
De novo Neurosteroidogenesis in Human Microglia: Involvement of the 18 kDa Translocator Protein

 ,
,  , , , , , , ,
, , , , , , ,  and
and
Abstract
1. Introduction
2. Results
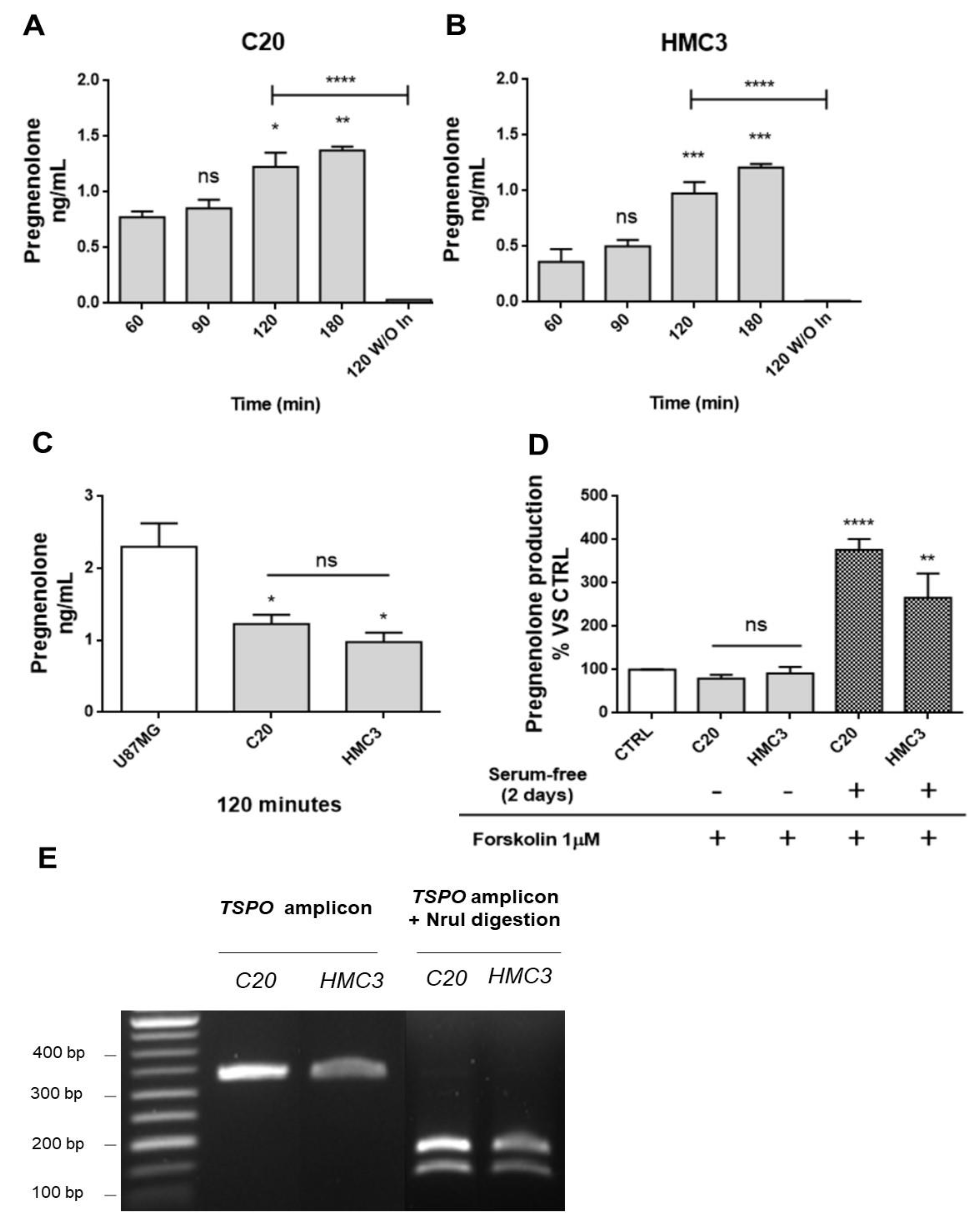
2.1. Human Microglia C20 and HMC3 Cells Produce Pregnenolone De novo
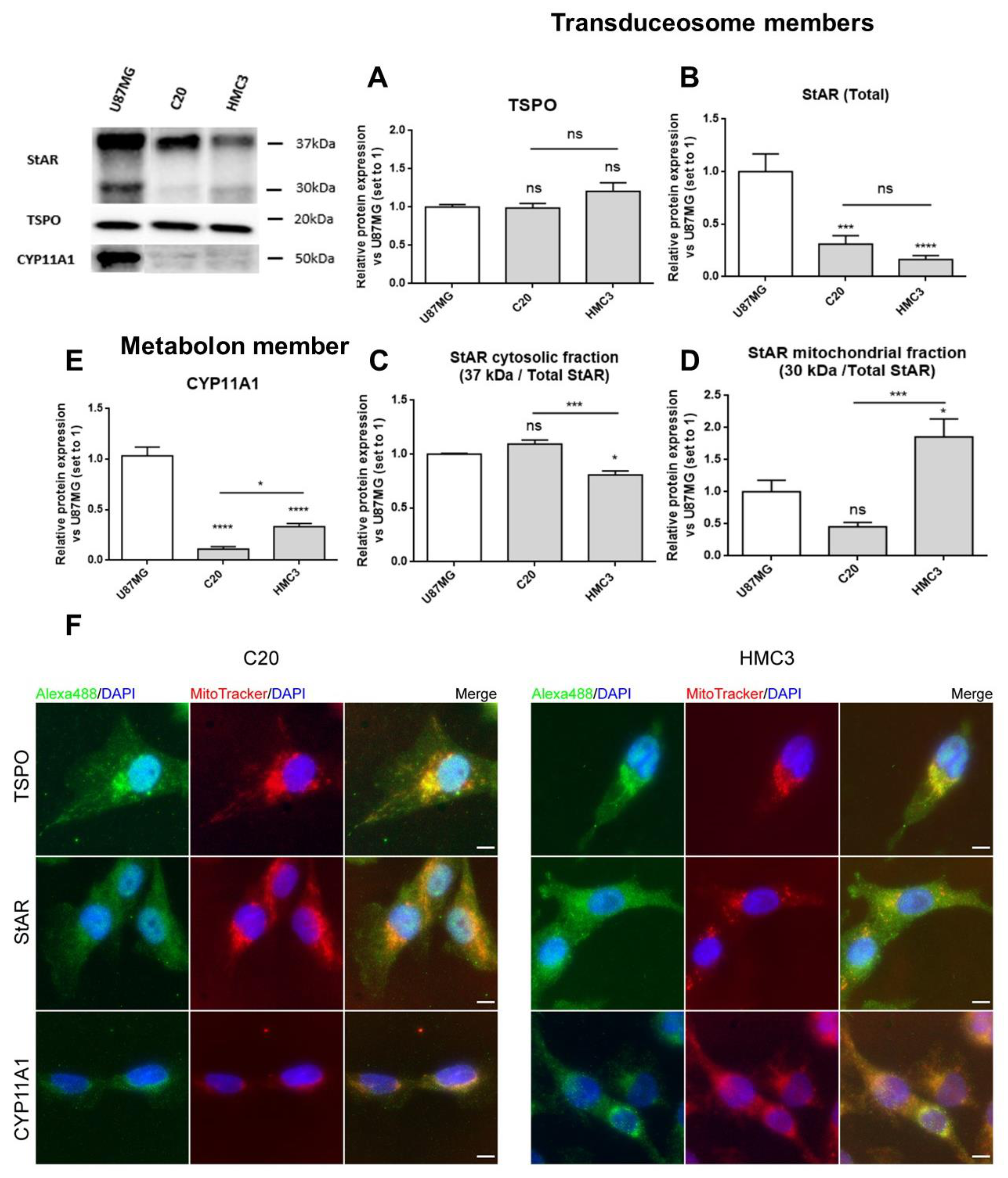
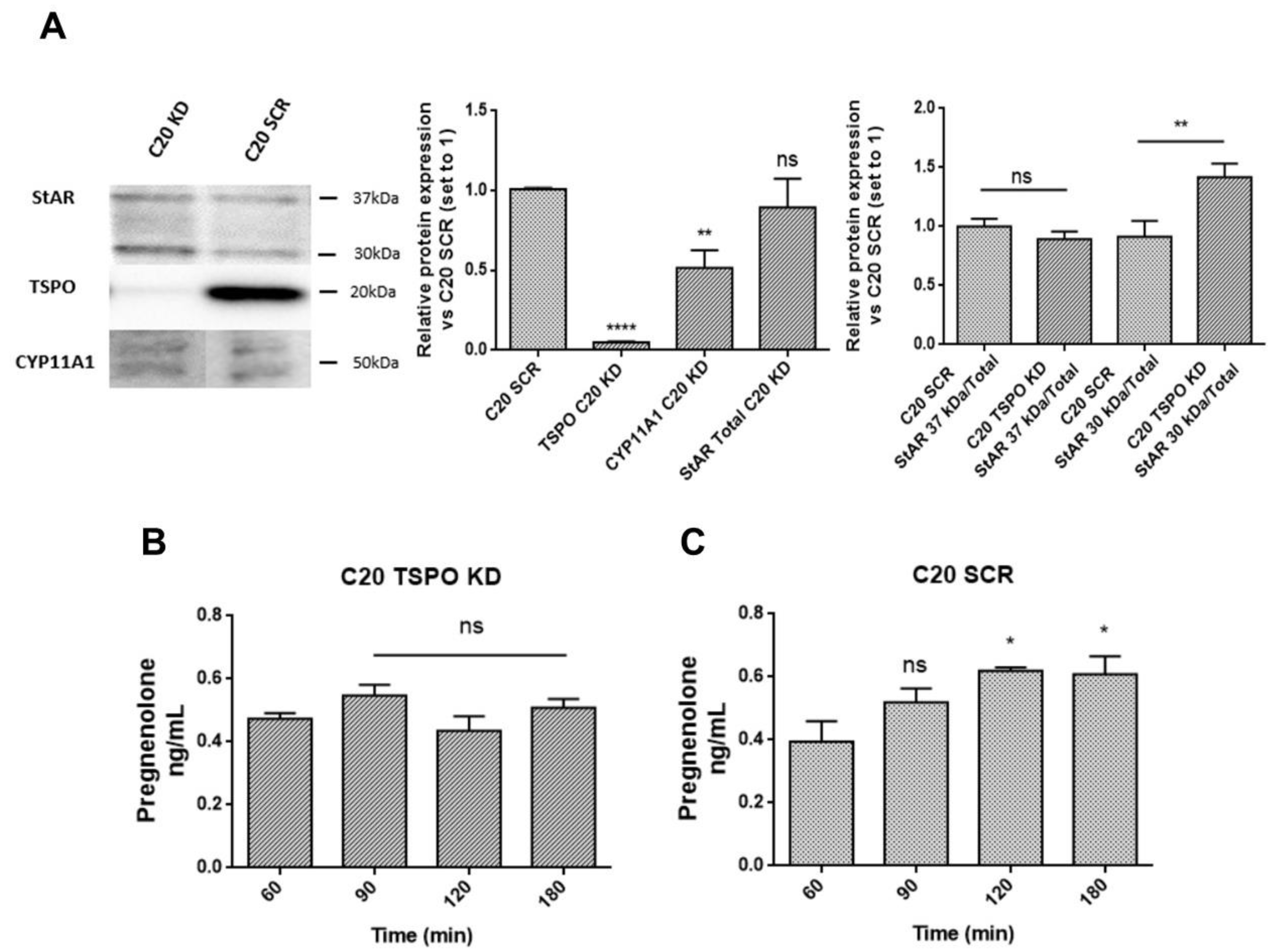
2.2. C20 and HMC3 Cells Express Transduceosome/Metabolon Key Proteins
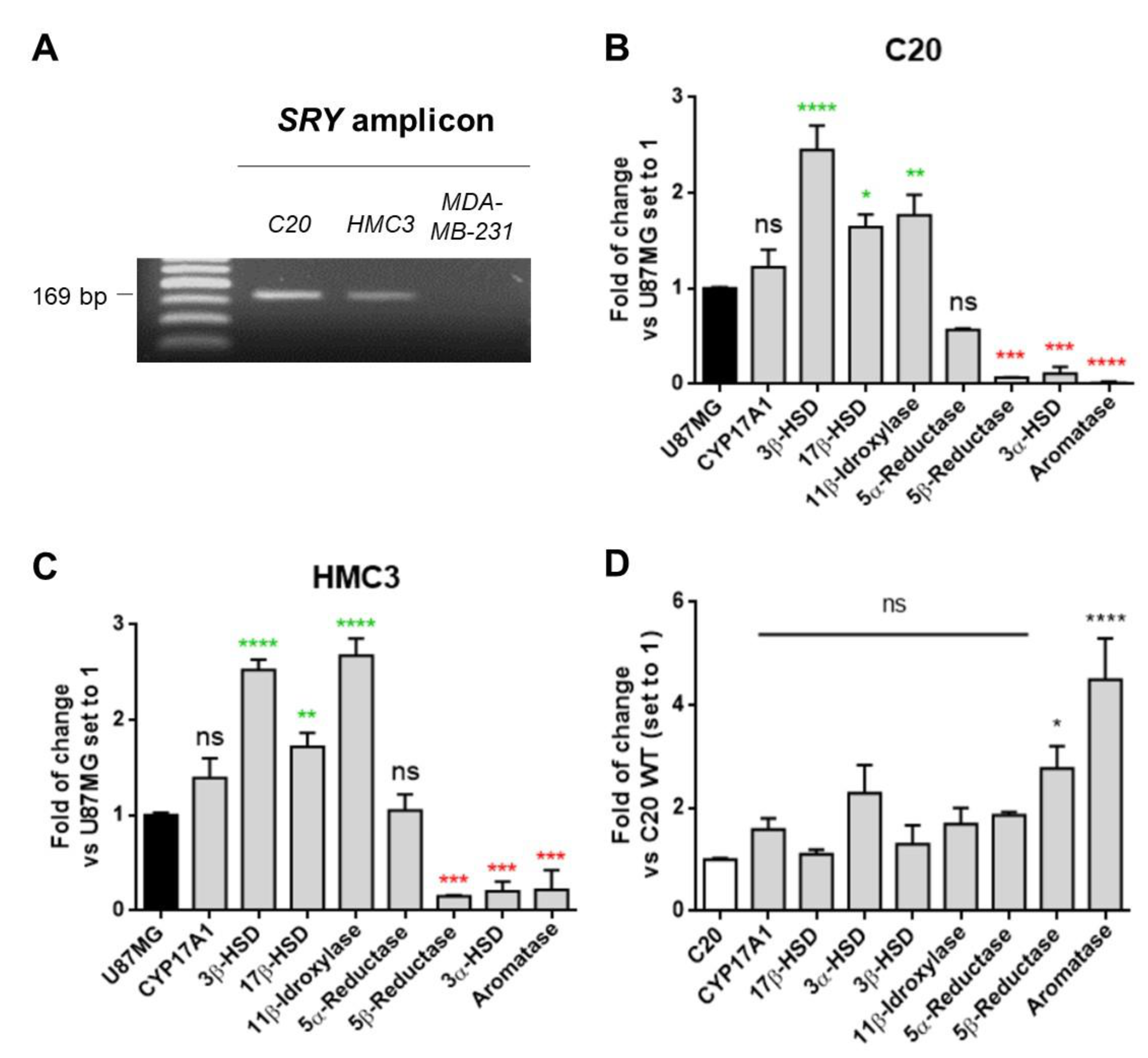
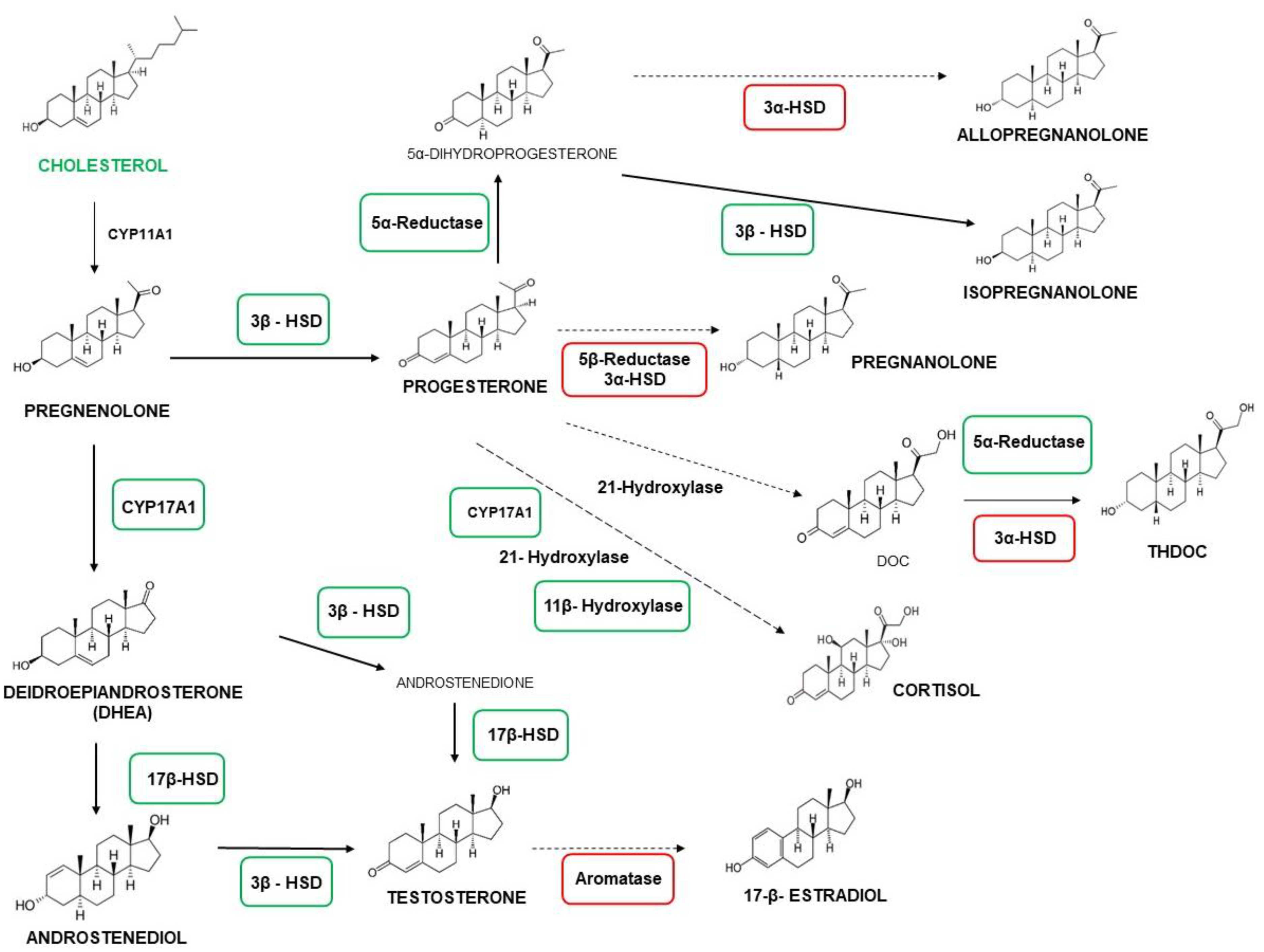
2.3. C20 and HMC3 Cells Showed Gene Expression of Key Steroidogenic Enzymes
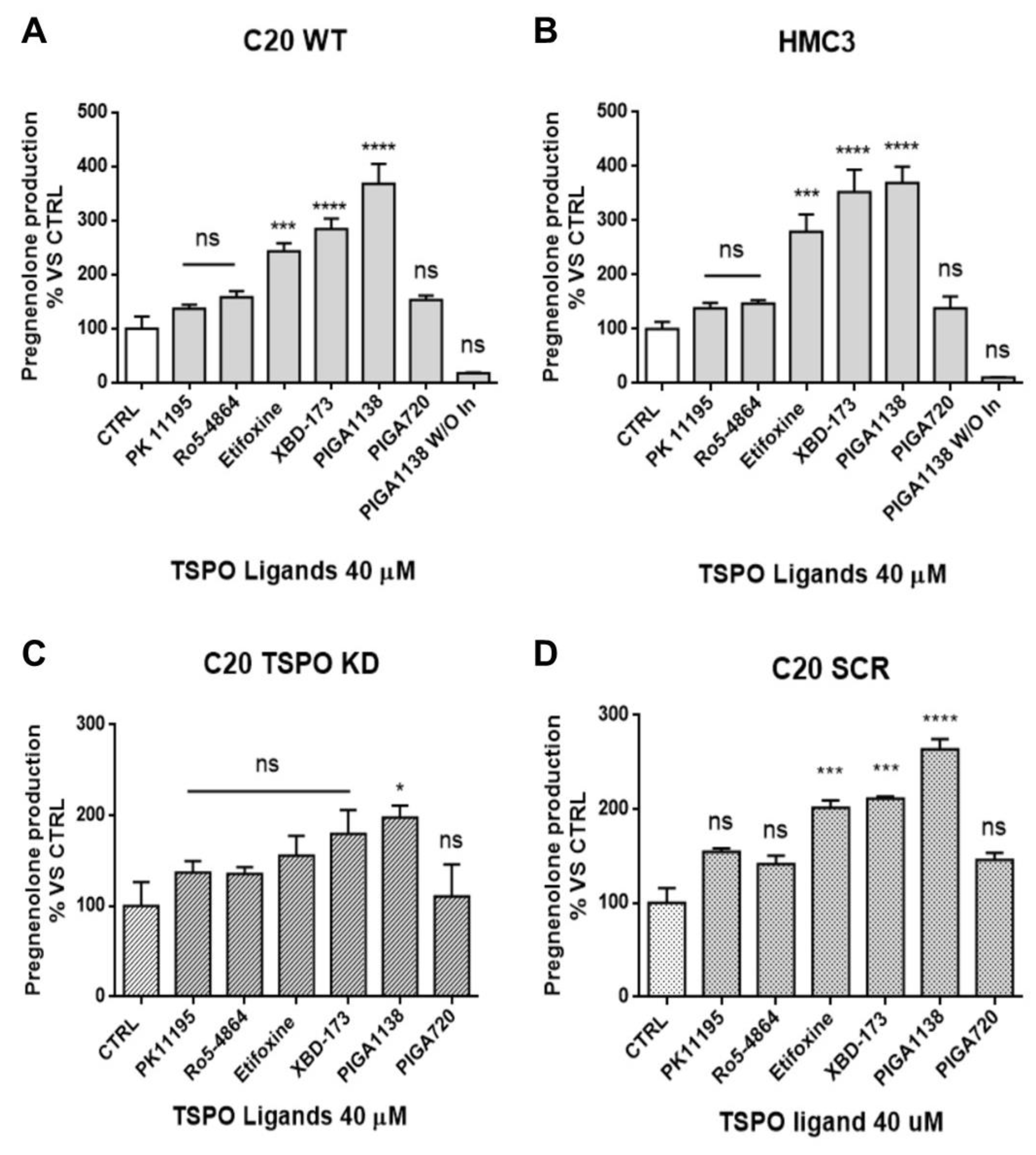
2.4. TSPO Is Required to Maintain Basal De novo Neurosteroidogenesis in C20 and HMC3 Cells
2.5. Potentiation of TSPO Function Increases Basal De novo Neurosteroidogenic Activity in C20 and HMC3 Cells
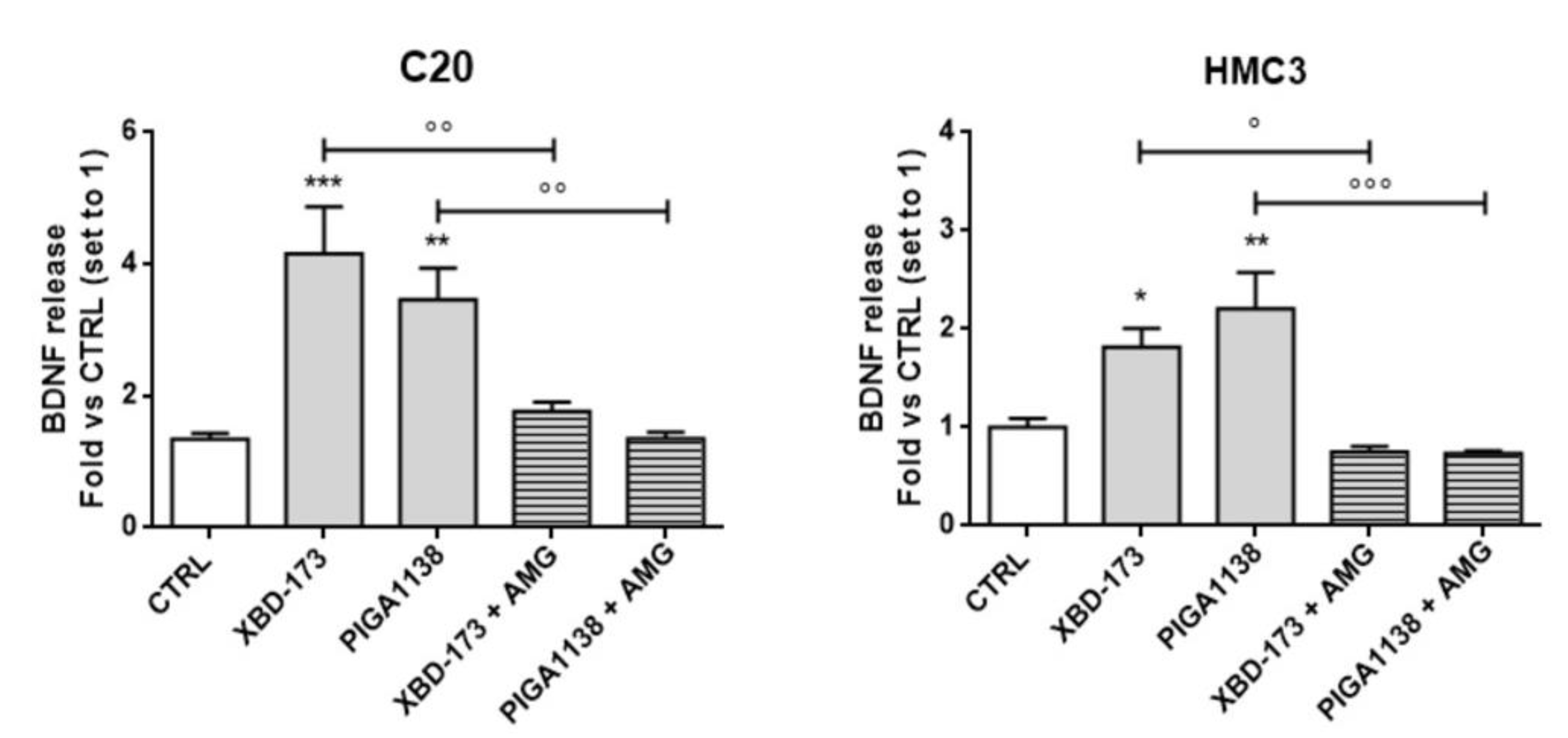
2.6. Potentiation of TSPO Function Increases Brain-Derived Neurotrophic Factor (BDNF) Production in C20 and HMC3 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Pregnenolone ELISA
4.4. Western Blot
4.5. Genotyping of Human Microglial Cells for TSPO rs6971 Polymorphism
4.6. Cell Sex Determination by SRY Gene PCR
4.7. Immunostaining of Transduceosome/Metabolon Protein
4.8. Real-Time RT-PCR of Human Microglial Neurosteroidogenic Enzymes
4.9. BDNF and TGF-β ELISA Quantification
4.10. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TSPO | Translocator Protein 18 KDa |
| StAR | Steroidogenic Acute Regulatory protein |
| CYP11A1 | Cytochrome P450 family 11 subfamily A member 1 |
| BDNF | Brain-derived neurotrophic factor |
| TGF-β | Transforming Growth Factor β |
| KD | Knockdown |
| CNS | Central Nervous System |
| CRAC | Cholesterol Recognition Aminoacidic Consensus |
| DHEA | Dehydroepiandrosterone |
| CYP17A1 | 17α-hydroxylase/C17-20 lyase |
| 3α-HSD | 3α-hydroxysteroid dehydrogenase |
| shRNA | Short Hairpin RNA |
| SCR | Scramble |
| AMG | Aminoglutethimide |
| OMM | Outer Mitochondrial Membrane |
| IMM | Inner Mitochondrial Membrane |
| R | Residence Time |
References
- Frost, J.L.; Schafer, D.P. Microglia: Architects of the Developing Nervous System. Trends Cell Biol. 2016, 26, 587–597. [Google Scholar] [CrossRef]
- Butovsky, O.; Weiner, H.L. Microglial signatures and their role in health and disease. Nat. Rev. Neurosci. 2018, 19, 622–635. [Google Scholar] [CrossRef]
- Augusto-Oliveira, M.; Arrifano, G.P.; Lopes-Araújo, A.; Santos-Sacramento, L.; Takeda, P.Y.; Anthony, D.C.; Malva, J.O.; Crespo-Lopez, M.E. What Do Microglia Really Do in Healthy Adult Brain? Cells 2019, 8, 1293. [Google Scholar] [CrossRef]
- Saijo, K.; Glass, C.K. Microglial cell origin and phenotypes in health and disease. Nat. Rev. Immunol. 2011, 11, 775–787. [Google Scholar] [CrossRef]
- Walker, D.G.; Lue, L.-F. Immune phenotypes of microglia in human neurodegenerative disease: Challenges to detecting microglial polarization in human brains. Alzheimer’s Res. Ther. 2015, 7, 56. [Google Scholar] [CrossRef]
- Franco, R.; Fernández-Suárez, D. Alternatively activated microglia and macrophages in the central nervous system. Prog. Neurobiol. 2015, 131, 65–86. [Google Scholar] [CrossRef]
- Chen, Z.; Trapp, B.D. Microglia and neuroprotection. J. Neurochem. 2016, 136, 10–17. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Wang, X.; Liu, C.; Zhang, H.-L. Pharmacological Targeting of Microglial Activation: New Therapeutic Approach. Front. Cell. Neurosci. 2019, 13, 514. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Zu, H.-B. Microglial polarization: Novel therapeutic mechanism against Alzheimer’s disease. Inflammopharmacology 2020, 28, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Beckers, L.; Ory, D.; Geric, I.; Declercq, L.; Koole, M.; Kassiou, M.; Bormans, G.; Baes, M. Increased Expression of Translocator Protein (TSPO) Marks Pro-inflammatory Microglia but Does Not Predict Neurodegeneration. Mol. Imaging Biol. 2018, 20, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Guilarte, T.R. TSPO in diverse CNS pathologies and psychiatric disease: A critical review and a way forward. Pharmacol. Ther. 2019, 194, 44–58. [Google Scholar] [CrossRef]
- Pannell, M.; Economopoulos, V.; Wilson, T.C.; Kersemans, V.; Isenegger, P.G.; Larkin, J.R.; Smart, S.; Gilchrist, S.; Gouverneur, V.; Sibson, N.R. Imaging of translocator protein upregulation is selective for pro-inflammatory polarized astrocytes and microglia. Glia 2020, 68, 280–297. [Google Scholar] [CrossRef]
- Liu, G.; Middleton, R.J.; Hatty, C.R.; Kam, W.W.; Chan, R.; Pham, T.; Harrison-Brown, M.; Dodson, E.; Veale, K.; Banati, R.B. The 18 kDa Translocator Protein, Microglia and Neuroinflammation. Brain Pathol. 2014, 24, 631–653. [Google Scholar] [CrossRef]
- Da Pozzo, E.; Tremolanti, C.; Costa, B.; Giacomelli, C.; Milenkovic, V.M.; Bader, S.; Wetzel, C.H.; Rupprecht, R.; Taliani, S.; Da Settimo, F.; et al. Microglial Pro-Inflammatory and Anti-Inflammatory Phenotypes Are Modulated by Translocator Protein Activation. Int. J. Mol. Sci. 2019, 20, 4467. [Google Scholar] [CrossRef]
- Werry, E.L.; Bright, F.M.; Piguet, O.; Ittner, L.M.; Halliday, G.M.; Hodges, J.R.; Kiernan, M.C.; Loy, C.T.; Kril, J.J.; Kassiou, M. Recent Developments in TSPO PET Imaging as A Biomarker of Neuroinflammation in Neurodegenerative Disorders. Int. J. Mol. Sci. 2019, 20, 3161. [Google Scholar] [CrossRef]
- Arbo, B.; Benetti, F.; Garcia-Segura, L.; Ribeiro, M. Therapeutic actions of translocator protein (18 kDa) ligands in experimental models of psychiatric disorders and neurodegenerative diseases. J. Steroid Biochem. Mol. Biol. 2015, 154, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Dimitrova-Shumkovska, J.; Krstanoski, L.; Veenman, L. Diagnostic and Therapeutic Potential of TSPO Studies Regarding Neurodegenerative Diseases, Psychiatric Disorders, Alcohol Use Disorders, Traumatic Brain Injury, and Stroke: An Update. Cells 2020, 9, 870. [Google Scholar] [CrossRef] [PubMed]
- Barresi, E.; Robello, M.; Costa, B.; Da Pozzo, E.; Baglini, E.; Salerno, S.; Da Settimo, F.; Martini, C.; Taliani, S. An update into the medicinal chemistry of translocator protein (TSPO) ligands. Eur. J. Med. Chem. 2021, 209, 112924. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, V.; Fan, J.; Zirkin, B. Translocator protein (18 kDa): An update on its function in steroidogenesis. J. Neuroendocr. 2018, 30, e12500. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Da Pozzo, E.; Martini, C. Translocator protein and steroidogenesis. Biochem. J. 2018, 475, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Porcu, P.; Barron, A.M.; Frye, C.A.; Walf, A.A.; Yang, S.-Y.; He, X.-Y.; Morrow, A.L.; Panzica, G.C.; Melcangi, R.C. Neurosteroidogenesis Today: Novel Targets for Neuroactive Steroid Synthesis and Action and Their Relevance for Translational Research. J. Neuroendocr. 2016, 28, 12351. [Google Scholar] [CrossRef]
- Yilmaz, C.; Karali, K.; Fodelianaki, G.; Gravanis, A.; Chavakis, T.; Charalampopoulos, I.; Alexaki, V.I. Neurosteroids as regulators of neuroinflammation. Front. Neuroendocr. 2019, 55, 100788. [Google Scholar] [CrossRef]
- Costa, B.; Da Pozzo, E.; Martini, C. 18-kDa translocator protein association complexes in the brain: From structure to function. Biochem. Pharmacol. 2020, 177, 114015. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. StAR Search—What We Know about How the Steroidogenic Acute Regulatory Protein Mediates Mitochondrial Cholesterol Import. Mol. Endocrinol. 2007, 21, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Rone, M.B.; Fan, J.; Papadopoulos, V. Cholesterol transport in steroid biosynthesis: Role of protein–protein interactions and implications in disease states. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2009, 1791, 646–658. [Google Scholar] [CrossRef]
- De Nicola, A.F.; Garay, L.I.; Meyer, M.; Guennoun, R.; Sitruk-Ware, R.; Schumacher, M.; Deniselle, M.C.G. Neurosteroidogenesis and progesterone anti-inflammatory/neuroprotective effects. J. Neuroendocr. 2018, 30, e12502. [Google Scholar] [CrossRef] [PubMed]
- Lanussa, O.H.; Ávila-Rodriguez, M.; García-Segura, L.M.; González, J.; Echeverria, V.; Aliev, G.; Barreto, G.E. Microglial dependent protective effects of neuroactive steroids. CNS Neurol. Disord. Drug Targets 2016, 15, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Zwain, I.H.; Yen, S.S.C. Neurosteroidogenesis in Astrocytes, Oligodendrocytes, and Neurons of Cerebral Cortex of Rat Brain. Endocrinology 1999, 140, 3843–3852. [Google Scholar] [CrossRef] [PubMed]
- Melcangi, R.C.; Azcoitia, I.; Galbiati, M.; Magnaghi, V.; Garcia-Ovejero, D.; Garcia-Segura, L.M. Non-neuronal cells in the nervous system: Sources and targets of neuroactive steroids. Gap Junctions 2003, 31, 535–559. [Google Scholar] [CrossRef]
- Gago, N.; El-Etr, M.; Sananès, N.; Cadepond, F.; Samuel, D.; Avellana-Adalid, V.; Evercooren, A.B.-V.; Schumacher, M. 3α,5α-tetrahydroprogesterone (allopregnanolone) and γ-aminobutyric acid: Autocrine/paracrine interactions in the control of neonatal PSA-NCAM+progenitor proliferation. J. Neurosci. Res. 2004, 78, 770–783. [Google Scholar] [CrossRef]
- Gottfried-Blackmore, A.; Sierra, A.; Jellinck, P.H.; McEwen, B.S.; Bulloch, K. Brain microglia express steroid-converting enzymes in the mouse. J. Steroid Biochem. Mol. Biol. 2008, 109, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Vegeto, E.; Villa, A.; Della Torre, S.; Crippa, V.; Rusmini, P.; Cristofani, R.; Galbiati, M.; Maggi, A.; Poletti, A. The Role of Sex and Sex Hormones in Neurodegenerative Diseases. Endocr. Rev. 2020, 41, 273–319. [Google Scholar] [CrossRef]
- Owen, D.R.; Narayan, N.; Wells, L.; Healy, L.; Smyth, E.; A Rabiner, E.; Galloway, D.; Williams, J.B.; Lehr, J.; Mandhair, H.; et al. Pro-inflammatory activation of primary microglia and macrophages increases 18 kDa translocator protein expression in rodents but not humans. Br. J. Pharmacol. 2017, 37, 2679–2690. [Google Scholar] [CrossRef]
- Avallone, R.; Lucchi, C.; Puja, G.; Codeluppi, A.; Filaferro, M.; Vitale, G.; Rustichelli, C.; Biagini, G. BV-2 Microglial Cells Respond to Rotenone Toxic Insult by Modifying Pregnenolone, 5α-Dihydroprogesterone and Pregnanolone Levels. Cells 2020, 9, 2091. [Google Scholar] [CrossRef] [PubMed]
- Saijo, K.; Collier, J.G.; Li, A.C.; Katzenellenbogen, J.A.; Glass, C.K. An ADIOL-ERβ-CtBP Transrepression Pathway Negatively Regulates Microglia-Mediated Inflammation. Cell 2011, 145, 584–595. [Google Scholar] [CrossRef]
- Sohrabji, F.; Miranda, R.C.; Toran-Allerand, C.D. Identification of a putative estrogen response element in the gene encoding brain-derived neurotrophic factor. Proc. Natl. Acad. Sci. USA 1995, 92, 11110–11114. [Google Scholar] [CrossRef]
- Pluchino, N.; Russo, M.; Santoro, A.; Litta, P.; Cela, V.; Genazzani, A. Steroid hormones and BDNF. Neuroscience 2013, 239, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-H.; Kato, H.; Chen, S.-T.; Kogure, K.; Itoyama, Y. Expression disparity of brain-derived neurotrophic factor immunoreactivity and mRNA in ischemic hippocampal neurons. NeuroReport 2002, 13, 2271–2275. [Google Scholar] [CrossRef] [PubMed]
- Shulga, A.; Thomas-Crusells, J.; Sigl, T.; Blaesse, A.; Mestres, P.; Meyer, M.; Yan, Q.; Kaila, K.; Saarma, M.; Rivera, C.; et al. Posttraumatic GABAA-Mediated [Ca2+]i Increase Is Essential for the Induction of Brain-Derived Neurotrophic Factor-Dependent Survival of Mature Central Neurons. J. Neurosci. 2008, 28, 6996–7005. [Google Scholar] [CrossRef]
- Parkhurst, C.N.; Yang, G.; Ninan, I.; Savas, J.N.; Yates, J.R., III; Lafaille, J.J.; Hempstead, B.L.; Littman, D.R.; Gan, W.-B. Microglia promote learning-dependent synapse formation through brain-derived neurotrophic factor. Cell 2013, 155, 1596–1609. [Google Scholar] [CrossRef] [PubMed]
- Russo, C.D.; Cappoli, N.; Coletta, I.; Mezzogori, D.; Paciello, F.; Pozzoli, G.; Navarra, P.; Battaglia, A. The human microglial HMC3 cell line: Where do we stand? A systematic literature review. J. Neuroinflammation 2018, 15, 1–24. [Google Scholar] [CrossRef]
- Garcia-Mesa, Y.; Jay, T.R.; Checkley, M.A.; Luttge, B.; Dobrowolski, C.; Valadkhan, S.; Landreth, G.E.; Karn, J.; Alvarez-Carbonell, D. Immortalization of primary microglia: A new platform to study HIV regulation in the central nervous system. J. Neurovirology 2017, 23, 47–66. [Google Scholar] [CrossRef]
- Davis, R.L.; Buck, D.J.; McCracken, K.; Cox, G.W.; Das, S. Interleukin-1β-induced inflammatory signaling in C20 human microglial cells. Neuroimmunol. Neuroinflammation 2018, 2018, 50. [Google Scholar] [CrossRef]
- Da Pozzo, E.; Giacomelli, C.; Costa, B.; Cavallini, C.; Taliani, S.; Barresi, E.; Da Settimo, F.; Martini, C. TSPO PIGA Ligands Promote Neurosteroidogenesis and Human Astrocyte Well-Being. Int. J. Mol. Sci. 2016, 17, 1028. [Google Scholar] [CrossRef]
- Karri, S.; Dertien, J.S.; Stocco, D.M.; Syapin, P.J. Steroidogenic Acute Regulatory Protein Expression and Pregnenolone Synthesis in Rat Astrocyte Cultures. J. Neuroendocr. 2007, 19, 860–869. [Google Scholar] [CrossRef]
- Liu, T.; Wimalasena, J.; Bowen, R.L.; Atwood, C.S. Luteinizing hormone receptor mediates neuronal pregnenolone production via up-regulation of steroidogenic acute regulatory protein expression. J. Neurochem. 2006, 100, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, V.M.; Slim, D.; Bader, S.; Koch, V.; Heinl, E.-S.; Alvarez-Carbonell, D.; Nothdurfter, C.; Rupprecht, R.; Wetzel, C.H. CRISPR-Cas9 Mediated TSPO Gene Knockout alters Respiration and Cellular Metabolism in Human Primary Microglia Cells. Int. J. Mol. Sci. 2019, 20, 3359. [Google Scholar] [CrossRef]
- Yao, Y.; Fu, K.-Y. Serum-deprivation leads to activation-like changes in primary microglia and BV-2 cells but not astrocytes. Biomed. Rep. 2020, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Pini, S.; Gabelloni, P.; Da Pozzo, E.; Abelli, M.; Lari, L.; Preve, M.; Lucacchini, A.; Cassano, G.B.; Martini, C. The Spontaneous Ala147Thr Amino Acid Substitution within the Translocator Protein Influences Pregnenolone Production in Lymphomonocytes of Healthy Individuals. Endocrinology 2009, 150, 5438–5445. [Google Scholar] [CrossRef] [PubMed]
- Rechichi, M.; Salvetti, A.; Chelli, B.; Costa, B.; Da Pozzo, E.; Spinetti, F.; Lena, A.; Evangelista, M.; Rainaldi, G.; Martini, C.; et al. TSPO over-expression increases motility, transmigration and proliferation properties of C6 rat glioma cells. Biochim. Biophys. Acta Mol. Basis Dis. 2008, 1782, 118–125. [Google Scholar] [CrossRef]
- Yasin, N.; Veenman, L.; Singh, S.; Azrad, M.; Bode, J.; Vainshtein, A.; Caballero, B.; Marek, I.; Gavish, M. Classical and Novel TSPO Ligands for the Mitochondrial TSPO Can Modulate Nuclear Gene Expression: Implications for Mitochondrial Retrograde Signaling. Int. J. Mol. Sci. 2017, 18, 786. [Google Scholar] [CrossRef]
- Utsumi, K.; Iritani, A. Embryo sexing by male specific antibody and by PCR using male specific (SRY) primer. Mol. Reprod. Dev. 1993, 36, 238–241. [Google Scholar] [CrossRef]
- Girdler, S.S.; Klatzkin, R. Neurosteroids in the context of stress: Implications for depressive disorders. Pharmacol. Ther. 2007, 116, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Da Pozzo, E.; Cavallini, C.; Taliani, S.; Da Settimo, F.; Martini, C. Long Residence Time at the Neurosteroidogenic 18 kDa Translocator Protein Characterizes the Anxiolytic Ligand XBD173. ACS Chem. Neurosci. 2016, 7, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Da Pozzo, E.; Giacomelli, C.; Barresi, E.; Taliani, S.; Da Settimo, F.; Martini, C. TSPO ligand residence time: A new parameter to predict compound neurosteroidogenic efficacy. Sci. Rep. 2016, 6, 18164. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Cavallini, C.; Da Pozzo, E.; Taliani, S.; Da Settimo, F.; Martini, C. The Anxiolytic Etifoxine Binds to TSPO Ro5-4864 Binding Site with Long Residence Time Showing a High Neurosteroidogenic Activity. ACS Chem. Neurosci. 2017, 8, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Barresi, E.; Bruno, A.; Taliani, S.; Cosconati, S.; Da Pozzo, E.; Salerno, S.; Simorini, F.; Daniele, S.; Giacomelli, C.; Marini, A.M.; et al. Deepening the Topology of the Translocator Protein Binding Site by Novel N,N-Dialkyl-2-arylindol-3-ylglyoxylamides. J. Med. Chem. 2015, 58, 6081–6092. [Google Scholar] [CrossRef] [PubMed]
- Wolf, L.; Bauer, A.; Melchner, D.; Hallof-Buestrich, H.; Stoertebecker, P.; Haen, E.; Kreutz, M.; Sarubin, N.; Milenkovic, V.M.; Wetzel, C.H.; et al. Enhancing Neurosteroid Synthesis—Relationship to the Pharmacology of Translocator Protein (18 kDa) (TSPO) Ligands and Benzodiazepines. Pharmacopsychiatry 2015, 48, 72–77. [Google Scholar] [CrossRef]
- Daugherty, D.J.; Selvaraj, V.; Chechneva, O.V.; Liu, X.-B.; Pleasure, D.E.; Deng, W. A TSPO ligand is protective in a mouse model of multiple sclerosis. EMBO Mol. Med. 2013, 5, 891–903. [Google Scholar] [CrossRef]
- Betlazar, C.; Middleton, R.J.; Banati, R.; Liu, G.-J. The Translocator Protein (TSPO) in Mitochondrial Bioenergetics and Immune Processes. Cells 2020, 9, 512. [Google Scholar] [CrossRef] [PubMed]
- Monga, S.; Nagler, R.; Amara, R.; Weizman, A.; Gavish, M. Inhibitory Effects of the Two Novel TSPO Ligands 2-Cl-MGV-1 and MGV-1 on LPS-induced Microglial Activation. Cells 2019, 8, 486. [Google Scholar] [CrossRef]
- Naert, G.; Maurice, T.; Tapia-Arancibia, L.; Givalois, L. Neuroactive steroids modulate HPA axis activity and cerebral brain-derived neurotrophic factor (BDNF) protein levels in adult male rats. Psychoneuroendocrinology 2007, 32, 1062–1078. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, C.C.; Baudry, M. Progesterone reverses 17β-estradiol-mediated neuroprotection and BDNF induction in cultured hippocampal slices. Eur. J. Neurosci. 2009, 29, 447–454. [Google Scholar] [CrossRef]
- Zhou, D.; Ji, L.; Chen, Y. TSPO Modulates IL-4-Induced Microglia/Macrophage M2 Polarization via PPAR-γ Pathway. J. Mol. Neurosci. 2019, 70, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, H.; Liu, N.; Wang, P.-Q.; Guo, W.-Z.; Fu, Q.; Jiao, L.-B.; Ma, Y.-Q.; Mi, W.-D. TSPO ligand PK11195 alleviates neuroinflammation and beta-amyloid generation induced by systemic LPS administration. Brain Res. Bull. 2016, 121, 192–200. [Google Scholar] [CrossRef]
- Kaushik, D.K.; Thounaojam, M.C.; Kumawat, K.L.; Gupta, M.; Basu, A. Interleukin-1β orchestrates underlying inflammatory responses in microglia via Krüppel-like factor 4. J. Neurochem. 2013, 127, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Garcia, C.; Atif, F.; Yousuf, S.; Sayeed, I.; Neigh, G.N.; Stein, D.G. Progesterone Attenuates Stress-Induced NLRP3 Inflammasome Activation and Enhances Autophagy Following Ischemic Brain Injury. Int. J. Mol. Sci. 2020, 21, 3740. [Google Scholar] [CrossRef]
- Aryanpour, R.; Pasbakhsh, P.; Zibara, K.; Namjoo, Z.; Boroujeni, F.B.; Shahbeigi, S.; Kashani, I.R.; Beyer, C.; Zendehdel, A. Progesterone therapy induces an M1 to M2 switch in microglia phenotype and suppresses NLRP3 inflammasome in a cuprizone-induced demyelination mouse model. Int. Immunopharmacol. 2017, 51, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Brotfain, E.; Gruenbaum, S.E.; Boyko, M.; Kutz, R.; Zlotnik, A.; Klein, M. Neuroprotection by Estrogen and Progesterone in Traumatic Brain Injury and Spinal Cord Injury. Curr. Neuropharmacol. 2016, 14, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.M.; Rozovsky, I.; Arimoto, J.M.; Du, Y.; Wei, M.; Morgan, T.E.; Finch, C.E. Progesterone Influence on Neurite Outgrowth Involves Microglia. Endocrinology 2008, 150, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Campioli, E.; Midzak, A.; Culty, M.; Papadopoulos, V. Conditional steroidogenic cell-targeted deletion of TSPO unveils a crucial role in viability and hormone-dependent steroid formation. Proc. Natl. Acad. Sci. USA 2015, 112, 7261–7266. [Google Scholar] [CrossRef]
- Fan, J.; Campioli, E.; Sottas, C.; Zirkin, B.; Papadopoulos, V. Amhr2-Cre–Mediated Global Tspo Knockout. J. Endocr. Soc. 2020, 4, bvaa001. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Wang, K.; Zirkin, B.; Papadopoulos, V. CRISPR/Cas9‒Mediated Tspo Gene Mutations Lead to Reduced Mitochondrial Membrane Potential and Steroid Formation in MA-10 Mouse Tumor Leydig Cells. Endocrinology 2018, 159, 1130–1146. [Google Scholar] [CrossRef] [PubMed]
- Balthazart, J.; Choleris, E.; Remage-Healey, L. Steroids and the brain: 50 years of research, conceptual shifts and the ascent of non-classical and membrane-initiated actions. Horm. Behav. 2018, 99, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tuem, K.B.; Atey, T.M. Neuroactive Steroids: Receptor Interactions and Responses. Front. Neurol. 2017, 8, 442. [Google Scholar] [CrossRef] [PubMed]
- Giatti, S.; Garcia-Segura, L.M.; Barreto, G.E.; Melcangi, R.C. Neuroactive steroids, neurosteroidogenesis and sex. Prog. Neurobiol. 2019, 176, 1–17. [Google Scholar] [CrossRef]
- Wang, T.; Ye, X.; Bian, W.; Chen, Z.; Du, J.; Li, M.; Zhou, P.; Cui, H.; Ding, Y.-Q.; Qi, S.; et al. Allopregnanolone Modulates GABAAR-Dependent CaMKIIδ3 and BDNF to Protect SH-SY5Y Cells Against 6-OHDA-Induced Damage. Front. Cell. Neurosci. 2020, 13, 569. [Google Scholar] [CrossRef]
- Meyer, M.; Garay, L.I.; Kruse, M.S.; Lara, A.; Gargiulo-Monachelli, G.; Schumacher, M.; Guennoun, R.; Coirini, H.; De Nicola, A.F.; Deniselle, M.C.G. Protective effects of the neurosteroid allopregnanolone in a mouse model of spontaneous motoneuron degeneration. J. Steroid Biochem. Mol. Biol. 2017, 174, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Nin, M.S.; Martinez, L.A.; Pibiri, F.; Nelson, M.; Pinna, G. Neurosteroids reduce social isolation-induced behavioral deficits: A proposed link with neurosteroid-mediated upregulation of BDNF expression. Front. Endocrinol. 2011, 2, 73. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.B.; Gomez, R.; Barros, H.M.T.; Nin, M.S. Hemisphere-dependent Changes in mRNA Expression of GABAA Receptor Subunits and BDNF after Intra-prefrontal Cortex Allopregnanolone Infusion in Rats. Neuroscience 2019, 397, 56–66. [Google Scholar] [CrossRef]
- Primofiore, G.; Da Settimo, F.; Taliani, S.; Simorini, F.; Patrizi, M.P.; Novellino, E.; Greco, G.; Abignente, E.; Costa, B.; Chelli, B.; et al. N,N-Dialkyl-2-phenylindol-3-ylglyoxylamides. A New Class of Potent and Selective Ligands at the Peripheral Benzodiazepine Receptor. J. Med. Chem. 2004, 47, 1852–1855. [Google Scholar] [CrossRef] [PubMed]
- Voets, M.; Antes, I.; Scherer, C.; Müller-Vieira, U.; Biemel, K.; Marchais-Oberwinkler, S.; Hartmann, R.W. Synthesis and Evaluation of Heteroaryl-Substituted Dihydronaphthalenes and Indenes: Potent and Selective Inhibitors of Aldosterone Synthase (CYP11B2) for the Treatment of Congestive Heart Failure and Myocardial Fibrosis. J. Med. Chem. 2006, 49, 2222–2231. [Google Scholar] [CrossRef] [PubMed]
- Gilda, J.E.; Gomes, A.V. Western Blotting Using In-Gel Protein Labeling as a Normalization Control: Stain-Free Technology. Methods Mol. Biol. 2015, 1295, 381–391. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Nucleotide Sequence | Anneal. Temp. | Product Size |
|---|---|---|---|
| CYP17A1 | F: TAGGGGACATCTTTGGGGCTGGC R: CACTGATAGTTGGTGTGCGGCT | 58 °C | 150 bp |
| 3α-HSD | F: GGTCACTTCATGCCTGTCCTGGG R: GCCAGTCCAACCTGCTCCTCA | 65 °C | 155 bp |
| 3β-HSD | F: TGTGAAAGGTACCCAGCTCCTGT R: GGAGCGGGCCATGTGTTTTCC | 55 °C | 159 bp |
| 17β-HSD | F: TGGGGCTGCCTTTCAATGACGT R: CGATCAGGCTCAAGTGGACCCCA | 55 °C | 111 bp |
| 5α-Reductase | F: TTGGCTTGTGGTTAACGGGC R: AAGCCGCGCCTTGGACAGAC | 58 °C | 198 bp |
| 5β-Reductase | F: GGTCCTCCAGCTAGATTATGTGG R: ACCATTTGCCATTCTCATCTCT | 67 °C | 101 bp |
| 11β-hydroxylase | F: TCGGAACCCCAACGTGCAGCA R: GCACCAAGTCTGAGCTCGCCA | 65 °C | 182 bp |
| 21-hydroxylase | F: AGGAGTTCTGTGAGCGCATGAG R: GGCAGGCATTAAGTTGTCGTCC | 68 °C | 137 bp |
| Aromatase | F: GCATGGCAAGCTCTCCTCATCA R: TCAACTCAGTGGCAAAGTCCA | 65 °C | 181 bp |
| β-actin | F: GCACTCTTCCAGCCTTCCTTCC R: GAGCCGCCGATCCACACG | 58 °C | 254 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germelli, L.; Da Pozzo, E.; Giacomelli, C.; Tremolanti, C.; Marchetti, L.; Wetzel, C.H.; Barresi, E.; Taliani, S.; Da Settimo, F.; Martini, C.; et al. De novo Neurosteroidogenesis in Human Microglia: Involvement of the 18 kDa Translocator Protein. Int. J. Mol. Sci. 2021, 22, 3115. https://doi.org/10.3390/ijms22063115
Germelli L, Da Pozzo E, Giacomelli C, Tremolanti C, Marchetti L, Wetzel CH, Barresi E, Taliani S, Da Settimo F, Martini C, et al. De novo Neurosteroidogenesis in Human Microglia: Involvement of the 18 kDa Translocator Protein. International Journal of Molecular Sciences. 2021; 22(6):3115. https://doi.org/10.3390/ijms22063115
Chicago/Turabian StyleGermelli, Lorenzo, Eleonora Da Pozzo, Chiara Giacomelli, Chiara Tremolanti, Laura Marchetti, Christian H. Wetzel, Elisabetta Barresi, Sabrina Taliani, Federico Da Settimo, Claudia Martini, and et al. 2021. "De novo Neurosteroidogenesis in Human Microglia: Involvement of the 18 kDa Translocator Protein" International Journal of Molecular Sciences 22, no. 6: 3115. https://doi.org/10.3390/ijms22063115
APA StyleGermelli, L., Da Pozzo, E., Giacomelli, C., Tremolanti, C., Marchetti, L., Wetzel, C. H., Barresi, E., Taliani, S., Da Settimo, F., Martini, C., & Costa, B. (2021). De novo Neurosteroidogenesis in Human Microglia: Involvement of the 18 kDa Translocator Protein. International Journal of Molecular Sciences, 22(6), 3115. https://doi.org/10.3390/ijms22063115