
Matrix Metalloproteinases and Their Role in Mechanisms Underlying Effects of Quercetin on Heart Function in Aged Zucker Diabetic Fatty Rats

 ,
,
Abstract
1. Introduction
2. Results
2.1. Effect of Quercetin on Biometric and Biochemical Characteristics of Lean and Obese ZDF Rats
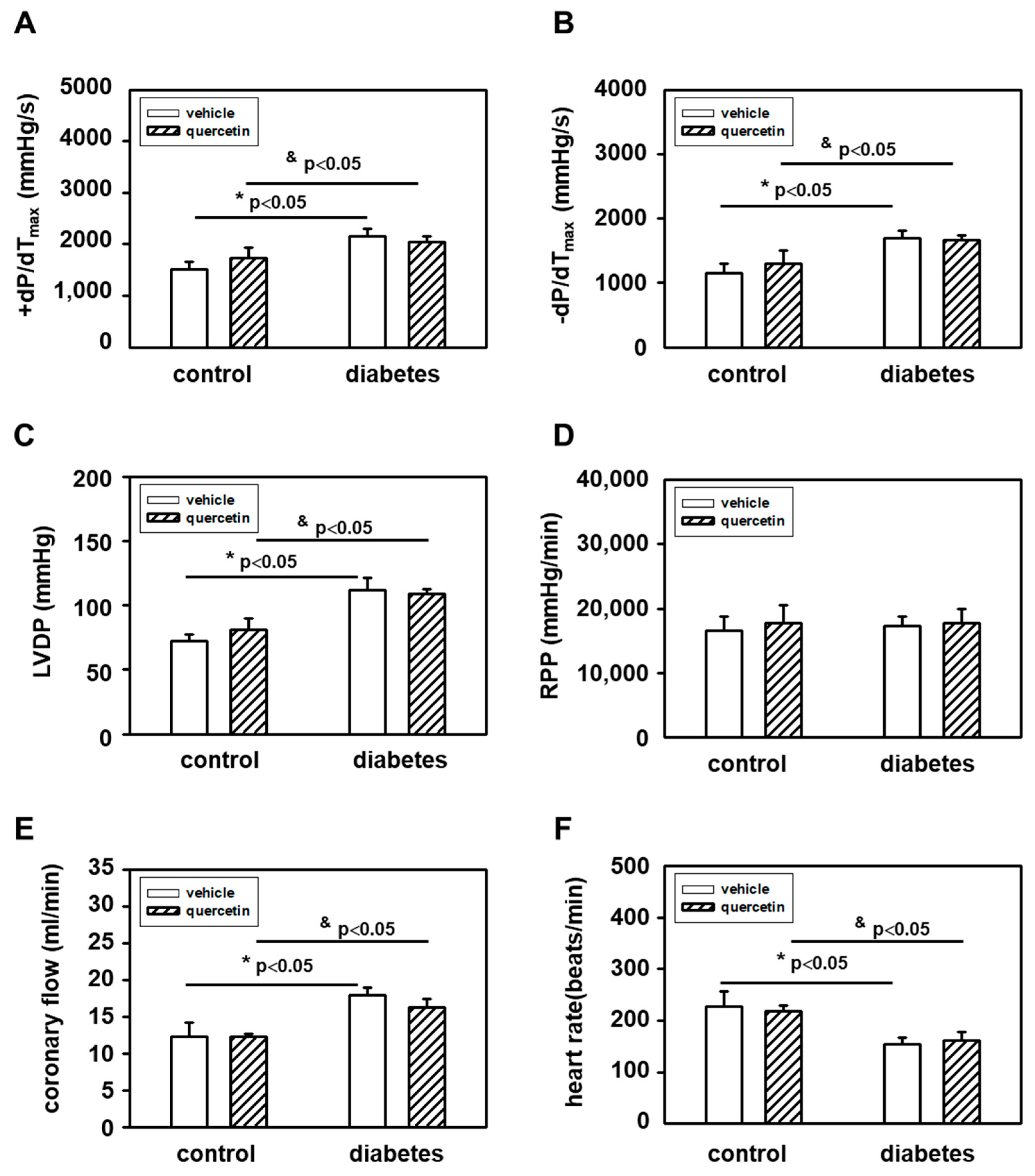
2.2. Effects of Quercetin on Functional Parameters of Isolated Langendorff-Perfused Hearts
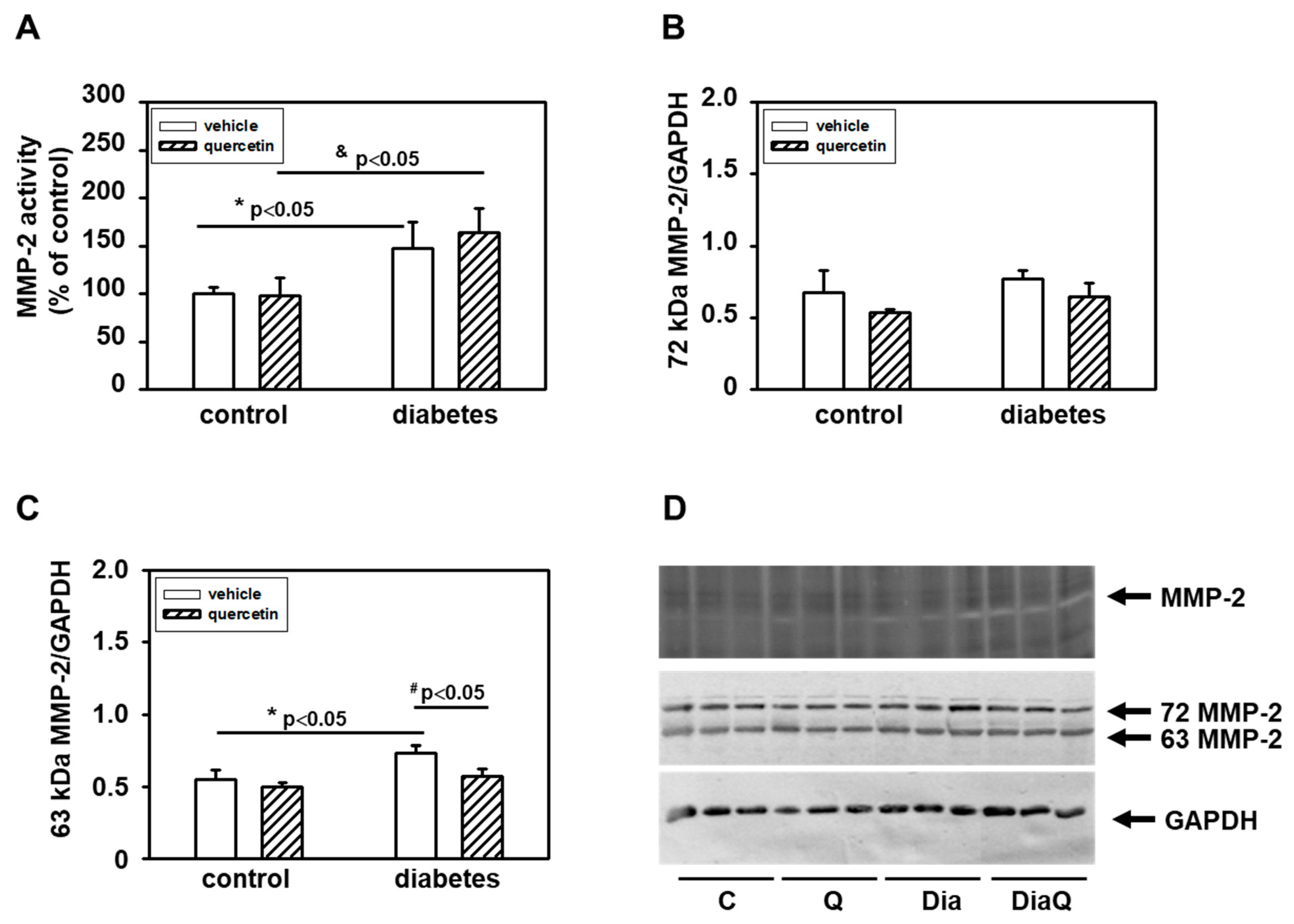
2.3. Effects of Quercetin on Modulation of Matrix Metalloproteinase-2 Activities and Protein Levels in Lean and Obese ZDF Rats
2.4. Effects of Quercetin on Matrix Metalloproteinase-9 and Tissue Inhibitor of Matrix Metalloproteinases-2 in Lean and Obese ZDF Rats
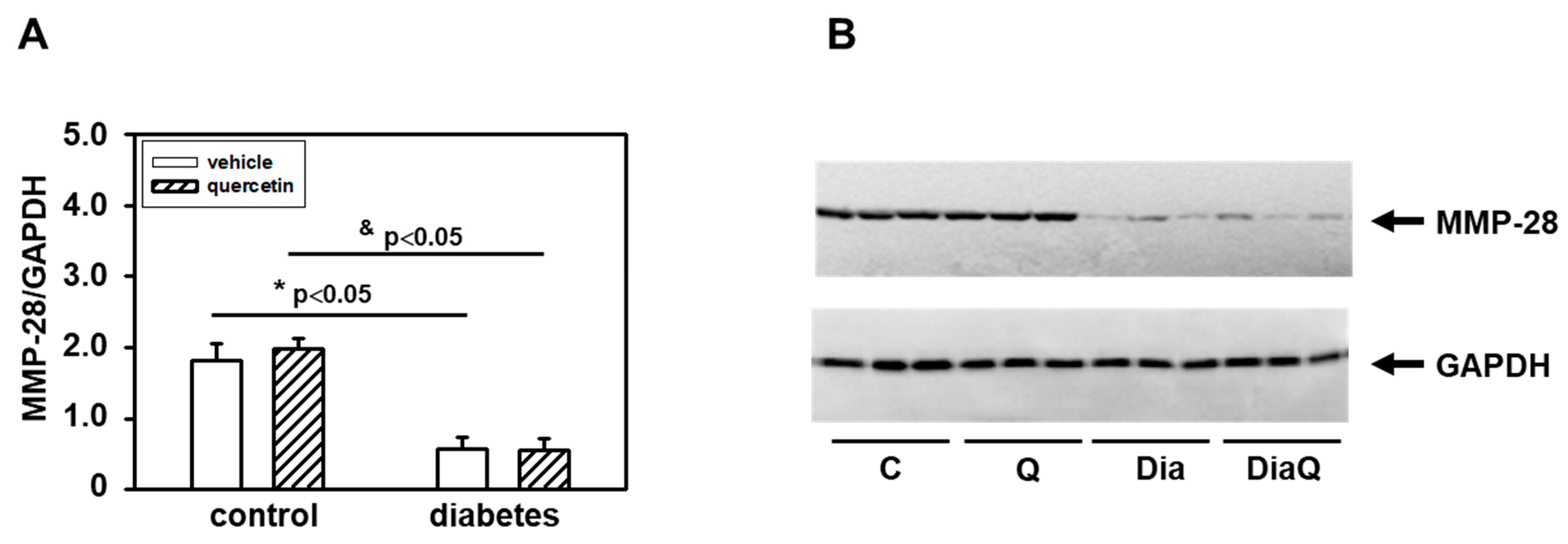
2.5. Effects of Quercetin on Matrix Metalloproteinase-28 in Lean and Obese ZDF Rats
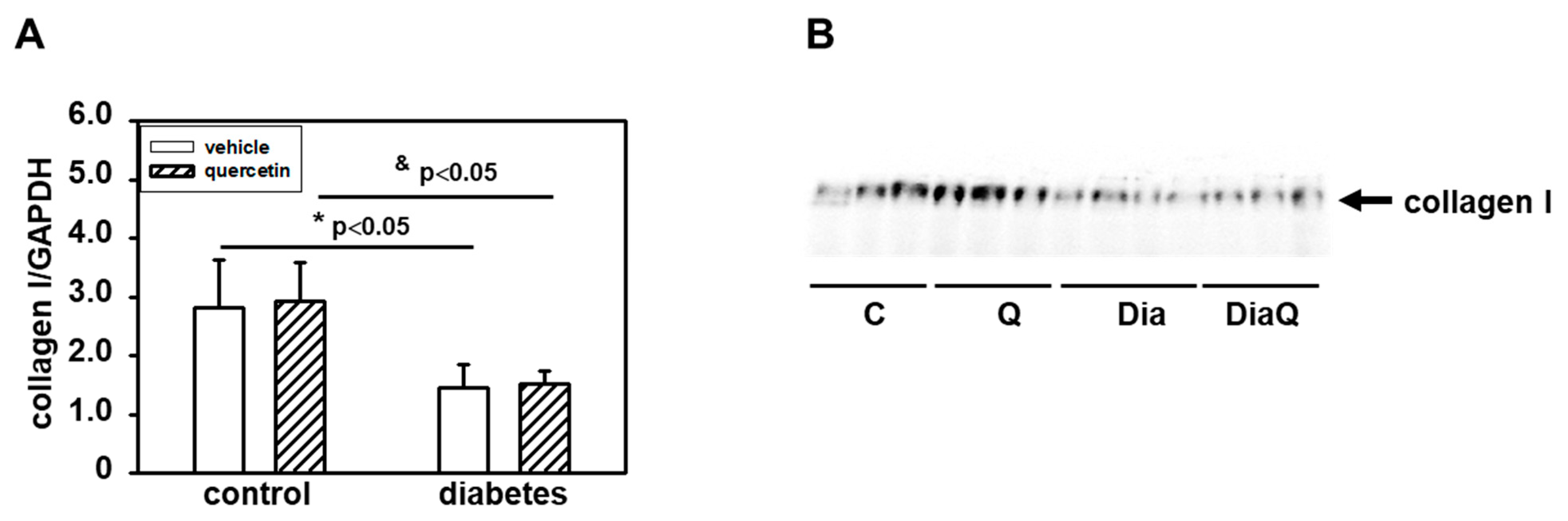
2.6. Effects of Quercetin on Collagen I Content in Lean and Obese ZDF Rats
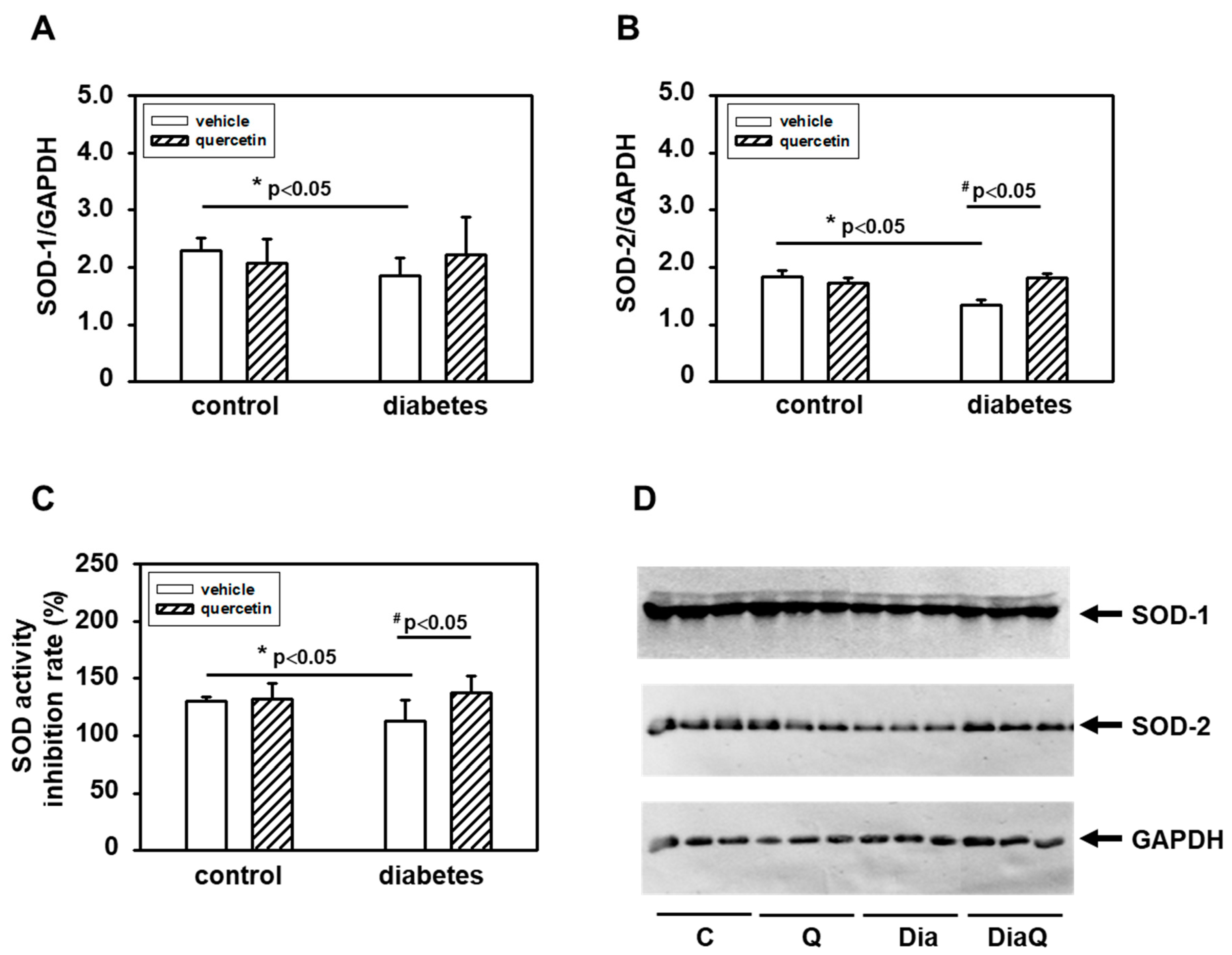
2.7. Quercetin Prevents Diabetes-induced Inhibition of Superoxide Dismutase
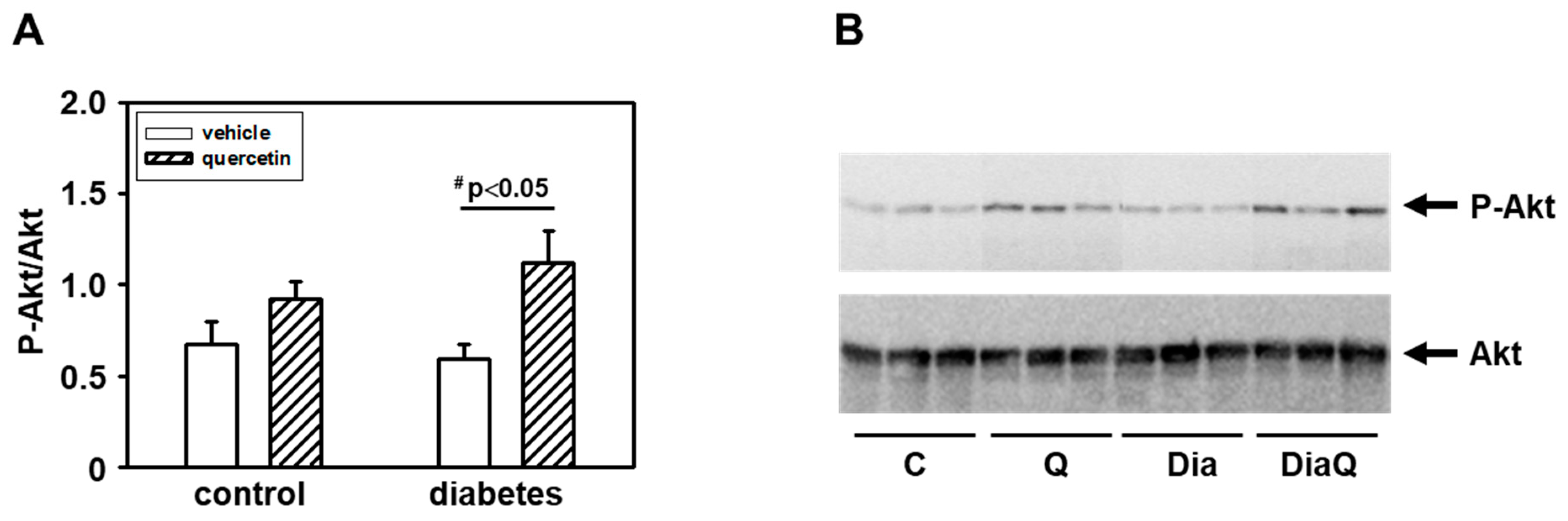
2.8. Quercetin Induced Akt Kinase Activation in Diabetic Rat Hearts
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Experimental Model
4.3. Assessment of Heart Functional Parameters in Langendorff-Perfused Hearts
4.4. Tissue Samples Collection
4.5. Preparation of Tissue Protein Fractions and Western Blot Analysis
4.6. Measurement of MMPs Activities by Gelatin Zymography
4.7. Determination of Superoxide Dismutase Activity
4.8. Statistical Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Schnurr, T.M.; Jakupović, H.; Carrasquilla, G.D.; Ängquist, L.; Grarup, N.; Sørensen, T.I.A.; Tjønneland, A.; Overvad, K.; Pedersen, O.; Hansen, T.; et al. Obesity, unfavourable lifestyle and genetic risk of type 2 diabetes: A case-cohort study. Diabetologia 2020, 63, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Kenchaiah, S.; Evans, J.C.; Levy, D.; Wilson, P.W.F.; Benjamin, E.J.; Larson, M.G.; Kannel, W.B.; Vasan, R.S. Obesity and the Risk of Heart Failure. N. Engl. J. Med. 2002, 347, 305–313. [Google Scholar] [CrossRef]
- Donahoe, S.M.; Stewart, G.C.; McCabe, C.H.; Mohanavelu, S.; Murphy, S.A.; Cannon, C.P.; Antman, E.M. Diabetes and Mortality Following Acute Coronary Syndromes. JAMA 2007, 298, 765. [Google Scholar] [CrossRef]
- Koek, H.L.; Soedamah-Muthu, S.S.; Kardaun, J.W.P.F.; Gevers, E.; de Bruin, A.; Reitsma, J.B.; Bots, M.L.; Grobbee, D.E. Short- and long-term mortality after acute myocardial infarction: Comparison of patients with and without diabetes mellitus. Eur. J. Epidemiol. 2007, 22, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Cavender, M.A.; Steg, P.G.; Smith, S.C.; Eagle, K.; Ohman, E.M.; Goto, S.; Kuder, J.; Im, K.; Wilson, P.W.F.; Bhatt, D.L. Impact of Diabetes Mellitus on Hospitalization for Heart Failure, Cardiovascular Events, and Death. Circulation 2015, 132, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Portik-Dobos, V.; Anstadt, M.P.; Hutchinson, J.; Bannan, M.; Ergul, A. Evidence for a Matrix Metalloproteinase Induction/Activation System in Arterial Vasculature and Decreased Synthesis and Activity in Diabetes. Diabetes 2002, 51, 3063–3068. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.K.; Hutchinson, J.R.; Sachidanandam, K.; Johnson, M.H.; Dorrance, A.M.; Stepp, D.W.; Fagan, S.C.; Ergul, A. Type 2 Diabetes Causes Remodeling of Cerebrovasculature via Differential Regulation of Matrix Metalloproteinases and Collagen Synthesis: Role of Endothelin-1. Diabetes 2005, 54, 2638–2644. [Google Scholar] [CrossRef]
- Banaszkiewicz, M.; Krzywonos-Zawadzka, A.; Olejnik, A.; Bil-Lula, I. Tissue Expression of Atrial and Ventricular Myosin Light Chains in the Mechanism of Adaptation to Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 8384. [Google Scholar] [CrossRef]
- Spinale, F.G. Myocardial Matrix Remodeling and the Matrix Metalloproteinases: Influence on Cardiac Form and Function. Physiol. Rev. 2007, 87, 1285–1342. [Google Scholar] [CrossRef]
- Lee, H.W.; Lee, S.J.; Lee, M.Y.; Park, M.W.; Kim, S.S.; Shin, N.; Lovett, D.H.; Bae, S.S.; Ahn, J.; Park, J.-S.; et al. Enhanced cardiac expression of two isoforms of matrix metalloproteinase-2 in experimental diabetes mellitus. PLoS ONE 2019, 14, e0221798. [Google Scholar] [CrossRef] [PubMed]
- Giannakos, E.; Vardali, E.; Bartekova, M.; Fogarassyova, M.; Barancik, M.; Radosinska, J. Changes in Activities of Circulating MMP-2 and MMP-9 in Patients Suffering From Heart Failure in Relation to Gender, Hypertension and Treatment: A Cross-Sectional Study. Physiol. Res. 2016, S149–S152. [Google Scholar] [CrossRef]
- Hlatky, M.A.; Ashley, E.; Quertermous, T.; Boothroyd, D.B.; Ridker, P.; Southwick, A.; Myers, R.M.; Iribarren, C.; Fortmann, S.P.; Go, A.S. Matrix metalloproteinase circulating levels, genetic polymorphisms, and susceptibility to acute myocardial infarction among patients with coronary artery disease. Am. Heart J. 2007, 154, 1043–1051. [Google Scholar] [CrossRef]
- Phatharajaree, W.; Phrommintikul, A.; Chattipakorn, N. Matrix metalloproteinases and myocardial infarction. Can. J. Cardiol. 2007, 23, 727–733. [Google Scholar] [CrossRef]
- Barteková, M.; Šimončíková, P.; Fogarassyová, M.; Ivanová, M.; Okruhlicová, Ľ.; Tribulová, N.; Dovinová, I.; Barančík, M. Quercetin Improves Postischemic Recovery of Heart Function in Doxorubicin-Treated Rats and Prevents Doxorubicin-Induced Matrix Metalloproteinase-2 Activation and Apoptosis Induction. Int. J. Mol. Sci. 2015, 16, 8168–8185. [Google Scholar] [CrossRef]
- Ivanová, M.; Dovinová, I.; Okruhlicová, Ľ.; Tribulová, N.; Šimončíková, P.; Barte-ková, M.; Vlkovičová, J.; Barančík, M. Chronic cardiotoxicity of doxorubicin involves activation of myocardial and circulating matrix metalloproteinases in rats. Acta Pharmacol. Sin. 2012, 33, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Camp, T.M.; Tyagi, S.C.; Senior, R.M.; Hayden, M.R.; Tyagi, S.C. Gelatinase B(MMP-9) an apoptotic factor in diabetic transgenic mice. Diabetologia 2003, 46, 1438–1445. [Google Scholar] [CrossRef] [PubMed]
- Yabluchanskiy, A.; Ma, Y.; Iyer, R.P.; Hall, M.E.; Lindsey, M.L. Matrix Metalloproteinase-9: Many Shades of Function in Cardiovascular Disease. Physiology 2013, 28, 391–403. [Google Scholar] [CrossRef]
- Iwasaka, T.; Takahashi, N.; Nakamura, S.; Sugiura, T.; Tarumi, N.; Kimura, Y.; Okubo, N.; Taniguchi, H.; Matsui, Y.; Inada, M. Residual Left Ventricular Pump Function After Acute Myocardial Infarction in NIDDM Patients. Diabetes Care 1992, 15, 1522–1526. [Google Scholar] [CrossRef]
- Uemura, S.; Matsushita, H.; Li, W.; Glassford, A.J.; Asagami, T.; Lee, K.-H.; Harrison, D.G.; Tsao, P.S. Diabetes Mellitus Enhances Vascular Matrix Metalloproteinase Activity. Circ. Res. 2001, 88, 1291–1298. [Google Scholar] [CrossRef]
- Annapurna, A.; Reddy, C.S.; Akondi, R.B.; Rao, S.R.C. Cardioprotective actions of two bioflavonoids, quercetin and rutin, in experimental myocardial infarction in both normal and streptozotocin-induced type I diabetic rats. J. Pharm. Pharmacol. 2009, 61, 1365–1374. [Google Scholar] [CrossRef]
- Jeong, S.-M.; Kang, M.-J.; Choi, H.-N.; Kim, J.-H.; Kim, J.-I. Quercetin ameliorates hyperglycemia and dyslipidemia and improves antioxidant status in type 2 diabetic db/db mice. Nutr. Res. Pract. 2012, 6, 201. [Google Scholar] [CrossRef]
- Pereira, T.; Pimenta, F.; Porto, M.; Baldo, M.; Campagnaro, B.; Gava, A.; Meyrelles, S.; Vasquez, E. Coadjuvants in the Diabetic Complications: Nutraceuticals and Drugs with Pleiotropic Effects. Int. J. Mol. Sci. 2016, 17, 1273. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [PubMed]
- Elbarbry, F.; Abdelkawy, K.; Moshirian, N.; Abdel-Megied, A.M. The Antihypertensive Effect of Quercetin in Young Spontaneously Hypertensive Rats; Role of Arachidonic Acid Metabolism. Int. J. Mol. Sci. 2020, 21, 6554. [Google Scholar] [CrossRef] [PubMed]
- Barteková, M.; Čarnická, S.; Pancza, D.; Ondrejčáková, M.; Breier, A.; Ravingerová, T. Acute treatment with polyphenol quercetin improves postischemic recovery of isolated perfused rat hearts after global ischemia. Can. J. Physiol. Pharmacol. 2010, 88, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Ferenczyova, K.; Kalocayova, B.; Bartekova, M. Potential Implications of Quercetin and its Derivatives in Cardioprotection. Int. J. Mol. Sci. 2020, 21, 1585. [Google Scholar] [CrossRef]
- Vessal, M.; Hemmati, M.; Vasei, M. Antidiabetic effects of quercetin in streptozocin-induced diabetic rats. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2003, 135, 357–364. [Google Scholar] [CrossRef]
- Marsh, S.A.; Powell, P.C.; Agarwal, A.; Dell’Italia, L.J.; Chatham, J.C. Cardiovascular dysfunction in Zucker obese and Zucker diabetic fatty rats: Role of hydronephrosis. Am. J. Physiol. Circ. Physiol. 2007, 293, H292–H298. [Google Scholar] [CrossRef] [PubMed]
- Daniels, A.; Linz, D.; van Bilsen, M.; Rütten, H.; Sadowski, T.; Ruf, S.; Juretschke, H.-P.; Neumann-Haefelin, C.; Munts, C.; van der Vusse, G.J.; et al. Long-term severe diabetes only leads to mild cardiac diastolic dysfunction in Zucker diabetic fatty rats. Eur. J. Heart Fail. 2012, 14, 193–201. [Google Scholar] [CrossRef]
- Borges, G.R.; de Oliveira, M.; Salgado, H.C.; Fazan, R. Myocardial performance in conscious streptozotocin diabetic rats. Cardiovasc. Diabetol. 2006, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, S.B.; Pælestik, K.B.; Johnsen, J.; Jespersen, N.R.; Pryds, K.; Hjortbak, M.V.; Jensen, R.V.; Bøtker, H.E. Impact of hyperglycemia on myocardial ischemia–reperfusion susceptibility and ischemic preconditioning in hearts from rats with type 2 diabetes. Cardiovasc. Diabetol. 2019, 18, 66. [Google Scholar] [CrossRef]
- Wang, P.; Chatham, J.C. Onset of diabetes in Zucker diabetic fatty (ZDF) rats leads to improved recovery of function after ischemia in the isolated perfused heart. Am. J. Physiol. Metab. 2004, 286, E725–E736. [Google Scholar] [CrossRef]
- Krebber, M.M.; van Dijk, C.G.M.; Vernooij, R.W.M.; Brandt, M.M.; Emter, C.A.; Rau, C.D.; Fledderus, J.O.; Duncker, D.J.; Verhaar, M.C.; Cheng, C.; et al. Matrix Metalloproteinases and Tissue Inhibitors of Metalloproteinases in Extracellular Matrix Remodeling during Left Ventricular Diastolic Dysfunction and Heart Failure with Preserved Ejection Fraction: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2020, 21, 6742. [Google Scholar] [CrossRef]
- Chan, B.Y.H.; Roczkowsky, A.; Cho, W.J.; Poirier, M.; Lee, T.Y.T.; Mahmud, Z.; Schulz, R. Junctophilin-2 is a target of matrix metalloproteinase-2 in myocardial ischemia–reperfusion injury. Basic Res. Cardiol. 2019, 114, 42. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhang, Q.; Pu, L.J.; Peng, W.H.; Yan, X.X.; Wang, L.J.; Chen, Q.J.; Zhu, Z.B.; Michel, J.-B.; Shen, W.F. Dysregulation of matrix metalloproteinases and their tissue inhibitors is related to abnormality of left ventricular geometry and function in streptozotocin-induced diabetic minipigs. Int. J. Exp. Pathol. 2008, 89, 125–137. [Google Scholar] [CrossRef]
- Gliozzi, M.; Malara, N.; Muscoli, S.; Mollace, V. The treatment of hyperuricemia. Int. J. Cardiol. 2016, 213, 23–27. [Google Scholar] [CrossRef]
- Scheepers, L.E.J.M.; Boonen, A.; Dagnelie, P.C.; Schram, M.T.; van der Kallen, C.J.H.; Henry, R.M.A.; Kroon, A.A.; Stehouwer, C.D.A.; Arts, I.C.W. Uric acid and blood pressure. J. Hypertens. 2017, 35, 1968–1975. [Google Scholar] [CrossRef]
- Bhole, V.; Choi, J.W.J.; Woo Kim, S.; de Vera, M.; Choi, H. Serum Uric Acid Levels and the Risk of Type 2 Diabetes: A Prospective Study. Am. J. Med. 2010, 123, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Gruszka, K.; Rajzer, M.; Drożdż, T.; Wojciechowska, W.; Pizoń, T.; Migacz-Gruszka, K.; Czarnecka, D. Selected matrix metalloproteinases activity and hypertension-mediated organ damage in relation to uric acid serum level. Cardiol. J. 2013. [Google Scholar] [CrossRef]
- Cancemi, P.; Aiello, A.; Accardi, G.; Caldarella, R.; Candore, G.; Caruso, C.; Ciaccio, M.; Cristaldi, L.; Di Gaudio, F.; Siino, V.; et al. The Role of Matrix Metalloproteinases (MMP-2 and MMP-9) in Ageing and Longevity: Focus on Sicilian Long-Living Individuals (LLIs). Mediat. Inflamm. 2020, 2020, 8635158. [Google Scholar] [CrossRef] [PubMed]
- Kanimozhi, S.; Bhavani, P.; Subramanian, P. Influence of the Flavonoid, Quercetin on Antioxidant Status, Lipid Peroxidation and Histopathological Changes in Hyperammonemic Rats. Indian J. Clin. Biochem. 2017, 32, 275–284. [Google Scholar] [CrossRef]
- Punithavathi, V.R.; Stanely Mainzen Prince, P. The cardioprotective effects of a combination of quercetin and α-tocopherol on isoproterenol-induced myocardial infarcted rats. J. Biochem. Mol. Toxicol. 2011, 25, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Halade, G.V.; Zhang, J.; Ramirez, T.A.; Levin, D.; Voorhees, A.; Jin, Y.-F.; Han, H.-C.; Manicone, A.M.; Lindsey, M.L. Matrix Metalloproteinase-28 Deletion Exacerbates Cardiac Dysfunction and Rupture After Myocardial Infarction in Mice by Inhibiting M2 Macrophage Activation. Circ. Res. 2013, 112, 675–688. [Google Scholar] [CrossRef]
- Ge, Z.; Wang, B.; Hu, J.; Sun, J.; Wang, S.; Chen, X.; Meng, S.; Liu, L.; Cheng, Z. IRAK3 gene silencing prevents cardiac rupture and ventricular remodeling through negative regulation of the NF-κB signaling pathway in a mouse model of acute myocardial infarction. J. Cell. Physiol. 2019, 234, 11722–11733. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Jiang, T.; Li, Y.; Kong, L. Endostatin attenuates heart failure via inhibiting reactive oxygen species in myocardial infarction rats. Biosci. Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Chiao, Y.A.; Zhang, J.; Manicone, A.M.; Jin, Y.-F.; Lindsey, M.L. Matrix Metalloproteinase-28 Deletion Amplifies Inflammatory and Extracellular Matrix Responses to Cardiac Aging. Microsc. Microanal. 2012, 18, 81–90. [Google Scholar] [CrossRef]
- Chang, J.-H.; Jin, M.-M.; Liu, J.-T. Dexmedetomidine pretreatment protects the heart against apoptosis in ischemia/reperfusion injury in diabetic rats by activating PI3K/Akt signaling in vivo and in vitro. Biomed. Pharmacother. 2020, 127, 110188. [Google Scholar] [CrossRef]
- Maciel, L.; de Oliveira, D.F.; Mesquita, F.; Souza, H.A.D.S.; Oliveira, L.; Christie, M.L.A.; Palhano, F.L.; Campos de Carvalho, A.C.; Nascimento, J.H.M.; Foguel, D. New Cardiomyokine Reduces Myocardial Ischemia/Reperfusion Injury by PI3K-AKT Pathway Via a Putative KDEL-Receptor Binding. J. Am. Heart Assoc. 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity: A Critical Component in Human Diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Lu, K.; Zhao, Z.; Wang, B.; Liu, H.; Zhang, S.; Liao, J.; Zeng, Y.; Dong, Q.; Zhao, N.; et al. Blockade of Transient Receptor Potential Vanilloid 4 Enhances Antioxidation after Myocardial Ischemia/Reperfusion. Oxidative Med. Cell. Longev. 2019, 2019, 7283683. [Google Scholar] [CrossRef] [PubMed]
- Ferenczyova, K.; Kalocayova, B.; Kindernay, L.; Jelemensky, M.; Balis, P.; Berenyiova, A.; Zemancikova, A.; Farkasova, V.; Sykora, M.; Tothova, L.; et al. Quercetin Exerts Age-Dependent Beneficial Effects on Blood Pressure and Vascular Function, But Is Inefficient in Preventing Myocardial Ischemia-Reperfusion Injury in Zucker Diabetic Fatty Rats. Molecules 2020, 25, 187. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | C | Q | Dia | DiaQ |
|---|---|---|---|---|
| BW (g) | 406 ± 10 | 413 ± 8 | 520 ± 25 * | 514 ± 24 & |
| Glucose (mmol/L) | 6.1 ± 0.1 | 5.9 ± 0.2 | 16.9 ± 1.6 * | 18.48 ± 2.1 & |
| TAG (mmol/L) | 0.24 ± 0.05 | 0.18 ± 0.03 | 3.66 ± 0.41 * | 3.67 ± 0.39 & |
| Cholesterol (mmol/L) | 2.85 ± 0.11 | 2.81 ± 0.08 | 4.86 ± 0.41 * | 4.79 ± 0.25 & |
| HDL-cholesterol (mmol/L) | 1.36 ± 0.04 | 1.37 ± 0.04 | 2.36 ± 0.13 * | 2.37 ± 0.07 & |
| LDL-cholesterol (mmol/L) | 0.82 ± 0.04 | 0.81 ± 0.02 | 1.05 ± 0.08 * | 0.91 ± 0.07 & |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boťanská, B.; Barteková, M.; Ferenczyová, K.; Fogarassyová, M.; Kindernay, L.; Barančík, M. Matrix Metalloproteinases and Their Role in Mechanisms Underlying Effects of Quercetin on Heart Function in Aged Zucker Diabetic Fatty Rats. Int. J. Mol. Sci. 2021, 22, 4457. https://doi.org/10.3390/ijms22094457
Boťanská B, Barteková M, Ferenczyová K, Fogarassyová M, Kindernay L, Barančík M. Matrix Metalloproteinases and Their Role in Mechanisms Underlying Effects of Quercetin on Heart Function in Aged Zucker Diabetic Fatty Rats. International Journal of Molecular Sciences. 2021; 22(9):4457. https://doi.org/10.3390/ijms22094457
Chicago/Turabian StyleBoťanská, Barbora, Monika Barteková, Kristína Ferenczyová, Mária Fogarassyová, Lucia Kindernay, and Miroslav Barančík. 2021. "Matrix Metalloproteinases and Their Role in Mechanisms Underlying Effects of Quercetin on Heart Function in Aged Zucker Diabetic Fatty Rats" International Journal of Molecular Sciences 22, no. 9: 4457. https://doi.org/10.3390/ijms22094457
APA StyleBoťanská, B., Barteková, M., Ferenczyová, K., Fogarassyová, M., Kindernay, L., & Barančík, M. (2021). Matrix Metalloproteinases and Their Role in Mechanisms Underlying Effects of Quercetin on Heart Function in Aged Zucker Diabetic Fatty Rats. International Journal of Molecular Sciences, 22(9), 4457. https://doi.org/10.3390/ijms22094457