
Quantitative Proteomics Reveals the Dynamic Regulation of the Tomato Proteome in Response to Phytophthora infestans

 ,
, 
Abstract
1. Introduction
2. Results and Discussion
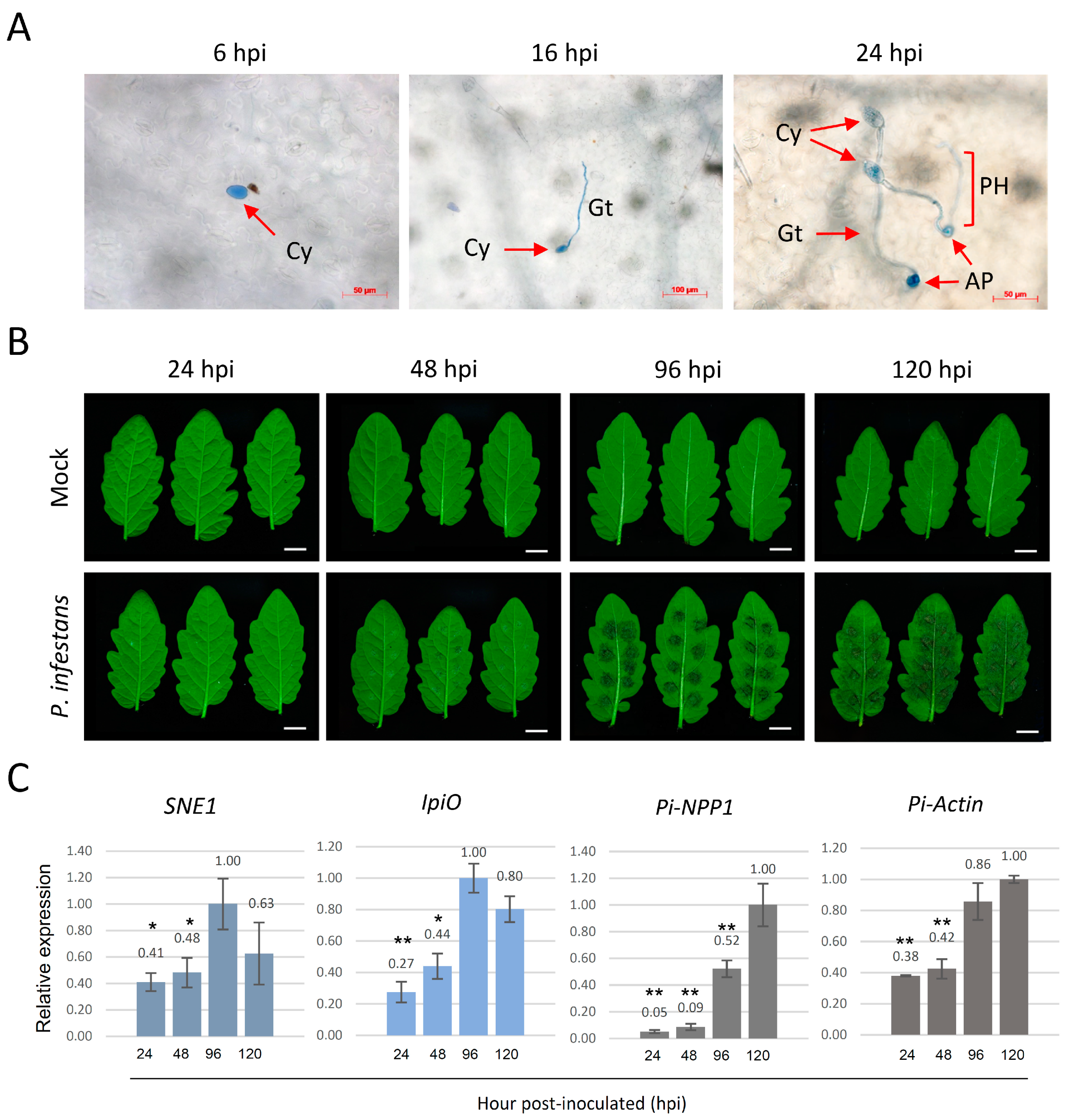
2.1. P. infestans Pathogenesis Assessment in Tomato Leaves
2.2. Proteomics Analysis and Quantification Using the DIA Approach
2.3. Functional Classification of the Proteins Regulated by P. infestans
2.4. Changes Associated with Direct Defense
2.5. Changes Associated with Immune Regulation
2.6. Changes Associated with the Regulation of Phytohormones
2.7. Changes Associated with Reactive Oxygen Species and Oxidation–Reduction Reactions
2.8. Differentially Regulated Proteins Involved in Carbohydrate and Energy Metabolism
2.9. Changes Associated with Secondary Metabolites
2.10. Novel P. infestans-Regulated Tomato Reponses Revealed by Time-Lapse Proteomics Studies
3. Materials and Methods
3.1. Plant Material, Growth Conditions and P. infestans Inoculation
3.2. Observation of P. infestans Growth by Trypan Blue Staining and Microscope
3.3. RNA Extraction and Marker Gene Expression Analysis of P. infestans-Inoculated Tomato Leaf
3.4. Sample Preparation for Proteome Analysis
3.5. Liquid Chromatography-Mass Spectrometry Analysis
3.6. MS Data Analysis
3.7. Quantitation Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| DIA | data-independent acquisition |
| DDA | data-dependent acquisition |
| ET | ethylene |
| ETI | effector-triggered immunity |
| ETS | effector-triggered susceptibility |
| HR | hypersensitive response |
| JA | jasmonic acid |
| LB | late blight |
| PAMP | pathogen-associated molecular pattern |
| PCD | programmed cell death |
| PTI | PAMP-triggered immunity |
| ROS | Reactive-oxidative species |
| SA | salicylic acid |
References
- Schoina, C.; Govers, F. The Oomycete Phytophthora Infestans, the Irish Potato Famine Pathogen. In Principles of Plant-Microbe Interactions; Springer: Cham, Switzerland, 2015; pp. 371–378. [Google Scholar]
- Nowicki, M.; Fooled, M.R.; Nowakowska, M.; Kozik, E.U. Potato and Tomato Late Blight Caused by Phytophthora Infestans: An Overview of Pathology and Resistance Breeding. Plant Dis. 2012, 96, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, A.Y.; da Silva, D.J.H.; Carneiro, P.C.S.; Mizubuti, E.S.G. The Inheritance of Late Blight Resistance Derived from Solanum Habrochaites. Crop. Breed. Appl. Biot. 2012, 12, 199–205. [Google Scholar] [CrossRef]
- Kamoun, S.; Smart, C.D. Late Blight of Potato and Tomato in the Genomics Era. Plant Dis. 2005, 89, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Hausbeck, M.K.; Lamour, K.H. Phytophthora Capsici on Vegetable Crops: Research Progress and Management Challenges. Plant Dis. 2004, 88, 1292–1303. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Panstruga, R.; Dodds, P.N. Terrific Protein Traffic: The Mystery of Effector Protein Delivery by Filamentous Plant Pathogens. Science 2009, 324, 748–750. [Google Scholar] [CrossRef]
- Koeck, M.; Hardham, A.R.; Dodds, P.N. The Role of Effectors of Biotrophic and Hemibiotrophic Fungi in Infection. Cellular Microbiol. 2011, 13, 1849–1857. [Google Scholar] [CrossRef]
- Wang, B.L.; Liu, J.; Tian, Z.D.; Song, B.T.; Xie, C.H. Monitoring the Expression Patterns of Potato Genes Associated with Quantitative Resistance to Late Blight During Phytophthora Infestans Infection Using Cdna Microarrays. Plant Sci. 2005, 169, 1155–1167. [Google Scholar] [CrossRef]
- Cai, G.; Restrepo, S.; Myers, K.; Zuluaga, P.; Danies, G.; Smart, C.; Fry, W. Gene Profiling in Partially Resistant and Susceptible near-Isogenic Tomatoes in Response to Late Blight in the Field. Mol. Plant Pathol. 2013, 14, 171–184. [Google Scholar] [CrossRef]
- Zuluaga, A.P.; Vega-Arreguin, J.C.; Fei, Z.; Matas, A.J.; Patev, S.; Fry, W.E.; Rose, J.K. Analysis of the Tomato Leaf Transcriptome During Successive Hemibiotrophic Stages of a Compatible Interaction with the Oomycete Pathogen Phytophthora Infestans. Mol. Plant Pathol. 2016, 17, 42–54. [Google Scholar] [CrossRef]
- Larsen, M.K.; Jorgensen, M.M.; Bennike, T.B.; Stensballe, A. Time-Course Investigation of Phytophthora Infestans Infection of Potato Leaf from Three Cultivars by Quantitative Proteomics. Data Brief. 2016, 6, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Alexandersson, E.; Sandin, M.; Resjo, S.; Lenman, M.; Hedley, P.; Levander, F.; Andreasson, E. Quantitative Proteomics and Transcriptomics of Potato in Response to Phytophthora Infestans in Compatible and Incompatible Interactions. BMC Genom. 2014, 15, 497. [Google Scholar] [CrossRef]
- Xiao, C.; Gao, J.; Zhang, Y.; Wang, Z.; Zhang, D.; Chen, Q.; Ye, X.; Xu, Y.; Yang, G.; Yan, L.; et al. Quantitative Proteomics of Potato Leaves Infected with Phytophthora Infestans Provides Insights into Coordinated and Altered Protein Expression During Early and Late Disease Stages. Int. J. Mol. Sci. 2019, 20, 136. [Google Scholar] [CrossRef]
- Laurindo, B.; Laurindo, R.; Fontes, P.; Vital, C.; Delazari, F.; Baracat-Pereira, M.; da Silva, D. Comparative Proteomics Reveals Set of Oxidative Stress and Thaumatin-Like Proteins Associated with Resistance to Late Blight of Tomato. Am. J. Plant Sci. 2018, 9, 789–816. [Google Scholar] [CrossRef]
- Kelley, B.S.; Lee, S.J.; Damasceno, C.M.; Chakravarthy, S.; Kim, B.D.; Martin, G.B.; Rose, J.K. A Secreted Effector Protein (Sne1) from Phytophthora Infestans Is a Broadly Acting Suppressor of Programmed Cell Death. Plant J. 2010, 62, 357–366. [Google Scholar] [CrossRef] [PubMed]
- van West, P.; de Jong, A.J.; Judelson, H.S.; Emons, A.M.C.; Govers, F. The Ipio Gene of Phytophthora Infestans Is Highly Expressed in Invading Hyphae During Infection. Fungal Genet. Biol. 1998, 23, 126–138. [Google Scholar] [CrossRef]
- Kanneganti, T.-D.; Huitema, E.; Cakir, C.; Kamoun, S. Synergistic Interactions of the Plant Cell Death Pathways Induced by Phytophthora Infestans Nep1-Like Protein Pinpp1. 1 and Inf1 Elicitin. Mol. Plant-Microbe Interact. 2006, 19, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Bartnicki-Garcia, S. Cell Wall Chemistry, Morphogenesis, and Taxonomy of Fungi. Annu. Rev. Microbiol. 1968, 22, 87–108. [Google Scholar] [CrossRef]
- Salzman, R.A.; Koiwa, H.; Ibeas, J.I.; Pardo, J.M.; Hasegawa, P.M.; Bressan, R.A. Inorganic Cations Mediate Plant Pr5 Protein Antifungal Activity through Fungal Mnn1- and Mnn4-Regulated Cell Surface Glycans. Mol. Plant Microbe Interact. 2004, 17, 780–788. [Google Scholar] [CrossRef]
- Batalia, M.A.; Monzingo, A.F.; Ernst, S.; Roberts, W.; Robertus, J.D. The Crystal Structure of the Antifungal Protein Zeamatin, a Member of the Thaumatin-Like, PR-5 Protein Family. Nat. Struct. Biol. 1996, 3, 19–23. [Google Scholar] [CrossRef]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-Related Proteins and Peptides as Promising Tools for Engineering Plants with Multiple Stress Tolerance. Microbiol. Res. 2018, 212, 29–37. [Google Scholar] [CrossRef]
- Bantignies, B.; Seguin, J.; Muzac, I.; Dedaldechamp, F.; Gulick, P.; Ibrahim, R. Direct Evidence for Ribonucleolytic Activity of a PR-10-Like Protein from White Lupin Roots. Plant Mol. Biol. 2000, 42, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.J.; Lu, S.; Xu, Y.H.; Wang, J.W.; Chen, X.Y. A Cotton Cdna (Gapr-10) Encoding a Pathogenesis-Related 10 Protein with in Vitro Ribonuclease Activity. Plant Sci. 2002, 162, 629–636. [Google Scholar] [CrossRef]
- Andrade, L.B.; Oliveira, A.S.; Ribeiro, J.K.; Kiyota, S.; Vasconcelos, I.M.; de Oliveira, J.T.; de Sales, M.P. Effects of a Novel Pathogenesis-Related Class 10 (PR-10) Protein from Crotalaria Pallida Roots with Papain Inhibitory Activity against Root-Knot Nematode Meloidogyne Incognita. J. Agric. Food Chem. 2010, 58, 4145–4152. [Google Scholar] [CrossRef]
- Guevara-Morato, M.A.; de Lacoba, M.G.; Garcia-Luque, I.; Serra, M.T. Characterization of a Pathogenesis-Related Protein 4 (PR-4) Induced in Capsicum Chinense L3 Plants with Dual Rnase and Dnase Activities. J. Exp. Bot. 2010, 61, 3259–3271. [Google Scholar] [CrossRef]
- Klarzynski, O.; Plesse, B.; Joubert, J.M.; Yvin, J.C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear Beta-1,3 Glucans Are Elicitors of Defense Responses in Tobacco. Plant Physiol. 2000, 124, 1027–1038. [Google Scholar] [CrossRef]
- Chen, Y.L.; Lee, C.Y.; Cheng, K.T.; Chang, W.H.; Huang, R.N.; Nam, H.G.; Chen, Y.R. Quantitative Peptidomics Study Reveals That a Wound-Induced Peptide from PR-1 Regulates Immune Signaling in Tomato. Plant Cell 2014, 26, 4135–4148. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Fan, K.T.; Hung, S.C.; Chen, Y.R. The Role of Peptides Cleaved from Protein Precursors in Eliciting Plant Stress Reactions. New Phytol. 2020, 225, 2267–2282. [Google Scholar] [CrossRef] [PubMed]
- Gamir, J.; Darwiche, R.; Van’t Hof, P.; Choudhary, V.; Stumpe, M.; Schneiter, R.; Mauch, F. The Sterol-Binding Activity of Pathogenesis-Related Protein 1 Reveals the Mode of Action of an Antimicrobial Protein. Plant J. 2017, 89, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Kruger, J.; Thomas, C.M.; Golstein, C.; Dixon, M.S.; Smoker, M.; Tang, S.K.; Mulder, L.; Jones, J.D.G. A Tomato Cysteine Protease Required for Cf-2-Dependent Disease Resistance and Suppression of Autonecrosis. Science 2002, 296, 744–747. [Google Scholar] [CrossRef]
- Tian, M.; Win, J.; Song, J.; van der Hoorn, R.; van der Knaap, E.; Kamoun, S. A Phytophthora Infestans Cystatin-Like Protein Targets a Novel Tomato Papain-Like Apoplastic Protease. Plant Physiol. 2007, 143, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Cheung, M.-Y.; Li, M.-W.; Fu, Y.; Sun, Z.; Sun, S.-M.; Lam, H.-M. Rice Hypersensitive Induced Reaction Protein 1 (Oshir1) Associates with Plasma Membrane and Triggers Hypersensitive Cell Death. BMC Plant Biol. 2010, 10, 290. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Tsuda, K.; Nguyen le, V.; Wang, X.; Lin, J.; Murphy, A.S.; Glazebrook, J.; Thordal-Christensen, H.; Katagiri, F. Physical Association of Arabidopsis Hypersensitive Induced Reaction Proteins (Hirs) with the Immune Receptor Rps2. J. Biol. Chem. 2011, 286, 31297–31307. [Google Scholar] [CrossRef] [PubMed]
- Lampl, N.; Alkan, N.; Davydov, O.; Fluhr, R. Set-Point Control of Rd21 Protease Activity by Atserpin1 Controls Cell Death in Arabidopsis. Plant J. 2013, 74, 498–510. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, K.; Cheng, Q.; Kong, D.; Zhang, X.; Wang, Z.; Wang, Q.; Xie, Q.; Yan, J.; Chu, J.; et al. Cysteine Protease Rd21a Regulated by E3 Ligase Sinat4 Is Required for Drought-Induced Resistance to Pseudomonas syringae in Arabidopsis. J. Exp. Bot. 2020, 71, 5562–5576. [Google Scholar] [CrossRef]
- DiMario, R.J.; Clayton, H.; Mukherjee, A.; Ludwig, M.; Moroney, J.V. Plant Carbonic Anhydrases: Structures, Locations, Evolution, and Physiological Roles. Mol. Plant 2017, 10, 30–46. [Google Scholar] [CrossRef]
- Zhou, Y.; Vroegop-Vos, I.A.; Van Dijken, A.J.H.; Van der Does, D.; Zipfel, C.; Pieterse, C.M.J.; Van Wees, S.C.M. Carbonic Anhydrases Ca1 and Ca4 Function in Atmospheric Co2-Modulated Disease Resistance. Planta 2020, 251, 75. [Google Scholar] [CrossRef]
- Slaymaker, D.H.; Navarre, D.A.; Clark, D.; del Pozo, O.; Martin, G.B.; Klessig, D.F. The Tobacco Salicylic Acid-Binding Protein 3 (Sabp3) Is the Chloroplast Carbonic Anhydrase, Which Exhibits Antioxidant Activity and Plays a Role in the Hypersensitive Defense Response. Proc. Natl. Acad. Sci. USA 2002, 99, 11640–11645. [Google Scholar] [CrossRef]
- Restrepo, S.; Myers, K.L.; del Pozo, O.; Martin, G.B.; Hart, A.L.; Buell, C.R.; Fry, W.E.; Smart, C.D. Gene Profiling of a Compatible Interaction between Phytophthora Infestans and Solanum Tuberosum Suggests a Role for Carbonic Anhydrase. Mol. Plant-Microbe Interact. 2005, 18, 913–922. [Google Scholar] [CrossRef]
- Wirthmueller, L.; Zhang, Y.; Jones, J.D.; Parker, J.E. Nuclear Accumulation of the Arabidopsis Immune Receptor Rps4 Is Necessary for Triggering Eds1-Dependent Defense. Curr. Biol. 2007, 17, 2023–2029. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, Eds1 and NPR1/Nim1 Like Genes Are Required for N-Mediated Resistance to Tobacco Mosaic Virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef]
- Hu, G.S.; deHart, A.K.A.; Li, Y.S.; Ustach, C.; Handley, V.; Navarre, R.; Hwang, C.F.; Aegerter, B.J.; Williamson, V.M.; Baker, B. Eds1 in Tomato Is Required for Resistance Mediated by Tir-Class R Genes and the Receptor-Like R Gene Ve. Plant J. 2005, 42, 376–391. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.E.; Holub, E.B.; Frost, L.N.; Falk, A.; Gunn, N.D.; Daniels, M.J. Characterization of Eds1, a Mutation in Arabidopsis Suppressing Resistance to Peronospora Parasitica Specified by Several Different Rpp Genes. Plant Cell 1996, 8, 2033–2046. [Google Scholar] [PubMed]
- Heidrich, K.; Wirthmueller, L.; Tasset, C.; Pouzet, C.; Deslandes, L.; Parker, J.E. Arabidopsis Eds1 Connects Pathogen Effector Recognition to Cell Compartment-Specific Immune Responses. Science 2011, 334, 1401–1404. [Google Scholar] [CrossRef]
- Glazebrook, J.; Rogers, E.E.; Ausubel, F.M. Isolation of Arabidopsis Mutants with Enhanced Disease Susceptibility by Direct Screening. Genetics 1996, 143, 973–982. [Google Scholar] [CrossRef]
- Cui, H.; Gobbato, E.; Kracher, B.; Qiu, J.; Bautor, J.; Parker, J.E. A Core Function of Eds1 with Pad4 Is to Protect the Salicylic Acid Defense Sector in Arabidopsis Immunity. New Phytol. 2017, 213, 1802–1817. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Brader, G.; Palva, E.T. Kunitz Trypsin Inhibitor: An Antagonist of Cell Death Triggered by Phytopathogens and Fumonisin B1 in Arabidopsis. Mol. Plant 2008, 1, 482–495. [Google Scholar] [CrossRef]
- Arnaiz, A.; Talavera-Mateo, L.; Gonzalez-Melendi, P.; Martinez, M.; Diaz, I.; Santamaria, M.E. Arabidopsis Kunitz Trypsin Inhibitors in Defense against Spider Mites. Front. Plant Sci. 2018, 9, 986. [Google Scholar] [CrossRef]
- Heitz, T.; Bergey, D.R.; Ryan, C.A. A Gene Encoding a Chloroplast-Targeted Lipoxygenase in Tomato Leaves Is Transiently Induced by Wounding, Systemin, and Methyl Jasmonate. Plant Physiol. 1997, 114, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Thilmony, R.; Bender, C.L.; Schaller, A.; He, S.Y.; Howe, G.A. Virulence Systems of Pseudomonas syringae pv. Tomato Promote Bacterial Speck Disease in Tomato by Targeting the Jasmonate Signaling Pathway. Plant J. 2003, 36, 485–499. [Google Scholar] [CrossRef]
- Pauwels, L.; Barbero, G.F.; Geerinck, J.; Tilleman, S.; Grunewald, W.; Perez, A.C.; Chico, J.M.; Vanden Bossche, R.; Sewell, J.; Gil, E.; et al. Ninja Connects the Co-Repressor Topless to Jasmonate Signalling. Nature 2010, 464, 788–U169. [Google Scholar] [CrossRef]
- Causier, B.; Ashworth, M.; Guo, W.; Davies, B. The Topless Interactome: A Framework for Gene Repression in Arabidopsis. Plant Physiol. 2012, 158, 423–438. [Google Scholar] [CrossRef]
- Harvey, S.; Kumari, P.; Lapin, D.; Griebel, T.; Hickman, R.; Guo, W.; Zhang, R.; Parker, J.E.; Beynon, J.; Denby, K. Downy Mildew Effector Harxl21 Interacts with the Transcriptional Repressor Topless to Promote Pathogen Susceptibility. bioRxiv 2020, 16, e1008835. [Google Scholar] [CrossRef]
- Mochizuki, N.; Brusslan, J.A.; Larkin, R.; Nagatani, A.; Chory, J. Arabidopsis Genomes Uncoupled 5 (Gun5) Mutant Reveals the Involvement of Mg-Chelatase H Subunit in Plastid-to-Nucleus Signal Transduction. Proc. Natl. Acad. Sci. USA 2001, 98, 2053–2058. [Google Scholar] [CrossRef]
- Ibata, H.; Nagatani, A.; Mochizuki, N. Chlh/Gun5 Function in Tetrapyrrole Metabolism Is Correlated with Plastid Signaling but Not Aba Responses in Guard Cells. Front. Plant Sci. 2016, 7, 1650. [Google Scholar] [CrossRef]
- Shang, Y.; Yan, L.; Liu, Z.Q.; Cao, Z.; Mei, C.; Xin, Q.; Wu, F.Q.; Wang, X.F.; Du, S.Y.; Jiang, T.; et al. The Mg-Chelatase H Subunit of Arabidopsis Antagonizes a Group of Wrky Transcription Repressors to Relieve Aba-Responsive Genes of Inhibition. Plant Cell 2010, 22, 1909–1935. [Google Scholar] [CrossRef] [PubMed]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of Hormone Signaling Networks in Plant Defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef] [PubMed]
- Sagi, M.; Davydov, O.; Orazova, S.; Yesbergenova, Z.; Ophir, R.; Stratmann, J.W.; Fluhr, R. Plant Respiratory Burst Oxidase Homologs Impinge on Wound Responsiveness and Development in Lycopersicon Esculentum. Plant Cell 2004, 16, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kaur, A.; Chattopadhyay, B.; Bachhawat, A.K. Defining the Cytosolic Pathway of Glutathione Degradation in Arabidopsis Thaliana: Role of the Chac/Gcg Family of Gamma-Glutamyl Cyclotransferases as Glutathione-Degrading Enzymes and Atlap1 as the Cys-Gly Peptidase. Biochem. J. 2015, 468, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.M.; Senthil-Kumar, M.; Tzin, V.; Mysore, K. Regulation of Primary Plant Metabolism during Plant-Pathogen Interactions and Its Contribution to Plant Defense. Front. Plant Sci. 2014, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.; Wang, L. The Phenylpropanoid Pathway and Plant Defence—A Genomics Perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- McManus, J.; Cheng, Z.; Vogel, C. Next-Generation Analysis of Gene Expression Regulation--Comparing the Roles of Synthesis and Degradation. Mol. Biosyst. 2015, 11, 2680–2689. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on Mrna Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Tsuzuki, T.; Takahashi, K.; Inoue, S.; Okigaki, Y.; Tomiyama, M.; Hossain, M.A.; Shimazaki, K.; Murata, Y.; Kinoshita, T. Mg-Chelatase H Subunit Affects Aba Signaling in Stomatal Guard Cells, but Is Not an Aba Receptor in Arabidopsis Thaliana. J. Plant Res. 2011, 124, 527–538. [Google Scholar] [CrossRef]
- Zhang, D.-P.; Wu, Z.-Y.; Li, X.-Y.; Zhao, Z.-X. Purification and Identification of a 42-Kilodalton Abscisic Acid-Specific-Binding Protein from Epidermis of Broad Bean Leaves. Plant Physiol. 2002, 128, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.Q.; Xin, Q.; Cao, Z.; Liu, Z.Q.; Du, S.Y.; Mei, C.; Zhao, C.X.; Wang, X.F.; Shang, Y.; Jiang, T.; et al. The Magnesium-Chelatase H Subunit Binds Abscisic Acid and Functions in Abscisic Acid Signaling: New Evidence in Arabidopsis. Plant Physiol. 2009, 150, 1940–1954. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.Y.; Wang, X.F.; Wu, F.Q.; Du, S.Y.; Cao, Z.; Shang, Y.; Wang, X.L.; Peng, C.C.; Yu, X.C.; Zhu, S.Y.; et al. The Mg-Chelatase H Subunit Is an Abscisic Acid Receptor. Nature 2006, 443, 823–826. [Google Scholar] [CrossRef]
- Muller, A.H.; Hansson, M. The Barley Magnesium Chelatase 150-Kd Subunit Is Not an Abscisic Acid Receptor. Plant Physiol. 2009, 150, 157–166. [Google Scholar] [CrossRef]
- Chan, K.X.; Phua, S.Y.; Crisp, P.; McQuinn, R.; Pogson, B.J. Learning the Languages of the Chloroplast: Retrograde Signaling and Beyond. Annu. Rev. Plant Biol. 2016, 67, 25–53. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2005; p. xxiii. 922p. [Google Scholar]
- Li, Y.; Han, Y.; Qu, M.; Chen, J.; Chen, X.; Geng, X.; Wang, Z.; Chen, S. Apoplastic Cell Death-Inducing Proteins of Filamentous Plant Pathogens: Roles in Plant-Pathogen Interactions. Front. Genet. 2020, 11, 661. [Google Scholar] [CrossRef]
- Caten, C.; Jinks, J. Spontaneous Variability of Single Isolates of Phytophthora Infestans. I. Cultural Variation. Can. J. Bot. 1968, 46, 329–348. [Google Scholar] [CrossRef]
- Wilson, U.; Coffey, M. Cytological Evaluation of General Resistance to Phytophthora Infestans in Potato Foliage. Ann. Bot. 1980, 45, 81–90. [Google Scholar]
- Yin, J.; Gu, B.; Huang, G.; Tian, Y.; Quan, J.; Lindqvist-Kreuze, H.; Shan, W. Conserved Rxlr Effector Genes of Phytophthora Infestans Expressed at the Early Stage of Potato Infection Are Suppressive to Host Defense. Front. Plant Sci. 2017, 8, 2155. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih Image to Imagej: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Faurobert, M.; Pelpoir, E.; Chaib, J. Phenol Extraction of Proteins for Proteomic Studies of Recalcitrant Plant Tissues. Methods Mol. Biol. 2007, 355, 9–14. [Google Scholar]
- Wu, X.; Xiong, E.; Wang, W.; Scali, M.; Cresti, M. Universal Sample Preparation Method Integrating Trichloroacetic Acid/Acetone Precipitation with Phenol Extraction for Crop Proteomic Analysis. Nat. Protoc. 2014, 9, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, R.S.; Rose, J.K. A Critical Evaluation of Sample Extraction Techniques for Enhanced Proteomic Analysis of Recalcitrant Plant Tissues. Proteomics 2004, 4, 2522–2532. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.T.; Wang, K.H.; Chang, W.H.; Yang, J.C.; Yeh, C.F.; Cheng, K.T.; Hung, S.C.; Chen, Y.R. Application of Data-Independent Acquisition Approach to Study the Proteome Change from Early to Later Phases of Tomato Pathogenesis Responses. Int. J. Mol. Sci. 2019, 20, 863. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, W.Y.; Tseng, M.C.; Chen, Y.R. Tunnel Frit: A Nonmetallic in-Capillary Frit for Nanoflow Ultra High-Performance Liquid Chromatography-Mass Spectrometryapplications. Anal. Chem. 2012, 84, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Scheltema, R.A.; Hauschild, J.P.; Lange, O.; Hornburg, D.; Denisov, E.; Damoc, E.; Kuehn, A.; Makarov, A.; Mann, M. The Q Exactive HF, a Benchtop Mass Spectrometer with a Pre-Filter, High-Performance Quadrupole and an Ultra-High-Field Orbitrap Analyzer. Mol. Cell. Proteom. MCP 2014, 13, 3698–3708. [Google Scholar] [CrossRef]
- Bruderer, R.; Bernhardt, O.M.; Gandhi, T.; Xuan, Y.; Sondermann, J.; Schmidt, M.; Gomez-Varela, D.; Reiter, L. Optimization of Experimental Parameters in Data-Independent Mass Spectrometry Significantly Increases Depth and Reproducibility of Results. Mol. Cell. Proteom. MCP 2017, 16, 2296–2309. [Google Scholar] [CrossRef]
- Craig, R.; Beavis, R.C. Tandem: Matching Proteins with Tandem Mass Spectra. Bioinformatics 2004, 20, 1466–1467. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical Statistical Model to Estimate the Accuracy of Peptide Identifications Made by MS/MS and Database Search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef] [PubMed]
- Shteynberg, D.; Deutsch, E.W.; Lam, H.; Eng, J.K.; Sun, Z.; Tasman, N.; Mendoza, L.; Moritz, R.L.; Aebersold, R.; Nesvizhskii, A.I. Iprophet: Multi-Level Integrative Analysis of Shotgun Proteomic Data Improves Peptide and Protein Identification Rates and Error Estimates. Mol. Cell. Proteom. MCP 2011, 10, M111-007690. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, E.W.; Mendoza, L.; Shteynberg, D.; Slagel, J.; Sun, Z.; Moritz, R.L. Trans-Proteomic Pipeline, a Standardized Data Processing Pipeline for Large-Scale Reproducible Proteomics Informatics. Proteom. Clin. Appl. 2015, 9, 745–754. [Google Scholar] [CrossRef]
- Reiter, L.; Claassen, M.; Schrimpf, S.P.; Jovanovic, M.; Schmidt, A.; Buhmann, J.M.; Hengartner, M.O.; Aebersold, R. Protein Identification False Discovery Rates for Very Large Proteomics Data Sets Generated by Tandem Mass Spectrometry. Mol. Cell. Proteom. MCP 2009, 8, 2405–2417. [Google Scholar] [CrossRef]
- Lam, H.; Deutsch, E.W.; Eddes, J.S.; Eng, J.K.; Stein, S.E.; Aebersold, R. Building Consensus Spectral Libraries for Peptide Identification in Proteomics. Nat. Methods 2008, 5, 873–875. [Google Scholar] [CrossRef]
- Röst, H.L.; Sachsenberg, T.; Aiche, S.; Bielow, C.; Weisser, H.; Aicheler, F.; Andreotti, S.; Ehrlich, H.C.; Gutenbrunner, P.; Kenar, E.; et al. OpenMS: A Flexible Open-Source Software Platform for Mass Spectrometry Data Analysis. Nat. Methods 2016, 13, 741–748. [Google Scholar] [CrossRef]
- Schubert, O.T.; Gillet, L.C.; Collins, B.C.; Navarro, P.; Rosenberger, G.; Wolski, W.E.; Lam, H.; Amodei, D.; Mallick, P.; MacLean, B.; et al. Building High-Quality Assay Libraries for Targeted Analysis of Swath MS Data. Nat. Protoc. 2015, 10, 426–441. [Google Scholar] [CrossRef]
- Röst, H.L.; Rosenberger, G.; Navarro, P.; Gillet, L.; Miladinović, S.M.; Schubert, O.T.; Wolski, W.; Collins, B.C.; Malmström, J.; Malmström, L.; et al. OpenSWATH Enables Automated, Targeted Analysis of Data- Independent Acquisition MS Data. Nat. Biotechnol. 2014, 32, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.P.; Ivosev, G.; Couzens, A.L.; Larsen, B.; Taipale, M.; Lin, Z.Y.; Zhong, Q.; Lindquist, S.; Vidal, M.; Aebersold, R.; et al. Mapping Differential Interactomes by Affinity Purification Coupled with Data-Independent Mass Spectrometry Acquisition. Nat. Methods 2013, 10, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE Database and Related Tools and Resources in 2019: Improving Support for Quantification Data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 24 h | 48 h | 96 h | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Accession | Protein Description | Arabidopsis Homolog | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc |
| Solyc10g055800 | chitinase | AT3G12500 | 3 | −0.121 | 0.804 | 3 | 0.454 | 0.347 | 3 | 1.816 | 0.023 |
| Solyc02g082920 | class II chitinase | AT3G12500 | 3 | 0.140 | 0.326 | 3 | 0.556 | 0.009 | 3 | 2.982 | 0.019 |
| Solyc01g008620 | glucan endo-1,3-beta-glucosidase | AT4G16260 | 3 | 0.072 | 0.133 | 3 | −0.022 | 0.933 | 3 | 2.563 | 0.001 |
| Solyc10g079860 | LEQB L.esculentum TomQ’b beta (1,3) glucanase | AT3G57270 | 3 | 0.089 | 0.496 | 3 | 0.612 | 0.344 | 3 | 4.055 | 0.007 |
| Solyc09g090990 | major allergen Pru ar.1 (PR-10 family) | AT5G45860 | 3 | 0.755 | 0.162 | 3 | 2.446 | 0.002 | 3 | 4.479 | 0.001 |
| Solyc07g005370 | norcoclaurine synthase (NCS; PR-10/Bet v 1 family) | AT2G26040 | 3 | 0.135 | 0.642 | 3 | 0.375 | 0.153 | 3 | −0.756 | 0.033 |
| Solyc08g080670 | pathogenesis-related 5-like protein (PR-5) | AT4G11650 | 3 | −1.633 | 0.005 | 3 | −0.298 | 0.623 | 3 | 0.394 | 0.436 |
| Solyc01g097240 | pathogenesis-related protein 4 (PR-4) | AT3G04720 | 3 | 0.199 | 0.027 | 3 | 0.522 | 0.058 | 3 | 3.745 | 0.007 |
| Solyc09g090980 | pathogenesis-related protein STH-2-like (PR-10 family) | AT1G24020 | 3 | 0.185 | 0.009 | 3 | 1.206 | 0.010 | 3 | 3.456 | 0.004 |
| 24 h | 48 h | 96 h | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Accession | Protein Description | Arabidopsis Homolog | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc |
| Solyc02g077880 | auxin-repressed protein (ARP) | AT2G33830 | 3 | −0.404 | 0.202 | 3 | −1.360 | 0.072 | 3 | −2.441 | 0.007 |
| Solyc03g033790 | P-loop containing nucleoside triphosphate hydrolases superfamily protein | AT3G50930 | 1 | 0.083 | NQ | 2 | 0.304 | NQ | 3 | 1.340 | 0.021 |
| Solyc06g068840 | calcium-dependent phospholipid-binding copine family protein | AT5G61900 | 2 | 0.262 | NQ | 3 | 0.534 | 0.181 | 3 | 1.015 | 0.030 |
| Solyc02g067750 | carbonic anhydrase (CA) | AT3G01500 | 3 | 1.884 | 0.719 | 3 | −1.904 | 0.374 | 3 | 8.330 | 0.012 |
| Solyc04g078540 | cysteine proteinase RD21a | AT1G47128 | 3 | 0.331 | 0.102 | 3 | −0.029 | 0.888 | 3 | 0.716 | 0.036 |
| Solyc06g071280 | enhanced disease susceptibility 1 (EDS1) | AT3G48090 | 3 | −0.015 | 0.100 | 3 | 0.092 | 0.594 | 3 | 0.930 | 0.037 |
| Solyc06g071050 | hypersensitive-induced response protein (HIR) | AT5G62740 | 2 | −0.334 | NQ | 3 | −0.430 | 0.525 | 3 | 0.686 | 0.016 |
| Solyc03g098730 | Kunitz-type protease inhibitor (KTI) | AT1G17860 | 3 | −0.607 | 0.220 | 3 | 0.076 | 0.744 | 3 | 2.740 | 0.008 |
| Solyc07g043320 | myosin heavy chain, embryonic smooth protein | AT2G32240 | 3 | 0.129 | 0.276 | 3 | 0.578 | 0.188 | 3 | 1.050 | 0.030 |
| Solyc00g174340 | pathogenesis-related protein 1b (PR-1b) | AT4G33720 | 3 | 0.058 | 0.484 | 3 | 0.591 | 0.025 | 3 | 2.902 | 0.008 |
| Solyc09g007010 | pathogenesis-related protein 1 (PR-1) | AT4G33720 | 3 | 0.097 | 0.354 | 3 | 0.411 | 0.302 | 3 | 2.296 | 0.043 |
| Solyc02g077040 | phytophthora-inhibited protease 1 (PIP1) | AT3G49340 | 3 | 0.183 | 0.052 | 3 | 0.291 | 0.316 | 3 | 1.585 | 0.024 |
| 24 h | 48 h | 96 h | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Accession | Protein Description | Arabidopsis Homolog | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc |
| Solyc07g049530 | 1-aminocyclopropane-1-carboxylate oxidase 1 (ACO1) | AT1G05010 | 3 | 0.109 | 0.521 | 3 | 0.877 | 0.007 | 3 | 2.288 | 0.012 |
| Solyc04g015750 | cobN/magnesium chelatase (CHLH/ABAR) | AT5G13630 | 3 | 0.629 | 0.042 | 3 | 0.155 | 0.625 | 3 | −1.239 | 0.031 |
| Solyc01g099160 | lipoxygenase (Lox) | AT1G55020 | 0 | ND | ND | 3 | 2.067 | 0.003 | 3 | 3.063 | 0.055 |
| Solyc08g029000 | lipoxygenase (Lox) | AT1G55020 | 3 | −0.291 | 0.527 | 3 | 1.889 | 0.003 | 3 | 3.804 | 0.008 |
| Solyc03g122340 | lipoxygenase D (LoxD) | AT1G17420 | 3 | −0.611 | 0.046 | 3 | −0.453 | 0.394 | 1 | −1.177 | NQ |
| Solyc12g062290 | protease do-like 9 (DEGP9) | AT5G40200 | 3 | 0.133 | 0.268 | 3 | −0.588 | 0.011 | 2 | 0.000 | NQ |
| Solyc01g100050 | topless 3 (TPL3) | AT5G27030 | 3 | 0.205 | 0.067 | 3 | 0.307 | 0.188 | 3 | 0.817 | 0.004 |
| 24 h | 48 h | 96 h | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Accession | Protein Description | Arabidopsis Homolog | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc |
| Solyc04g073990 | annexin p34 (ANXP34) | AT1G35720 | 3 | −0.122 | 0.603 | 3 | 0.107 | 0.883 | 3 | 0.690 | 0.002 |
| Solyc04g082460 | catalase (CAT) | AT4G35090 | 3 | 0.223 | 0.122 | 3 | −0.182 | 0.724 | 3 | −0.845 | 0.046 |
| Solyc01g107860 | CBS domain-containing protein-like (CDCP-like) | AT4G36910 | 3 | −0.026 | 0.345 | 3 | −0.046 | 0.777 | 3 | 1.068 | 0.010 |
| Solyc11g012910 | gamma-glutamylcyclotransferase (GGCG) | AT1G44790 | 3 | −0.564 | 0.046 | 3 | 0.369 | 0.418 | 3 | −1.588 | 0.003 |
| Solyc08g080940 | glutathione peroxidase -like encoding 1 (GPX-1) | AT4G11600 | 3 | −0.037 | 0.498 | 3 | 0.241 | 0.319 | 3 | 0.957 | 0.019 |
| Solyc06g083770 | glutathione S-transferase (GST) | AT5G44000 | 3 | −0.008 | 0.967 | 3 | −0.260 | 0.366 | 3 | −1.300 | 0.016 |
| Solyc10g084400 | glutathione S-transferase (GST) | AT5G02790 | 3 | −0.099 | 0.596 | 3 | 0.221 | 0.100 | 3 | 1.572 | 0.000 |
| Solyc09g011590 | glutathione S-transferase-like protein (GST-like) | AT3G09270 | 3 | −1.121 | 0.316 | 3 | 0.987 | 0.272 | 3 | 2.891 | 0.007 |
| Solyc02g081430 | microsomal Glutathione s-Transferase (MGST) | AT1G65820 | 3 | 0.082 | 0.263 | 3 | 0.201 | 0.644 | 3 | −0.790 | 0.006 |
| Solyc01g096430 | NADPH:quinone oxidoreductase-like (NQR-like) | AT3G27890 | 2 | −0.040 | NQ | 3 | 0.140 | 0.814 | 3 | 1.794 | 0.034 |
| Solyc03g111720 | peptide methionine sulfoxide reductase (MSR) | AT5G61640 | 3 | −0.440 | 0.079 | 3 | 0.135 | 0.396 | 3 | 0.829 | 0.010 |
| Solyc10g076240 | peroxidase (POX) | AT5G05340 | 0 | NQ | NQ | 0 | NQ | NQ | 3 | 3.731 | 0.037 |
| Solyc08g069040 | peroxidase (POX) | AT4G37530 | 2 | −0.165 | NQ | 3 | −0.831 | 0.041 | 2 | −0.428 | NQ |
| Solyc01g067740 | superoxide dismutase 1 (CSD1) | AT1G08830 | 3 | 0.320 | 0.060 | 3 | 0.074 | 0.820 | 3 | 1.113 | 0.038 |
| Solyc04g081970 | thioredoxin (TRX) | AT1G76080 | 3 | 0.021 | 0.805 | 3 | 0.000 | 1.000 | 3 | −1.087 | 0.011 |
| Solyc02g082250 | thioredoxin reductase (TRXR) | AT2G17420 | 3 | 0.015 | 0.883 | 3 | 0.217 | 0.262 | 3 | 0.607 | 0.013 |
| Solyc03g117980 | whitefly-induced gp91-phox (GP91phox) | AT5G47910 | 3 | 0.039 | 0.777 | 3 | 0.765 | 0.038 | 3 | 2.345 | 0.016 |
| 24 h | 48 h | 96 h | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Accession | Protein Description | Arabidopsis Homolog | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc |
| Metabolism-Primary-Energy Metabolisms | |||||||||||
| Solyc11g007720 | pyruvate dehydrogenase complex (PDC) | AT3G52200 | 3 | −0.121 | 0.358 | 3 | 0.082 | 0.842 | 3 | 0.645 | 0.017 |
| Solyc01g073740 | citrate synthase (CSY) | AT2G44350 | 3 | −0.112 | 0.410 | 3 | −0.198 | 0.283 | 3 | 0.639 | 0.017 |
| Solyc07g052350 | aconitate hydratase (ACO) | AT2G05710 | 3 | 0.019 | 0.872 | 3 | 0.110 | 0.544 | 3 | 0.708 | 0.047 |
| Solyc12g005860 | aconitate hydratase (ACO) | AT2G05710 | 3 | −0.084 | 0.054 | 3 | −0.100 | 0.605 | 3 | 0.968 | 0.023 |
| Solyc01g005560 | isocitrate dehydrogenase (IDH) | AT1G65930 | 3 | −0.125 | 0.650 | 3 | 0.311 | 0.369 | 3 | 0.753 | 0.049 |
| Solyc07g064800 | oxoglutarate dehydrogenase complex (ODC) | AT4G26910 | 3 | 0.049 | 0.497 | 3 | 0.150 | 0.700 | 3 | 0.607 | 0.046 |
| Solyc12g005080 | oxoglutarate dehydrogenase complex (ODC) | AT5G55070 | 3 | −0.073 | 0.661 | 3 | 0.107 | 0.043 | 3 | 1.003 | 0.023 |
| Solyc06g083790 | succinyl-CoA ligase (SCoAL) | AT2G20420 | 3 | 0.058 | 0.107 | 3 | 0.039 | 0.888 | 3 | 0.602 | 0.016 |
| Solyc01g007910 | succinyl-CoA ligase (SCoAL) | AT5G23250 | 3 | −0.131 | 0.132 | 3 | 0.295 | 0.529 | 3 | 0.630 | 0.035 |
| Solyc09g075450 | fumarase (FUM) | AT2G47510 | 3 | 0.088 | 0.362 | 3 | 0.091 | 0.691 | 3 | 0.953 | 0.004 |
| Solyc08g066360 | malic enzyme (ME) | AT1G79750 | 3 | −0.007 | 0.978 | 3 | −0.016 | 0.957 | 3 | 0.903 | 0.045 |
| Metabolism-Primary-Glycolysis | |||||||||||
| Solyc07g045160 | ATP-dependent 6-phosphofructokinase (PFK) | AT4G26270 | 3 | 0.217 | 0.106 | 3 | 0.107 | 0.497 | 3 | 1.052 | 0.037 |
| Solyc10g005510 | glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | AT1G16300 | 3 | 0.097 | 0.413 | 3 | 0.220 | 0.713 | 3 | 1.096 | 0.045 |
| Solyc03g114500 | enolase (ENO1) | AT1G74030 | 3 | 0.599 | 0.049 | 3 | −1.053 | 0.430 | 2 | 1.073 | NQ |
| Solyc03g097680 | pyruvate dehydrogenase 1 (PDH) | AT5G50850 | 3 | −0.048 | 0.857 | 3 | −0.270 | 0.724 | 3 | 1.251 | 0.038 |
| Metabolism-Primary-Carbohydrate Metabolisms-PPP | |||||||||||
| Solyc04g005160 | 6-phosphogluconate dehydrogenase (6PGD) | AT3G02360 | 3 | −0.046 | 0.033 | 3 | 0.524 | 0.031 | 3 | 1.886 | 0.016 |
| Solyc05g008370 | ribose 5-phosphate isomerase A (Rpi) | AT1G71100 | 3 | 0.068 | 0.366 | 3 | 0.211 | 0.715 | 3 | 0.790 | 0.036 |
| Solyc11g033288 | transaldolase (TAL) | AT5G13420 | 3 | 0.029 | 0.751 | 3 | −0.206 | 0.729 | 3 | 0.667 | 0.032 |
| Metabolism-Primary-Fatty Acid/Lipids | |||||||||||
| Solyc10g076600 | acyl-CoA oxidase/dehydrogenase (ACAD) | AT3G51840 | 3 | −0.057 | 0.872 | 3 | 0.050 | 0.928 | 3 | 2.085 | 0.014 |
| Solyc01g059830 | enoyl-CoA hydratase (ECH) | AT4G16210 | 3 | 0.469 | 0.386 | 2 | −0.156 | NQ | 3 | 0.750 | 0.041 |
| Solyc12g094450 | enoyl-CoA hydratase (ECH) | AT1G76150 | 2 | −0.278 | NQ | 3 | 0.202 | 0.216 | 3 | 1.663 | 0.002 |
| 24 h | 48 h | 96 h | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Accession | Protein Description | Arabidopsis Homolog | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc |
| Solyc06g063370 | chlorophyll a-b binding protein (LHCB) | AT4G10340 | 3 | −0.061 | 0.659 | 3 | 0.391 | 0.212 | 3 | −0.662 | 0.014 |
| Solyc12g005630 | cytochrome b6-f complex iron-sulfur subunit | AT4G03280 | 3 | ‒0.368 | 0.077 | 3 | ‒0.001 | 0.998 | 3 | ‒0.739 | 0.001 |
| Solyc06g009940 | photosystem I (PSI) P700 chlorophyll a apoprotein | ATCG00350 | 3 | 0.029 | 0.810 | 3 | ‒0.121 | 0.869 | 3 | ‒0.836 | 0.015 |
| Solyc02g069450 | photosystem I (PSI) reaction center subunit III | AT1G31330 | 3 | 0.107 | 0.605 | 3 | 0.048 | 0.930 | 3 | ‒0.661 | 0.038 |
| Solyc08g013670 | photosystem I (PSI) reaction center subunit N | AT5G64040 | 3 | 0.416 | 0.244 | 3 | 0.412 | 0.498 | 3 | ‒0.854 | 0.033 |
| Solyc05g150152 | photosystem I (PSI) reaction centre subunit N protein | AT1G49975 | 2 | 0.073 | NQ | 3 | ‒0.279 | 0.587 | 3 | ‒0.599 | 0.048 |
| Solyc06g060340 | photosystem II (PSII) 22 kDa protein | AT1G44575 | 3 | ‒0.091 | 0.339 | 3 | 0.234 | 0.063 | 3 | ‒0.910 | 0.008 |
| Solyc05g052600 | sedoheptulose-1,7-bisphosphatase (SBPase) | AT3G55800 | 3 | 0.077 | 0.001 | 3 | ‒0.292 | 0.626 | 3 | ‒0.758 | 0.039 |
| 24 h | 48 h | 96 h | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Accession | Protein Description | Arabidopsis Homolog | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc | # rep a | log2 ratio b | pc |
| Solyc06g150137 | cinnamate 4-hydroxylase (C4H) | AT2G30490 | 3 | −0.075 | 0.505 | 3 | 0.164 | 0.736 | 3 | 1.390 | 0.004 |
| Solyc03g117870 | 4-coumarate:CoA ligase (4CL) | AT3G21240 | 3 | 0.185 | 0.366 | 3 | 0.622 | 0.178 | 3 | 2.293 | 0.004 |
| Solyc01g091190 | 5-enolpyruvylshikimate-3-phosphate synthase (ESPS) | AT2G45300 | 3 | −0.124 | 0.369 | 3 | 0.208 | 0.229 | 3 | 1.165 | 0.022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, K.-T.; Hsu, Y.; Yeh, C.-F.; Chang, C.-H.; Chang, W.-H.; Chen, Y.-R. Quantitative Proteomics Reveals the Dynamic Regulation of the Tomato Proteome in Response to Phytophthora infestans. Int. J. Mol. Sci. 2021, 22, 4174. https://doi.org/10.3390/ijms22084174
Fan K-T, Hsu Y, Yeh C-F, Chang C-H, Chang W-H, Chen Y-R. Quantitative Proteomics Reveals the Dynamic Regulation of the Tomato Proteome in Response to Phytophthora infestans. International Journal of Molecular Sciences. 2021; 22(8):4174. https://doi.org/10.3390/ijms22084174
Chicago/Turabian StyleFan, Kai-Ting, Yang Hsu, Ching-Fang Yeh, Chi-Hsin Chang, Wei-Hung Chang, and Yet-Ran Chen. 2021. "Quantitative Proteomics Reveals the Dynamic Regulation of the Tomato Proteome in Response to Phytophthora infestans" International Journal of Molecular Sciences 22, no. 8: 4174. https://doi.org/10.3390/ijms22084174
APA StyleFan, K.-T., Hsu, Y., Yeh, C.-F., Chang, C.-H., Chang, W.-H., & Chen, Y.-R. (2021). Quantitative Proteomics Reveals the Dynamic Regulation of the Tomato Proteome in Response to Phytophthora infestans. International Journal of Molecular Sciences, 22(8), 4174. https://doi.org/10.3390/ijms22084174