
Vitamin D and Beta-Glucans Synergically Stimulate Human Macrophage Activity



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Vitamin D and Beta-Glucans Stimulate the Synthesis and Secretion of IL-8 in Human Macrophages
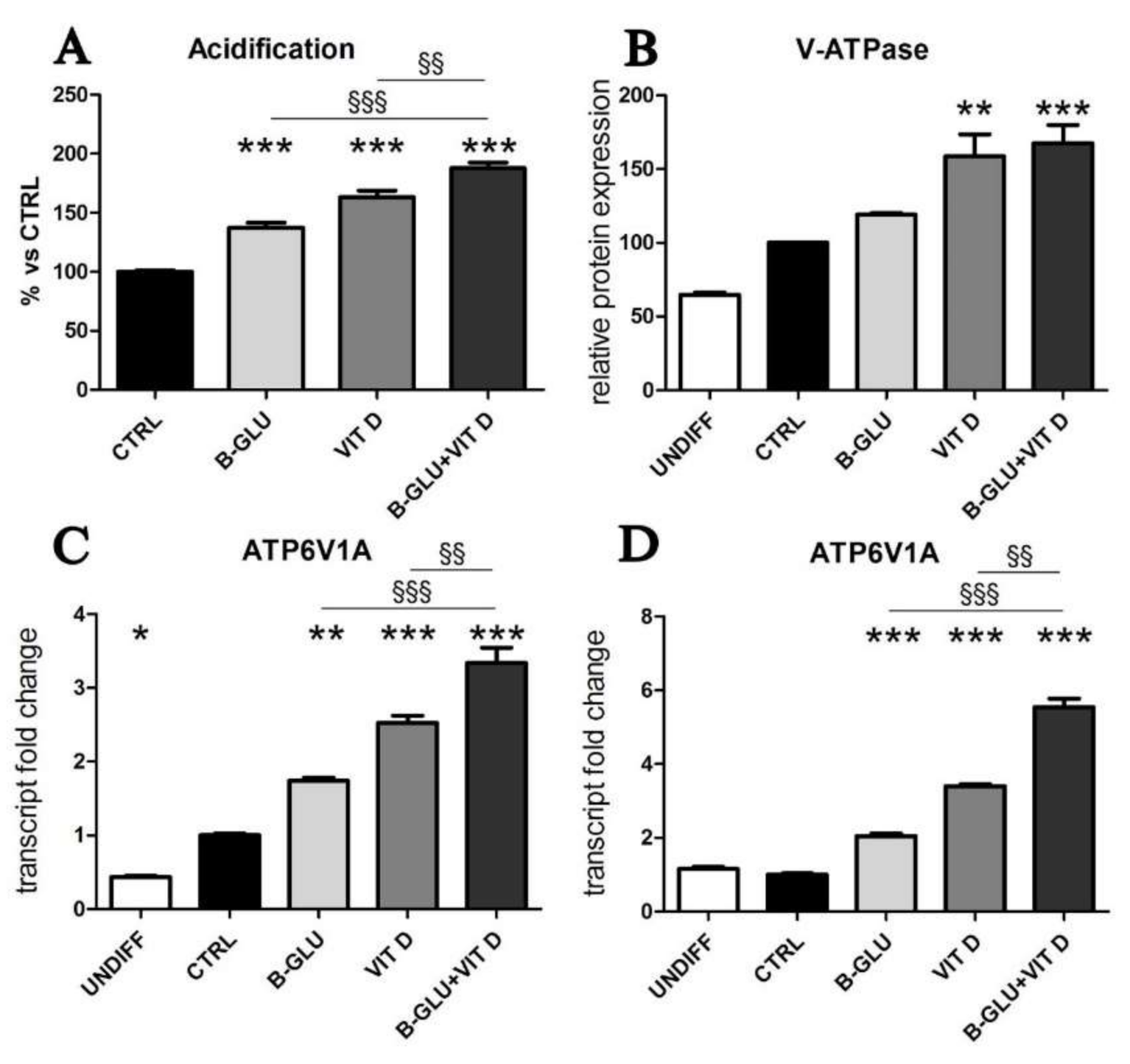
2.2. Vitamin D and Beta-Glucans Stimulate the Intracellular Vesicle Acidification of Human Macrophages
2.3. Vitamin D and Beta-Glucans Stimulate the Mitochondrial Metabolism of Human Macrophages
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatments
4.2. Real-Time Polymerase Chain Reaction (qRT-PCR)
4.3. IL-8 Quantification
4.4. Acidification Assay
4.5. Western Blotting Analysis
4.6. ROS Essay
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Dixit, N.; Simon, S.I. Chemokines, Selectins and Intracellular Calcium Flux: Temporal and Spatial Cues for Leukocyte Arrest. Front. Immunol. 2012, 3. [Google Scholar] [CrossRef]
- Cafforio, P.; D’Oronzo, S.; Felici, C.; Sigala, S.; Fragni, M.; Silvestris, F. 1,25(OH)2 Vitamin D(3) Contributes to Osteoclast-like Trans-Differentiation of Malignant Plasma Cells. Exp. Cell Res. 2017, 358, 260–268. [Google Scholar] [CrossRef]
- Chung, M.; Balk, E.M.; Brendel, M.; Ip, S.; Lau, J.; Lee, J.; Lichtenstein, A.; Patel, K.; Raman, G.; Tatsioni, A.; et al. Vitamin D and Calcium: A Systematic Review of Health Outcomes. Evid. Rep. Technol. Assess. 2009, 183, 1–420. [Google Scholar]
- Chen, L.; Eapen, M.S.; Zosky, G.R. Vitamin D Both Facilitates and Attenuates the Cellular Response to Lipopolysaccharide. Sci. Rep. 2017, 7, 45172. [Google Scholar] [CrossRef]
- Adams, J.; Hewison, M. Extrarenal Expression of the 25-Hydroxyvitamin D-1-Hydroxylase. Arch. Biochem. Biophys. 2012, 523, 95–102. [Google Scholar] [CrossRef]
- Muñoz Garcia, A.; Kutmon, M.; Eijssen, L.; Hewison, M.; Evelo, C.T.; Coort, S.L. Pathway Analysis of Transcriptomic Data Shows Immunometabolic Effects of Vitamin D. J. Mol. Endocrinol 2018, 60, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Calton, E.K.; Keane, K.N.; Newsholme, P.; Soares, M.J. The Impact of Vitamin D Levels on Inflammatory Status: A Systematic Review of Immune Cell Studies. PLoS ONE 2015, 10, e0141770. [Google Scholar] [CrossRef]
- Ryynänen, J.; Carlberg, C. Primary 1,25-Dihydroxyvitamin D3 Response of the Interleukin 8 Gene Cluster in Human Monocyte- and Macrophage-like Cells. PLoS ONE 2013, 8, e78170. [Google Scholar] [CrossRef]
- Kim, H.S.; Hong, J.T.; Kim, Y.; Han, S.-B. Stimulatory Effect of β-Glucans on Immune Cells. Immune Netw. 2011, 11, 191–195. [Google Scholar] [CrossRef]
- Xiang, D.; Sharma, V.R.; Freter, C.E.; Yan, J. Anti-Tumor Monoclonal Antibodies in Conjunction with β-Glucans: A Novel Anti-Cancer Immunotherapy. Curr. Med. Chem. 2012, 19, 4298–4305. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Yan, J.; Baran, J.T.; Allendorf, D.J.; Hansen, R.D.; Ostroff, G.R.; Xing, P.X.; Cheung, N.-K.V.; Ross, G.D. Mechanism by Which Orally Administered Beta-1,3-Glucans Enhance the Tumoricidal Activity of Antitumor Monoclonal Antibodies in Murine Tumor Models. J. Immunol. 2004, 173, 797–806. [Google Scholar] [CrossRef]
- Zheng, X.; Zou, S.; Xu, H.; Liu, Q.; Song, J.; Xu, M.; Xu, X.; Zhang, L. The Linear Structure of β-Glucan from Baker’s Yeast and Its Activation of Macrophage-like RAW264.7 Cells. Carbohydr. Polym. 2016, 148, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Toll-like Receptors and Their Crosstalk with Other Innate Receptors in Infection and Immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Rice, P.J.; Adams, E.L.; Ozment-Skelton, T.; Gonzalez, A.J.; Goldman, M.P.; Lockhart, B.E.; Barker, L.A.; Breuel, K.F.; Deponti, W.K.; Kalbfleisch, J.H.; et al. Oral Delivery and Gastrointestinal Absorption of Soluble Glucans Stimulate Increased Resistance to Infectious Challenge. J. Pharmacol. Exp. Ther. 2005, 314, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Gabig, T.G.; Babior, B.M. The Killing of Pathogens by Phagocytes. Annu. Rev. Med. 1981, 32, 313–326. [Google Scholar] [CrossRef]
- Wang, T.-T.; Nestel, F.P.; Bourdeau, V.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.W.; Mader, S.; et al. Cutting Edge: 1,25-Dihydroxyvitamin D3 Is a Direct Inducer of Antimicrobial Peptide Gene Expression. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human Cathelicidin Antimicrobial Peptide (CAMP) Gene Is a Direct Target of the Vitamin D Receptor and Is Strongly up-Regulated in Myeloid Cells by 1,25-Dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef]
- Verway, M.; Bouttier, M.; Wang, T.-T.; Carrier, M.; Calderon, M.; An, B.-S.; Devemy, E.; McIntosh, F.; Divangahi, M.; Behr, M.A.; et al. Vitamin D Induces Interleukin-1β Expression: Paracrine Macrophage Epithelial Signaling Controls M. Tuberculosis Infection. PLoS Pathog 2013, 9, e1003407. [Google Scholar] [CrossRef]
- Chanput, W.; Reitsma, M.; Kleinjans, L.; Mes, J.J.; Savelkoul, H.F.J.; Wichers, H.J. β-Glucans Are Involved in Immune-Modulation of THP-1 Macrophages. Mol. Nutr. Food Res. 2012, 56, 822–833. [Google Scholar] [CrossRef] [PubMed]
- Ikewaki, N.; Fujii, N.; Onaka, T.; Ikewaki, S.; Inoko, H. Immunological Actions of Sophy Beta-Glucan (Beta-1,3-1,6 Glucan), Currently Available Commercially as a Health Food Supplement. Microbiol. Immunol. 2007, 51, 861–873. [Google Scholar] [CrossRef]
- Lee, B.S.; Underhill, D.M.; Crane, M.K.; Gluck, S.L. Transcriptional Regulation of the Vacuolar H(+)-ATPase B2 Subunit Gene in Differentiating THP-1 Cells. J. Biol. Chem. 1995, 270, 7320–7329. [Google Scholar] [CrossRef] [PubMed]
- Futai, M.; Sun-Wada, G.-H.; Wada, Y.; Matsumoto, N.; Nakanishi-Matsui, M. Vacuolar-Type ATPase: A Proton Pump to Lysosomal Trafficking. Proc. Jpn. Aca.d Ser. B Phys. Biol. Sci. 2019, 95, 261–277. [Google Scholar] [CrossRef]
- Bartkiewicz, M.; Hernando, N.; Reddy, S.V.; Roodman, G.D.; Baron, R. Characterization of the Osteoclast Vacuolar H(+)-ATPase B-Subunit. Gene 1995, 160, 157–164. [Google Scholar] [CrossRef]
- Lee, B.S.; Holliday, L.S.; Krits, I.; Gluck, S.L. Vacuolar H+-ATPase Activity and Expression in Mouse Bone Marrow Cultures. J. Bone Miner. Res. 1999, 14, 2127–2136. [Google Scholar] [CrossRef]
- Harant, H.; Andrew, P.J.; Reddy, G.S.; Foglar, E.; Lindley, I.J. 1alpha,25-Dihydroxyvitamin D3 and a Variety of Its Natural Metabolites Transcriptionally Repress Nuclear-Factor-KappaB-Mediated Interleukin-8 Gene Expression. Eur J. Biochem 1997, 250, 63–71. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, J.; Ge, X.; Du, J.; Deb, D.K.; Li, Y.C. Vitamin D Receptor Inhibits Nuclear Factor ΚB Activation by Interacting with IκB Kinase β Protein. J. Biol. Chem. 2013, 288, 19450–19458. [Google Scholar] [CrossRef]
- Wu, S.; Liao, A.P.; Xia, Y.; Li, Y.C.; Li, J.-D.; Sartor, R.B.; Sun, J. Vitamin D Receptor Negatively Regulates Bacterial-Stimulated NF-KappaB Activity in Intestine. Am. J. Pathol. 2010, 177, 686–697. [Google Scholar] [CrossRef]
- Mai, S.; Brehm, N.; Auburger, G.; Bereiter-Hahn, J.; Jendrach, M. Age-Related Dysfunction of the Autophago-Lysosomal Pathway in Human Endothelial Cells. Pflugers Arch. 2019, 471, 1065–1078. [Google Scholar] [CrossRef]
- Brown, D.; Breton, S. Mitochondria-Rich, Proton-Secreting Epithelial Cells. J. Exp. Biol. 1996, 199, 2345–2358. [Google Scholar] [CrossRef]
- Yamamoto, T.; Sakaguchi, N.; Hachiya, M.; Nakayama, F.; Yamakawa, M.; Akashi, M. Role of Catalase in Monocytic Differentiation of U937 Cells by TPA: Hydrogen Peroxide as a Second Messenger. Leukemia 2009, 23, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Barendsen, N.; Mueller, M.; Chen, B. Inhibition of TPA-Induced Monocytic Differentiation in THP-1 Human Monocytic Leukemic Cells by Staurosporine, a Potent Protein Kinase C Inhibitor. Leuk. Res. 1990, 14, 467–474. [Google Scholar] [CrossRef]
- Chorváth, B.; Sedlák, J.; Duraj, J.; Klobusická, M.; Plesková, I.; Augustinová, M.; Suliková, M. Biochemical and Immunological Phenotype Alterations in Human Monoblastoid Cell Line U-937 Induced by Physiological (Interferon-Alpha, Retinoic Acid) and Nonphysiological (Phorbol Ester) Inducers. Neoplasma 1990, 37, 317–331. [Google Scholar]
- Tsuchiya, S.; Kobayashi, Y.; Goto, Y.; Okumura, H.; Nakae, S.; Konno, T.; Tada, K. Induction of Maturation in Cultured Human Monocytic Leukemia Cells by a Phorbol Diester. Cancer Res. 1982, 42, 1530–1536. [Google Scholar]
- Chanput, W.; Peters, V.; Wichers, H. THP-1 and U937 Cells. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerlan, 2015; ISBN 978-3-319-15791-7. [Google Scholar]
- Hass, R.; Giese, G.; Meyer, G.; Hartmann, A.; Dörk, T.; Köhler, L.; Resch, K.; Traub, P.; Goppelt-Strübe, M. Differentiation and Retrodifferentiation of U937 Cells: Reversible Induction and Suppression of Intermediate Filament Protein Synthesis. Eur. J. Cell Biol. 1990, 51, 265–271. [Google Scholar]
- Soderholm, A.T.; Barnett, T.C.; Korn, O.; Rivera-Hernandez, T.; Seymour, L.M.; Schulz, B.L.; Nizet, V.; Wells, C.A.; Sweet, M.J.; Walker, M.J. Group A Streptococcus M1T1 Intracellular Infection of Primary Tonsil Epithelial Cells Dampens Levels of Secreted IL-8 Through the Action of SpyCEP. Front. Cell Infect. Microbiol. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Ricca, C.; Aillon, A.; Bergandi, L.; Alotto, D.; Castagnoli, C.; Silvagno, F. Vitamin D Receptor Is Necessary for Mitochondrial Function and Cell Health. Int. J. Mol. Sci. 2018, 19, 1672. [Google Scholar] [CrossRef]
- Seglen, P.O. Inhibitors of Lysosomal Function. Methods Enzymol. 1983, 96, 737–764. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergandi, L.; Apprato, G.; Silvagno, F. Vitamin D and Beta-Glucans Synergically Stimulate Human Macrophage Activity. Int. J. Mol. Sci. 2021, 22, 4869. https://doi.org/10.3390/ijms22094869
Bergandi L, Apprato G, Silvagno F. Vitamin D and Beta-Glucans Synergically Stimulate Human Macrophage Activity. International Journal of Molecular Sciences. 2021; 22(9):4869. https://doi.org/10.3390/ijms22094869
Chicago/Turabian StyleBergandi, Loredana, Giulia Apprato, and Francesca Silvagno. 2021. "Vitamin D and Beta-Glucans Synergically Stimulate Human Macrophage Activity" International Journal of Molecular Sciences 22, no. 9: 4869. https://doi.org/10.3390/ijms22094869
APA StyleBergandi, L., Apprato, G., & Silvagno, F. (2021). Vitamin D and Beta-Glucans Synergically Stimulate Human Macrophage Activity. International Journal of Molecular Sciences, 22(9), 4869. https://doi.org/10.3390/ijms22094869