrossRef]
- Feng, H.; Yin, S.H.; Tang, A.Z.; Tan, S.H. Salicylate initiates apoptosis in the spiral ganglion neuron of guinea pig cochlea by activating caspase-3. Neurochem. Res. 2011, 36, 1108–1115. [Google Scholar] [CrossRef]
- Basselin, M.; Chang, L.; Chen, M.; Bell, J.M.; Rapoport, S.I. Chronic administration of valproic acid reduces brain NMDA signaling via arachidonic acid in unanesthetized rats. Neurochem. Res. 2008, 33, 2229–2240. [Google Scholar] [CrossRef] [Green Version]
- Bosetti, F.; Bell, J.M.; Manickam, P. Microarray analysis of rat brain gene expression after chronic administration of sodium valproate. Brain Res. Bull. 2005, 65, 331–338. [Google Scholar] [CrossRef]
- Niu, H.; Ding, S.; Li, H.; Wei, J.; Ren, C.; Wu, X.; Huma, T.; Zhang, Q. Effect of Long-Term Sodium Salicylate Administration on Learning, Memory, and Neurogenesis in the Rat Hippocampus. Biomed. Res. Int. 2018, 2018, 7807426. [Google Scholar] [CrossRef]
- Cui, W.; Wang, H.; Cheng, Y.; Ma, X.; Lei, Y.; Ruan, X.; Shi, L.; Lv, M. Long-term treatment with salicylate enables NMDA receptors and impairs AMPA receptors in C57BL/6J mice inner hair cell ribbon synapse. Mol. Med. Rep. 2019, 19, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.H.; Chen, J.C.; Chan, Y.C. Effects of C-phycocyanin and Spirulina on salicylate-induced tinnitus, expression of NMDA receptor and inflammatory genes. PLoS ONE 2013, 8, e58215. [Google Scholar] [CrossRef]
- Hu, S.S.; Mei, L.; Chen, J.Y.; Huang, Z.W.; Wu, H. Expression of immediate-early genes in the dorsal cochlear nucleus in salicylate-induced tinnitus. Eur. Arch. Otorhinolaryngol. 2016, 273, 325–332. [Google Scholar] [CrossRef]
- Qin, D.; Liu, P.; Chen, H.; Huang, X.; Ye, W.; Lin, X.; Wei, F.; Su, J. Salicylate-Induced Ototoxicity of Spiral Ganglion Neurons: Ca(2+)/CaMKII-Mediated Interaction Between NMDA Receptor and GABA(A) Receptor. Neurotox Res. 2019, 35, 838–847. [Google Scholar] [CrossRef]
- Amidfar, M.; de Oliveira, J.; Kucharska, E.; Budni, J.; Kim, Y.K. The role of CREB and BDNF in neurobiology and treatment of Alzheimer’s disease. Life Sci. 2020, 257, 118020. [Google Scholar] [CrossRef] [PubMed]
- Nanau, R.M.; Neuman, M.G. Adverse drug reactions induced by valproic acid. Clin. Biochem. 2013, 46, 1323–1338. [Google Scholar] [CrossRef]
- Bing, D.; Lee, S.C.; Campanelli, D.; Xiong, H.; Matsumoto, M.; Panford-Walsh, R.; Wolpert, S.; Praetorius, M.; Zimmermann, U.; Chu, H. Cochlear NMDA receptors as a therapeutic target of noise-induced tinnitus. Cell. Physiol. Biochem. 2015, 35, 1905–1923. [Google Scholar] [CrossRef]
- Li, M.; Zhao, Y.; Zhan, Y.; Yang, L.; Feng, X.; Lu, Y.; Lei, J.; Zhao, T.; Wang, L.; Zhao, H. Enhanced white matter reorganization and activated brain glucose metabolism by enriched environment following ischemic stroke: Micro PET/CT and MRI study. Neuropharmacology 2020, 176, 108202. [Google Scholar] [CrossRef] [PubMed]
- Jang, D.-P.; Lee, S.-H.; Lee, S.-Y.; Park, C.-W.; Cho, Z.-H.; Kim, Y.-B. Neural responses of rats in the forced swimming test: [F-18] FDG micro PET study. Behav. Brain Res. 2009, 203, 43–47. [Google Scholar] [CrossRef]
- Li, Y.-Y.; Zhang, B.; Yu, K.-W.; Li, C.; Xie, H.-Y.; Bao, W.-Q.; Kong, Y.-Y.; Jiao, F.-Y.; Guan, Y.-H.; Bai, Y.-L. Effects of constraint-induced movement therapy on brain glucose metabolism in a rat model of cerebral ischemia: A micro PET/CT study. Int. J. Neurosci. 2018, 128, 736–745. [Google Scholar] [CrossRef]
- Lu, J.; Lobarinas, E.; Deng, A.; Goodey, R.; Stolzberg, D.; Salvi, R.J.; Sun, W. GABAergic neural activity involved in salicylate-induced auditory cortex gain enhancement. Neuroscience 2011, 189, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Bauer, C.A.; Brozoski, T.J.; Holder, T.M.; Caspary, D.M. Effects of chronic salicylate on GABAergic activity in rat inferior colliculus. Hear. Res. 2000, 147, 175–182. [Google Scholar] [CrossRef]
- Liu, J.; Li, X.; Wang, L.; Dong, Y.; Han, H.; Liu, G. Effects of salicylate on serotoninergic activities in rat inferior colliculus and auditory cortex. Hear. Res. 2003, 175, 45–53. [Google Scholar] [CrossRef]
- Wang, H.T.; Luo, B.; Huang, Y.N.; Zhou, K.Q.; Chen, L. Sodium salicylate suppresses serotonin-induced enhancement of GABAergic spontaneous inhibitory postsynaptic currents in rat inferior colliculus in vitro. Hear. Res. 2008, 236, 42–51. [Google Scholar] [CrossRef]
- Sheppard, A.; Hayes, S.H.; Chen, G.D.; Ralli, M.; Salvi, R. Review of salicylate-induced hearing loss, neurotoxicity, tinnitus and neuropathophysiology. Acta Otorhinolaryngol. Ital. 2014, 34, 79–93. [Google Scholar] [PubMed]
- Shahsavarani, S.; Khan, R.A.; Husain, F.T. Tinnitus and the brain: A review of functional and anatomical magnetic resonance imaging studies. Perspect. ASHA Spec. Interest Groups 2019, 4, 896–909. [Google Scholar] [CrossRef]
- Bojić, T.; Perović, V.R.; Senćanski, M.; Glišić, S. Identification of candidate allosteric modulators of the m1 muscarinic acetylcholine receptor which may improve vagus nerve stimulation in chronic tinnitus. Front. Neurosci. 2017, 11, 636. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, A.; Cho, G.-W.; Vijayakumar, K.A.; Moon, C.; Ang, M.J.; Kim, J.; Park, I.; Jang, C.H. Neuroprotective Effect of Valproic Acid on Salicylate-Induced Tinnitus. Int. J. Mol. Sci. 2022, 23, 23. https://doi.org/10.3390/ijms23010023
Song A, Cho G-W, Vijayakumar KA, Moon C, Ang MJ, Kim J, Park I, Jang CH. Neuroprotective Effect of Valproic Acid on Salicylate-Induced Tinnitus. International Journal of Molecular Sciences. 2022; 23(1):23. https://doi.org/10.3390/ijms23010023
Chicago/Turabian StyleSong, Anji, Gwang-Won Cho, Karthikeyan A. Vijayakumar, Changjong Moon, Mary Jasmin Ang, Jahae Kim, Ilyong Park, and Chul Ho Jang. 2022. "Neuroprotective Effect of Valproic Acid on Salicylate-Induced Tinnitus" International Journal of Molecular Sciences 23, no. 1: 23. https://doi.org/10.3390/ijms23010023
APA StyleSong, A., Cho, G.-W., Vijayakumar, K. A., Moon, C., Ang, M. J., Kim, J., Park, I., & Jang, C. H. (2022). Neuroprotective Effect of Valproic Acid on Salicylate-Induced Tinnitus. International Journal of Molecular Sciences, 23(1), 23. https://doi.org/10.3390/ijms23010023