
Targeting the Reconsolidation of Licit Drug Memories to Prevent Relapse: Focus on Alcohol and Nicotine


Abstract
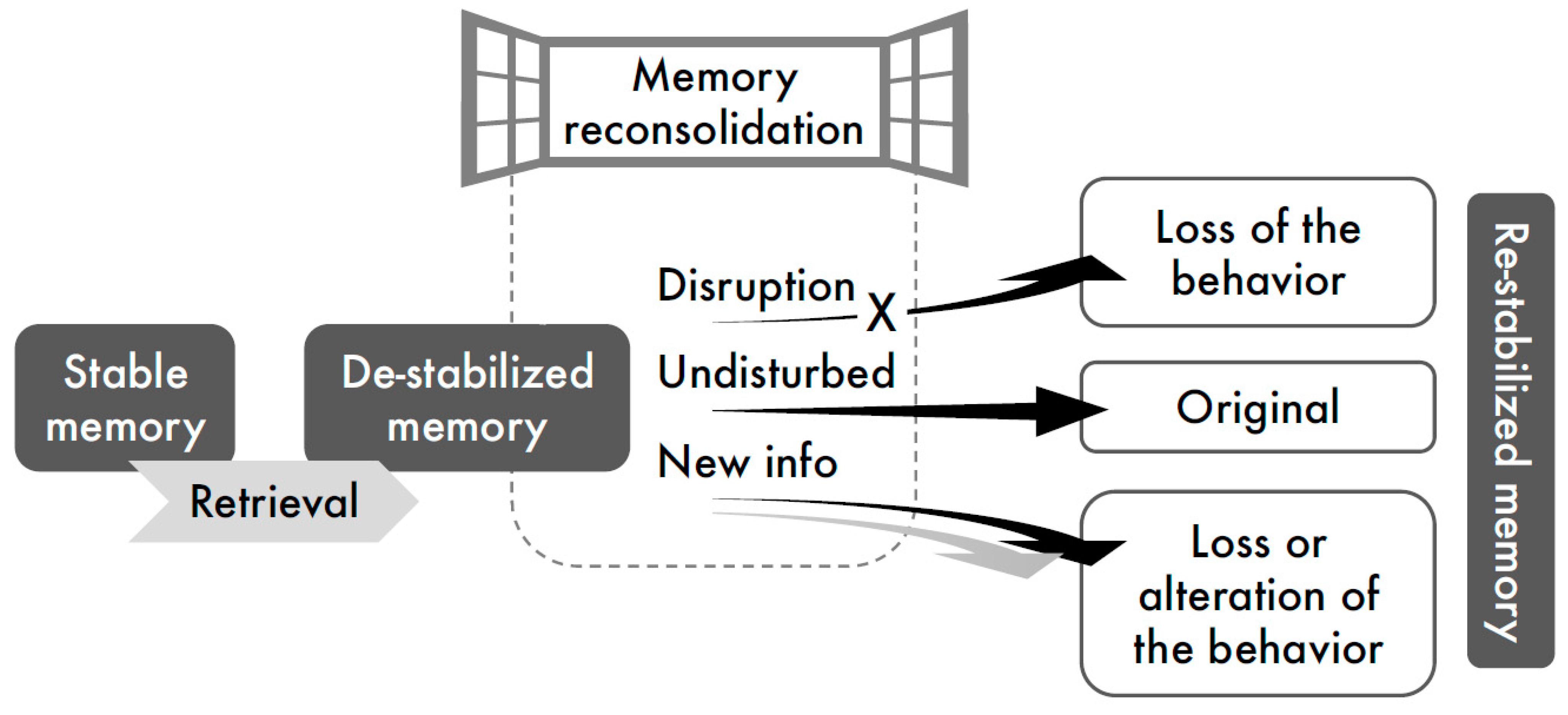
1. Introduction
2. Alcohol
2.1. Pharmacological Interference with Alcohol-Memory Reconsolidation
2.1.1. NMDA Receptor Blockade
2.1.2. β-Adrenergic Receptor Blockade
2.1.3. Protein Synthesis Inhibition
2.2. Behavioral Interference with Alcohol-Memory Reconsolidation
3. Nicotine and Tobacco
3.1. Pharmacological Interference with Nicotine-Memory Reconsolidation
3.1.1. NMDA Receptor Blockade
3.1.2. Adrenergic-Receptors Blockade
3.1.3. Additional Pharmacological Manipulations
3.2. Behavioral Manipulations
4. Conclusive and Critical Remarks
4.1. Methodological Standardization and Replicability of Effects
4.1.1. Classical vs. Operant Conditioning Paradigms, and Limited Effectivity
4.1.2. Standardization of Protocols and Parameters
4.1.3. Retrieval Procedures
4.1.4. Timing of the Manipulation
4.2. Translational/Clinical Limitations and Possible Solutions
4.2.1. Strong Memories in Self-Administration Procedures and Implications for Addiction
4.2.2. Which Memory Is Targeted?
4.2.3. Context- and State-Dependency of the Treatment
4.2.4. Generalization of Results
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peacock, A.; Leung, J.; Larney, S.; Colledge, S.; Hickman, M.; Rehm, J.; Giovino, G.A.; West, R.; Hall, W.; Griffiths, P. Global statistics on alcohol, tobacco and illicit drug use: 2017 status report. Addiction 2018, 113, 1905–1926. [Google Scholar] [CrossRef]
- World Health Organization. Global Status Report on Alcohol and Health 2018; WHO: Geneva, Switzerland, 2018; Available online: https://www.who.int/substance_abuse/publications/global_alcohol_report/en/ (accessed on 16 February 2021).
- García-Rodríguez, O.; Secades-Villa, R.; Flórez-Salamanca, L.; Okuda, M.; Liu, S.-M.; Blanco, C. Probability and predictors of relapse to smoking: Results of the National Epidemiologic Survey on Alcohol and Related Conditions (NESARC). Drug Alcohol Depend. 2013, 132, 479–485. [Google Scholar] [CrossRef]
- Cooney, N.L.; Litt, M.D.; Sevarino, K.A.; Levy, L.; Kranitz, L.S.; Sackler, H.; Cooney, J.L. Concurrent alcohol and tobacco treatment: Effect on daily process measures of alcohol relapse risk. J. Consult. Clin. Psychol. 2015, 83, 346. [Google Scholar] [CrossRef] [PubMed]
- Venniro, M.; Caprioli, D.; Shaham, Y. Animal models of drug relapse and craving. In Neuroscience for Addiction Medicine: From Prevention to Rehabilitation—Methods and Interventions; Ekhtiari, H., Paulus, M.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 224, pp. 25–52. [Google Scholar]
- Witteman, J.; Post, H.; Tarvainen, M.; de Bruijn, A.; Perna Ede, S.; Ramaekers, J.G.; Wiers, R.W. Cue reactivity and its relation to craving and relapse in alcohol dependence: A combined laboratory and field study. Psychopharmacology 2015, 232, 3685–3696. [Google Scholar] [CrossRef] [PubMed]
- Niaura, R.S.; Rohsenow, D.J.; Binkoff, J.A.; Monti, P.M.; Pedraza, M.; Abrams, D.B. Relevance of cue reactivity to understanding alcohol and smoking relapse. J. Abnorm. Psychol. 1988, 97, 133. [Google Scholar] [CrossRef] [PubMed]
- Bouton, M.E. Context, ambiguity, and unlearning: Sources of relapse after behavioral extinction. Biol. Psychiatry 2002, 52, 976–986. [Google Scholar] [CrossRef]
- Lee, J.L.C.; Nader, K.; Schiller, D. An update on memory reconsolidation updating. Trends Cogn. Sci. 2017, 21, 531–545. [Google Scholar] [CrossRef]
- Nader, K.; Hardt, O. A single standard for memory: The case for reconsolidation. Nat. Rev. Neurosci. 2009, 10, 224–234. [Google Scholar] [CrossRef]
- Nader, K.; Schafe, G.E.; Le Doux, J.E. Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval. Nature 2000, 406, 722–726. [Google Scholar] [CrossRef]
- Przybyslawski, J.; Roullet, P.; Sara, S.J. Attenuation of emotional and nonemotional memories after their reactivation: Role of beta adrenergic receptors. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 6623–6628. [Google Scholar] [CrossRef]
- Valjent, E.; Corbille, A.G.; Bertran-Gonzalez, J.; Herve, D.; Girault, J.A. Inhibition of ERK pathway or protein synthesis during reexposure to drugs of abuse erases previously learned place preference. Proc. Natl. Acad. Sci. USA 2006, 103, 2932–2937. [Google Scholar] [CrossRef] [PubMed]
- Barak, S.; Liu, F.; Ben Hamida, S.; Yowell, Q.V.; Neasta, J.; Kharazia, V.; Janak, P.H.; Ron, D. Disruption of alcohol-related memories by mTORC1 inhibition prevents relapse. Nat. Neurosci. 2013, 16, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, A.B.; Taylor, J.R. Reconsolidation and psychopathology: Moving towards reconsolidation-based treatments. Neurobiol. Learn. Mem. 2017, 142, 162–171. [Google Scholar] [CrossRef]
- Taujanskaite, U.; Cahill, E.N.; Milton, A.L. Targeting drug memory reconsolidation: A neural analysis. Curr. Opin. Pharm. 2020, 56, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Exton-McGuinness, M.T.J.; Milton, A.L. Reconsolidation blockade for the treatment of addiction: Challenges, new targets, and opportunities. Learn. Mem. 2018, 25, 492–500. [Google Scholar] [CrossRef]
- Olshavsky, M.E.; Song, B.J.; Powell, D.J.; Jones, C.E.; Monfils, M.H.; Lee, H.J. Updating appetitive memory during reconsolidation window: Critical role of cue-directed behavior and amygdala central nucleus. Front. Behav. Neurosci. 2013, 7, 186. [Google Scholar] [CrossRef][Green Version]
- Auber, A.; Tedesco, V.; Jones, C.E.; Monfils, M.H.; Chiamulera, C. Post-retrieval extinction as reconsolidation interference: Methodological issues or boundary conditions? Psychopharmacology 2013, 226, 631–647. [Google Scholar] [CrossRef]
- Schiller, D.; Monfils, M.H.; Raio, C.M.; Johnson, D.C.; Ledoux, J.E.; Phelps, E.A. Preventing the return of fear in humans using reconsolidation update mechanisms. Nature 2010, 463, 49–53. [Google Scholar] [CrossRef]
- Monfils, M.H.; Cowansage, K.K.; Klann, E.; LeDoux, J.E. Extinction-reconsolidation boundaries: Key to persistent attenuation of fear memories. Science 2009, 324, 951–955. [Google Scholar] [CrossRef]
- Gordon, W.C. Similarities of recently acquired and reactivated memories in interference. Am. J. Psychol. 1977, 90, 231–242. [Google Scholar] [CrossRef]
- Paulus, D.J.; Kamboj, S.K.; Das, R.K.; Saladin, M.E. Prospects for reconsolidation-focused treatments of substance use and anxiety-related disorders. Curr. Opin. Psychol. 2019, 30, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Goltseker, K.; Bolotin, L.; Barak, S. Counterconditioning during reconsolidation prevents relapse of cocaine memories. Neuropsychopharmacology 2017, 42, 716–726. [Google Scholar] [CrossRef]
- Goltseker, K.; Handrus, H.; Barak, S. Disruption of relapse to alcohol seeking by aversive counterconditioning following memory retrieval. Addict. Biol. 2020, e12935. [Google Scholar] [CrossRef] [PubMed]
- Von der Goltz, C.; Vengeliene, V.; Bilbao, A.; Perreau-Lenz, S.; Pawlak, C.R.; Kiefer, F.; Spanagel, R. Cue-induced alcohol-seeking behaviour is reduced by disrupting the reconsolidation of alcohol-related memories. Psychopharmacology 2009, 205, 389–397. [Google Scholar] [CrossRef]
- Lin, J.; Liu, L.; Wen, Q.; Zheng, C.; Gao, Y.; Peng, S.; Tan, Y.; Li, Y. Rapamycin prevents drug seeking via disrupting reconsolidation of reward memory in rats. Int. J. Neuropsychopharmacol. 2014, 17, 127–136. [Google Scholar] [CrossRef]
- Milton, A.L.; Schramm, M.J.; Wawrzynski, J.R.; Gore, F.; Oikonomou-Mpegeti, F.; Wang, N.Q.; Samuel, D.; Economidou, D.; Everitt, B.J. Antagonism at NMDA receptors, but not beta-adrenergic receptors, disrupts the reconsolidation of pavlovian conditioned approach and instrumental transfer for ethanol-associated conditioned stimuli. Psychopharmacology 2012, 219, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Das, R.K.; Gale, G.; Walsh, K.; Hennessy, V.E.; Iskandar, G.; Mordecai, L.A.; Brandner, B.; Kindt, M.; Curran, H.V.; Kamboj, S.K. Ketamine can reduce harmful drinking by pharmacologically rewriting drinking memories. Nat. Commun. 2019, 10, 5187. [Google Scholar] [CrossRef]
- Puaud, M.; Ossowska, Z.; Barnard, J.; Milton, A.L. Saccharin fading is not required for the acquisition of alcohol self-administration, and can alter the dynamics of cue-alcohol memory reconsolidation. Psychopharmacology 2018, 235, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Vengeliene, V.; Olevska, A.; Spanagel, R. Long-lasting effect of NMDA receptor antagonist memantine on ethanol-cue association and relapse. J. Neurochem. 2015, 135, 1080–1085. [Google Scholar] [CrossRef]
- Wouda, J.A.; Diergaarde, L.; Riga, D.; van Mourik, Y.; Schoffelmeer, A.N.; De Vries, T.J. Disruption of Long-Term Alcohol-Related Memory Reconsolidation: Role of beta-Adrenoceptors and NMDA Receptors. Front. Behav. Neurosci. 2010, 4, 179. [Google Scholar] [CrossRef]
- Schramm, M.J.; Everitt, B.J.; Milton, A.L. Bidirectional modulation of alcohol-associated memory reconsolidation through manipulation of adrenergic signaling. Neuropsychopharmacology 2016, 41, 1103–1111. [Google Scholar] [CrossRef]
- Chesworth, R.; Corbit, L.H. Noradrenergic beta-receptor antagonism in the basolateral amygdala impairs reconsolidation, but not extinction, of alcohol self-administration: Intra-BLA propranolol impairs reconsolidation of alcohol self-administration. Neurobiol. Learn. Mem. 2018, 151, 59–70. [Google Scholar] [CrossRef]
- Font, L.; Cunningham, C.L. Post-retrieval propranolol treatment does not modulate reconsolidation or extinction of ethanol-induced conditioned place preference. Pharmacol. Biochem. Behav. 2012, 101, 222–230. [Google Scholar] [CrossRef]
- Lonergan, M.; Saumier, D.; Tremblay, J.; Kieffer, B.; Brown, T.G.; Brunet, A. Reactivating addiction-related memories under propranolol to reduce craving: A pilot randomized controlled trial. J. Behav. Ther. Exp. Psychiatry 2016, 50, 245–249. [Google Scholar] [CrossRef]
- Exton-McGuinness, M.T.J.; Drame, M.L.; Flavell, C.R.; Lee, J.L.C. On the resistance to relapse to cocaine-seeking following impairment of instrumental cocaine memory reconsolidation. Front. Behav. Neurosci. 2019, 13, 242. [Google Scholar] [CrossRef]
- Exton-McGuinness, M.T.; Patton, R.C.; Sacco, L.B.; Lee, J.L. Reconsolidation of a well-learned instrumental memory. Learn. Mem. 2014, 21, 468–477. [Google Scholar] [CrossRef]
- Exton-McGuinness, M.T.; Lee, J.L. Reduction in responding for sucrose and cocaine reinforcement by disruption of memory reconsolidation. eNeuro 2015, 2. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.X.; Xue, Y.X.; Liu, J.F.; Shi, H.S.; Jian, M.; Han, Y.; Zhu, W.L.; Bao, Y.P.; Wu, P.; Ding, Z.B.; et al. A novel UCS memory retrieval-extinction procedure to inhibit relapse to drug seeking. Nat. Commun. 2015, 6, 7675. [Google Scholar] [CrossRef]
- Xue, Y.X.; Luo, Y.X.; Wu, P.; Shi, H.S.; Xue, L.F.; Chen, C.; Zhu, W.L.; Ding, Z.B.; Bao, Y.P.; Shi, J.; et al. A memory retrieval-extinction procedure to prevent drug craving and relapse. Science 2012, 336, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Forcato, C.; Fernandez, R.S.; Pedreira, M.E. Strengthening a consolidated memory: The key role of the reconsolidation process. J. Physiol. Paris 2014, 108, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Milekic, M.H.; Alberini, C.M. Temporally graded requirement for protein synthesis following memory reactivation. Neuron 2002, 36, 521–525. [Google Scholar] [CrossRef]
- Suzuki, A.; Josselyn, S.A.; Frankland, P.W.; Masushige, S.; Silva, A.J.; Kida, S. Memory reconsolidation and extinction have distinct temporal and biochemical signatures. J. Neurosci. 2004, 24, 4787–4795. [Google Scholar] [CrossRef] [PubMed]
- Frankland, P.W.; Ding, H.K.; Takahashi, E.; Suzuki, A.; Kida, S.; Silva, A.J. Stability of recent and remote contextual fear memory. Learn. Mem. 2006, 13, 451–457. [Google Scholar] [CrossRef]
- Bustos, S.G.; Maldonado, H.; Molina, V.A. Disruptive effect of midazolam on fear memory reconsolidation: Decisive influence of reactivation time span and memory age. Neuropsychopharmacology 2009, 34, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Roullet, P.; Sara, S. Consolidation of memory after its reactivation: Involvement of beta noradrenergic receptors in the late phase. Neural Plast. 1998, 6, 63–68. [Google Scholar] [CrossRef]
- Milton, A.L.; Lee, J.L.C.; Everitt, B.J. Reconsolidation of appetitive memories for both natural and drug reinforcement is dependent on beta-adrenergic receptors. Learn. Mem. 2008, 15, 88–92. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sofuoglu, M.; Sewell, R.A. Norepinephrine and stimulant addiction. Addict. Biol. 2009, 14, 119–129. [Google Scholar] [CrossRef]
- Ramos, B.P.; Arnsten, A.F.T. Adrenergic pharmacology and cognition: Focus on the prefrontal cortex. Pharmacol. Ther. 2007, 113, 523–536. [Google Scholar] [CrossRef] [PubMed]
- Hoeffer, C.A.; Klann, E. mTOR signaling: At the crossroads of plasticity, memory and disease. Trends Neurosci. 2010, 33, 67–75. [Google Scholar] [CrossRef]
- Neasta, J.; Barak, S.; Hamida, S.B.; Ron, D. mTOR complex 1: A key player in neuroadaptations induced by drugs of abuse. J. Neurochem. 2014, 130, 172–184. [Google Scholar] [CrossRef]
- Tronson, N.C.; Taylor, J.R. Addiction: A drug-induced disorder of memory reconsolidation. Curr. Opin. Neurobiol. 2013, 23, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Das, R.K.; Lawn, W.; Kamboj, S.K. Rewriting the valuation and salience of alcohol-related stimuli via memory reconsolidation. Transl Psychiatry 2015, 5, e645. [Google Scholar] [CrossRef] [PubMed]
- Millan, E.Z.; Milligan-Saville, J.; McNally, G.P. Memory retrieval, extinction, and reinstatement of alcohol seeking. Neurobiol. Learn. Mem. 2013, 101, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Gera, R.; Barak, S.; Schonberg, T. Counterconditioning following memory retrieval diminishes the reinstatement of appetitive memories in humans. Sci. Rep. 2019, 9, 9213. [Google Scholar] [CrossRef]
- Cofresi, R.U.; Lewis, S.M.; Chaudhri, N.; Lee, H.J.; Monfils, M.H.; Gonzales, R.A. Postretrieval extinction attenuates alcohol cue reactivity in rats. Alcohol. Clin. Exp. Res. 2017, 41, 608–617. [Google Scholar] [CrossRef]
- Hutton-Bedbrook, K.; McNally, G.P. The promises and pitfalls of retrieval-extinction procedures in preventing relapse to drug seeking. Front. Psychiatry 2013, 4, 14. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, J.J.; Yu, L.C. Post-retrieval extinction training enhances or hinders the extinction of morphine-induced conditioned place preference in rats dependent on the retrieval-extinction interval. Psychopharmacology 2012, 221, 19–26. [Google Scholar] [CrossRef]
- Sartor, G.C.; Aston-Jones, G. Post-retrieval extinction attenuates cocaine memories. Neuropsychopharmacology 2014, 39, 1059–1065. [Google Scholar] [CrossRef]
- Barak, S.; Ben Hamida, S. Memory erasure, enhanced extinction and disrupted reconsolidation. J. Neurosci. 2012, 32, 2250–2251. [Google Scholar] [CrossRef]
- Luyten, L.; Beckers, T. A preregistered, direct replication attempt of the retrieval-extinction effect in cued fear conditioning in rats. Neurobiol. Learn. Mem. 2017, 144, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Cannon, D.S.; Baker, T.B.; Wehl, C.K. Emetic and electric shock alcohol aversion therapy: Six- and twelve-month follow-up. J. Consult. Clin. Psychol. 1981, 49, 360–368. [Google Scholar] [CrossRef]
- Tunstall, B.J.; Verendeev, A.; Kearns, D.N. A comparison of therapies for the treatment of drug cues: Counterconditioning vs. extinction in male rats. Exp. Clin. Psychopharmacol. 2012, 20, 447–453. [Google Scholar] [CrossRef]
- Van Gucht, D.; Baeyens, F.; Vansteenwegen, D.; Hermans, D.; Beckers, T. Counterconditioning reduces cue-induced craving and actual cue-elicited consumption. Emotion 2010, 10, 688–695. [Google Scholar] [CrossRef]
- Elkins, R.L.; Richards, T.L.; Nielsen, R.; Repass, R.; Stahlbrandt, H.; Hoffman, H.G. The neurobiological mechanism of chemical aversion (emetic) therapy for alcohol use disorder: An fMRI study. Front. Behav. Neurosci. 2017, 11, 182. [Google Scholar] [CrossRef]
- Bouton, M.E.; Peck, C.A. Spontaneous-recovery in cross-motivational transfer (counterconditioning). Anim. Learn. Behav. 1992, 20, 313–321. [Google Scholar] [CrossRef]
- Brooks, D.C.; Hale, B.; Nelson, J.B.; Bouton, M.E. Reinstatement after counterconditioning. Anim. Learn. Behav. 1995, 23, 383–390. [Google Scholar] [CrossRef]
- Van Dis, E.A.; Hagenaars, M.A.; Bockting, C.L.; Engelhard, I.M. Reducing negative stimulus valence does not attenuate the return of fear: Two counterconditioning experiments. Behav. Res. Ther. 2019, 120, 103416. [Google Scholar] [CrossRef]
- Goltseker, K.; Barak, S. Flood-conditioned place aversion as a novel non-pharmacological aversive learning procedure in mice. Sci. Rep. 2018, 8, 7280. [Google Scholar] [CrossRef]
- Gisquet-Verrier, P.; Riccio, D.C. Memory integration: An alternative to the consolidation/reconsolidation hypothesis. Prog. Neurobiol. 2018, 171, 15–31. [Google Scholar] [CrossRef]
- Goltseker, K.; Hopf, F.W.; Barak, S. Advances in behavioral animal models of alcohol use disorder. Alcohol 2019, 74, 73–82. [Google Scholar] [CrossRef]
- Burattini, C.; Gill, T.M.; Aicardi, G.; Janak, P.H. The ethanol self-administration context as a reinstatement cue: Acute effects of naltrexone. Neuroscience 2006, 139, 877–887. [Google Scholar] [CrossRef]
- Das, R.K.; Gale, G.; Hennessy, V.; Kamboj, S.K. A prediction error-driven retrieval procedure for destabilizing AND Rewriting maladaptive reward memories in hazardous drinkers. J. Vis. Exp. 2018, 131, 56097. [Google Scholar] [CrossRef]
- Gale, G.; Walsh, K.; Hennessy, V.E.; Stemerding, L.E.; Ni, K.S.; Thomas, E.; Kamboj, S.K.; Das, R.K. Long-term behavioural rewriting of maladaptive drinking memories via reconsolidation-update mechanisms. Psychol. Med. 2020, 1–11. [Google Scholar] [CrossRef]
- Hon, T.; Das, R.K.; Kamboj, S.K. The effects of cognitive reappraisal following retrieval-procedures designed to destabilize alcohol memories in high-risk drinkers. Psychopharmacology 2016, 233, 851–861. [Google Scholar] [CrossRef]
- Kaag, A.M.; Goudriaan, A.E.; De Vries, T.J.; Pattij, T.; Wiers, R.W. A high working memory load prior to memory retrieval reduces craving in non-treatment seeking problem drinkers. Psychopharmacology 2018, 235, 695–708. [Google Scholar] [CrossRef]
- Xue, Y.X.; Chen, Y.Y.; Zhang, L.B.; Zhang, L.Q.; Huang, G.D.; Sun, S.C.; Deng, J.H.; Luo, Y.X.; Bao, Y.P.; Wu, P.; et al. Selective inhibition of amygdala neuronal ensembles encoding nicotine-associated memories inhibits nicotine preference and relapse. Biol. Psychiatry 2017, 82, 781–793. [Google Scholar] [CrossRef]
- Tedesco, V.; Mutti, A.; Auber, A.; Chiamulera, C. Nicotine-seeking reinstatement is reduced by inhibition of instrumental memory reconsolidation. Behav. Pharmacol. 2014, 25, 725–731. [Google Scholar] [CrossRef]
- Das, R.K.; Hindocha, C.; Freeman, T.P.; Lazzarino, A.I.; Curran, H.V.; Kamboj, S.K. Assessing the translational feasibility of pharmacological drug memory reconsolidation blockade with memantine in quitting smokers. Psychopharmacology 2015, 232, 3363–3374. [Google Scholar] [CrossRef]
- Pachas, G.N.; Gilman, J.; Orr, S.P.; Hoeppner, B.; Carlini, S.V.; Grasser, E.B.; Loebl, T.; Nino, J.; Pitman, R.K.; Evins, A.E. Single dose propranolol does not affect physiologic or emotional reactivity to smoking cues. Psychopharmacology 2015, 232, 1619–1628. [Google Scholar] [CrossRef]
- Forget, B.; Hamon, M.; Thiebot, M.H. Involvement of alpha1-adrenoceptors in conditioned place preference supported by nicotine in rats. Psychopharmacology 2009, 205, 503–515. [Google Scholar] [CrossRef]
- Xue, Y.X.; Deng, J.H.; Chen, Y.Y.; Zhang, L.B.; Wu, P.; Huang, G.D.; Luo, Y.X.; Bao, Y.P.; Wang, Y.M.; Shaham, Y.; et al. Effect of selective inhibition of reactivated nicotine-associated memories with propranolol on nicotine craving. JAMA Psychiatry 2017, 74, 224–232. [Google Scholar] [CrossRef]
- Briggs, S.B.; Hafenbreidel, M.; Young, E.J.; Rumbaugh, G.; Miller, C.A. The role of nonmuscle myosin II in polydrug memories and memory reconsolidation. Learn. Mem. 2018, 25, 391–398. [Google Scholar] [CrossRef]
- Fang, Q.; Li, F.Q.; Li, Y.Q.; Xue, Y.X.; He, Y.Y.; Liu, J.F.; Lu, L.; Wang, J.S. Cannabinoid CB1 receptor antagonist rimonabant disrupts nicotine reward-associated memory in rats. Pharmacol. Biochem. Behav. 2011, 99, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Zipori, D.; Sadot-Sogrin, Y.; Goltseker, K.; Even-Chen, O.; Rahamim, N.; Shaham, O.; Barak, S. Re-exposure to nicotine-associated context from adolescence enhances alcohol intake in adulthood. Sci. Rep. 2017, 7, 2479. [Google Scholar] [CrossRef]
- Struik, R.F.; De Vries, T.J.; Peters, J. Detrimental effects of a retrieval-extinction procedure on nicotine seeking, but not cocaine seeking. Front. Behav. Neurosci. 2019, 13, 243. [Google Scholar] [CrossRef]
- Germeroth, L.J.; Carpenter, M.J.; Baker, N.L.; Froeliger, B.; LaRowe, S.D.; Saladin, M.E. Effect of a brief memory updating intervention on smoking behavior: A randomized clinical trial. JAMA Psychiatry 2017, 74, 214–223. [Google Scholar] [CrossRef]
- Luyten, L.; Schnell, A.E.; Schroyens, N.; Beckers, T. Lack of drug-induced post-retrieval amnesia for auditory fear memories in rats. BMC Biol. 2021, 19, 17. [Google Scholar] [CrossRef]
- Schroyens, N.; Sigwald, E.L.; Van Den Noortgate, W.; Beckers, T.; Luyten, L. Reactivation-dependent amnesia for contextual fear memories: Evidence for publication bias. eNeuro 2021, 8. [Google Scholar] [CrossRef]
- Brown, T.E.; Lee, B.R.; Sorg, B.A. The NMDA antagonist MK-801 disrupts reconsolidation of a cocaine-associated memory for conditioned place preference but not for self-administration in rats. Learn. Mem. 2008, 15, 857–865. [Google Scholar] [CrossRef]
- Hernandez, P.J.; Kelley, A.E. Long-term memory for instrumental responses does not undergo protein synthesis-dependent reconsolidation upon retrieval. Learn. Mem. 2004, 11, 748–754. [Google Scholar] [CrossRef]
- Mierzejewski, P.; Korkosz, A.; Rogowski, A.; Korkosz, I.; Kostowski, W.; Scinska, A. Post-session injections of a protein synthesis inhibitor, cycloheximide do not alter saccharin self-administration. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 286–289. [Google Scholar] [CrossRef]
- Piva, A.; Pintori, N.; Padovani, L.; Chiamulera, C. Protocols for instrumental memory reconsolidation in rodents: A methodological review. J. Neurosci. Methods 2020, 342, 108766. [Google Scholar] [CrossRef]
- Duvarci, S.; Nader, K. Characterization of fear memory reconsolidation. J. Neurosci. 2004, 24, 9269–9275. [Google Scholar] [CrossRef] [PubMed]
- Monfils, M.H.; Holmes, E.A. Memory boundaries: Opening a window inspired by reconsolidation to treat anxiety, trauma-related, and addiction disorders. Lancet Psychiatry 2018, 5, 1032–1042. [Google Scholar] [CrossRef]
- Milton, A.L. Drink, drugs and disruption: Memory manipulation for the treatment of addiction. Curr. Opin. Neurobiol. 2013, 23, 706–712. [Google Scholar] [CrossRef]
- Treanor, M.; Brown, L.A.; Rissman, J.; Craske, M.G. Can memories of traumatic experiences or addiction be erased or modified? A critical review of research on the disruption of memory reconsolidation and its applications. Perspect. Psychol. Sci. 2017, 12, 290–305. [Google Scholar] [CrossRef]
- Cappell, H.; LeBlanc, A.; Endrenyi, L. Aversive conditioning by psychoactive drugs: Effects of morphine, alcohol and chlordiazepoxide. Psychopharmacologia 1973, 29, 239–246. [Google Scholar] [CrossRef]
- Shram, M.J.; Funk, D.; Li, Z.; Lê, A.D. Periadolescent and adult rats respond differently in tests measuring the rewarding and aversive effects of nicotine. Psychopharmacology 2006, 186, 201–208. [Google Scholar] [CrossRef]
- Carnicella, S.; Ron, D.; Barak, S. Intermittent ethanol access schedule in rats as a preclinical model of alcohol abuse. Alcohol 2014, 48, 243–252. [Google Scholar] [CrossRef]
- Jobes, M.L.; Aharonovich, E.; Epstein, D.H.; Phillips, K.A.; Reamer, D.; Anderson, M.; Preston, K.L. Effects of prereactivation propranolol on cocaine craving elicited by imagery script/cue sets in opioid-dependent polydrug users: A randomized study. J. Addict. Med. 2015, 9, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Kindt, M.; Soeter, M.; Vervliet, B. Beyond extinction: Erasing human fear responses and preventing the return of fear. Nat. Neurosci. 2009, 12, 256. [Google Scholar] [CrossRef] [PubMed]
- Soeter, M.; Kindt, M. Dissociating response systems: Erasing fear from memory. Neurobiol. Learn. Mem. 2010, 94, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Cogan, E.S.; Shapses, M.A.; Robinson, T.E.; Tronson, N.C. Disrupting reconsolidation: Memory erasure or blunting of emotional/motivational value? Neuropsychopharmacology 2019, 44, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Marchant, N.J.; Campbell, E.J.; Kaganovsky, K. Punishment of alcohol-reinforced responding in alcohol preferring P rats reveals a bimodal population: Implications for models of compulsive drug seeking. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 87 Pt A, 68–77. [Google Scholar] [CrossRef]
- Marchant, N.J.; Kaganovsky, K. A critical role of nucleus accumbens dopamine D1-family receptors in renewal of alcohol seeking after punishment-imposed abstinence. Behav. Neurosci. 2015, 129, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Marchant, N.J.; Campbell, E.J.; Pelloux, Y.; Bossert, J.M.; Shaham, Y. Context-induced relapse after extinction versus punishment: Similarities and differences. Psychopharmacology 2019, 236, 439–448. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Pharmacological Manipulations | ||||||
| NMDA-Receptor Antagonists | ||||||
| Species | Procedure | Retrieval | Treatment relative to retrieval | Treatment | Results | Reference |
| Rats | OSA | 5 min extinction | Immediately after | MK801 (0.1 mg/kg, ip) | +? | [26] |
| Rats | OSA | 5 min extinction | Immediately after | Acamprosate (200 mg/kg, ip) | - | [26] |
| Rats | OSA | 20 min of cues only | Immediately after | MK801 (0.1 mg/kg, ip) | +? | [32] |
| Rats | OSA | 10–25 cues only | 30 min before | MK801 (0.1 mg/kg, ip) | + | [28] |
| Rats | OSA | 5 min extinction | Immediately after | Memantine (20 mg/kg, ip) | ? | [31] |
| Rats | OSA | 30 extinction trials | 30 min before | MK801 (0.1 mg/kg, ip) | - | [30] |
| Humans | Visual and olfactory cues | 5 min after | Ketamine (350 ng/mL, 30 min, iv) | ++ | [29] | |
| β-adrenergic receptors antagonists | ||||||
| Species | Procedure | Retrieval | Treatment relative to retrieval | Treatment | Results | Reference |
| Rats | OSA | 20 min extinction | Immediately after | Propranolol (10 mg/kg, ip) | + | [32] |
| Rats | OSA | 10–25 cues only | 30 min before | Propranolol (10 mg/kg, ip) | - | [28] |
| Mice | CPP | 15 min extinction | Immediately after | Propranolol (10 or 30 mg/kg, ip) | - | [35] |
| Rats | OSA | 15 min extinction | 10–30 min before | Propranolol (10 mg/kg, ip) | + | [33] |
| Rats | OSA | 15 min extinction | 10–30 min before | Nadolol (peripheral effect) (20 mg/kg, ip) | - | [33] |
| Rats | OSA | 15 min extinction | 10–30 min before | Dipivefrine (adrenergic prodrug) (10 µg/kg, ip) | +* | [33] |
| Rats | OSA | A training session | Immediately after | Propranolol (0.2 µg/µL, BLA) | +? | [34] |
| Humans | Recap of alcohol memories | 1 h before | Propranolol (1 mg/kg, po) | + | [36] | |
| Protein synthesis inhibitors | ||||||
| Species | Procedure | Retrieval | Treatment relative to retrieval | Treatment | Results | Reference |
| Rats | OSA | 5 min extinction | Immediately after | Anisomycin (400 µg, icv) | ++ | [26] |
| Rats | OSA | 5 min extinction | Immediately after | Rapamycin (20 mg/kg, ip; 50 µg/side, cea) | ++ | [14] |
| Rats | OSA | 5 min extinction | Immediately after | Anisomycin (62.5 µg/side, cea) | + | [14] |
| Rats | CPP | 10 min extinction | Immediately after | Rapamycin (0.1 or 10 mg/kg, ip) | ++ | [27] |
| Behavioral manipulations | ||||||
| Species | Procedure | Retrieval | Treatment relative to retrieval | Treatment | Results | Reference |
| Rats | OSA | 10 min extinction | 70 min after | Extinction | ? | [55] |
| Rats | Cue reactivity | Exposure to alcohol cues | 1 h after | Extinction | + | [57] |
| Mice | CPP | 3 min extinction | 45 min after | CC with water-flood | + | [25] |
| Rats | OSA | Odor-taste cues in home-cages | 45 min after | Punishment with footshocks | + | [25] |
| Humans | Visual and olfactory cues | 10 min after | CC with disgust | + | [54,74] | |
| Humans | Visual and olfactory cues | 10 min after | CC with disgust | ++ | [75] | |
| Humans | Visual and olfactory cues | 10 min after | Alcohol memory reappraisal | + | [76] | |
| Humans | Recap of alcohol memories | Immediately before | High working memory load task | + | [77] | |
| Humans | Recap of alcohol memories | Immediately after | High working memory load task | - | [77] | |
| Pharmacological Manipulations | ||||||
| Species | Procedure | Retrieval | Treatment Relative to Retrieval | Treatment | Results | Reference |
| Rats | OSA | 20 extinction trials | 30 min before | MK-801 (0.1 mg/kg, ip) | - | [79] |
| Rats | OSA | 20 extinction trials | 1 h after | MK-801 (0.1 mg/kg, ip) | + | [79] |
| Mice | CPP | 15 min extinction | 30 min before | Blebbistatin (nonmuscle myosin II (NMII) inhibitor) (10 mg/kg, ip) | + | [84] |
| Rats | CPP | 10 min extinction | Immediately after | Rimonabant (CB1 ant.) (0.3, 3 mg/kg, ip) | + | [85] |
| Rats | CPP, OSA | 5 min extinction or nicotine | Immediately after | Anisomycin (synthesis inhibitor) (400 µg, icv) | + | [78] |
| Rats | CPP, OSA | 5 min extinction or nicotine | Immediately after | Intra-BLA neuronal ensembles inactivation (with Daun02) | + | [78] |
| Rats | CPP | 20 min extinction | 30 min before | Prazosin (α1-adrenergic antagonist) (0.125–1 mg/kg, ip) | ++ | [82] |
| Rats | CPP | 20 min extinction | Immediately after | Prazosin (α1-adrenergic antagonist) (0.125–1 mg/kg, ip) | - | [82] |
| Rats | CPP, OSA | 5 min extinction or nicotine | Immediately after | Propranolol (10 mg/kg, ip) | + | [83] |
| Humans | 2 cigarette puffs | 1 h before | Propranolol (40 mg, po) | + | [83] | |
| Humans | Recap of smoking experience | Immediately before | Propranolol (1 mg/kg, po) | - | [81] | |
| Humans | Smoking cues and video | Memantine (10 mg, po) | - | [80] | ||
| Behavioral manipulations | ||||||
| Species | Procedure | Retrieval | Treatment relative to retrieval | Treatment | Results | Reference |
| Rats | OSA | 10 min extinction | 10 min after | 60 min extinction | +* | [87] |
| Humans | Smoking cues and video | 10 min after | 60 min extinction | ++ | [88] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barak, S.; Goltseker, K. Targeting the Reconsolidation of Licit Drug Memories to Prevent Relapse: Focus on Alcohol and Nicotine. Int. J. Mol. Sci. 2021, 22, 4090. https://doi.org/10.3390/ijms22084090
Barak S, Goltseker K. Targeting the Reconsolidation of Licit Drug Memories to Prevent Relapse: Focus on Alcohol and Nicotine. International Journal of Molecular Sciences. 2021; 22(8):4090. https://doi.org/10.3390/ijms22084090
Chicago/Turabian StyleBarak, Segev, and Koral Goltseker. 2021. "Targeting the Reconsolidation of Licit Drug Memories to Prevent Relapse: Focus on Alcohol and Nicotine" International Journal of Molecular Sciences 22, no. 8: 4090. https://doi.org/10.3390/ijms22084090
APA StyleBarak, S., & Goltseker, K. (2021). Targeting the Reconsolidation of Licit Drug Memories to Prevent Relapse: Focus on Alcohol and Nicotine. International Journal of Molecular Sciences, 22(8), 4090. https://doi.org/10.3390/ijms22084090