
Genetic and Epigenomic Modifiers of Diabetic Neuropathy

 , ,
, ,
Abstract
1. Introduction
2. Anatomical, Histological and Clinical Considerations
3. Peripheral Nervous System Dysfunction in Diabetic Neuropathy
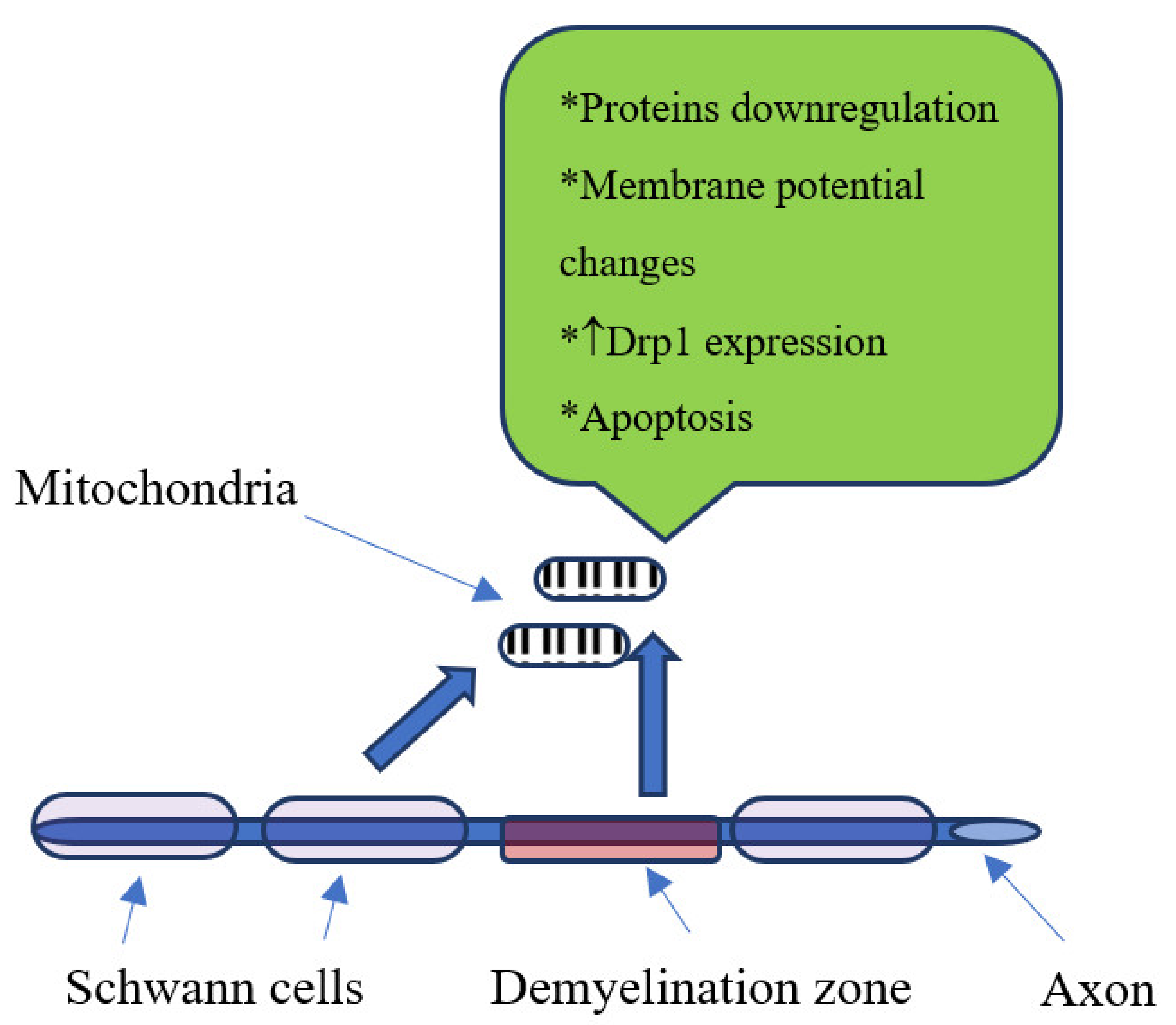
4. Biochemical Changes in Diabetic Neuropathy
5. Inflammation, Cytokines and Neurotrophins in Diabetic Neuropathy
6. The Role of Dyslipidemia in Diabetic Neuropathy
7. Genetic Basis of Polyol Pathway
8. Other Genetic Risk Factors for Diabetic Neuropathy
9. Epigenomic Modifications in Diabetic Neuropathy

{kind=link}
{kind=link}
{kind=link}
| Non-Coding RNAs | Potential Mechanism in PDN | Model System | References |
|---|---|---|---|
| MiRNA | |||
| miR199a3p | Increased expression, Serpin E2 downregulation | DM Patients (peripheral blood) | [91] |
| mir9 | Increased expression, CALHM1 upregulated | PDN rat model (spinal dorsal horn) | [92] |
| miR25 | Decreased expression, regulation of oxidative/nitrative stress | Diabetic mouse (sciatic nerves) | [93] |
| miR146 | Decreased expression, negatively correlated with the levels of IL-1β, TNF-α and NF-κB | T2DM and PDN rat | [94,95] |
| miR190a5p | Decreased expression, SLC17A6 upregulated | Diabetic mouse | [96] |
| LncRNA | |||
| 48+ | Increased expression, P2X3 Receptor upregulated | Diabetic rat (dorsal root ganglia) | [102] |
| Increased expression, P2X7 Receptor upregulated | Diabetic rat (superior cervical ganglia) | [107] | |
| NONRATT021972 | Increased expression, neuropathic pain higher scores/TNF-a upregulated | DM2 Patients (peripheral blood)/diabetic rat | [103] |
| Increased expression, P2X7 Receptor upregulated | PC12 Cells | [106] | |
| Diabetic rat (dorsal root ganglia) | [108] | ||
| BC168687 | Increased expression, TRPV1 Receptor upregulated Increased expression, P2X7 Receptor upregulated | Diabetic rat (dorsal root ganglia) | [104,109] |
| CCNT2-AS1 | Neurotrophin-MAPK signaling pathway affecting | DM/PDN patients (peripheral blood) | [110] |
| RP1-249H1.2 | |||
| CTD-3239E11.2 | |||
| RP11-51B23.3 | |||
| STAM-AS1 | |||
| LINC00629 | |||
| MALAT1 (NEAT2) | Increased expression in DM and PDN patients, affecting multiple physiological processes | ||
| MIR143HG | Decreased expression in PDN patients | ||
| PVT1 | |||
| H19 | Decreased expression in PDN patients VDAC1 downregulation in diabetic cardiomyopathy | DM/PDN patients (peripheral blood) Cultured neonatal rat cardiomyocytes | [110,113] |
10. Potential Treatment Targets
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Witzel, I.-I.; Jelinek, H.F.; Khalaf, K.; Lee, S.; Khandoker, A.H.; Alsafar, H. Identifying Common Genetic Risk Factors of Diabetic Neuropathies. Front. Endocrinol. (Lausanne) 2015, 6, 88. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, R.; Wang, D.; Liu, X. Genetics of Diabetic Neuropathy: Systematic Review, Meta-Analysis and Trial Sequential Analysis. Ann. Clin. Transl. Neurol. 2019, 6, 1996–2013. [Google Scholar] [CrossRef]
- Kallinikou, D.; Soldatou, A.; Tsentidis, C.; Louraki, M.; Kanaka-Gantenbein, C.; Kanavakis, E.; Karavanaki, K. Diabetic Neuropathy in Children and Adolescents with Type 1 Diabetes Mellitus: Diagnosis, Pathogenesis, and Associated Genetic Markers. Diabetes. Metab. Res. Rev. 2019, 35, e3178. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Han, Y.; Hu, Q.; Zhang, X.; Cui, G.; Li, Z.; Guan, Y. Effects of Common Polymorphisms in the MTHFR and ACE Genes on Diabetic Peripheral Neuropathy Progression: A Meta-Analysis. Mol. Neurobiol. 2017, 54, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Verrotti, A.; Giuva, P.T.; Morgese, G.; Chiarelli, F. New trends in the etiopathogenesis of diabetic peripheral neuropathy. J. Child. Neurol. 2001, 16, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Novakovic, I.; Maksimovic, N.; Pavlovic, A.; Zarkovic, M.; Rovcanin, B.; Mirkovic, D.; Pekmezovic, T.; Cvetkovic, D. Introduction to Molecular Genetic Diagnostics. J. Med. Biochem. 2014, 33, 3–7. [Google Scholar] [CrossRef][Green Version]
- Wilson, N.M.; Wright, D.E. Inflammatory Mediators in Diabetic Neuropathy. J. Diabetes Metab. 2011, S5, 4. [Google Scholar] [CrossRef]
- Vinik, A.I.; Nevoret, M.L.; Casellini, C.; Parson, H. Diabetic neuropathy. Endocrinol. Metab. Clin. N. Am. 2013, 42, 747–787. [Google Scholar] [CrossRef]
- Groener, J.B.; Jende, J.M.E.; Kurz, F.T.; Kender, Z.; Treede, R.D.; Schuh-Hofer, S.; Nawroth, P.P.; Bendszus, M.; Kopf, S. Understanding Diabetic Neuropathy-From Subclinical Nerve Lesions to Severe Nerve Fiber Deficits: A Cross-Sectional Study in Patients With Type 2 Diabetes and Healthy Control Subjects. Diabetes 2020, 69, 436–447. [Google Scholar] [CrossRef]
- Galosi, E.; Di Pietro, G.; La Cesa, S.; Di Stefano, G.; Leone, C.; Fasolino, A.; Di Lionardo, A.; Leonetti, F.; Buzzetti, R.; Mollica, C.; et al. Differential involvement of myelinated and unmyelinated nerve fibers in painful diabetic polyneuropathy. Muscle Nerve 2021, 63, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L. Small Fiber Neuropathy. Semin. Neurol. 2019, 39, 570–577. [Google Scholar] [CrossRef]
- Vinik, A.I.; Park, T.S.; Stansberry, K.B.; Pittenger, G.L. Diabetic neuropathies. Diabetologia 2000, 43, 957–973. [Google Scholar] [CrossRef] [PubMed]
- Freeman, R. Diabetic autonomic neuropathy. Handb. Clin. Neurol. 2014, 126, 63–79. [Google Scholar]
- Azmi, S.; Petropoulos, I.N.; Ferdousi, M.; Ponirakis, G.; Alam, U.; Malik, R.A. An update on the diagnosis and treatment of diabetic somatic and autonomic neuropathy. F1000Research 2019, 8, 186. [Google Scholar] [CrossRef]
- Gatopoulou, A.; Papanas, N.; Maltezos, E. Diabetic gastrointestinal autonomic neuropathy: Current status and new achievements for everyday clinical practice. Eur. J. Intern. Med. 2012, 23, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Bissinger, A. Cardiac Autonomic Neuropathy: Why Should Cardiologists Care about That? J. Diabetes Res. 2017, 2017, 5374176. [Google Scholar] [CrossRef] [PubMed]
- Hecht, M.J.; Neundörfer, B.; Kiesewetter, F.; Hilz, M.J. Neuropathy is a major contributing factor to diabetic erectile dysfunction. Neurol. Res. 2001, 23, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Agochukwu-Mmonu, N.; Pop-Busui, R.; Wessells, H.; Sarma, A.V. Autonomic neuropathy and urologic complications in diabetes. Auton. Neurosci. 2020, 229, 102736. [Google Scholar] [CrossRef]
- Nielsen, F.S.; Hansen, H.P.; Jacobsen, P.; Rossing, P.; Smidt, U.M.; Christensen, N.J.; Pevet, P.; Vivien-Roels, B.; Parving, H.H. Increased sympathetic activity during sleep and nocturnal hypertension in Type 2 diabetic patients with diabetic nephropathy. Diabet. Med. 1999, 16, 555–562. [Google Scholar] [CrossRef]
- Gandhi, P.G.; Gundu, H.R. Detection of neuropathy using a sudomotor test in type 2 diabetes. Degener. Neurol. Neuromuscul. Dis. 2015, 5, 1–7. [Google Scholar] [PubMed]
- Schmidt, R.E. Neuropathology and pathogenesis of diabetic autonomic neuropathy. Int. Rev. Neurobiol. 2002, 50, 257–292. [Google Scholar]
- Naruse, K. Schwann Cells as Crucial Players in Diabetic Neuropathy. Adv. Exp. Med. Biol. 2019, 1190, 345–356. [Google Scholar]
- De Gregorio, C.; Contador, D.; Campero, M.; Ezquer, M.; Ezquer, F. Characterization of diabetic neuropathy progression in a mouse model of type 2 diabetes mellitus. Biol. Open. 2018, 7, bio036830. [Google Scholar] [CrossRef] [PubMed]
- Younger, D.S. Diabetic neuropathy: A clinical and neuropathological study of 107 patients. Neurol. Res. Int. 2010, 2010, 140379. [Google Scholar] [CrossRef] [PubMed]
- Zenker, J.; Ziegler, D.; Chrast, R. Novel pathogenic pathways in diabetic neuropathy. Trends Neurosci. 2013, 36, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Mizisin, A.P. Mechanisms of diabetic neuropathy: Schwann cells. Handb. Clin. Neurol. 2014, 126, 401–428. [Google Scholar] [PubMed]
- Robertson, D.M.; Sima, A.A. Diabetic Neuropathy in the Mutant Mouse [C57BL/Ks(Db/Db)]: A Morphometric Study. Diabetes 1980, 29, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, N.P.; Vægter, C.B.; Andersen, H.; Østergaard, L.; Calcutt, N.A.; Jensen, T.S. Schwann cell interactions with axons and microvessels in diabetic neuropathy. Nat. Rev. Neurol. 2017, 13, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Akude, E.; Zherebitskaya, E.; Chowdhury, S.K.R.; Smith, D.R.; Dobrowsky, R.T.; Fernyhough, P. Diminished Superoxide Generation Is Associated with Respiratory Chain Dysfunction and Changes in the Mitochondrial Proteome of Sensory Neurons from Diabetic Rats. Diabetes 2011, 60, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-J.; Price, S.A.; Chilton, L.; Calcutt, N.A.; Tomlinson, D.R.; Verkhratsky, A.; Fernyhough, P. Insulin Prevents Depolarization of the Mitochondrial Inner Membrane in Sensory Neurons of Type 1 Diabetic Rats in the Presence of Sustained Hyperglycemia. Diabetes 2003, 52, 2129–2136. [Google Scholar] [CrossRef] [PubMed]
- Roy Chowdhury, S.K.; Smith, D.R.; Saleh, A.; Schapansky, J.; Marquez, A.; Gomes, S.; Akude, E.; Morrow, D.; Calcutt, N.A.; Fernyhough, P. Impaired adenosine monophosphate-activated protein kinase signalling in dorsal root ganglia neurons is linked to mitochondrial dysfunction and peripheral neuropathy in diabetes. Brain 2012, 135 Pt 6, 1751–1766. [Google Scholar] [CrossRef]
- Sifuentes-Franco, S.; Pacheco-Moisés, F.P.; Rodríguez-Carrizalez, A.D.; Miranda-Díaz, A.G. The Role of Oxidative Stress, Mitochondrial Function, and Autophagy in Diabetic Polyneuropathy. J. Diabetes Res. 2017, 2017, 1673081. [Google Scholar] [CrossRef]
- Román-Pintos, L.M.; Villegas-Rivera, G.; Rodríguez-Carrizalez, A.D.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. Diabetic Polyneuropathy in Type 2 Diabetes Mellitus: Inflammation, Oxidative Stress, and Mitochondrial Function. J. Diabetes. Res. 2016, 2016, 3425617. [Google Scholar] [CrossRef] [PubMed]
- Verge, V.M.K.; Andreassen, C.S.; Arnason, T.G.; Andersen, H. Mechanisms of Disease: Role of Neurotrophins in Diabetes and Diabetic Neuropathy. Handb. Clin. Neurol. 2014, 126, 443–460. [Google Scholar]
- Singh, R.; Kishore, L.; Kaur, N. Diabetic Peripheral Neuropathy: Current Perspective and Future Directions. Pharmacol. Res. 2014, 80, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Dewanjee, S.; Das, S.; Das, A.K.; Bhattacharjee, N.; Dihingia, A.; Dua, T.K.; Kalita, J.; Manna, P. Molecular Mechanism of Diabetic Neuropathy and Its Pharmacotherapeutic Targets. Eur. J. Pharmacol. 2018, 833, 472–523. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Kato, N.; Makino, M.; Suzuki, T.; Shindo, M. Continuous Inhibition of Excessive Polyol Pathway Flux in Peripheral Nerves by Aldose Reductase Inhibitor Fidarestat Leads to Improvement of Diabetic Neuropathy. J. Diabetes Complicat. 1999, 13, 141–150. [Google Scholar] [CrossRef]
- Vincent, A.M.; Russell, J.W.; Low, P.; Feldman, E.L. Oxidative Stress in the Pathogenesis of Diabetic Neuropathy. Endocr. Rev. 2004, 25, 612–628. [Google Scholar] [CrossRef]
- Buyukaydin, B.; Guler, E.M.; Karaaslan, T.; Olgac, A.; Zorlu, M.; Kiskac, M.; Kocyigit, A. Relationship between Diabetic Polyneuropathy, Serum Visfatin, and Oxidative Stress Biomarkers. World J. Diabetes 2020, 11, 309–321. [Google Scholar] [CrossRef]
- Pop-Busui, R.; Sima, A.; Stevens, M. Diabetic Neuropathy and Oxidative Stress. Diabetes. Metab. Res. Rev. 2006, 22, 257–273. [Google Scholar] [CrossRef]
- Jin, H.Y.; Park, T.S. Role of Inflammatory Biomarkers in Diabetic Peripheral Neuropathy. J. Diabetes Investig. 2018, 9, 1016–1018. [Google Scholar] [CrossRef] [PubMed]
- Doupis, J.; Lyons, T.E.; Wu, S.; Gnardellis, C.; Dinh, T.; Veves, A. Microvascular Reactivity and Inflammatory Cytokines in Painful and Painless Peripheral Diabetic Neuropathy. J. Clin. Endocrinol. Metab. 2009, 94, 2157–2163. [Google Scholar] [CrossRef]
- Bönhof, G.J.; Herder, C.; Strom, A.; Papanas, N.; Roden, M.; Ziegler, D. Emerging Biomarkers, Tools, and Treatments for Diabetic Polyneuropathy. Endocr. Rev. 2019, 40, 153–192. [Google Scholar] [CrossRef]
- Nikolic, D.; Jankovic, M.; Petrovic, B.; Novakovic, I. Genetic Aspects of Inflammation and Immune Response in Stroke. Int. J. Mol. Sci. 2020, 21, 7409. [Google Scholar] [CrossRef] [PubMed]
- Bonini, S.; Rasi, G.; Bracci-Laudiero, M.L.; Procoli, A.; Aloe, L. Nerve Growth Factor: Neurotrophin or Cytokine? Int. Arch. Allergy Immunol. 2003, 131, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Stino, A.M.; Rumora, A.E.; Kim, B.; Feldman, E.L. Evolving concepts on the role of dyslipidemia, bioenergetics, and inflammation in the pathogenesis and treatment of diabetic peripheral neuropathy. J. Peripher. Nerv. Syst. 2020, 25, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.M.; Callaghan, B.C.; Smith, A.L.; Feldman, E.L. Diabetic neuropathy: Cellular mechanisms as therapeutic targets. Nat. Rev. Neurol. 2011, 7, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Feldman, E.L.; Callaghan, B.C.; Pop-Busui, R.; Zochodne, D.W.; Wright, D.E.; Bennett, D.L.; Bril, V.; Russel, J.W.; Viswanathan, V. Diabetic neuropathy. Nat. Rev. Dis. Primers 2019, 5, 41. [Google Scholar] [CrossRef]
- Cai, Z.; Yang, Y.; Zhang, J. A systematic review and meta-analysis of the serum lipid profile in prediction of diabetic neuropathy. Sci. Rep. 2021, 11, 499. [Google Scholar] [CrossRef]
- Samadi, A.; Gurlek, A.; Sendur, S.N.; Karahan, S.; Akbiyik, F.; Lay, I. Oxysterol species: Reliable markers of oxidative stress in diabetes mellitus. J. Endocrinol. Investig. 2019, 42, 7–17. [Google Scholar] [CrossRef]
- Mutemberezi, V.; Guillemot-Legris, O.; Muccioli, G.G. Oxysterols: From cholesterol metabolites to key mediators. Prog. Lipid. Res. 2016, 64, 152–169. [Google Scholar] [CrossRef] [PubMed]
- Ferderbar, S.; Pereira, E.C.; Apolinário, E.; Bertolami, M.C.; Faludi, A.; Monte, O.; Calliari, L.E.; Sales, J.E.; Gagliardi, A.R.; Xavier, H.T.; et al. Cholesterol oxides as biomarkers of oxidative stress in type 1 and type 2 diabetes mellitus. Diabetes Metab. Res. Rev. 2007, 23, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.S.M.; Chung, S.K. Genetic Analysis of Aldose Reductase in Diabetic Complications. Curr. Med. Chem. 2003, 10, 1375–1387. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, S.; Mei, Z.; Zhao, F.; Xiang, Y. Polymorphisms in Sorbitol-Aldose Reductase (Polyol) Pathway Genes and Their Influence on Risk of Diabetic Retinopathy among Han Chinese. Med. Sci. Monit. 2019, 25, 7073–7078. [Google Scholar] [CrossRef] [PubMed]
- Sivenius, K.; Pihlajamäki, J.; Partanen, J.; Niskanen, L.; Laakso, M.; Uusitupa, M. Aldose Reductase Gene Polymorphisms and Peripheral Nerve Function in Patients with Type 2 Diabetes. Diabetes Care 2004, 27, 2021–2026. [Google Scholar] [CrossRef]
- Heesom, A.E.; Millward, A.; Demaine, A.G. Susceptibility to Diabetic Neuropathy in Patients with Insulin Dependent Diabetes Mellitus Is Associated with a Polymorphism at the 5′ End of the Aldose Reductase Gene. J. Neurol. Neurosurg. Psychiatry 1998, 64, 213–216. [Google Scholar] [CrossRef]
- Boulton, A.J.M.; Malik, R.A.; Arezzo, J.C.; Sosenko, J.M. Diabetic Somatic Neuropathies. Diabetes Care 2004, 27, 1458–1486. [Google Scholar] [CrossRef]
- Vinik, A.; Maser, R.; Nakave, A. Diabetic cardiovascular autonomic nerve dysfunction. US Endocr. Dis. 2007, 2, 66–74. [Google Scholar] [CrossRef]
- Forbes, J.M.; Cooper, M.E. Mechanisms of Diabetic Complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef]
- Li, Y.; Tong, N. Angiotensin-converting enzyme I/D polymorphism and diabetic peripheral neuropathy in type 2 diabetes mellitus: A meta-analysis. J. Renin Angiotensin. Aldosterone Syst. 2015, 16, 787–792. [Google Scholar] [CrossRef]
- Rahimi, Z. ACE insertion/deletion (I/D) polymorphism and diabetic nephropathy. J. Nephropathol. 2012, 1, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Fan, D.; Zhang, Y. Angiogenin gene polymorphism: A risk factor for diabetic peripheral neuropathy in the northern Chinese Han population. Neural. Regen. Res. 2013, 8, 3434–3440. [Google Scholar]
- Mooyaart, A.L.; Valk, E.J.; van Es, L.A.; Bruijn, J.A.; de Heer, E.; Freedman, B.I.; Dekkers, O.M.; Baelde, H.J. Genetic associations in diabetic nephropathy: A meta-analysis. Diabetologia 2011, 54, 544–553. [Google Scholar] [CrossRef]
- Bedlack, R.S.; Edelman, D.; Gibbs, J.W., 3rd; Kelling, D.; Strittmatter, W.; Saunders, A.M.; Morgenlander, J. APOE genotype is a risk factor for neuropathy severity in diabetic patients. Neurology 2003, 60, 1022–1024. [Google Scholar] [CrossRef] [PubMed]
- Monastiriotis, C.; Papanas, N.; Veletza, S.; Maltezos, E. APOE gene polymorphisms and diabetic peripheral neuropathy. Arch. Med. Sci. 2012, 8, 583–588. [Google Scholar] [CrossRef]
- Yigit, S.; Karakus, N.; Inanir, A. Association of MTHFR gene C677T mutation with diabetic peripheral neuropathy and diabetic retinopathy. Mol. Vis. 2013, 19, 1626–1630. [Google Scholar] [PubMed]
- Tutuncu, N.B.; Erbas, T.; Alikasifoglu, M.; Tuncbilek, E. Thermolabile methylenetet-rahydrofolate reductase enzyme genotype is frequent in type 2 diabetic patients with normal fasting homocysteine levels. J. Intern. Med. 2005, 257, 446–453. [Google Scholar] [CrossRef]
- Wang, H.; Fan, D.; Hong, T. Is the C677T polymorphism in methylenetetrahydrofo-late reductase gene or plasma homocysteine a risk factor for diabetic peripheral neuropathy in Chinese individuals? Neural Regen. Res. 2012, 7, 2384–2391. [Google Scholar] [PubMed]
- Rahimi, M.; Hasanvand, A.; Rahimi, Z.; Vaisi-Raygani, A.; Mozafari, H.; Rezaei, M.; Zargooshi, J.; Najafi, F.; Shakiba, E. Synergistic effects of the MTHFR C677T and A1298C polymorphisms on the increased risk of micro-and macro-albuminuria and progression of diabetic nephropathy among Iranians with type 2 diabetes mellitus. Clin. Biochem. 2010, 43, 1333–1339. [Google Scholar] [CrossRef]
- Ezzidi, I.; Mtiraoui, N.; Mohamed, M.; Mahjoub, T.; Kacem, M.; Almawi, W. Association of endothelial nitric oxide synthase Glu298Asp, 4b/a, and-786T>C gene vari-ants with diabetic nephropathy. J. Diabetes Complicat. 2008, 22, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Dellamea, B.S.; Pinto, L.C.; Leitão, C.B.; Santos, K.G.; Canani, L.H. Endothelial nitric oxide synthase gene polymorphisms and risk of diabetic nephropa-thy: A systematic review and meta-analysis. BMC Med. Genet. 2014, 15, 9. [Google Scholar] [CrossRef] [PubMed]
- Bonnefond, A.; Saulnier, P.J.; Stathopoulou, M.G.; Grarup, N.; Ndiaye, N.C.; Roussel, R.; Nezhad, M.A.; Dechaume, A.; Lantieri, O.; Hercberg, S.; et al. What is the contribution of two genetic variants regulating VEGF levels to type 2 diabetes risk and to microvascular complications? PLoS ONE 2013, 8, e55921. [Google Scholar]
- Babizhayev, M.A.; Strokov, I.A.; Nosikov, V.V.; Savel’yeva, E.L.; Sitnikov, V.F.; Yegorov, Y.E.; Lankin, V.Z. The Role of Oxidative Stress in Diabetic Neuropathy: Generation of Free Radical Species in the Glycation Reaction and Gene Polymorphisms Encoding Antioxidant Enzymes to Genetic Susceptibility to Diabetic Neuropathy in Population of Type I Diabetic Patients. Cell Biochem. Biophys. 2015, 71, 1425–1443. [Google Scholar] [CrossRef]
- Meng, W.; Deshmukh, H.A.; Donnelly, L.A.; Torrance, N.; Colhoun, H.M.; Palmer, C.N.; Smith, B.H.; Wellcome Trust Case Control Consortium 2 (WTCCC2); Surrogate Markers for Micro-and Macro-Vascular Hard Endpoints for Innovative Diabetes Tools (SUMMIT) Study Group. A Genome-wide Association Study Provides Evidence of Sex-specific Involvement of Chr1p35.1 (ZSCAN20-TLR12P) and Chr8p23.1 (HMGB1P46) With Diabetic Neuropathic Pain. EBioMedicine 2015, 2, 1386–1393. [Google Scholar] [CrossRef] [PubMed]
- Testa, R.; Bonfigli, A.R.; Prattichizzo, F.; La Sala, L.; De Nigris, V.; Ceriello, A. The “Metabolic Memory” Theory and the Early Treatment of Hyperglycemia in Prevention of Diabetic Complications. Nutrients 2017, 9, 437. [Google Scholar] [CrossRef]
- Berger, S.L.; Kouzarides, T.; Shiekhattar, R.; Shilatifard, A. An Operational Definition of Epigenetics. Genes Dev. 2009, 23, 781–783. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.A.; Zhang, E.; Natarajan, R. Epigenetic Mechanisms in Diabetic Complications and Metabolic Memory. Diabetologia 2015, 58, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Luo, C.; Hajkova, P.; Ecker, J.R. Dynamic DNA methylation: In the right place at the right time. Science 2018, 361, 1336–1340. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Miao, F.; Paterson, A.D.; Lachin, J.M.; Zhang, L.; Schones, D.E.; Wu, X.; Wang, J.; Tompkins, J.D.; Genuth, S.; et al. DCCT/EDIC Research Group. Epigenomic Profiling Reveals an Association between Persistence of DNA Methylation and Metabolic Memory in the DCCT/EDIC Type 1 Diabetes Cohort. Proc. Natl. Acad. Sci. USA 2016, 113, E3002–E3011. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Han, X.; Wang, M.; Hu, Q.; Li, S.; Wang, M.; Hu, J. The Association between Genomic DNA Methylation and Diabetic Peripheral Neuropathy in Patients with Type 2 Diabetes Mellitus. J. Diabetes Res. 2019, 2019, 2494057. [Google Scholar] [CrossRef] [PubMed]
- Gastoł, J.; Kapusta, P.; Polus, A.; Pitera, E.; Biela, M.; Wołkow, P.; Pawliński, Ł.; Kieć-Wilk, B. Epigenetic Mechanism in Search for the Pathomechanism of Diabetic Neuropathy Development in Diabetes Mellitus Type 1 (T1DM). Endocrine 2020, 68, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Araki, T.; Milbrandt, J. Ninjurin2, a Novel Homophilic Adhesion Molecule, Is Expressed in Mature Sensory and Enteric Neurons and Promotes Neurite Outgrowth. J. Neurosci. 2000, 20, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Blattler, A.; Farnham, P.J. Cross-talk between site-specific transcription factors and DNA methylation states. J. Biol. Chem. 2013, 288, 34287–34294. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Elzinga, S.; Eid, S.; Figueroa-Romero, C.; Hinder, L.M.; Pacut, C.; Feldman, E.L.; Hur, J. Genome-Wide DNA Methylation Profiling of Human Diabetic Peripheral Neuropathy in Subjects with Type 2 Diabetes Mellitus. Epigenetics 2019, 14, 766–779. [Google Scholar] [CrossRef]
- Guo, K.; Eid, S.A.; Elzinga, S.E.; Pacut, C.; Feldman, E.L.; Hur, J. Genome-Wide Profiling of DNA Methylation and Gene Expression Identifies Candidate Genes for Human Diabetic Neuropathy [Published Correction Appears in Clin Epigenetics. Clin. Epigenet. 2020, 12, 123. [Google Scholar] [CrossRef]
- Hur, J.; Sullivan, K.A.; Pande, M.; Hong, Y.; Sima, A.A.F.; Jagadish, H.V.; Kretzler, M.; Feldman, E.L. The Identification of Gene Expression Profiles Associated with Progression of Human Diabetic Neuropathy. Brain 2011, 134, 3222–3235. [Google Scholar] [CrossRef]
- Liu, J. Control of protein synthesis and mRNA degradation by microRNAs. Curr. Opin. Cell Biol. 2008, 20, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Xourgia, E.; Papazafiropoulou, A.; Melidonis, A. Circulating MicroRNAs as Biomarkers for Diabetic Neuropathy: A Novel Approach. World, J. Exp. Med. 2018, 8, 18–23. [Google Scholar] [CrossRef]
- Simeoli, R.; Fierabracci, A. Insights into the Role of MicroRNAs in the Onset and Development of Diabetic Neuropathy. Int. J. Mol. Sci. 2019, 20, 4627. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.B.; Wu, Q.; Liu, J.; Fan, Y.Z.; Yu, K.F.; Cai, Y. miR-199a-3p is involved in the pathogenesis and progression of diabetic neuropathy through downregulation of SerpinE2. Mol. Med. Rep. 2017, 16, 2417–2424. [Google Scholar] [CrossRef]
- Liu, W.; Ao, Q.; Guo, Q.; He, W.; Peng, L.; Jiang, J.; Hu, X. miR-9 Mediates CALHM1-Activated ATP-P2X7R Signal in Painful Diabetic Neuropathy Rats. Mol. Neurobiol. 2017, 54, 922–929. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, C.; Liu, J.; Bi, Y.; Li, H. Inhibition of miR-25 aggravates diabetic peripheral neuropathy. Neuroreport 2018, 29, 945–953. [Google Scholar] [CrossRef]
- Feng, Y.; Chen, L.; Luo, Q.; Wu, M.; Chen, Y.; Shi, X. Involvement of microRNA-146a in diabetic peripheral neuropathy through the regulation of inflammation. Drug Des. Devel. Ther. 2018, 12, 171–177. [Google Scholar] [CrossRef]
- Wang, L.; Chopp, M.; Lu, X.; Szalad, A.; Jia, L.; Liu, X.S.; Wu, K.H.; Lu, M.; Zhang, Z.G. miR-146a mediates thymosin β4 induced neurovascular remodeling of diabetic peripheral neuropathy in type-II diabetic mice. Brain Res. 2019, 1707, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Yang, Q.; Wei, X.; Liu, Y.; Ma, D.; Li, J.; Wan, Y.; Luo, Y. The role of miR-190a-5p contributes to diabetic neuropathic pain via targeting SLC17A6. J. Pain Res. 2017, 10, 2395–2403. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.A.; Liang, H.; Li, W.-H. Human Polymorphism at MicroRNAs and MicroRNA Target Sites. Proc. Natl. Acad. Sci. USA 2007, 104, 3300–3305. [Google Scholar] [CrossRef]
- Ciccacci, C.; Morganti, R.; Di Fusco, D.; D’Amato, C.; Cacciotti, L.; Greco, C.; Rufini, S.; Novelli, G.; Sangiuolo, F.; Marfia, G.A.; et al. Common Polymorphisms in MIR146a, MIR128a and MIR27a Genes Contribute to Neuropathy Susceptibility in Type 2 Diabetes. Acta Diabetol. 2014, 51, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Ciccacci, C.; Latini, A.; Greco, C.; Politi, C.; D’Amato, C.; Lauro, D.; Novelli, G.; Borgiani, P.; Spallone, V. Association between a MIR499A Polymorphism and Diabetic Neuropathy in Type 2 Diabetes. J. Diabetes Complicat. 2018, 32, 11–17. [Google Scholar] [CrossRef]
- Latini, A.; Borgiani, P.; De Benedittis, G.; D’Amato, C.; Greco, C.; Lauro, D.; Novelli, G.; Spallone, V.; Ciccacci, C. Mitochondrial DNA Copy Number in Peripheral Blood Is Reduced in Type 2 Diabetes Patients with Polyneuropathy and Associated with a MIR499A Gene Polymorphism. DNA Cell Biol. 2020, 39, 1467–1472. [Google Scholar] [CrossRef]
- Batista, P.J.; Chang, H.Y. Long noncoding RNAs: Cellular address codes in development and disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef]
- Wang, S.; Xu, H.; Zou, L.; Xie, J.; Wu, H.; Wu, B.; Yi, Z.; Lv, Q.; Zhang, X.; Ying, M.; et al. 48+ Is Involved in Diabetic Neuropathic Pain Mediated by the P2X3 Receptor in the Dorsal Root Ganglia. Purinergic Signal. 2016, 12, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Zhao, G.Q.; Cao, R.J.; Zhu, Z.H.; Li, K. LncRNANONRATT021972 Was Associated with Neuropathic Pain Scoring in Patients with Type 2 Diabetes. Behav. Neurol. 2017, 2017, 2941297. [Google Scholar] [CrossRef]
- Liu, C.; Li, C.; Deng, Z.; Du, E.; Xu, C. Long non-coding RNA BC168687 is involved in TRPV1-mediated diabetic neuropathic pain in rats. Neuroscience 2018, 15, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, X.; Chen, X.; Li, S.; Ho, I.H.T.; Liu, X.; Chan, M.T.V.; Wu, W.K.K. Emerging Roles of Long Non-Coding RNAs in Neuropathic Pain. Cell Prolif 2019, 52, e12528. [Google Scholar] [CrossRef]
- Xu, H.; He, L.; Liu, C.; Tang, L.; Xu, Y.; Xiong, M.; Yang, M.; Fan, Y.; Hu, F.; Liu, X.; et al. LncRNA NONRATT021972 SiRNA Attenuates P2X7 Receptor Expression and Inflammatory Cytokine Production Induced by Combined High Glucose and Free Fatty Acids in PC12 Cells. Purinergic Signal. 2016, 12, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Zhang, C.; Zou, L.; Ma, Y.; Huang, K.; Lv, Q.; Zhang, X.; Wang, S.; Xue, Y.; Yi, Z.; et al. 48+ SiRNA Improved Diabetic Sympathetic Neuropathy in Type 2 Diabetic Rats Mediated By P2X7 Receptor in SCG. Auton. Neurosci. 2016, 197, 14–18. [Google Scholar] [CrossRef]
- Peng, H.; Zou, L.; Xie, J.; Wu, H.; Wu, B.; Zhu, G.; Lv, Q.; Zhang, X.; Liu, S.; Li, G.; et al. LncRNA NONRATT021972 SiRNA Decreases Diabetic Neuropathic Pain Mediated by the P2x(3) Receptor in Dorsal Root Ganglia. Mol. Neurobiol. 2017, 54, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Tao, J.; Wu, H.; Yang, Y.; Chen, Q.; Deng, Z.; Liu, J.; Xu, C. Effects of LncRNA BC168687 siRNA on diabetic neuropathic pain mediated by P2X(7) Receptor on SGCs in DRG of rats. Biomed. Res. Int. 2017, 2017, 7831251. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Ji, L.-D.; Cai, J.-J.; Feng, M.; Zhou, M.; Hu, S.-P.; Xu, J.; Zhou, W.-H. Microarray Analysis of Long Noncoding RNAs in Female Diabetic Peripheral Neuropathy Patients. Cell. Physiol. Biochem. 2018, 46, 1209–1217. [Google Scholar] [CrossRef]
- Du, H.; Liu, Z.; Tan, X.; Ma, Y.; Gong, Q. Identification of the Genome-Wide Expression Patterns of Long Non-Coding RNAs and mRNAs in Mice with Streptozotocin-Induced Diabetic Neuropathic Pain. Neuroscience 2019, 402, 90–103. [Google Scholar] [CrossRef]
- Guo, G.; Ren, S.; Kang, Y.; Liu, Y.; Duscher, D.; Machens, H.-G.; Chen, Z. Microarray Analyses of LncRNAs and mRNAs Expression Profiling Associated with Diabetic Peripheral Neuropathy in Rats. J. Cell. Biochem. 2019, 120, 15347–15359. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Yao, B.; Xu, W.; Chen, J.; Zhou, X. lncRNA H19/miR-675 axis regulates cardiomyocyte apoptosis by targeting VDAC1 in diabetic cardiomyopathy. Sci. Rep. 2016, 6, 36340. [Google Scholar] [CrossRef] [PubMed]
- Margueron, R.; Reinberg, D. Chromatin structure and the inheritance of epigenetic information. Nat. Rev. Genet. 2010, 11, 285–296. [Google Scholar] [CrossRef]
- Denk, F.; Huang, W.; Sidders, B.; Bithell, A.; Crow, M.; Grist, J.; Sharma, S.; Ziemek, D.; Rice, A.S.C.; Buckley, N.J.; et al. HDAC Inhibitors Attenuate the Development of Hypersensitivity in Models of Neuropathic Pain. Pain 2013, 154, 1668–1679. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, X.; Xu, Y.; Xu, S.; Xia, F.; Zhu, B.; Liu, Y.; Wang, W.; Wu, H.; Wang, F. The Etiological Changes of Acetylation in Peripheral Nerve Injury-Induced Neuropathic Hypersensitivity. Mol. Pain 2018, 14, 1744806918798408. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, C.; Sun, G. Histone Acetylation and Its Modifiers in the Pathogenesis of Diabetic Nephropathy. J. Diabetes Res. 2016, 2016, 4065382. [Google Scholar] [CrossRef] [PubMed]
- Abouhish, H.; Thounaojam, M.C.; Jadeja, R.N.; Gutsaeva, D.R.; Powell, F.L.; Khriza, M.; Martin, P.M.; Bartoli, M. Inhibition of HDAC6 Attenuates Diabetes-Induced Retinal Redox Imbalance and Microangiopathy. Antioxidants (Basel) 2020, 9, 599. [Google Scholar] [CrossRef]
- Du, W.; Wang, N.; Li, F.; Jia, K.; An, J.; Liu, Y.; Wang, Y.; Zhu, L.; Zhao, S.; Hao, J. STAT3 Phosphorylation Mediates High Glucose-Impaired Cell Autophagy in an HDAC1-Dependent and-Independent Manner in Schwann Cells of Diabetic Peripheral Neuropathy. FASEB J. 2019, 33, 8008–8021. [Google Scholar] [CrossRef]
- Boulton, A.J.; Vinik, A.I.; Arezzo, J.C.; Bril, V.; Feldman, E.L.; Freeman, R.; Malik, R.A.; Maser, R.E.; Sosenko, J.M.; American Diabetes Association; et al. Diabetic neuropathies: A statement by the American Diabetes Association. Diabetes Care 2005, 28, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic, M.; Keaton, J.M.; Lynch, J.A.; Miller, D.R.; Zhou, J.; Tcheandjieu, C.; Huffman, J.E.; Assimes, T.L.; Lorenz, K.; Zhu, X.; et al. Discovery of 318 new risk loci for type 2 diabetes and related vascular outcomes among 1.4 million participants in a multi-ancestry meta-analysis. Nat. Genet. 2020, 52, 680–691. [Google Scholar] [CrossRef]
- Pollastro, C.; Ziviello, C.; Costa, V.; Ciccodicola, A. Pharmacogenomics of Drug Response in Type 2 Diabetes: Toward the Definition of Tailored Therapies? PPAR Res. 2015, 2015, 415149. [Google Scholar] [CrossRef]
- Becker, M.L.; Pearson, E.R.; Tkáč, I. Pharmacogenetics of oral antidiabetic drugs. Int. J. Endocrinol. 2013, 2013, 686315. [Google Scholar] [CrossRef]
- Faber, C.G.; Hoeijmakers, J.G.; Ahn, H.S.; Cheng, X.; Han, C.; Choi, J.S.; Estacion, M.; Lauria, G.; Vanhoutte, E.K.; Gerrits, M.M.; et al. Gain of function Naν1.7 mutations in idiopathic small fiber neuropathy. Ann. Neurol. 2012, 71, 26–39. [Google Scholar] [CrossRef]
- Li, Q.S.; Cheng, P.; Favis, R.; Wickenden, A.; Romano, G.; Wang, H. SCN9Avariants may be implicated in neuropathic pain associated with diabetic peripheral neuropathy and pain severity. Clin. J. Pain. 2015, 31, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Blesneac, I.; Themistocleous, A.C.; Fratter, C.; Conrad, L.J.; Ramirez, J.D.; Cox, J.J.; Tesfaye, S.; Shillo, P.R.; Rice, A.S.C.; Tucker, S.J.; et al. Rare NaV1.7 variants associated with painful diabetic peripheral neuropathy. Pain 2018, 159, 469–480. [Google Scholar] [CrossRef] [PubMed]
- de Greef, B.T.A.; Hoeijmakers, J.G.J.; Geerts, M.; Oakes, M.; Church, T.J.E.; Waxman, S.G.; Dib-Hajj, S.D.; Faber, C.G.; Merkies, I.S.J. Lacosamide in patients with Nav1.7 mutations-related small fibre neuropathy: A randomized controlled trial. Brain 2019, 142, 263–275. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Zhang, X.; Jia, K.; Zhang, C.; Zhu, L.; Cheng, M.; Li, F.; Zhao, S.; Hao, J. Trichostatin A Increases BDNF Protein Expression by Improving XBP-1s/ATF6/GRP78 Axis in Schwann Cells of Diabetic Peripheral Neuropathy. Biomed. Pharmacother. 2021, 133, 111062. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Feng, Y.; Xie, Y.; Shao, Y.; Wu, M.; Deng, X.; Yuan, W.E.; Chen, Y.; Shi, X. Nanoparticle-microRNA-146a-5p polyplexes ameliorate diabetic peripheral neuropathy by modulating inflammation and apoptosis. Nanomedicine 2019, 17, 188–197. [Google Scholar] [CrossRef] [PubMed]



| Type of Neuropathies due to Fiber Size | Autonomic Neuropathy [12,13,14,15,16,17,18,19,20] | ||||
|---|---|---|---|---|---|
| Small Fiber [11,12] | Large Fiber [12] | Systems | Presentation | Possible Pathogenesis and Origin | |
| Motor deficits | P | + | Cardiovascular | Myocardial infarctions | Coronary artery disease |
| Nocturnal hypertension | Sustained adrenergic activity during sleep | ||||
| Orthostatic hypotension | Efferent sympathetic vasomotor denervation | ||||
| Tachycardia | Vagal cardiac neuropathy | ||||
| Sensory loss | + | + | Sudden death | Modified perception of myocardial ischemia; impaired hemodynamic response to cardiovascular stresses; cardiac arrhythmias; impaired sympathetic–parasympathetic cardiac innervation balance | |
| Pain | +; Sharp;of C fiber type | + | Gastrointestinal | Gastroparesis | Impaired vagal activity and intrinsic enteric autonomic nerves, reduced nerve fiber amount in antral biopsies, loss of neuronal nitric oxide synthase (nNOS) |
| Diarrhea | Visceral hypersensitivity | ||||
| Constipation | Not fully understood; altered secretion of gastrointestinal hormones | ||||
| Fecal incontinence | Incompetence of anal sphincter or ↓ rectal sensation | ||||
| Vibration sensation | ↓ | ↓ | Genitourinary | Bladder dysfunction | Afferent and efferent autonomic nerves dysfunction; bladder smooth muscle dysfunction; and urothelial abnormalities |
| Erectile dysfunction | Morphological alterations of autonomic nerve fibers | ||||
| Thermal sensation | ↓ | ↓ | Ejaculatory failure | Sympathetic nervous system dysfunction | |
| Tendon reflex | P(∗) | Depressed | ↓ Sexual desire in females | Sexual dysfunction in diabetic females might be related more to psychogenic factors, probably due to the disease burden | |
| Origin | Dorsal root ganglion | ↑ Pain during intercourse in females | |||
| Presentation | Length-dependent ** | Various sensory and motor signs | Sudomotor | Loss of sweating, dry cold skin | Impaired postsympathetic cholinergic nerve fibers activity, which release acetylcholine |
| Autonomous symptoms | +/−; impaired blood flow | ↑ blood flow | |||
| Gene/Locus | Polymorphism | Risk Allele | Mechanism | References |
|---|---|---|---|---|
| ALR2 | -106C/T | T | Polyol pathway, oxidative stress | [55] |
| 5′-(CA)n | Z-2 (CA22) | [53,56] | ||
| ACE | I/D (intron 16) | D | oxidative stress, vascular changes | [4,60,61,63] |
| APOE | e2, e3, e4 | e4 | lipid metabolism, nerve repair and regeneration | [1,2,64,65] |
| MTHFR | 677C/T | T | hyperhomocysteinemia, lipid metabolism | [1,2,4,66,67,68,69] |
| 1298A/C | C | |||
| NOS3 | -786T/C | C | vascular changes, oxidative stress | [1,70,71] |
| 894G/T | T | |||
| 4a/b | 4a | |||
| 6p21.1 (VEGF–related) | rs6921438 A/G | A | vascular changes | [1,2,72] |
| 9p24.2 (VEGF–related) | rs10738760 A/G | A | vascular changes | |
| GPx-1 | 599C/T | T | oxidative stress | [2,73] |
| CAT | -262C/T | T | oxidative stress | [2,73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankovic, M.; Novakovic, I.; Nikolic, D.; Mitrovic Maksic, J.; Brankovic, S.; Petronic, I.; Cirovic, D.; Ducic, S.; Grajic, M.; Bogicevic, D. Genetic and Epigenomic Modifiers of Diabetic Neuropathy. Int. J. Mol. Sci. 2021, 22, 4887. https://doi.org/10.3390/ijms22094887
Jankovic M, Novakovic I, Nikolic D, Mitrovic Maksic J, Brankovic S, Petronic I, Cirovic D, Ducic S, Grajic M, Bogicevic D. Genetic and Epigenomic Modifiers of Diabetic Neuropathy. International Journal of Molecular Sciences. 2021; 22(9):4887. https://doi.org/10.3390/ijms22094887
Chicago/Turabian StyleJankovic, Milena, Ivana Novakovic, Dejan Nikolic, Jasmina Mitrovic Maksic, Slavko Brankovic, Ivana Petronic, Dragana Cirovic, Sinisa Ducic, Mirko Grajic, and Dragana Bogicevic. 2021. "Genetic and Epigenomic Modifiers of Diabetic Neuropathy" International Journal of Molecular Sciences 22, no. 9: 4887. https://doi.org/10.3390/ijms22094887
APA StyleJankovic, M., Novakovic, I., Nikolic, D., Mitrovic Maksic, J., Brankovic, S., Petronic, I., Cirovic, D., Ducic, S., Grajic, M., & Bogicevic, D. (2021). Genetic and Epigenomic Modifiers of Diabetic Neuropathy. International Journal of Molecular Sciences, 22(9), 4887. https://doi.org/10.3390/ijms22094887