
Identification of Anti-Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-CoV-2) Oxysterol Derivatives In Vitro

 ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Results
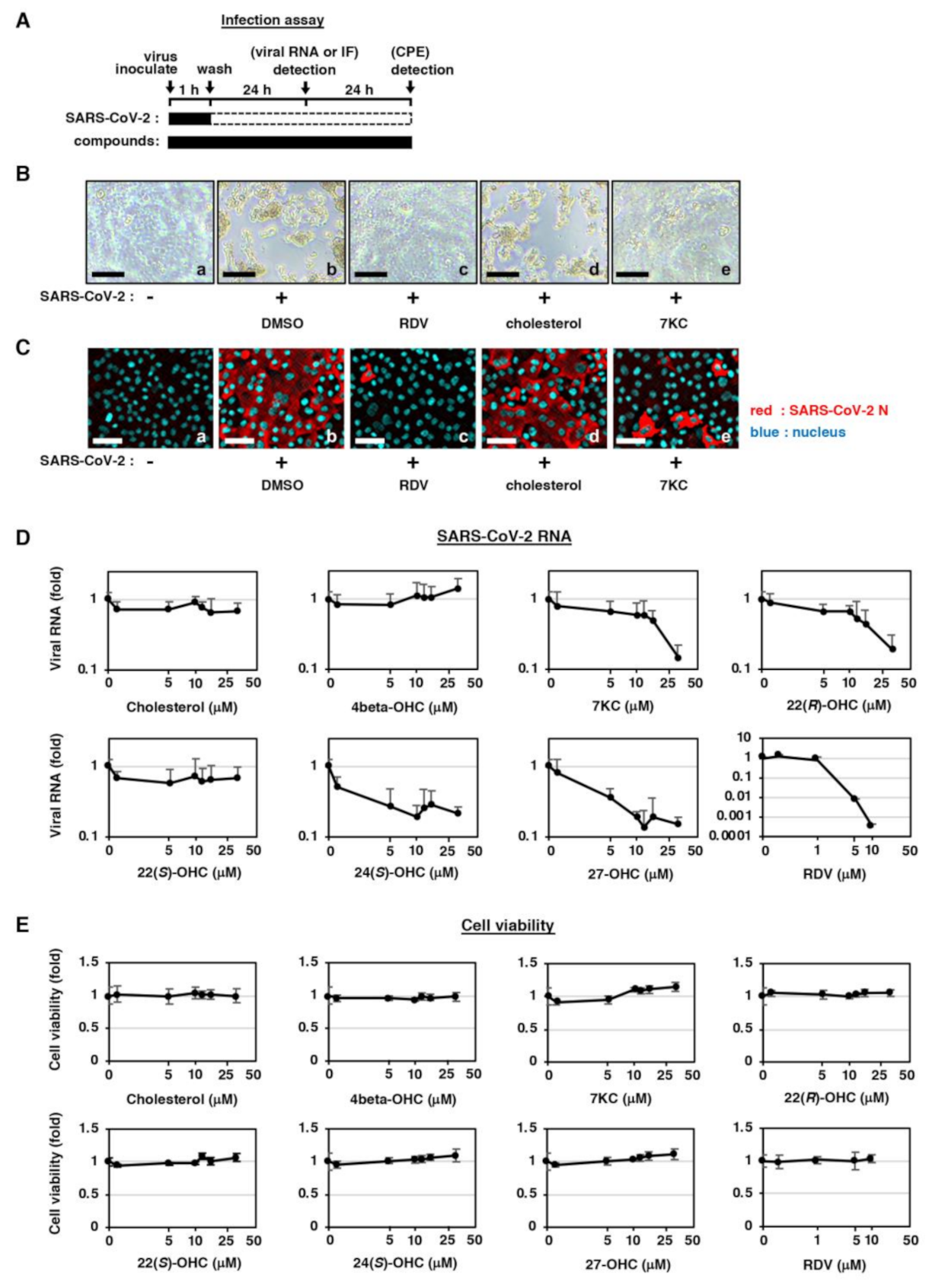
2.1. Natural Oxysterols Have Antiviral Activity against SARS-CoV-2 Infection
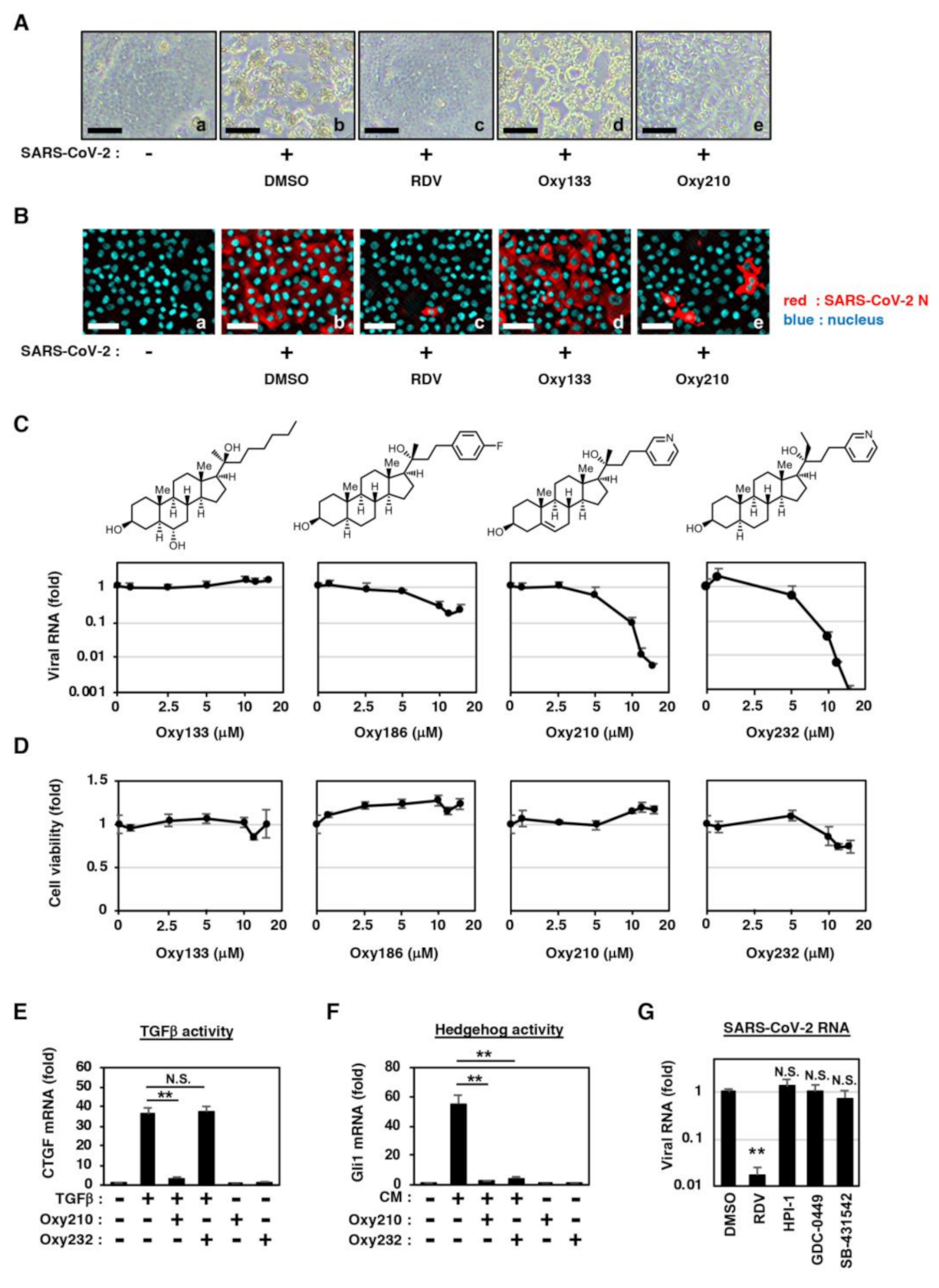
2.2. Semi-Synthetic Oxysterol Derivatives, Oxy210, Oxy186, and Oxy232 Inhibit SARS-CoV-2 Production
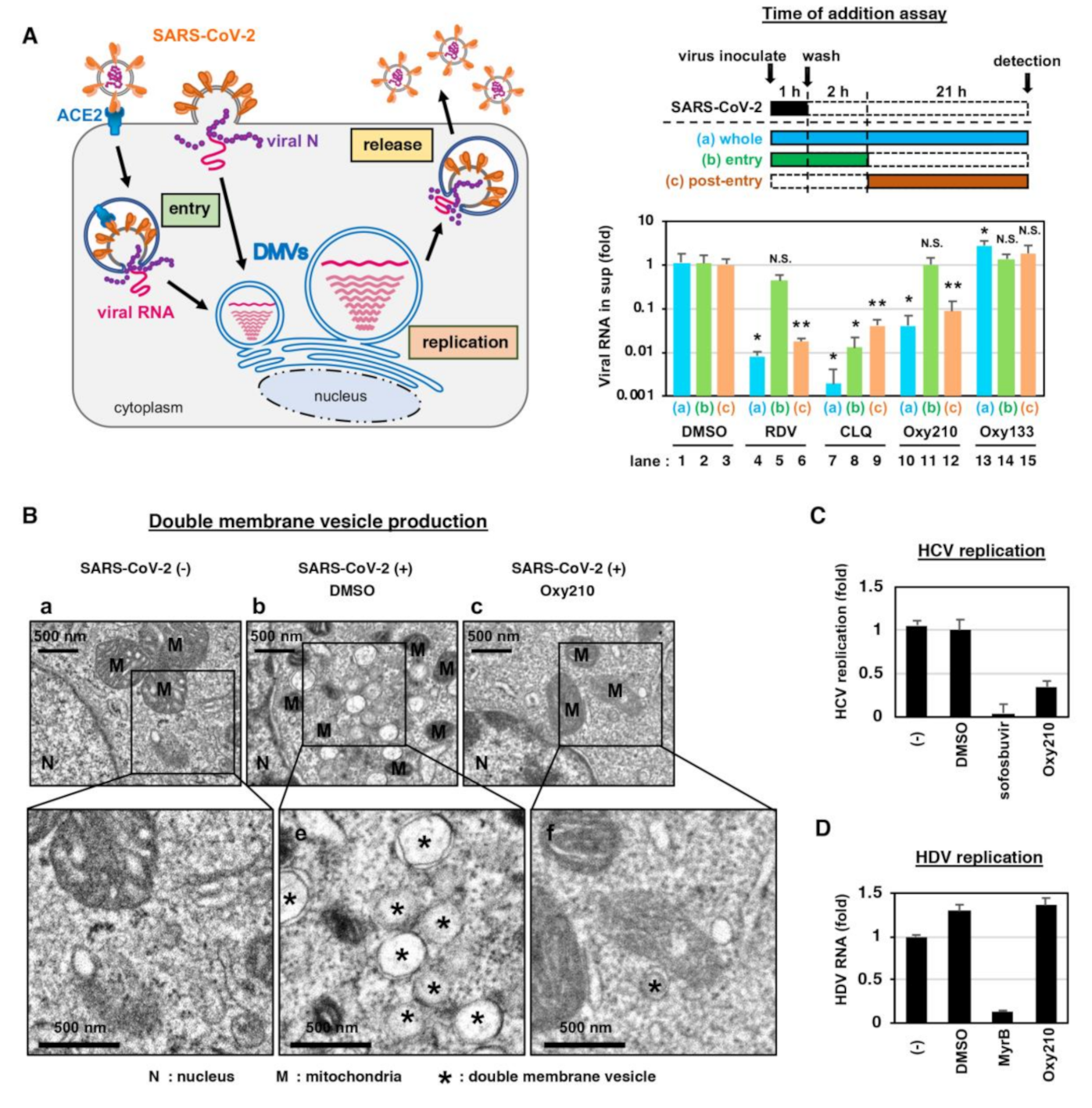
2.3. Oxy210 Inhibits the Intracellular SARS-CoV-2 Replication and Formation of Double Membrane Vesicles
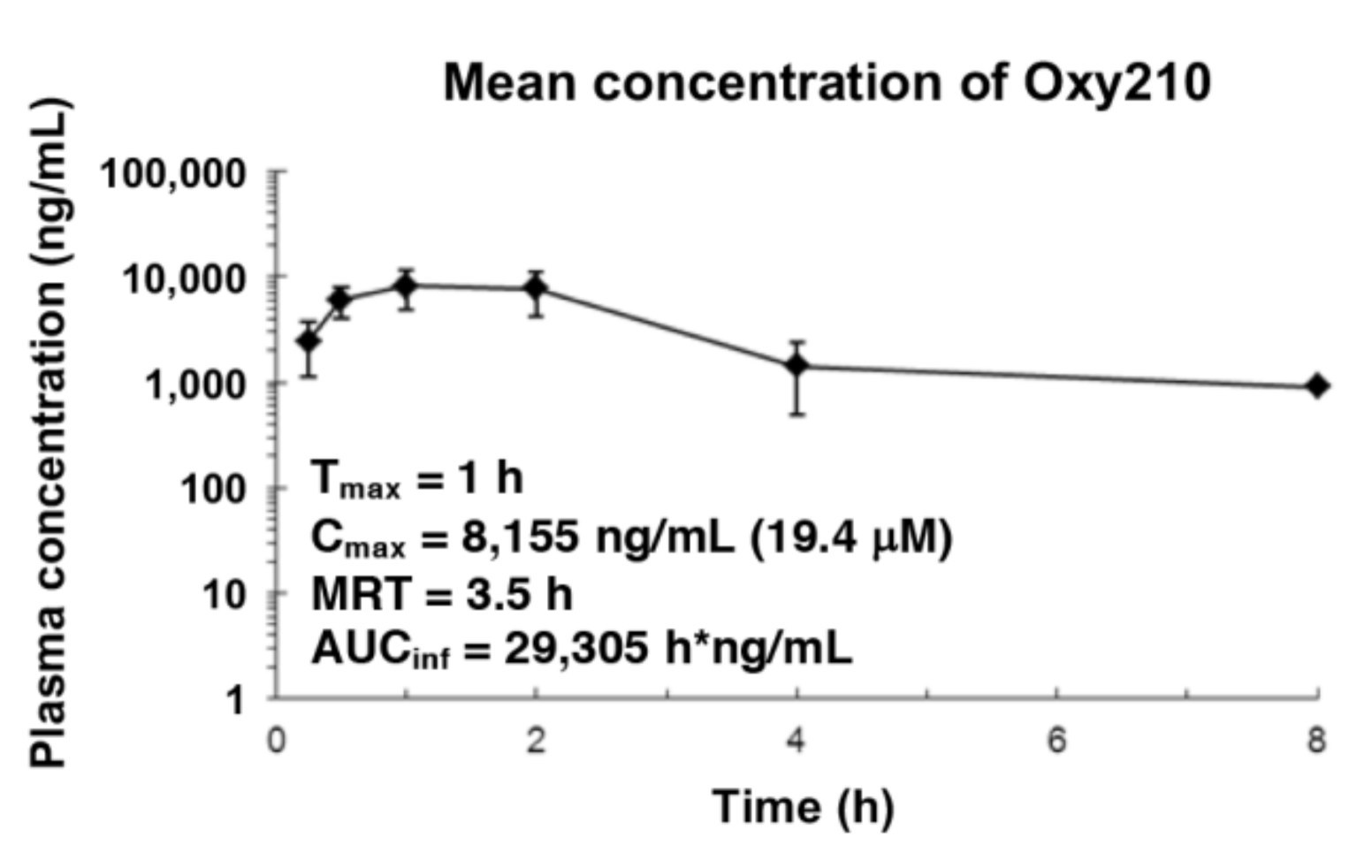
2.4. Pharmacokinetics of Oxy210 in Mice
3. Discussion
4. Materials and Methods
4.1. Compounds and the Synthesis of Oxysterol Derivatives
4.2. Cell Culture
4.3. SARS-CoV-2 Infection Assay
4.4. Quantification of Viral RNA
4.5. Detection of Viral N Protein
4.6. Quantification of Cell Viability
4.7. Quantification of Transforming Growth Factor β (TGFβ) and Hedgehog (Hh) Activity
4.8. Electron Microscopic Analysis
4.9. Hepatitis C Virus (HCV) Replication Assay
4.10. Hepatitis D Virus (HDV) Replication Assay
4.11. Pharmacokinetics of Oxy210 in Mice
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SARS-CoV-2 | severe acute respiratory syndrome-related coronavirus 2 |
| COVID-19 | coronavirus disease 2019 |
| PK | pharmacokinetics |
| DMVs | double-membrane vesicles |
| RDV | remdesivir |
| CPE | cytopathic effect |
| N protein | nucleocapsid protein |
| 7-KC | 7-ketocholesterol |
| 4beta-OHC | 4beta-hydroxycholesterol |
| 22(S)-OHC | 22(S)-hydroxycholesterol |
| 22(R)-OHC | 22(R)-hydroxycholesterol |
| 24(S)-OHC | 24(S)-hydroxycholesterol |
| 27-OHC | 27-hydroxycholesterol |
| IC50 | 50% maximal inhibitory concentration |
| IC90 | 90% maximal inhibitory concentration |
| CC50 | 50% maximal cytotoxic concentration |
| CLQ | chloroquine |
| HCV | hepatitis C virus |
| HDV | hepatitis D virus |
| CTGF | connective tissue growth factor |
References
- Watashi, K. Identifying and repurposing antiviral drugs against severe acute respiratory syndrome coronavirus 2 with in silico and in vitro approaches. Biochem. Biophys. Res. Commun. 2020, 538, 137–144. [Google Scholar] [CrossRef]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Edwards, A. What Are the Odds of Finding a COVID-19 Drug from a Lab Repurposing Screen? J. Chem. Inf. Model. 2020, 60, 5727–5729. [Google Scholar] [CrossRef]
- Griffiths, W.J.; Wang, Y. Oxysterol research: A brief review. Biochem. Soc. Trans. 2019, 47, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, F.; Crinelli, R.; Cesarini, E.; Canonico, B.; Guidi, L.; Zerbinati, C.; Di Sario, G.; Zamai, L.; Magnani, M.; Papa, S.; et al. Endothelial cells, endoplasmic reticulum stress and oxysterols. Redox Biol. 2017, 13, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Watashi, K.; Tsukuda, S.; Aly, H.H.; Fukasawa, M.; Fujimoto, A.; Suzuki, R.; Aizaki, H.; Ito, T.; Koiwai, O.; et al. Evaluation and identification of hepatitis B virus entry inhibitors using HepG2 cells overexpressing a membrane transporter NTCP. Biochem. Biophys. Res. Commun. 2014, 443, 808–813. [Google Scholar] [CrossRef]
- Cagno, V.; Civra, A.; Rossin, D.; Calfapietra, S.; Caccia, C.; Leoni, V.; Dorma, N.; Biasi, F.; Poli, G.; Lembo, D. Inhibition of herpes simplex-1 virus replication by 25-hydroxycholesterol and 27-hydroxycholesterol. Redox Biol. 2017, 12, 522–527. [Google Scholar] [CrossRef]
- Civra, A.; Cagno, V.; Donalisio, M.; Biasi, F.; Leonarduzzi, G.; Poli, G.; Lembo, D. Inhibition of pathogenic non-enveloped viruses by 25-hydroxycholesterol and 27-hydroxycholesterol. Sci. Rep. 2014, 4, 7487. [Google Scholar] [CrossRef] [PubMed]
- Shawli, G.T.; Adeyemi, O.O.; Stonehouse, N.J.; Herod, M.R. The Oxysterol 25-Hydroxycholesterol Inhibits Replication of Murine Norovirus. Viruses 2019, 11, 97. [Google Scholar] [CrossRef] [PubMed]
- Civra, A.; Francese, R.; Gamba, P.; Testa, G.; Cagno, V.; Poli, G.; Lembo, D. 25-Hydroxycholesterol and 27-hydroxycholesterol inhibit human rotavirus infection by sequestering viral particles into late endosomes. Redox Biol. 2018, 19, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Deng, Y.-Q.; Wang, S.; Ma, F.; Aliyari, R.; Huang, X.-Y.; Zhang, N.-N.; Watanabe, M.; Dong, H.-L.; Liu, P.; et al. 25-Hydroxycholesterol Protects Host against Zika Virus Infection and Its Associated Microcephaly in a Mouse Model. Immunity 2017, 46, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, S.R.; Nargizyan, T.; Meliton, V.; Nachtergaele, S.; Rohatgi, R.; Stappenbeck, F.; Jung, M.E.; Johnson, J.S.; Aghdasi, B.; Tian, H.; et al. A Novel Osteogenic Oxysterol Compound for Therapeutic Development to Promote Bone Growth: Activation of Hedgehog Signaling and Osteogenesis Through Smoothened Binding. J. Bone Miner. Res. 2014, 29, 1872–1885. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.P.; Phan, K.H.; Tian, H.; Suzuki, A.; Montgomery, S.R.; Johnson, J.S.; Atti, E.; Tetratis, S.; Pereira, R.C.; Wang, J.C.; et al. Comparison of a novel oxysterol molecule and rhBMP2 fusion rates in a rabbit posterolateral lumbar spine model. Spine J. 2015, 15, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Buser, Z.; Drapeau, S.; Stappenbeck, F.; Pereira, R.C.; Parhami, F.; Wang, J.C. Effect of Oxy133, an osteogenic oxysterol, on new bone formation in rat two-level posterolateral fusion model. Eur. Spine J. 2017, 26, 2763–2772. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Stappenbeck, F.; Parhami, F. Inhibition of Hedgehog Signaling in Fibroblasts, Pancreatic, and Lung Tumor Cells by Oxy186, an Oxysterol Analogue with Drug-Like Properties. Cells 2019, 8, 509. [Google Scholar] [CrossRef]
- Stappenbeck, F.; Wang, F.; Tang, L.-Y.; Zhang, Y.E.; Parhami, F. Inhibition of Non-Small Cell Lung Cancer Cells by Oxy210, an Oxysterol-Derivative that Antagonizes TGFβ and Hedgehog Signaling. Cells 2019, 8, 1297. [Google Scholar] [CrossRef]
- Du Toit, A. Coronavirus replication factories. Nat. Rev. Microbiol. Genet. 2020, 18, 411. [Google Scholar] [CrossRef] [PubMed]
- Wolff, G.; Limpens, R.W.A.L.; Zevenhoven-Dobbe, J.C.; Laugks, U.; Zheng, S.; De Jong, A.W.M.; Koning, R.I.; Agard, D.A.; Grünewald, K.; Koster, A.J.; et al. A molecular pore spans the double membrane of the coronavirus replication organelle. Science 2020, 369, 1395–1398. [Google Scholar] [CrossRef]
- Blanchard, E.; Roingeard, P. Virus-induced double-membrane vesicles. Cell. Microbiol. 2015, 17, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Nao, N.; Shirato, K.; Kawase, M.; Saito, S.; Takayama, I.; Nagata, N.; Sekizuka, V.O.R.C.I.D.P.; Katoh, V.O.R.C.I.D.P.; Kato, F. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. USA 2020, 117, 7001–7003. [Google Scholar] [CrossRef] [PubMed]
- Marcello, A.; Marcelloa, A.; Civra, A.; Bonottoa, R.M.; Alves, L.N.; Rajasekharan, S.; Giacobone, C.; Caccia, C.; Cavalli, R.; Adami, M.; et al. The cholesterol metabolite 27-hydroxycholesterol inhibits SARS-CoV-2 and is markedly decreased in COVID-19 patients. Redox. Biol. 2020, 36, 101682. [Google Scholar] [CrossRef]
- Arca, M.; Natoli, S.; Micheletta, F.; Riggi, S.; Di Angelantonio, E.; Montali, A.; Antonini, T.M.; Antonini, R.; Diczfalusy, U.; Iuliano, L. Increased plasma levels of oxysterols, in vivo markers of oxidative stress, in patients with familial combined hyperlipidemia: Reduction during atorvastatin and fenofibrate therapy. Free. Radic. Biol. Med. 2007, 42, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Akpovwa, H. Chloroquine could be used for the treatment of filoviral infections and other viral infections that emerge or emerged from viruses requiring an acidic pH for infectivity. Cell Biochem. Funct. 2016, 34, 191–196. [Google Scholar] [CrossRef]
- Liu, J.; Cao, R.; Xu, M.; Wang, X.; Zhang, H.; Hu, H.; Li, Y.; Hu, Z.; Zhong, W.; Wang, M. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov. 2020, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Kokic, G.; Hillen, H.S.; Tegunov, D.; Dienemann, C.; Seitz, F.; Schmitzova, J.; Farnung, L.; Siewert, A.; Höbartner, C.; Cramer, P. Mechanism of SARS-CoV-2 polymerase stalling by remdesivir. Nat. Commun. 2021, 12, 279. [Google Scholar] [CrossRef] [PubMed]
- Paul, D.; Bartenschlager, R. Architecture and biogenesis of plus-strand RNA virus replication factories. World J. Virol. 2013, 2, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Urban, S. New insights into HDV persistence: The role of interferon response and implications for upcoming novel therapies. J. Hepatol. 2021, 74, 686–699. [Google Scholar] [CrossRef] [PubMed]
- Ogando, N.S.; Dalebout, T.J.; Zevenhoven-Dobbe, J.C.; Limpens, R.W.; Van Der Meer, Y.; Caly, L.; Druce, J.; De Vries, J.J.C.; Kikkert, M.; Bárcena, M.; et al. SARS-coronavirus-2 replication in Vero E6 cells: Replication kinetics, rapid adaptation and cytopathology. J. Gen. Virol. 2020, 101, 925–940. [Google Scholar] [CrossRef]
- Nao, N.; Sato, K.; Yamagishi, J.; Tahara, M.; Nakatsu, Y.; Seki, F.; Katoh, H.; Ohnuma, A.; Shirogane, Y.; Hayashi, M.; et al. Consensus and variations in cell line specificity among human metapneumovirus strains. PLoS ONE 2019, 14, e0215822. [Google Scholar] [CrossRef]
- Goto, K.; Watashi, K.; Murata, T.; Hishiki, T.; Hijikata, M.; Shimotohno, K. Evaluation of the anti-hepatitis C virus effects of cyclophilin inhibitors, cyclosporin A, and NIM811. Biochem. Biophys. Res. Commun. 2006, 343, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, T.; Fukushi, S.; Saijo, M.; Kurane, I.; Morikawa, S. Phosphorylation of p38 MAPK and its downstream targets in SARS coronavirus-infected cells. Biochem. Biophys. Res. Commun. 2004, 319, 1228–1234. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, H.; Nishioka, K.; Nakajima, S.; Kim, S.; Suzuki, R.; Aizaki, H.; Fukasawa, M.; Kamisuki, S.; Sugawara, F.; Ohtani, N.; et al. The aryl hydrocarbon receptor–cytochrome P450 1A1 pathway controls lipid accumulation and enhances the permissiveness for hepatitis C virus assembly. J. Biol. Chem. 2018, 293, 19559–19571. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Watashi, K.; Kamisuki, S.; Matsunaga, H.; Iwamoto, M.; Kawai, F.; Ohashi, H.; Tsukuda, S.; Shimura, S.; Suzuki, R.; et al. A Novel Tricyclic Polyketide, Vanitaracin A, Specifically Inhibits the Entry of Hepatitis B and D Viruses by Targeting Sodium Taurocholate Cotransporting Polypeptide. J. Virol. 2015, 89, 11945–11953. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 (μM) | IC90 (μM) | CC50 (μM) |
|---|---|---|---|
| Cholesterol | >30 | >30 | >30 |
| 4-beta-OHC | >30 | >30 | >30 |
| 7-KC | 14.5 | >30 | >30 |
| 22(S)-OHC | >30 | >30 | >30 |
| 22(R)-OHC | 13.2 | >30 | >30 |
| 24(S)-OHC | 1.3 | >30 | >30 |
| 27-OHC | 3.5 | >30 | >30 |
| RDV | 1.5 | 2.5 | >10 |
| Oxy133 | >15 | >15 | >15 |
| Oxy186 | 7.4 | >15 | >15 |
| Oxy210 | 5.5 | 8.6 | >15 |
| Oxy232 | 5.4 | 7.8 | >15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohashi, H.; Wang, F.; Stappenbeck, F.; Tsuchimoto, K.; Kobayashi, C.; Saso, W.; Kataoka, M.; Yamasaki, M.; Kuramochi, K.; Muramatsu, M.; et al. Identification of Anti-Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-CoV-2) Oxysterol Derivatives In Vitro. Int. J. Mol. Sci. 2021, 22, 3163. https://doi.org/10.3390/ijms22063163
Ohashi H, Wang F, Stappenbeck F, Tsuchimoto K, Kobayashi C, Saso W, Kataoka M, Yamasaki M, Kuramochi K, Muramatsu M, et al. Identification of Anti-Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-CoV-2) Oxysterol Derivatives In Vitro. International Journal of Molecular Sciences. 2021; 22(6):3163. https://doi.org/10.3390/ijms22063163
Chicago/Turabian StyleOhashi, Hirofumi, Feng Wang, Frank Stappenbeck, Kana Tsuchimoto, Chisa Kobayashi, Wakana Saso, Michiyo Kataoka, Masako Yamasaki, Kouji Kuramochi, Masamichi Muramatsu, and et al. 2021. "Identification of Anti-Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-CoV-2) Oxysterol Derivatives In Vitro" International Journal of Molecular Sciences 22, no. 6: 3163. https://doi.org/10.3390/ijms22063163
APA StyleOhashi, H., Wang, F., Stappenbeck, F., Tsuchimoto, K., Kobayashi, C., Saso, W., Kataoka, M., Yamasaki, M., Kuramochi, K., Muramatsu, M., Suzuki, T., Sureau, C., Takeda, M., Wakita, T., Parhami, F., & Watashi, K. (2021). Identification of Anti-Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-CoV-2) Oxysterol Derivatives In Vitro. International Journal of Molecular Sciences, 22(6), 3163. https://doi.org/10.3390/ijms22063163