
Molecular and Biochemical Basis of Minocycline-Induced Hyperpigmentation—The Study on Normal Human Melanocytes Exposed to UVA and UVB Radiation

 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Minocycline Inhibits the Proliferation of Human Melanocytes
2.2. The Evaluation of Cell Number, Viability and Morphology of Human Melanocytes Treated with Minocycline and Exposed to UVA and UVB Radiation
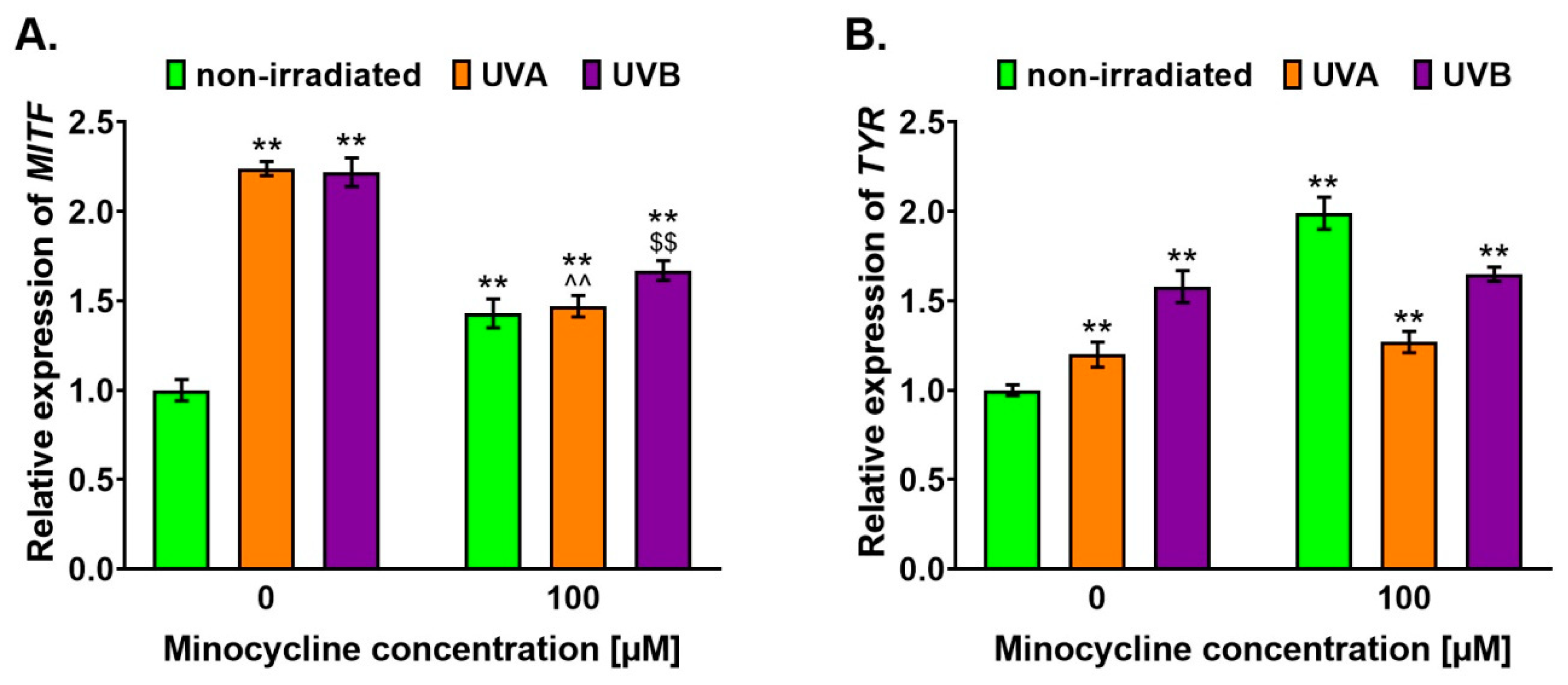
2.3. Minocycline Stimulates the Expression of MITF and TYR Genes in Melanocytes Exposed to UVA and UVB Radiation
2.4. Minocycline Increases Melanin Level in Human Melanocytes Irradiated with UVA and UVB
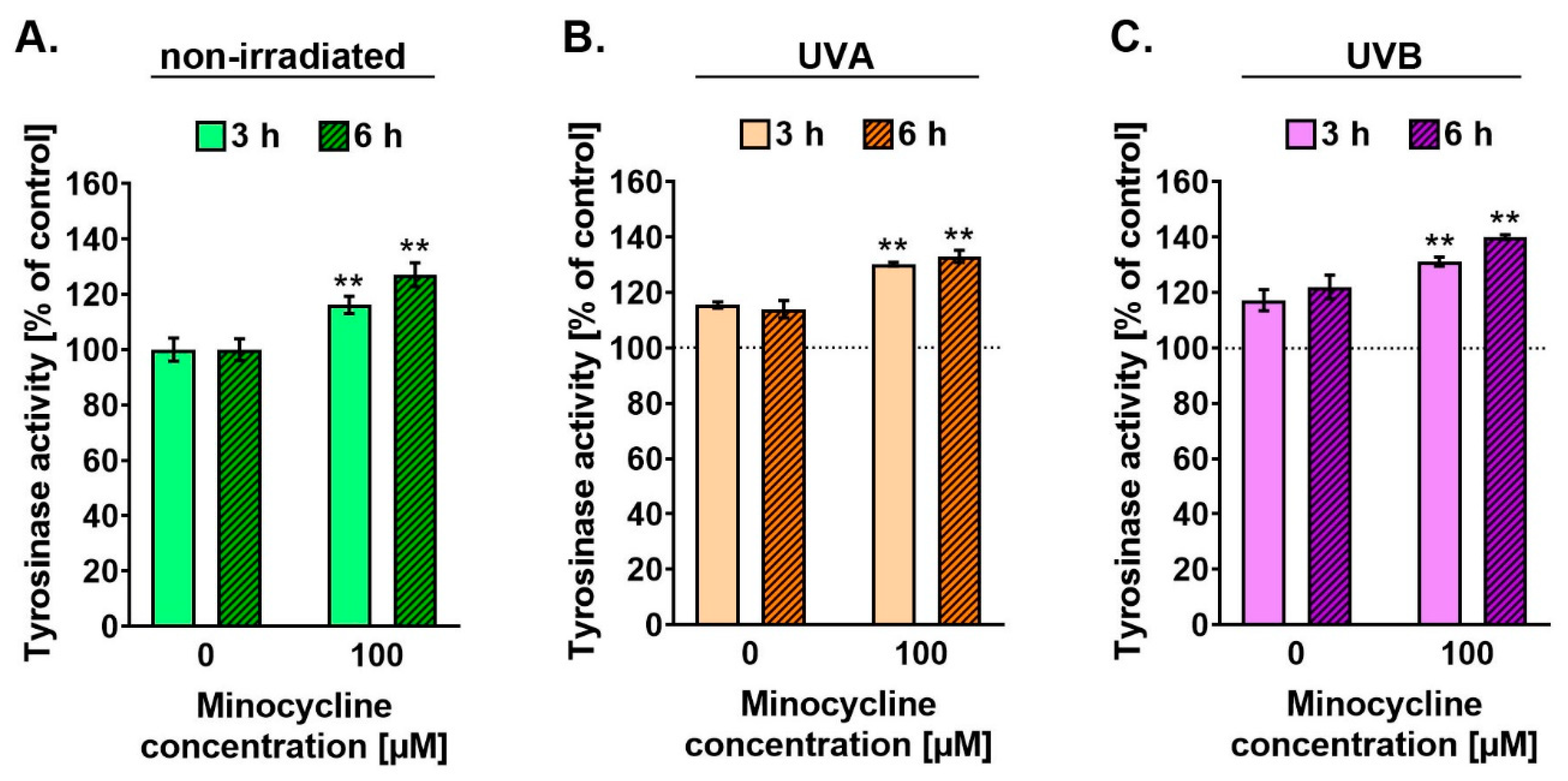
2.5. Minocycline Stimulates the Activity of Tyrosinase in Human Melanocytes Irradiated with UVA and UVB
2.6. Minocycline Increases Level of MITF and Tyrosinase but Decreases Level of Pmel17 in Human Melanocytes Exposed to UVA and UVB Radiation
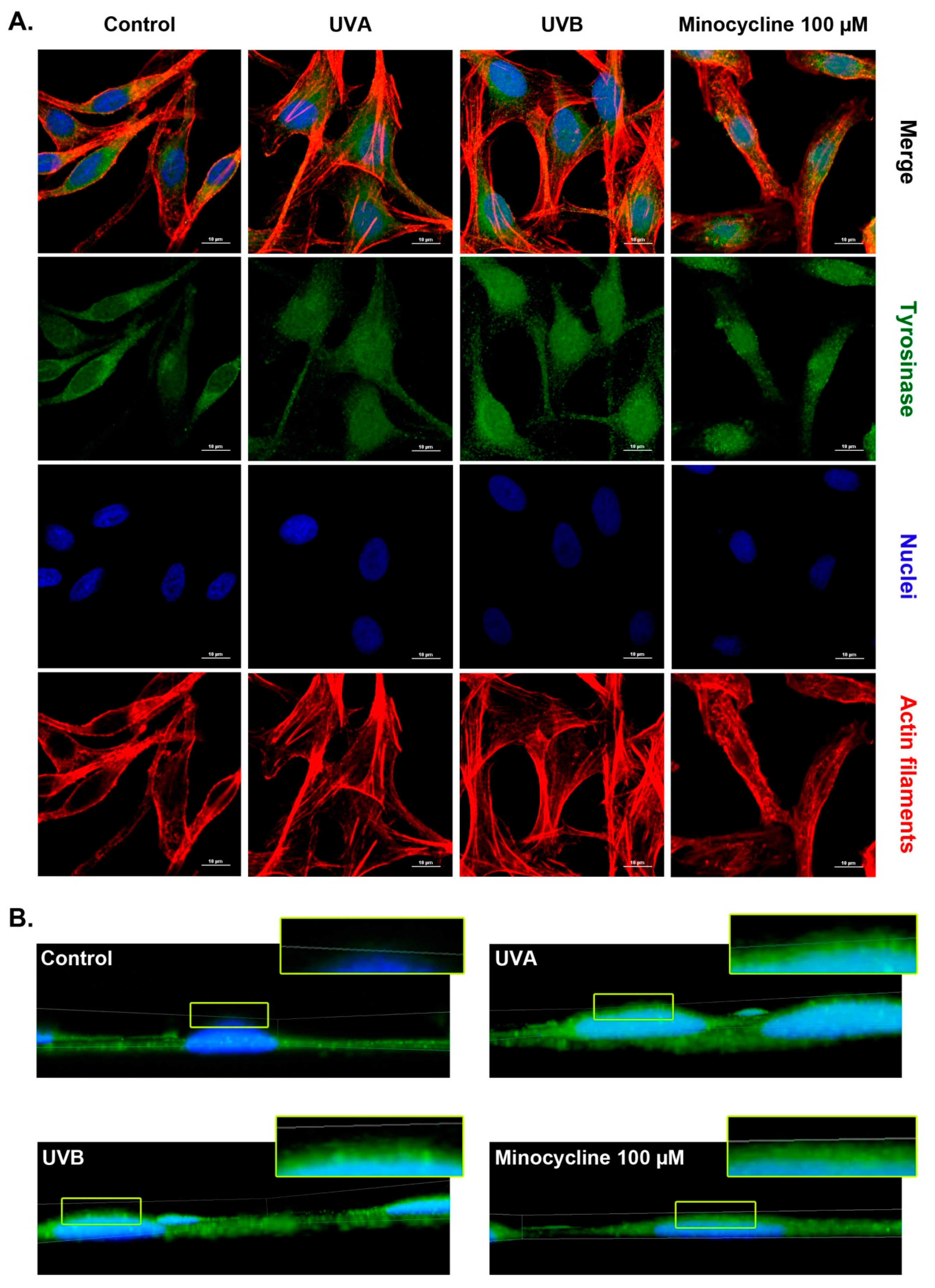
2.7. Confocal Microscopy Analysis of Tyrosinase in Human Melanocytes Treated with Minocycline and Exposed to UVA and UVB Radiation
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Cell Culture
5.3. Melanocyte Treatment
5.4. Screening Test of Cells Proliferation
5.5. The Assessment of Cell Number and Cell Viability
5.6. Real-Time Quantitative PCR Analysis
5.7. Preparation of Cell Lysates
5.8. The Analysis of Protein Concentration
5.9. The Analysis of Melanin Content
5.10. The Analysis of Tyrosinase Activity
5.11. Western Blotting Analysis
5.12. Microscopic Assessment of Melanocytes
5.13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nayak, S.; Acharjya, B. Adverse cutaneous drug reaction. Indian J. Dermatol. 2008, 53, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Carr, D.F.; Pirmohamed, M. Biomarkers of adverse drug reactions. Exp. Biol. Med. 2018, 243, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Bouvy, J.C.; De Bruin, M.L.; Koopmanschap, M.A. Epidemiology of adverse drug reactions in Europe: A review of recent observational studies. Drug Saf. 2015, 38, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Gerogianni, K.; Tsezou, A.; Dimas, K. Drug-induced skin adverse reactions: The role of pharmacogenomics in their prevention. Mol. Diagn. Ther. 2018, 22, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Kiepurska, N.; Paluchowska, E.; Owczarek, W.; Szkultecka-Dębek, M.; Jahnz-Różyk, K. The direct costs of drug-induced skin reactions. Ann. Agric. Environ. Med. 2017, 24, 190–193. [Google Scholar] [CrossRef]
- Giménez García, R.M.; Carrasco Molina, S. Drug-induced hyperpigmentation: Review and case series. J. Am. Board Fam. Med. 2019, 32, 628–638. [Google Scholar] [CrossRef]
- Dereure, O. Drug-induced skin pigmentation. Epidemiology, diagnosis and treatment. Am. J. Clin. Dermatol. 2001, 2, 253–262. [Google Scholar] [CrossRef]
- Nahhas, A.F.; Braunberger, T.L.; Hamzavi, I.H. An update on drug-induced pigmentation. Am. J. Clin. Dermatol. 2019, 20, 75–96. [Google Scholar] [CrossRef]
- Nelson, M.L.; Levy, S.B. The history of the tetracyclines. Ann. N. Y. Acad. Sci. 2011, 1241, 17–32. [Google Scholar] [CrossRef]
- Agwuh, K.N.; MacGowan, A. Pharmacokinetics and pharmacodynamics of the tetracyclines including glycylcyclines. J. Antimicrob. Chemother. 2006, 58, 256–265. [Google Scholar] [CrossRef]
- Domercq, M.; Matute, C. Neuroprotection by tetracyclines. Trends Pharmacol. Sci. 2004, 25, 609–612. [Google Scholar] [CrossRef]
- Asadi, A.; Abdi, M.; Kouhsari, E.; Panahi, P.; Sholeh, M.; Sadeghifard, N.; Amiriani, T.; Ahmadi, A.; Maleki, A.; Gholami, M. Minocycline, focus on mechanisms of resistance, antibacterial activity, and clinical effectiveness: Back to the future. J. Glob. Antimicrob. Resist. 2020, 22, 161–174. [Google Scholar] [CrossRef]
- Garrido-Mesa, N.; Zarzuelo, A.; Gálvez, J. What is behind the non-antibiotic properties of minocycline? Pharmacol. Res. 2013, 67, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Mesa, N.; Zarzuelo, A.; Gálvez, J. Minocycline: Far beyond an antibiotic. Br. J. Pharmacol. 2013, 169, 337–352. [Google Scholar] [CrossRef]
- Plane, J.M.; Shen, Y.; Pleasure, D.E.; Deng, W. Prospects for minocycline neuroprotection. Arch. Neurol. 2010, 67, 1442–1448. [Google Scholar] [CrossRef] [PubMed]
- Brenes-Salazar, J.A. Minocycline: A bacteriostatic antibiotic with pleiotropic cardioprotective effects. Can. J. Physiol. Pharmacol. 2015, 93, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Quinn, B.A.; Dash, R.; Sarkar, S.; Azab, B.; Bhoopathi, P.; Das, S.K.; Emdad, L.; Wei, J.; Pellecchia, M.; Sarkar, D.; et al. Pancreatic cancer combination therapy using a BH3 mimetic and a synthetic tetracycline. Cancer Res. 2015, 75, 2305–2315. [Google Scholar] [CrossRef]
- Zhou, Y.Q.; Liu, D.Q.; Chen, S.P.; Sun, J.; Wang, X.M.; Tian, Y.K.; Wu, W.; Ye, D.W. Minocycline as a promising therapeutic strategy for chronic pain. Pharmacol. Res. 2018, 134, 305–310. [Google Scholar] [CrossRef]
- Chen, X.; Ma, X.; Jiang, Y.; Pi, R.; Liu, Y.; Ma, L. The prospects of minocycline in multiple sclerosis. J. Neuroimmunol. 2011, 235, 1–8. [Google Scholar] [CrossRef]
- Chen, Y.; Cai, Z.; Ke, Z. Antineuroinflammation of minocycline in stroke. Neurologist 2017, 22, 120–126. [Google Scholar] [CrossRef]
- Sasaki, S.; Kato, M.; Nakamura, K.; Namba, Y.; Nagashima, O.; Takahashi, K. Management of skin sarcoidosis with minocycline monotherapy. Respirol. Case Rep. 2019, 7, e00413. [Google Scholar] [CrossRef]
- Dabas, G.; Guliani, A.; Vinay, K.; Radotra, B.D. Nevoid hyperkeratosis of male breast: Successful treatment with minocycline. Dermatol. Ther. 2019, 32, e13019. [Google Scholar] [CrossRef] [PubMed]
- Dean, O.M.; Data-Franco, J.; Giorlando, F.; Berk, M. Minocycline: Therapeutic potential in psychiatry. CNS Drugs 2012, 26, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Fiscus, V.; Hankinson, A.; Alweis, R. Minocycline-induced hyperpigmentation. J. Community Hosp. Intern. Med. Perspect. 2014, 4. [Google Scholar] [CrossRef]
- Berbari, H.; Berbari, E.F.; Sia, I.G. Diffuse skin hyperpigmentation associated with chronic minocycline use in a patient with prosthetic joint infection. IDCases. 2016, 7, 30–31. [Google Scholar] [CrossRef] [PubMed]
- Krause, W. Drug-induced hperpigemntation: A systematic review. J. Dtsch. Dermatol. Ges. 2013, 11, 644–651. [Google Scholar] [CrossRef]
- Shute, L.; Walkty, A.; Embil, J.M. Minocycline-induced cutaneous hyperpigmentation. CMAJ 2020, 192, E981. [Google Scholar] [CrossRef]
- Geria, A.N.; Tajirian, A.L.; Kihiczak, G.; Schwartz, R.A. Minocycline-induced skin pigmentation: An update. Acta Dermatovenerol. Croat. 2009, 17, 123–126. [Google Scholar]
- Banning, T.P.; Heard, C.M. Binding of doxycycline to keratin, melanin and human epidermal tissue. Int. J. Pharm. 2002, 235, 219–227. [Google Scholar] [CrossRef]
- Rok, J.; Rzepka, Z.; Respondek, M.; Beberok, A.; Wrześniok, D. Chlortetracycline and melanin biopolymer—The risk of accumulation and implications for phototoxicity: An in vitro study on normal human melanocytes. Chem. Biol. Interact. 2019, 303, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef]
- ElObeid, A.S.; Kamal-Eldin, A.; Abdelhalim, M.A.K.; Haseeb, A.M. Pharmacological properties of melanin and its function in health. Basic Clin. Pharmacol. Toxicol. 2017, 120, 515–522. [Google Scholar] [CrossRef]
- Solano, F. Photoprotection and skin pigmentation: Melanin-related molecules and some other new agents obtained from natural sources. Molecules 2020, 25, 1537. [Google Scholar] [CrossRef]
- Hearing, V.J. Determination of melanin synthetic pathways. J. Investig. Dermatol. 2011, 131, E8–E11. [Google Scholar] [CrossRef] [PubMed]
- Solano, F. Melanins: Skin pigments and much more-Types, structural models, biological functions, and formation routes. New J. Sci. 2014, 2014, 498276. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Maddodi, N.; Jayanthy, A.; Setaluri, V. Shining light on skin pigmentation: The darker and the brighter side of effects of UV radiation. Photochem. Photobiol. 2012, 88, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV light touches the brain and endocrine system through skin, and why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Gaddameedhi, S. Solar ultraviolet-induced DNA damage response: Melanocytes story in transformation to environmental melanomagenesis. Environ. Mol. Mutagen. 2020, 61, 736–751. [Google Scholar] [CrossRef]
- Emri, G.; Paragh, G.; Tósaki, Á.; Janka, E.; Kollár, S.; Hegedűs, C.; Gellén, E.; Horkay, I.; Koncz, G.; Remenyik, É. Ultraviolet radiation-mediated development of cutaneous melanoma: An update. J. Photochem. Photobiol. B 2018, 185, 169–175. [Google Scholar] [CrossRef]
- Nelson, M.L. Chemical and biological dynamics of tetracyclines. Adv. Dent. Res. 1998, 12, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Li, A.S.; Chignell, C.F.; Hall, R.D. Cutaneous phototoxicity of tetracycline antibiotics: Generation of free radicals and singlet oxygen during photolysis as measured by spin-trapping and the phosphorescence of singlet molecular oxygen. Photochem. Photobiol. 1987, 46, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Hasan, T.; Khan, A.U. Phototoxicity of the tetracyclines: Photosensitized emission of singlet delta dioxygen. Proc. Natl. Acad. Sci. USA 1986, 83, 4604–4606. [Google Scholar] [CrossRef]
- Rok, J.; Rzepka, Z.; Maszczyk, M.; Beberok, A.; Wrześniok, D. Minocycline impact on redox homeostasis of normal human melanocytes HEMn-LP exposed to UVA radiation and hydrogen peroxide. Int. J. Mol. Sci. 2021, 22, 1642. [Google Scholar] [CrossRef] [PubMed]
- Rok, J.; Buszman, E.; Beberok, A.; Delijewski, M.; Otręba, M.; Wrześniok, D. Modulation of melanogenesis and antioxidant status of melanocytes in response to phototoxic action of doxycycline. Photochem. Photobiol. 2015, 91, 1429–1434. [Google Scholar] [CrossRef] [PubMed]
- Rok, J.; Wrześniok, D.; Beberok, A.; Otręba, M.; Delijewski, M.; Buszman, E. Phototoxic effect of oxytetracycline on normal human melanocytes. Toxicol. In Vitro 2018, 48, 26–32. [Google Scholar] [CrossRef]
- Rok, J.; Buszman, E.; Delijewski, M.; Otręba, M.; Beberok, A.; Wrześniok, D. Effect of tetracycline and UV radiation on melanization and antioxidant status of melanocytes. J. Photochem. Photobiol. B 2015, 148, 168–173. [Google Scholar] [CrossRef]
- Kawakami, A.; Fisher, D.E. The master role of microphthalmia-associated transcription factor in melanocyte and melanoma biology. Lab. Investig. 2017, 97, 649–656. [Google Scholar] [CrossRef]
- Jang, E.J.; Shin, Y.; Park, H.J.; Kim, D.; Jung, C.; Hong, J.Y.; Kim, S.; Lee, S.K. Anti-melanogenic activity of phytosphingosine via the modulation of the microphthalmia-associated transcription factor signaling pathway. J. Dermatol. Sci. 2017, 87, 19–28. [Google Scholar] [CrossRef]
- Seo, E.Y.; Jin, S.P.; Sohn, K.C.; Park, C.H.; Lee, D.H.; Chung, J.H. UCHL1 regulates melanogenesis through controlling MITF stability in human melanocytes. J. Investig. Dermatol. 2017, 137, 1757–1765. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Fisher, D.E. MITF and UV responses in skin: From pigmentation to addiction. Pigment Cell Melanoma Res. 2019, 32, 224–236. [Google Scholar] [CrossRef]
- Wicks, N.L.; Chan, J.W.; Najera, J.A.; Ciriello, J.M.; Oancea, E. UVA phototransduction drives early melanin synthesis in human melanocytes. Curr. Biol. 2011, 21, 1906–1911. [Google Scholar] [CrossRef]
- Bellono, N.W.; Kammel, L.G.; Zimmerman, A.L.; Oancea, E. UV light phototransduction activates transient receptor potential A1 ion channels in human melanocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 2383–2388. [Google Scholar] [CrossRef]
- Bellono, N.W.; Oancea, E. UV light phototransduction depolarizes human melanocytes. Channels 2013, 7, 243–248. [Google Scholar] [CrossRef]
- Bellono, N.W.; Najera, J.A.; Oancea, E. UV light activates a Gαq/11-coupled phototransduction pathway in human melanocytes. J. Gen. Physiol. 2014, 143, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Wichers, H.J.; Soler-Lopez, M.; Dijkstra, B.W. Structure and function of human tyrosinase and tyrosinase-related proteins. Chemistry 2018, 24, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Ebanks, J.P.; Wickett, R.R.; Boissy, R.E. Mechanisms regulating skin pigmentation: The rise and fall of complexion coloration. Int. J. Mol. Sci. 2009, 10, 4066–4087. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Namasivayam, V.; Manickam, M.; Jung, S.H. Inhibitors of melanogenesis: An updated review. J. Med. Chem. 2018, 61, 7395–7418. [Google Scholar] [CrossRef]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [PubMed]
- Bissig, C.; Rochin, L.; van Niel, G. PMEL amyloid fibril formation: The bright steps of pigmentation. Int. J. Mol. Sci. 2016, 17, 1438. [Google Scholar] [CrossRef] [PubMed]
- Hida, T.; Kamiya, T.; Kawakami, A.; Ogino, J.; Sohma, H.; Uhara, H.; Jimbow, K. Elucidation of melanogenesis cascade for identifying pathophysiology and therapeutic approach of pigmentary disorders and melanoma. Int. J. Mol. Sci. 2020, 21, 6129. [Google Scholar] [CrossRef]
- Theos, A.C.; Berson, J.F.; Theos, S.C.; Herman, K.E.; Harper, D.C.; Tenza, D.; Sviderskaya, E.V.; Lamoreux, M.L.; Bennett, D.C.; Raposo, G.; et al. Dual loss of ER export and endocytic signals with altered melanosome morphology in the silver mutation of Pmel17. Mol. Biol. Cell. 2006, 17, 3598–3612. [Google Scholar] [CrossRef]
- Sun, L.; Hu, L.; Zhang, P.; Li, H.; Sun, J.; Wang, H.; Xie, X.; Hu, J. Silencing of PMEL attenuates melanization via activating lysosomes and degradation of tyrosinase by lysosomes. Biochem. Biophys. Res. Commun. 2018, 503, 2536–2542. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Marks, M.S. Melanosomes-dark organelles enlighten endosomal membrane transport. Nat. Rev. Mol. Cell. Biol. 2007, 8, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Hellström, A.R.; Watt, B.; Fard, S.S.; Tenza, D.; Mannström, P.; Narfström, K.; Ekesten, B.; Ito, S.; Wakamatsu, K.; Larsson, J.; et al. Inactivation of Pmel alters melanosome shape but has only a subtle effect on visible pigmentation. PLoS Genet. 2011, 7, e1002285. [Google Scholar] [CrossRef] [PubMed]
- Judson, T.; Mihara, K. Minocycline-induced hyperpigmentation. J. Gen. Intern. Med. 2017, 32, 133. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rivers, J.K.; Zarbafian, M.; Vestvik, B.; Kawamura, S.; Ulmer, M.; Kuritzky, L.A. Minocycline-induced hyperpigmentation: Rapid resolution after 755nm alexandrite picosecond laser treatment. J. Cosmet. Laser Ther. 2020, 22, 96–99. [Google Scholar] [CrossRef]
- Nisar, M.S.; Iyer, K.; Brodell, R.T.; Lloyd, J.R.; Shin, T.M.; Ahmad, A. Minocycline-induced hyperpigmentation: Comparison of 3 Q-switched lasers to reverse its effects. Clin. Cosmet. Investig. Dermatol. 2013, 6, 159–162. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| MITF | CAGTACCTTTCTACCACTTTAG | CCTCTTTTTCACAGTTGGAG |
| TYR | CAACAGCCATCAGTCTTTATG | CCTTCCAGTGTATTTCTAAAGC |
| GAPDH | CTTTTGCGTCGCCAG | TTGATGGCAACAATATCCAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rok, J.; Rzepka, Z.; Kowalska, J.; Banach, K.; Beberok, A.; Wrześniok, D. Molecular and Biochemical Basis of Minocycline-Induced Hyperpigmentation—The Study on Normal Human Melanocytes Exposed to UVA and UVB Radiation. Int. J. Mol. Sci. 2021, 22, 3755. https://doi.org/10.3390/ijms22073755
Rok J, Rzepka Z, Kowalska J, Banach K, Beberok A, Wrześniok D. Molecular and Biochemical Basis of Minocycline-Induced Hyperpigmentation—The Study on Normal Human Melanocytes Exposed to UVA and UVB Radiation. International Journal of Molecular Sciences. 2021; 22(7):3755. https://doi.org/10.3390/ijms22073755
Chicago/Turabian StyleRok, Jakub, Zuzanna Rzepka, Justyna Kowalska, Klaudia Banach, Artur Beberok, and Dorota Wrześniok. 2021. "Molecular and Biochemical Basis of Minocycline-Induced Hyperpigmentation—The Study on Normal Human Melanocytes Exposed to UVA and UVB Radiation" International Journal of Molecular Sciences 22, no. 7: 3755. https://doi.org/10.3390/ijms22073755
APA StyleRok, J., Rzepka, Z., Kowalska, J., Banach, K., Beberok, A., & Wrześniok, D. (2021). Molecular and Biochemical Basis of Minocycline-Induced Hyperpigmentation—The Study on Normal Human Melanocytes Exposed to UVA and UVB Radiation. International Journal of Molecular Sciences, 22(7), 3755. https://doi.org/10.3390/ijms22073755