Expression of AhATL1, an ABA Transport Factor Gene from Peanut, Is Affected by Altered Memory Gene Expression Patterns and Increased Tolerance to Drought Stress in Arabidopsis

Abstract
1. Introduction
2. Results
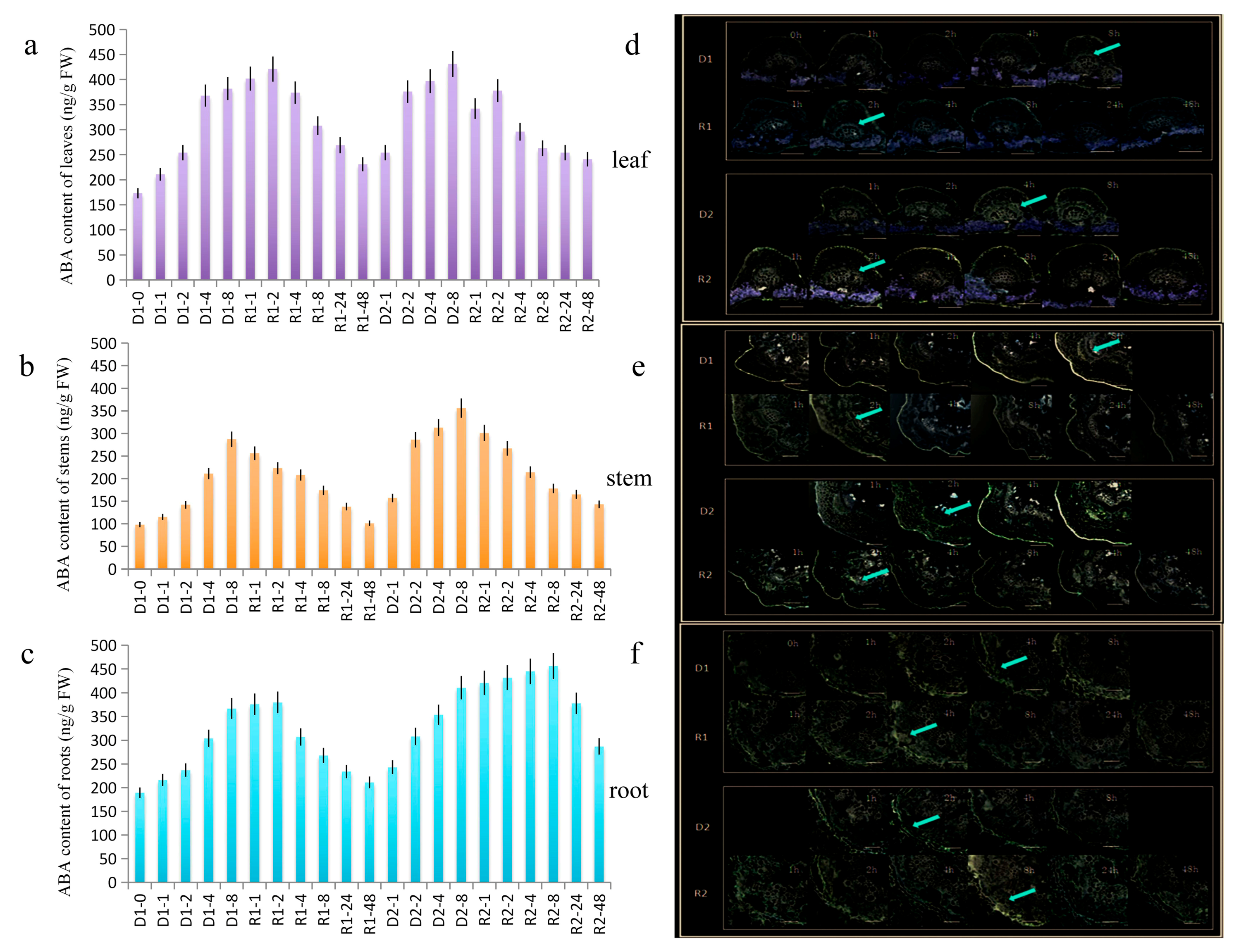
2.1. High Level of AhATL1 Expression and AhATL1 Distribution Resulted in Rapid Increase in ABA Content in Peanut during Drought Stress Memory
2.2. Heterologous Overexpression of AhATL1 Alters the Expression Patterns of Some Memory Genes in Arabidopsis Thaliana
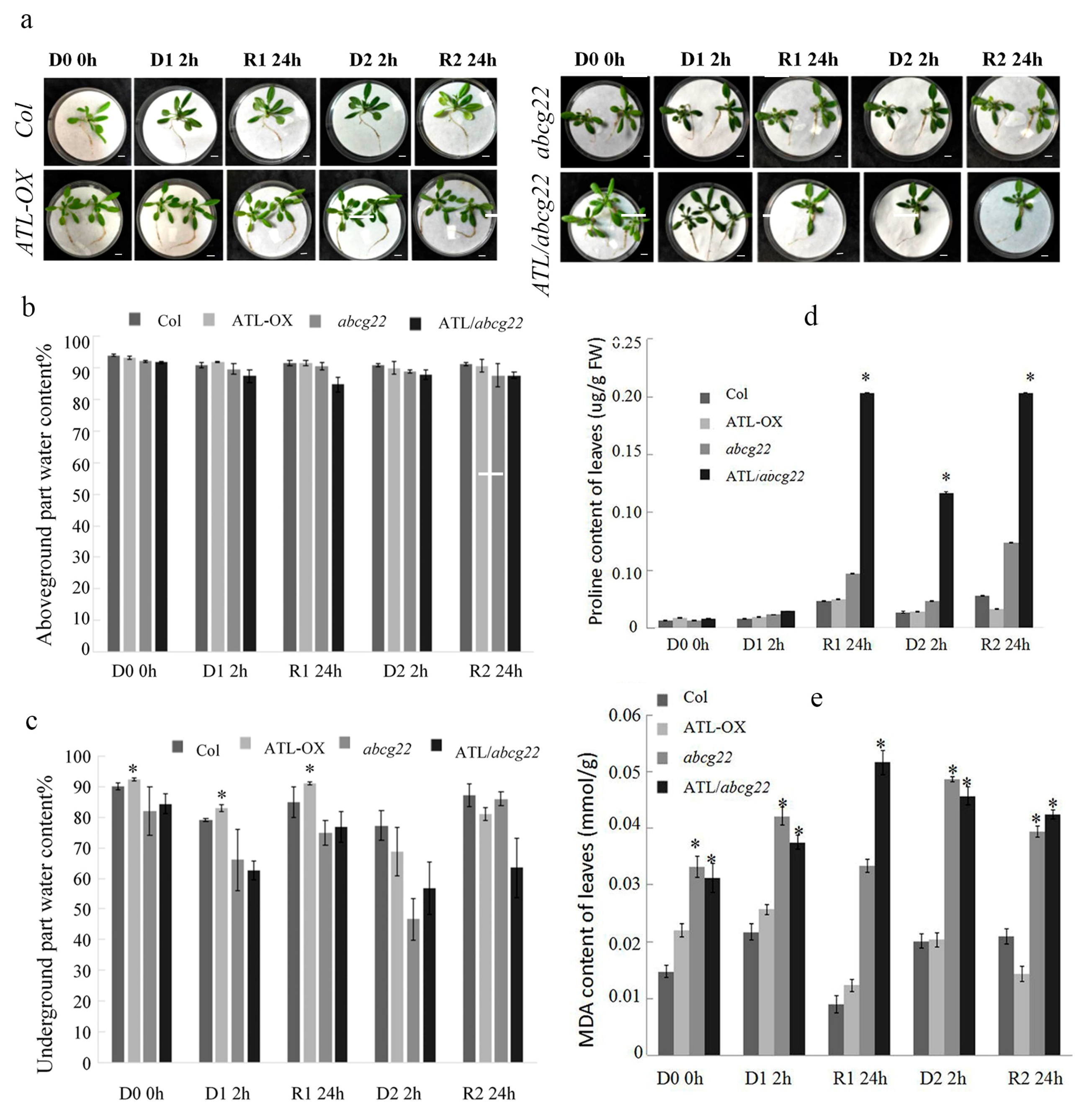
2.3. Overexpression of AhATL1 in Arabidopsis Enhances Tolerance Ability in the Drought Stress Memory Process
3. Discussion
4. Material and Methods
4.1. Plant Material and Growth Conditions
4.2. Measurement of Endogenous ABA Content
4.3. Immunofluorescence Localization Assays
4.4. AhATL1 Overexpressing Arabidopsis Plants
4.5. Measurement of Water Content
4.6. Measurement of Proline Content
4.7. Measurement of Malondialdehyde (MDA) Content
4.8. Expression Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Conrath, U.; Beckers, G.J.M.; Langenbach, C.J.G.; Jaskiewicz, A.M.R. Priming for Enhanced Defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Kumar, A. Plant behavior: An evolutionary response to the environment? Plant Biol. 2020, 22, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Stief, A.; Altmann, S.; Hoffmann, K.; Pant, B.D.; Scheible, W.-R.; Bäurle, I. Arabidopsis miR156 Regulates Tolerance to Recurring Environmental Stress through SPL Transcription Factors. Plant Cell 2014, 26, 1792–1807. [Google Scholar] [CrossRef]
- Hilker, M.; Schmülling, T. Stress priming, memory, and signalling in plants. Plant Cell Environ. 2019, 42, 753–761. [Google Scholar] [CrossRef]
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef]
- Ebe, M.; Pirko, J.; Hannes, K. The Role of ABA Recycling and Transporter Proteins in Rapid Stomatal Responses to Reduced Air Humidity, Elevated CO2 and Exogenous ABA. Mol. Plant 2016, 8, 657–659. [Google Scholar]
- Kang, J.; Yim, S.; Choi, H.; Kim, A.; Lee, K.P.; Lopez-Molina, L.; Martinoia, E.; Lee, Y. Abscisic acid transporters cooperate to control seed germination. Nat. Commun. 2015, 6, 8113. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Hwang, J.-U.; Lee, M.; Kim, Y.-Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef]
- Kuromori, T.; Miyaji, T.; Yabuuchi, H.; Shimizu, H.; Sugimoto, E.; Kamiya, A.; Moriyama, Y.; Shinozaki, K. ABC transporter AtABCG25 is involved in abscisic acid transport and responses. Proc. Natl. Acad. Sci. USA 2010, 107, 2361–2366. [Google Scholar] [CrossRef] [PubMed]
- Ge, K.; Liu, X.; Li, X.; Hu, B.; Li, L. Isolation of an ABA Transporter-Like 1 Gene from Arachis hypogaea That Affects ABA Import and Reduces ABA Sensitivity in Arabidopsis. Front. Plant Sci. 2017, 8, 1150. [Google Scholar] [CrossRef]
- Chen, R.Q.; Shu, W.; Ge, K.; Hu, B.; Li, L. Effect on growth and expressions of stress-related genes in peanut under drought stress training. Plant Physiol. J. 2017, 10, 1921–1927. [Google Scholar]
- Nambara, E.; Marion, P.A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R. Abscisic Acid Synthesis and Response. Arab. Book 2013, 11, e0166. [Google Scholar] [CrossRef] [PubMed]
- Boursiac, Y.; Léran, S.; Corratgé-Faillie, C.; Gojon, A.; Krouk, G.; Lacombe, B. ABA transport and transporters. Trends Plant Sci. 2013, 18, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Puértolas, J.; Conesa, M.R.; Ballester, C.; Dodd, I.C. Local root abscisic acid (ABA) accumulation depends on the spatial distribution of soil moisture in potato: Implications for ABA signalling under heterogeneous soil drying. J. Exp. Bot. 2015, 66, 2325–2334. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Cao, J.; Ge, K.; Li, L. The site of water stress governs the pattern of ABA synthesis and transport in peanut. Sci. Rep. 2016, 6, 32143. [Google Scholar] [CrossRef]
- Colaneri, A.C.; Jones, A.M. Genome-wide quantitative identifi cation of DNA differentially methylated sites in Arabidopsis seedlings growing at different water potential. PLoS ONE 2013, 8, e59878. [Google Scholar] [CrossRef]
- Liao, W.-B.; Huang, G.-B.; Yu, J.-H.; Zhang, M.-L. Nitric oxide and hydrogen peroxide alleviate drought stress in marigold explants and promote its adventitious root development. Plant Physiol. Biochem. 2012, 58, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Virlouvet, L.; Liu, N.; Riethoven, J.-J.; Fromm, M.; Avramova, Z. Dehydration stress memory genes of Zea mays; comparison with Arabidopsis thaliana. BMC Plant Biol. 2014, 14, 141. [Google Scholar] [CrossRef] [PubMed]
- Kuromori, T.; Sugimoto, E.; Shinozaki, K. Arabidopsis mutants of AtABCG22, an ABC transporter gene, increase water transpiration and drought susceptibility. Plant J. 2011, 67, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Fromm, M.E.; Avramova, Z. Multiple exposures to drought ’train’ transcriptional responses in Arabidopsis. Nat. Commun. 2012, 3, 740. [Google Scholar] [CrossRef] [PubMed]
- Forlani, G.; Bertazzini, M.; Cagnano, G. Stress-driven increase in proline levels, and not proline levels themselves, correlates with the ability to withstand excess salt in a group of 17 Italian rice genotypes. Plant Biol. 2019, 21, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Pál, M.; Tajti, J.; Szalai, G.; Peeva, V.; Végh, B.; Janda, T. Interaction of polyamines, abscisic acid and proline under osmotic stress in the leaves of wheat plants. Sci. Rep. 2018, 8, 12839. [Google Scholar] [CrossRef] [PubMed]
- Slatyer, R.O.; Markus, D.K. Plant-Water Relationships. Soil Sci. 1967, 106, 478. [Google Scholar] [CrossRef]
- Byun, Y.-J.; Koo, M.-Y.; Joo, H.-J.; Ha-Lee, Y.-M.; Lee, D.-H. Comparative analysis of gene expression under cold acclimation, deacclimation and reacclimation in Arabidopsis. Physiol. Plant. 2014, 152, 256–274. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Munné-Bosch, S. Stress memory and the inevitable effects of drought: A physiological perspective. Front. Plant Sci. 2016, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Nicotra, A.B.; Davidson, A. Adaptive phenotypic plasticity and plant water use. Funct. Plant Biol. 2010, 37, 117–127. [Google Scholar] [CrossRef]
- Cavanagh, A.P.; Kubien, D.S. Can phenotypic plasticity in Rubisco performance contribute to photosynthetic acclimation? Photosynth. Res. 2014, 119, 203–214. [Google Scholar] [CrossRef]
- Cao, J.J.; Lü, Y.; Hu, B.; Li, L. Changers of AhCYP707A1 proteins and ABA distribution in peanut leaves under water stress. Plant Physiol. J. 2016, 52, 436–444. [Google Scholar]
- Hu, B.; Xiao, S.; Lü, Y.; Li, L. Distribution of ABA and AhNCED 1 in peanut leaves of different drought resistant cultivars subjected to drought stress. Chin. J. Cell Biol. 2012, 34, 992–997. [Google Scholar]
- Baldwin, I.T.; Schmelz, E.A. Immunological “Memory” in the Induced Accumulation of Nicotine in Wild Tobacco. Ecology 1996, 77, 236–246. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, N.; Virlouvet, L.; Riethoven, J.-J.; Fromm, M.; Avramova, Z. Four distinct types of dehydration stress memory genes in Arabidopsis thaliana. BMC Plant Biol. 2013, 13, 229. [Google Scholar] [CrossRef]
- Virlouvet, L.; Fromm, M. Physiological and transcriptional memory in guard cells during repetitive dehydration stress. New Phytol. 2015, 205, 596–607. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; Nagy, L.; Hein, R.; Rascher, U.; Beierkuhnlein, C.; Willner, E.; Jentsch, A. Do plants remember drought? Hints towards a drought-memory in grasses. Environ. Exp. Bot. 2011, 71, 34–40. [Google Scholar] [CrossRef]
- Avramova, Z. Transcriptional ‘memory’ of a stress: Transient chromatin and memory (epigenetic) marks at stress-response genes. Plant J. 2015, 83, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Q.; Feng, H.; Deng, J.; Zhang, R.; Wen, J.; Dong, J.; Wang, T. Dehydrin MtCAS31 promotes autophagic degradation under drought stress. Autophagy 2019, 16, 862–877. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Lv, Y.; Wan, X.R.; Li, L.M.; Hu, B.; Li, L. Cloning and expression analysis of cDNAs encodingABA 8′-hydroxylase in peanut plants in response to osmotic stress. PLoS ONE 2014, 9, e97025. [Google Scholar]
- Kandasamy, M.K.; McKinney, E.C.; Meagher, R.B. Functional Nonequivalency of Actin Isovariants inArabidopsis. Mol. Biol. Cell 2002, 13, 251–261. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Col | abcg22 | AhATL1-OX | ||
|---|---|---|---|---|
| Memory gene | [+/+] D1-0h<D1-2h<D2-2h | AtOST1, AtABI5, AtAREB1, AtBG2 | AtOST1, AtABI5, AtAREB1, AtRD29A,AtNCED3, AtSLAC1,AtALMT12, AtABCG11 | AtOST1, AtABI5, AtAREB1, AtRD29A,AtNCED3, AtSLAC1,AtALMT12, AtABCG11 |
| [−/−] D1-0h>D1-2h>D2-2h | AtUGT71C5 | |||
| [+/−] D1-0h<D1-2h>D2-2h | AtRD29A,AtNCED3, AtPP2C5,AtSLAC1, AtALMT12, AtABCG11, AtABCG22,AtABCG40, AtABCG25 | AtABCG22 | ||
| Post response gene | [−/+] D1-0h>D1-2h<D2-2h | AtUGT71C5 | AtBG2,AtPP2C5,AtABCG40, AtABCG25,AtUGT71C5, AtCYP707A1 | AtBG2,AtPP2C5, AtABCG40,AtABCG25, AtCYP707A1 |
| [=/+] D1-0h=D1-2h<D2-2h | AtCYP707A1 | |||
| [=/−] D1-0h=D1-2h>D2-2h |
| Gene | Primer Sequence (5′→3′) |
|---|---|
| AtActin | F: GCTGTTGACTACGAGCAGGA |
| R: TTCCATTCCCACAAACGAG | |
| AtRD29A | F: CAAAGCAATGAGCATGAGCAAG |
| R: CGGAAGACACGACAGGAAACAC | |
| AtNCED3 | F: AAGGTCGCAAGATTCGGGATT |
| R: CGTTGAAAATTGAGTCTGGTGGAG | |
| AtCYP707A1 | F: TCTTCCAAACTCCCACTCCCT |
| R: GCACGAACTTAGCAGCCTCTG | |
| AtUGT71C5 | F: CCGATGAAATAGCCACAGCC |
| R: CCACCGTAGAAGACCCACCA | |
| AtBG2 | F: AAGAATGGATCACCGAGAAGGC |
| R: TGGATGAAACAGTCCCCAAAACT | |
| AtPP2C5 | F: CGACCACCGATGCTTGACTT |
| R: ACAACTTCCGCTCCTTTCTCC | |
| AtOST1 | F: TGGAGTTGCGAGATTGATGAGA |
| R: AATGGCTAAATGGGTTGGTGTT | |
| AtABI5 | F: CACTTCCAGCTCCGCTTTGT |
| R: GGTTGTCTAGCCGCAGTCTCA | |
| AtAREB1 | F: GTGTCGCCTGTTACGCCATT |
| R: CGGTTCTTTATCATTCTCCTTTGC | |
| AtSLAC1 | F: GCGGGTTTGAATCAGGTGG |
| R: TTTGCTGAGCGTTGATTTAGTCC | |
| AtALMT12 | F: TGAGCAAGACGAAGTGGATGG |
| R: AGCAAAGAAACGAGTGTCAAGGA | |
| AtABCG11 | F: TTTGACAGCCAAGGGAGTGC |
| R: TGATGAAGAAGATGATGCGGTAGA | |
| AtABCG22 | F: CGTGCGTGGTGTATCAGGTG |
| R: ATGGTTCTAAGAGCAGTGGTGGA | |
| AtABCG40 | F: CCCAGGAAATGATAGAGCAAGG |
| R: GGGTAACCGGAAATGGTGATG | |
| AtABCG25 | F: TAAACGCAGTCGCAGGGAGA |
| R: TCCGAGGAAGACGAAGCAAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, M.; Li, X.; Tang, S.; Huang, Y.; Li, L.; Hu, B. Expression of AhATL1, an ABA Transport Factor Gene from Peanut, Is Affected by Altered Memory Gene Expression Patterns and Increased Tolerance to Drought Stress in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 3398. https://doi.org/10.3390/ijms22073398
Qin M, Li X, Tang S, Huang Y, Li L, Hu B. Expression of AhATL1, an ABA Transport Factor Gene from Peanut, Is Affected by Altered Memory Gene Expression Patterns and Increased Tolerance to Drought Stress in Arabidopsis. International Journal of Molecular Sciences. 2021; 22(7):3398. https://doi.org/10.3390/ijms22073398
Chicago/Turabian StyleQin, Ming, Xiaoyan Li, Shaohua Tang, Yinglin Huang, Ling Li, and Bo Hu. 2021. "Expression of AhATL1, an ABA Transport Factor Gene from Peanut, Is Affected by Altered Memory Gene Expression Patterns and Increased Tolerance to Drought Stress in Arabidopsis" International Journal of Molecular Sciences 22, no. 7: 3398. https://doi.org/10.3390/ijms22073398
APA StyleQin, M., Li, X., Tang, S., Huang, Y., Li, L., & Hu, B. (2021). Expression of AhATL1, an ABA Transport Factor Gene from Peanut, Is Affected by Altered Memory Gene Expression Patterns and Increased Tolerance to Drought Stress in Arabidopsis. International Journal of Molecular Sciences, 22(7), 3398. https://doi.org/10.3390/ijms22073398