
Asthma-Associated Long TSLP Inhibits the Production of IgA


 ,
,  and
and 
Abstract
1. Introduction
2. Results
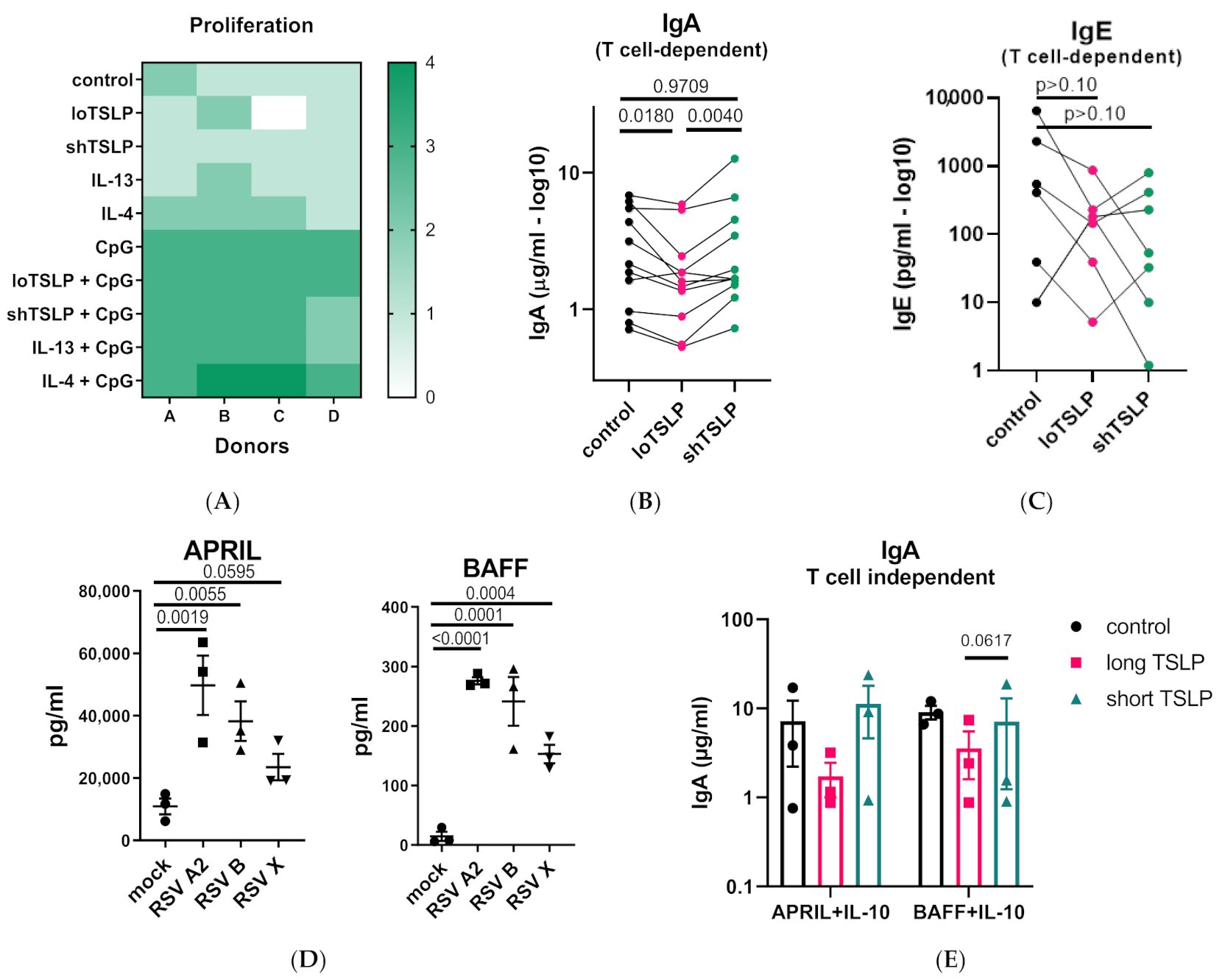
2.1. The Effect of TSLP on IgA Production
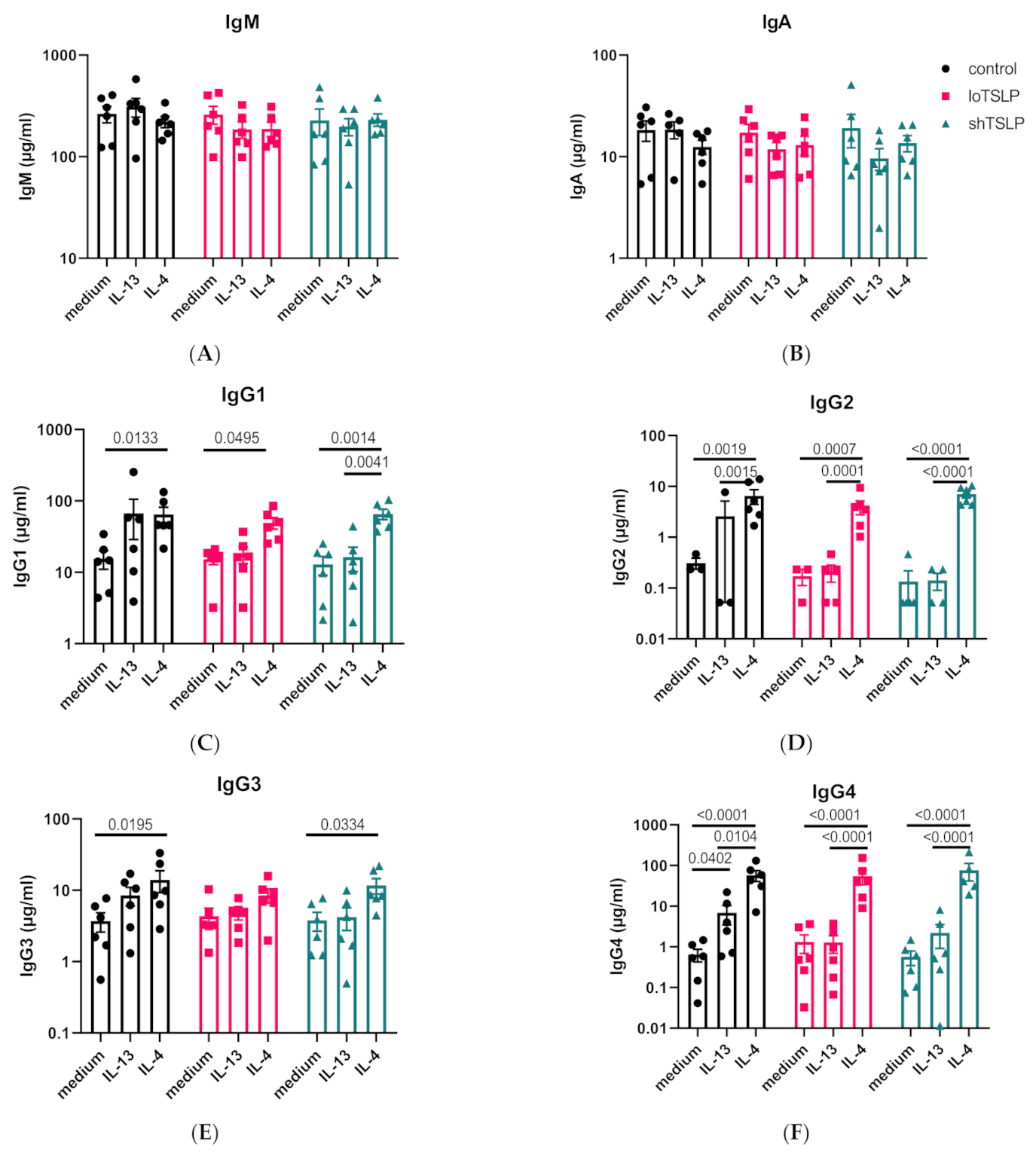
2.2. Regulation of IgA and IgG Production by the Type 2 Cytokines IL-4 and IL-13
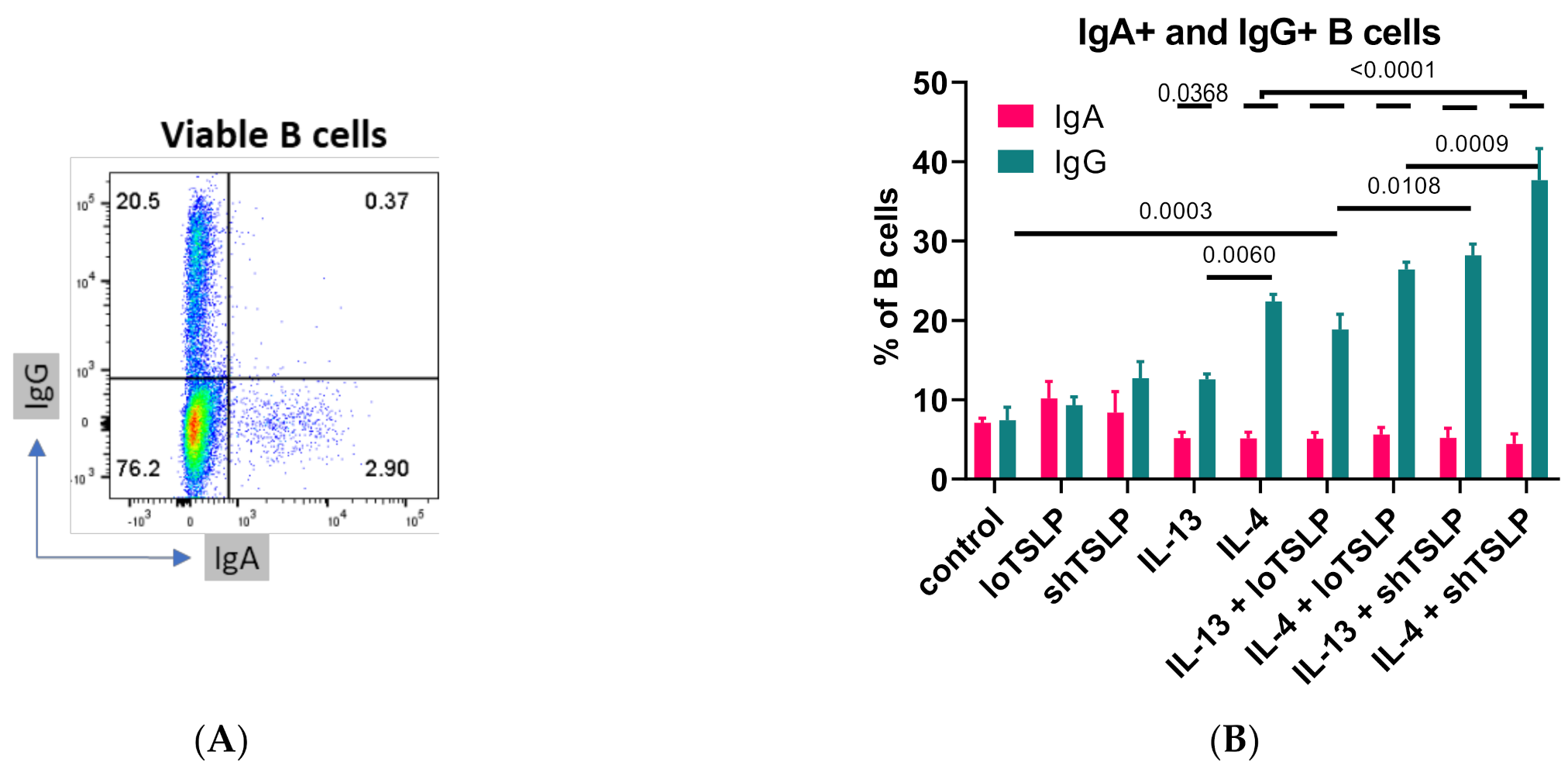
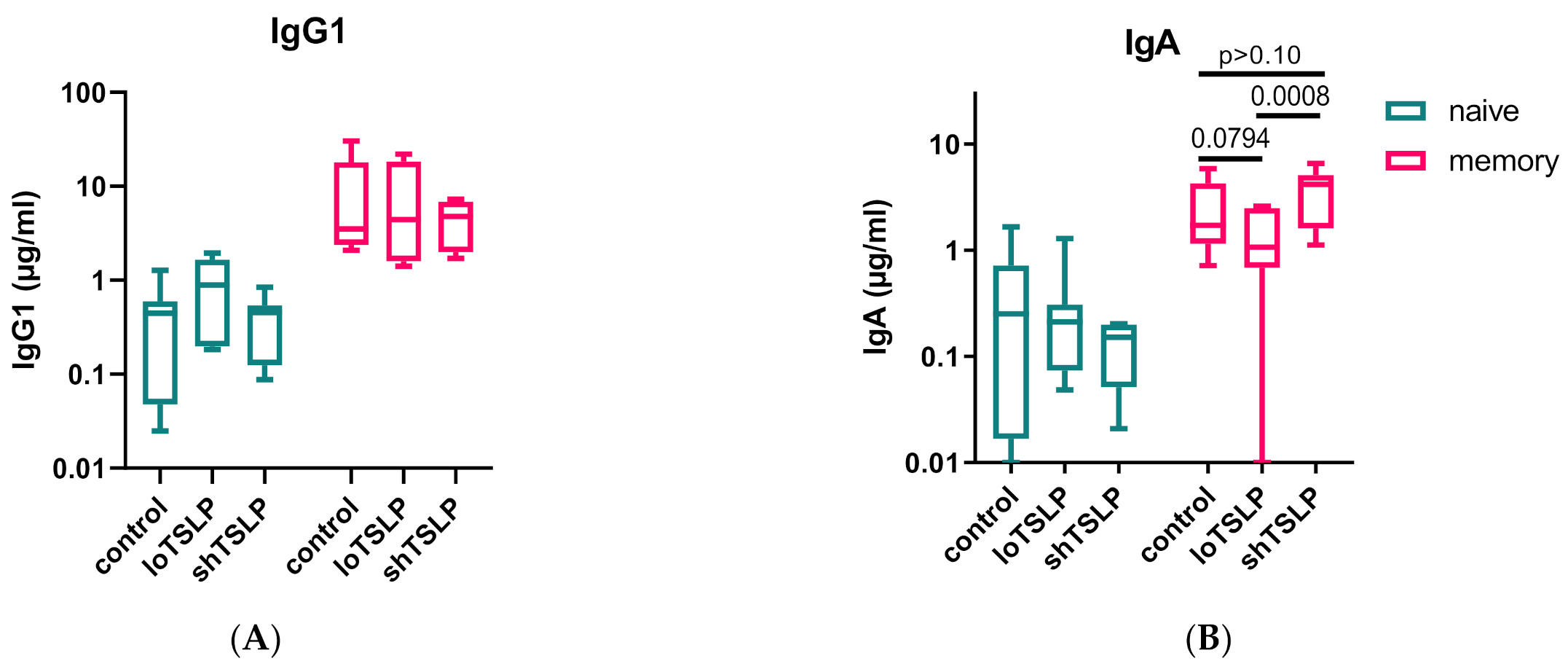
2.3. FACS Sorting Shows That TSLP Regulates IgA Production by Memory B Cells
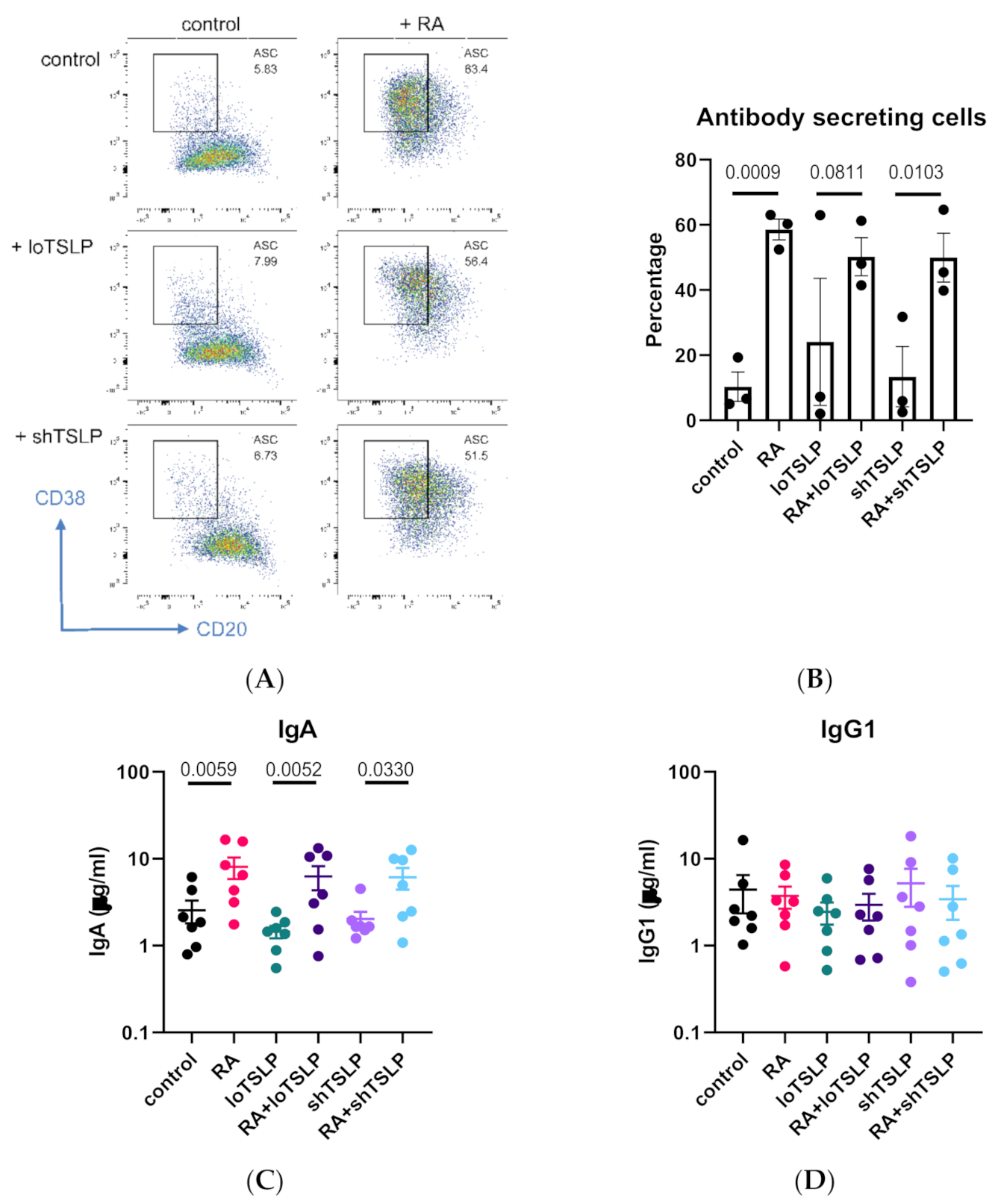
2.4. Restoration of loTSLP-Suppressed IgA Production by the Vitamin A Metabolite Retinoic Acid
3. Discussion
4. Materials and Methods
4.1. B Cell Isolation and Stimulation
4.2. Analysis of APRIL, BAFF and Ig Concentrations
4.3. Flow Cytometry
4.4. Cell Sorting
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uphoff, E.P.; Bird, P.K.; Antó, J.M.; Basterrechea, M.; Von Berg, A.; Bergström, A.; Bousquet, J.; Chatzi, L.; Fantini, M.P.; Ferrero, A.; et al. Variations in the prevalence of childhood asthma and wheeze in MeDALL cohorts in Europe. ERJ Open Res. 2017, 3, 00150–02016. [Google Scholar] [CrossRef] [PubMed]
- Calvén, J.; Ax, E.; Rådinger, M. The Airway Epithelium—A Central Player in Asthma Pathogenesis. Int. J. Mol. Sci. 2020, 21, 8907. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The Cytokines of Asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef]
- Lemanske, R.F.; Busse, W.W. Asthma: Clinical expression and molecular mechanisms. J. Allergy Clin. Immunol. 2010, 125, S95–S102. [Google Scholar] [CrossRef]
- Pullan, C.R.; Hey, E.N. Wheezing, asthma, and pulmonary dysfunction 10 years after infection with respiratory syncytial virus in infancy. BMJ 1982, 284, 1665–1669. [Google Scholar] [CrossRef]
- Corne, J.M.; Marshall, C.; Smith, S.; Schreiber, J.; Sanderson, G.; Holgate, S.T.; Johnston, S.L. Frequency, severity, and duration of rhinovirus infections in asthmatic and non-asthmatic individuals: A longitudinal cohort study. Lancet 2002, 359, 831–834. [Google Scholar] [CrossRef]
- Mohan, A.; Chandra, S.; Agarwal, D.; Guleria, R.; Broor, S.; Gaur, B.; Pandey, R.M. Prevalence of viral infection detected by PCR and RT-PCR in patients with acute exacerbation of COPD: A systematic review. Respirology 2010, 15, 536–542. [Google Scholar] [CrossRef]
- Bagga, B.; Cehelsky, J.E.; Vaishnaw, A.; Wilkinson, T.; Meyers, R.; Harrison, L.M.; Roddam, P.L.; Walsh, E.E.; DeVincenzo, J.P. Effect of Preexisting Serum and Mucosal Antibody on Experimental Respiratory Syncytial Virus (RSV) Challenge and Infection of Adults. J. Infect. Dis. 2015, 212, 1719–1725. [Google Scholar] [CrossRef]
- Berbers, G.; Mollema, L.; Van Der Klis, F.; Hartog, G.D.; Schepp, R. Antibody Responses to Respiratory Syncytial Virus: A Cross-Sectional Serosurveillance Study in the Dutch Population Focusing on Infants Younger Than 2 Years. J. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Habibi, M.S.; Jozwik, A.; Makris, S.; Dunning, J.; Paras, A.; DeVincenzo, J.P.; De Haan, C.A.M.; Wrammert, J.; Openshaw, P.J.M.; Chiu, C. Impaired Antibody-mediated Protection and Defective IgA B-Cell Memory in Experimental Infection of Adults with Respiratory Syncytial Virus. Am. J. Respir. Crit. Care Med. 2015, 191, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Balzar, S.; Strand, M.; Nakano, T.; Wenzel, S.E. Subtle Immunodeficiency in Severe Asthma: IgA and IgG2 Correlate with Lung Function and Symptoms. Int. Arch. Allergy Immunol. 2006, 140, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Hartog, G.D.; Van Neerven, R.J.J.; Boot, J.D.; Jansen, A.P.H.; Savelkoul, H.F.J. House dust mite-specific IgA2 is associated with protection against eczema in allergic patients. Allergy 2016, 71, 563–566. [Google Scholar] [CrossRef] [PubMed]
- MacPherson, A.J.; McCoy, K.D.; Johansen, F.-E.; Brandtzaeg, P. The immune geography of IgA induction and function. Mucosal Immunol. 2007, 1, 11–22. [Google Scholar] [CrossRef]
- Gloudemans, A.K.; Lambrecht, B.N.; Smits, H.H. Potential of Immunoglobulin A to Prevent Allergic Asthma. Clin. Dev. Immunol. 2013, 2013, 1–12. [Google Scholar] [CrossRef]
- Lee, H.-C.; Headley, M.B.; Loo, Y.-M.; Berlin, A.; Gale, M.; Debley, J.S.; Lukacs, N.W.; Ziegler, S.F. Thymic stromal lymphopoietin is induced by respiratory syncytial virus–infected airway epithelial cells and promotes a type 2 response to infection. J. Allergy Clin. Immunol. 2012, 130, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Gauvreau, G.M.; O’Byrne, P.M.; Boulet, L.-P.; Wang, Y.; Cockcroft, D.; Bigler, J.; Fitzgerald, J.M.; Boedigheimer, M.; Davis, B.E.; Dias, C.; et al. Effects of an Anti-TSLP Antibody on Allergen-Induced Asthmatic Responses. N. Engl. J. Med. 2014, 370, 2102–2110. [Google Scholar] [CrossRef]
- Corren, J.; Parnes, J.R.; Wang, L.; Mo, M.; Roseti, S.L.; Griffiths, J.M.; Van Der Merwe, R. Tezepelumab in Adults with Uncontrolled Asthma. N. Engl. J. Med. 2017, 377, 936–946. [Google Scholar] [CrossRef] [PubMed]
- Matera, M.G.; Rogliani, P.; Calzetta, L.; Cazzola, M. TSLP Inhibitors for Asthma: Current Status and Future Prospects. Drugs 2020, 80, 449–458. [Google Scholar] [CrossRef]
- Fornasa, G.; Tsilingiri, K.; Caprioli, F.; Botti, F.; Mapelli, M.; Meller, S.; Kislat, A.; Homey, B.; Di Sabatino, A.; Sonzogni, A.; et al. Dichotomy of short and long thymic stromal lymphopoietin isoforms in inflammatory disorders of the bowel and skin. J. Allergy Clin. Immunol. 2015, 136, 413–422. [Google Scholar] [CrossRef]
- Harada, M.; Hirota, T.; Jodo, A.I.; Doi, S.; Kameda, M.; Fujita, K.; Miyatake, A.; Enomoto, T.; Noguchi, E.; Yoshihara, S.; et al. Functional Analysis of the Thymic Stromal Lymphopoietin Variants in Human Bronchial Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2009, 40, 368–374. [Google Scholar] [CrossRef]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell–mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.; Fukuyama, Y.; Park, E.J.; Yuki, Y.; Kurashima, Y.; Ouchida, R.; Ziegler, S.F.; Kiyono, H. Critical role of TSLP-responsive mucosal dendritic cells in the induction of nasal antigen-specific IgA response. Mucosal Immunol. 2016, 10, 901–911. [Google Scholar] [CrossRef]
- He, B.; Xu, W.; Santini, P.A.; Polydorides, A.D.; Chiu, A.; Estrella, J.; Shan, M.; Chadburn, A.; Villanacci, V.; Plebani, A.; et al. Intestinal Bacteria Trigger T Cell-Independent Immunoglobulin A2 Class Switching by Inducing Epithelial-Cell Secretion of the Cytokine APRIL. Immunity 2007, 26, 812–826. [Google Scholar] [CrossRef]
- den Hartog, G.; van Osch, T.L.; Vos, M.; Meijer, B.; Savelkoul, H.F.; van Neerven, R.J.; Brugman, S. BAFF augments IgA2 and IL-10 production by TLR7/8 stimulated total peripheral blood B cells. Eur. J. Immunol. 2017, 48, 283–292. [Google Scholar] [CrossRef]
- Ladjemi, M.Z.; Lecocq, M.; Weynand, B.; Bowen, H.; Gould, H.J.; Van Snick, J.; Detry, B.; Pilette, C. Increased IgA production by B-cells in COPD via lung epithelial interleukin-6 and TACI pathways. Eur. Respir. J. 2015, 45, 980–993. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, Y.; Liu, Q.; Zhong, W.; Xia, Z. All-trans retinoic acid attenuates airway inflammation by inhibiting Th2 and Th17 response in experimental allergic asthma. BMC Immunol. 2013, 14, 28. [Google Scholar] [CrossRef]
- Mora, R.J.; von Andrian, U.H. Role of retinoic acid in the imprinting of gut-homing IgA-secreting cells. In Seminars in Immunology; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Meng, H.; Li, H.; Ohe, R.; Naing, Y.A.; Yang, S.; Kabasawa, T.; Kato, T.; Osakabe, M.; Ohtake, H.; Ishida, A.; et al. Thymic stromal lymphopoietin in tonsillar follicular dendritic cells correlates with elevated serum immunoglobulin A titer by promoting tonsillar immunoglobulin A class switching in immunoglobulin A nephropathy. Transl. Res. 2016, 176, 1–17. [Google Scholar] [CrossRef]
- Ladjemi, M.Z.; Gras, D.; Dupasquier, S.; Detry, B.; Lecocq, M.; Garulli, C.; Fregimilicka, C.; Bouzin, C.; Gohy, S.; Chanez, P.; et al. Bronchial Epithelial IgA Secretion Is Impaired in Asthma. Role of IL-4/IL-13. Am. J. Respir. Crit. Care Med. 2018, 197, 1396–1409. [Google Scholar] [CrossRef]
- Dzidic, M.; Abrahamsson, T.R.; Artacho, A.; Björkstén, B.; Collado, M.C.; Mira, A.; Jenmalm, M.C. Aberrant IgA responses to the gut microbiota during infancy precede asthma and allergy development. J. Allergy Clin. Immunol. 2017, 139, 1017–1025. [Google Scholar] [CrossRef]
- Rao, J.; Qian, X.; Wang, P.; Pu, L.; Zhai, Y.; Wang, X.; Zhang, F.; Lu, L. All-trans retinoic acid preconditioning protects against liver ischemia/reperfusion injury by inhibiting the nuclear factor kappa B signaling pathway. J. Surg. Res. 2013, 180, e99–e106. [Google Scholar] [CrossRef]
- Arora, P.; Kumar, V.; Batra, S. Vitamin A status in children with asthma. Pediatr. Allergy Immunol. 2002, 13, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, Y.; Furusho, T.; Yoshida, A.; Nakamura, H.; Matsuura, T.; Eto, Y. Serum vitamin A concentrations in asthmatic children in Japan. Pediatr. Int. 2006, 48, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Fahey, L.M.; Guan, S.; Liacouras, C.A.; Spergel, J.; Sleiman, P.M.; Hakonarson, H.; Cianferoni, A. Food Allergen Triggers are Increased in Children with the TSLP Risk Allele and Eosinophilic Esophagitis. Clin. Transl. Gastroenterol. 2017, 152, S854–S855. [Google Scholar] [CrossRef][Green Version]
- Mena, A.M.; Langlois, A.; Speca, S.; Schneider, L.; Desreumaux, P.; Dubuquoy, L.; Bertin, B. The Expression of the Short Isoform of Thymic Stromal Lymphopoietin in the Colon Is Regulated by the Nuclear Receptor Peroxisome Proliferator Activated Receptor-Gamma and Is Impaired during Ulcerative Colitis. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Gao, P.; Leung, D.; Rafaels, N.; Hand, T.; Boguniewicz, M.; Hata, T.; Schneider, L.; Hanifin, J.; Gallo, R.; Gao, L. Genetic Variants in TSLP and its Receptor, IL7R, Contribute to an Increased Risk for Atopic Dermatitis and Eczema Herpeticum in Two American Populations. J. Allergy Clin. Immunol. 2009, 123, S70. [Google Scholar] [CrossRef]
- Miyake, Y.; Hitsumoto, S.; Tanaka, K.; Arakawa, M. Association between TSLP Polymorphisms and Eczema in Japanese Women: The Kyushu Okinawa Maternal and Child Health Study. Inflammation 2015, 38, 1663–1668. [Google Scholar] [CrossRef]
- Sun, Y.; O’Connor, B.; Ratoff, J.; Meng, Q.; Fang, C.; Cousins, D.; Zhang, G.; Gu, S.; Gao, Z.; Shamji, B.; et al. Expression and Cellular Provenance of Thymic Stromal Lymphopoietin and Chemokines in Patients with Severe Asthma and Chronic Obstructive Pulmonary Disease. J. Immunol. 2008, 181, 2790–2798. [Google Scholar] [CrossRef]
- Wang, W.; Li, Y.; Lv, Z.; Chen, Y.; Li, Y.; Huang, K.; Corrigan, C.J.; Ying, S. Bronchial Allergen Challenge of Patients with Atopic Asthma Triggers an Alarmin (IL-33, TSLP, and IL-25) Response in the Airways Epithelium and Submucosa. J. Immunol. 2018, 201, 2221–2231. [Google Scholar] [CrossRef] [PubMed]
- Brandt, E.B.; Bolcas, P.E.; Ruff, B.P.; Hershey, G.K.K. TSLP contributes to allergic airway inflammation induced by diesel exhaust particle exposure in an experimental model of severe asthma. Clin. Exp. Allergy 2020, 50, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Segawa, R.; Hirasawa, N. Exacerbation of Allergic Diseases by Chemicals: Role of TSLP. J. Pharmacol. Sci. 2014, 124, 301–306. [Google Scholar] [CrossRef]
- Fahey, L.M.; Chandramouleeswaran, P.M.; Guan, S.; Benitez, A.J.; Furuta, G.T.; Aceves, S.S.; Wang, M.-L.; Liacouras, C.A.; Muir, A.B.; Sleiman, P.M.; et al. Food allergen triggers are increased in children with the TSLP risk allele and eosinophilic esophagitis. Clin. Transl. Gastroenterol. 2018, 9, e139. [Google Scholar] [CrossRef]
- Malinczak, C.-A.; Fonseca, W.; Rasky, A.J.; Ptaschinski, C.; Morris, S.; Ziegler, S.F.; Lukacs, N.W. Sex-associated TSLP-induced immune alterations following early-life RSV infection leads to enhanced allergic disease. Mucosal Immunol. 2019, 12, 969–979. [Google Scholar] [CrossRef]
- Hunninghake, G.M.; Soto-Quirós, M.E.; Avila, L.; Kim, H.P.; Lasky-Su, J.; Rafaels, N.; Ruczinski, I.; Beaty, T.H.; Mathias, R.A.; Barnes, K.C.; et al. TSLP polymorphisms are associated with asthma in a sex-specific fashion. Allergy 2010, 65, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Hirota, T.; Jodo, A.I.; Hitomi, Y.; Sakashita, M.; Tsunoda, T.; Miyagawa, T.; Doi, S.; Kameda, M.; Fujita, K.; et al. Thymic Stromal Lymphopoietin Gene Promoter Polymorphisms Are Associated with Susceptibility to Bronchial Asthma. Am. J. Respir. Cell Mol. Biol. 2011, 44, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, W.; Lv, Z.; Li, Y.; Chen, Y.; Huang, K.; Corrigan, C.J.; Ying, S. Elevated Expression of IL-33 and TSLP in the Airways of Human Asthmatics In Vivo: A Potential Biomarker of Severe Refractory Disease. J. Immunol. 2018, 7, 2253–2262. [Google Scholar] [CrossRef]
- Demehri, S.; Yockey, L.J.; Visness, C.M.; Jaffee, K.F.; Turkoz, A.; Wood, R.A.; O’Connor, G.T.; Kattan, M.; Gern, J.E.; Gergen, P.J.; et al. Circulating TSLP associates with decreased wheezing in non-atopic preschool children: Data from the URECA birth cohort. Clin. Exp. Allergy 2014, 44, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Westerhuis, B.; Ten Hulscher, H.; Jacobi, R.; van Beek, J.; Koopmans, M.; Rimmelzwaan, G.; Meijer, A.; van Binnendijk, R. Specific memory B cell response in humans upon infection with highly pathogenic H7N7 avian influenza virus. Sci. Rep. 2020, 10, 3152. [Google Scholar] [CrossRef]
- Yu, X.; Lakerveld, A.J.; Imholz, S.; Hendriks, M.; Brink, S.C.A.T.; Mulder, H.L.; De Haan, K.; Schepp, R.M.; Luytjes, W.; De Jong, M.D.; et al. Antibody and Local Cytokine Response to Respiratory Syncytial Virus Infection in Community-Dwelling Older Adults. mSphere 2020, 5, 00577-20. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stimulus | Added | |||
|---|---|---|---|---|
| Concentration | Supplier | Day 0 | Day 5 | |
| T Cell Dependent Cell Culture | ||||
| CD40L | 500 irradiated CD40L cells per well | RIVM | X | |
| CpG (ODN 2006) | 3 µg/mL | Invivogen | X | |
| IL-2 | 10 ng/mL | R&D Systems | X | X |
| IL-21 | 100 ng/mL | Myltenyi Biotec | X | |
| IL-10 | 10 ng/mL | BD Biosciences | X | X |
| T Cell Independent Cell Culture | ||||
| APRIL | 200 ng/mL | Peprotech | X | X |
| BAFF | 200 ng/mL | Peprotech | X | X |
| IL-10 | 10 ng/mL | BD Biosciences | X | X |
| Stimuli Investigated for Effect on IgA Production | ||||
| Short TSLP | 10 ng/mL | Maria Rescignio/Giulia Fornasa, HUNIMED, Italy * | X | X |
| Long TSLP | 10 ng/mL | BioLegend | X | X |
| IL-13 | 10 ng/mL | Peprotech | X | X |
| IL-4 | 10 ng/mL | Peprotech | X | X |
| RA | 100 µM | Sigma Aldrich | X | |
| Target Protein | Fluorochrome | Clone | Manufacturer |
|---|---|---|---|
| FACS Cell Analyses | |||
| CD19 | BV786 | SJ25C1 | BD Biosciences, San Jose, CA, USA |
| CD20 | BV510 | 2H7 | BioLegend, San Diego, CA, USA |
| CD27 | BV711 | M-T271 | BioLegend |
| CD38 | APC-H7 | HB7 | BD Biosciences |
| IgA | FITC or APC | IS11-8E10 | Myltenyi Biotec, Bergisch-Gladbach, Germany |
| IgG | BV421 | M1310G05 | BioLegend |
| FACS Cell Sorting | |||
| CD3 | PerCP-Cy5.5 | UCHT1 | BioLegend |
| CD19 | PE-Cy7 | HIB19 | BioLegend |
| IgD | FITC or AF700 | IA6-2 | BioLegend |
| CD27 | BV711 | M-T271 | BioLegend |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Heerden, D.; van Binnendijk, R.S.; Tromp, S.A.M.; Savelkoul, H.F.J.; van Neerven, R.J.J.; den Hartog, G. Asthma-Associated Long TSLP Inhibits the Production of IgA. Int. J. Mol. Sci. 2021, 22, 3592. https://doi.org/10.3390/ijms22073592
van Heerden D, van Binnendijk RS, Tromp SAM, Savelkoul HFJ, van Neerven RJJ, den Hartog G. Asthma-Associated Long TSLP Inhibits the Production of IgA. International Journal of Molecular Sciences. 2021; 22(7):3592. https://doi.org/10.3390/ijms22073592
Chicago/Turabian Stylevan Heerden, Dorianne, Robert S. van Binnendijk, Samantha A. M. Tromp, Huub F. J. Savelkoul, R. J. Joost van Neerven, and Gerco den Hartog. 2021. "Asthma-Associated Long TSLP Inhibits the Production of IgA" International Journal of Molecular Sciences 22, no. 7: 3592. https://doi.org/10.3390/ijms22073592
APA Stylevan Heerden, D., van Binnendijk, R. S., Tromp, S. A. M., Savelkoul, H. F. J., van Neerven, R. J. J., & den Hartog, G. (2021). Asthma-Associated Long TSLP Inhibits the Production of IgA. International Journal of Molecular Sciences, 22(7), 3592. https://doi.org/10.3390/ijms22073592