
Transcriptional Changes in Potato Sprouts upon Interaction with Rhizoctonia solani Indicate Pathogen-Induced Interference in the Defence Pathways of Potato

 ,
,
Abstract
1. Introduction
2. Results
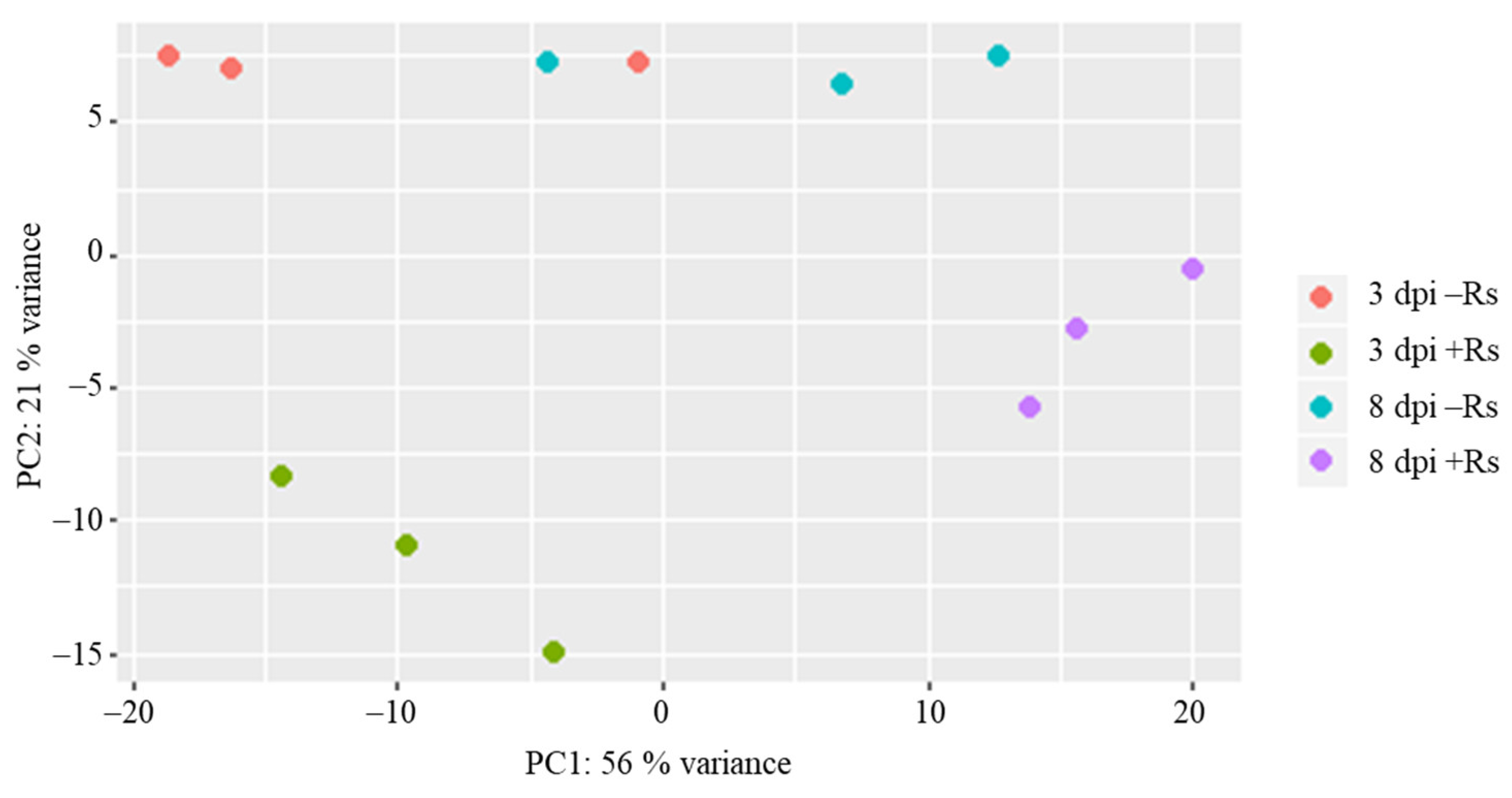
2.1. Sequencing and Transcriptome Assembly Statistics of Potato cv. Arkula
2.2. Overall Expressed Transcripts in Potato cv. Arkula
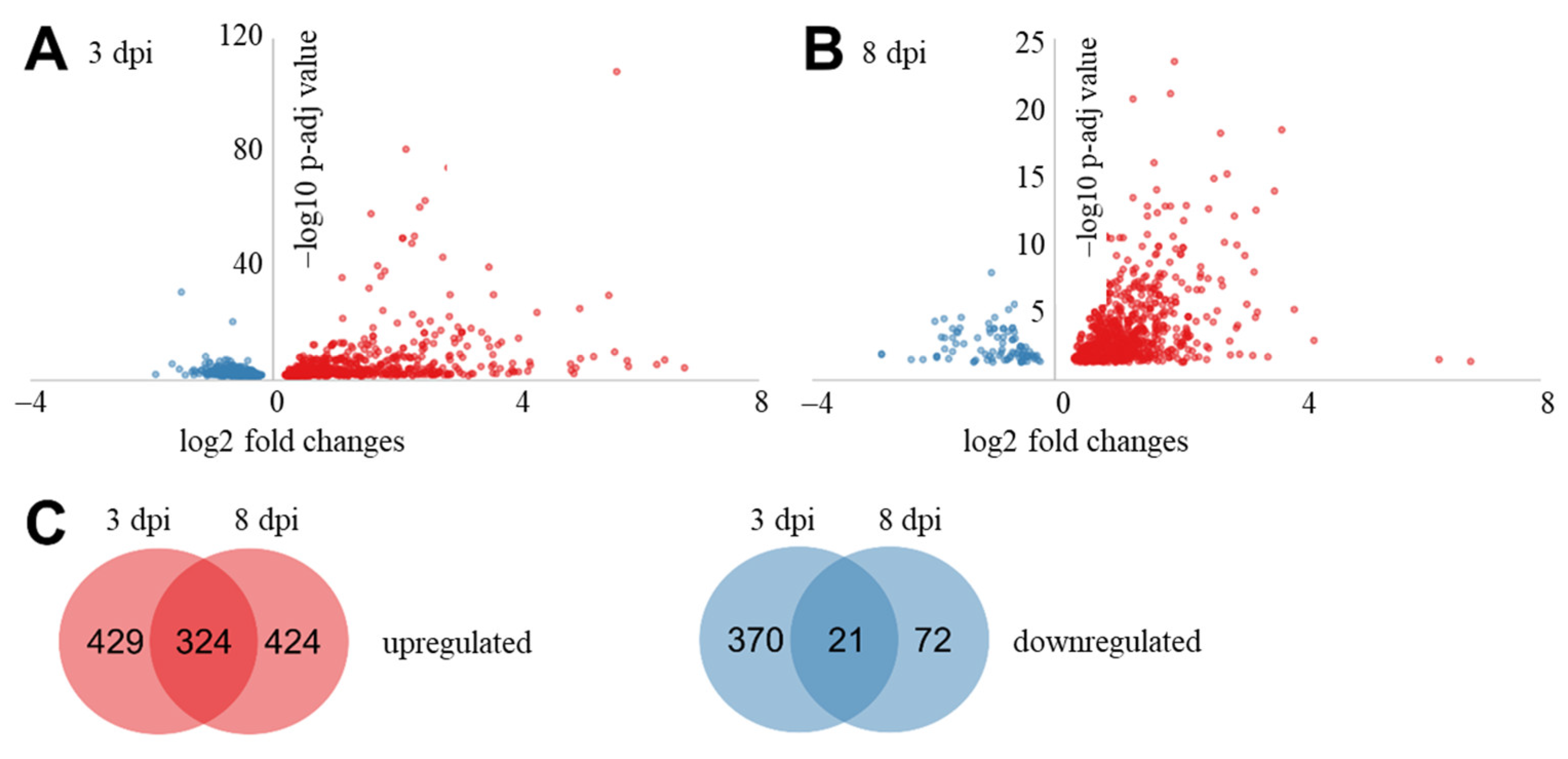
2.3. Differentially Expressed Potato Genes during Interaction with R. solani AG3-PT
2.4. Biological Functions of DEGs
3. Discussion
3.1. Biotic Stress Response Signalling
3.2. Increase of Antifungal Properties
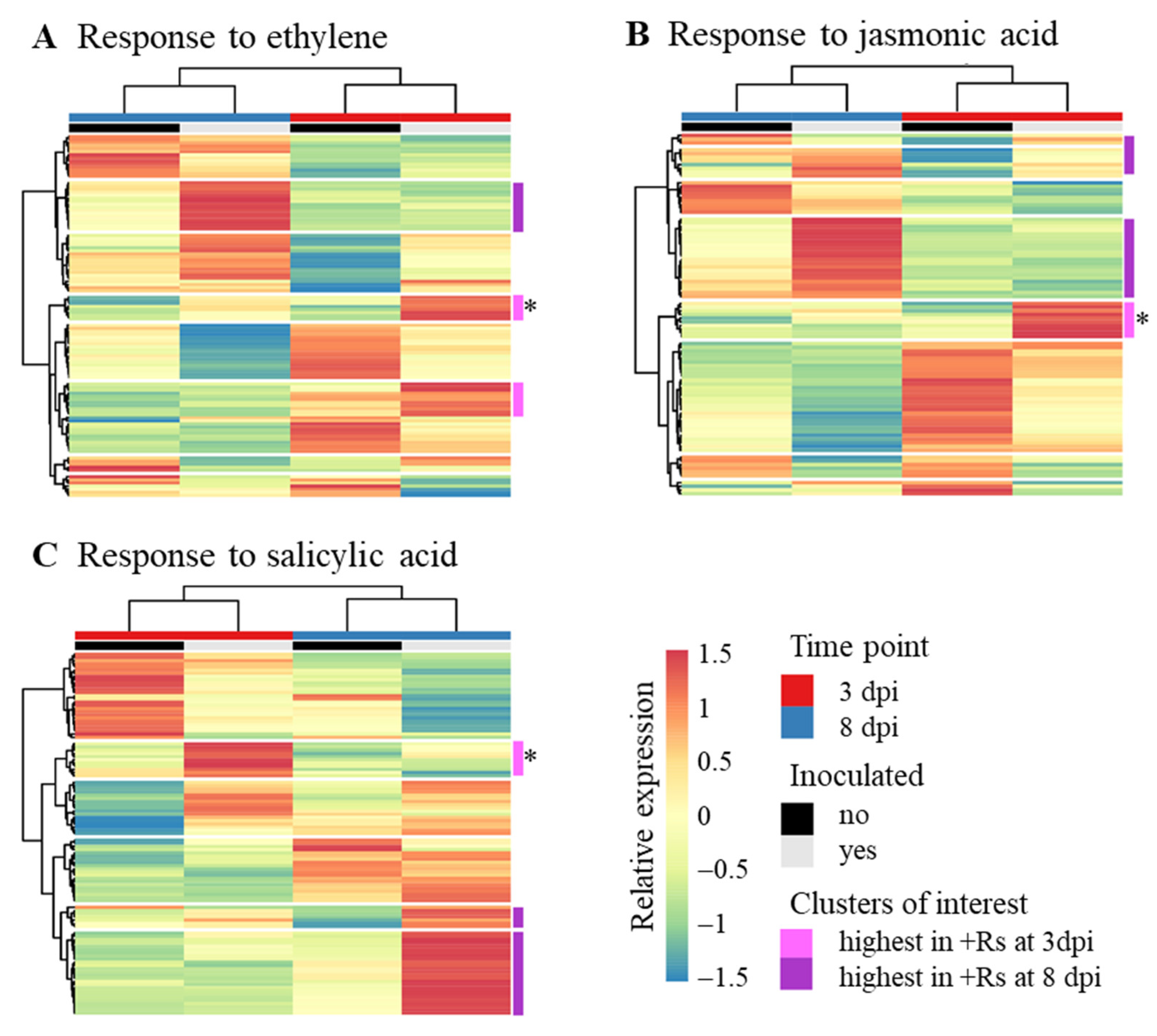
3.3. Responses Associated with Ethylene, Salicylic Acid and Jasmonic Acid
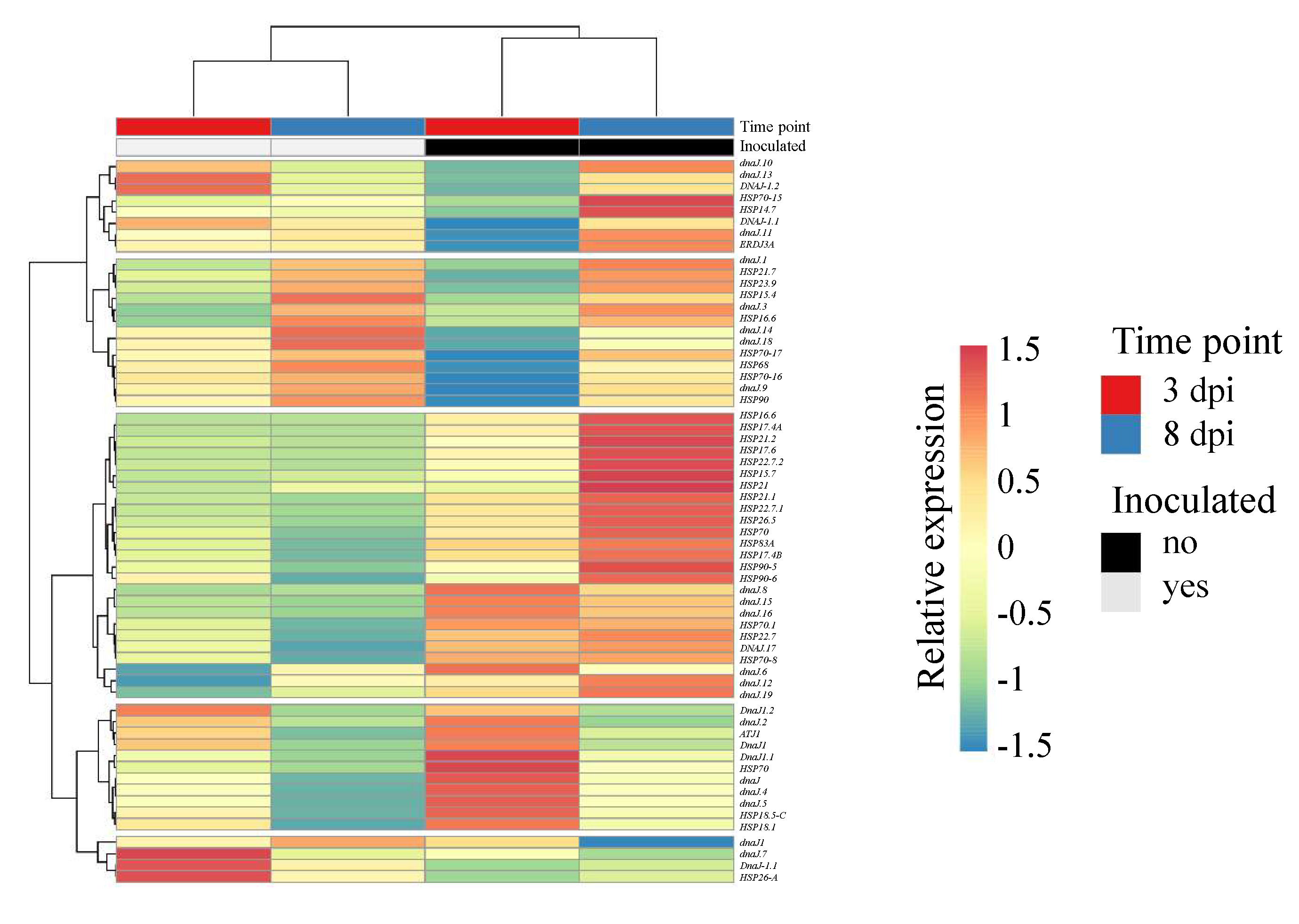
3.4. Involvement of Heat Shock Proteins in the Interaction of Potato with R. solani
4. Materials and Methods
4.1. Plant Treatment and Sampling
4.2. RNA Extraction
4.3. Sequencing of cDNA Libraries
4.4. Mapping and Gene Prediction
4.5. Expression Analysis and Functional Enrichment
4.6. Validating Expression Differences with RT-qPCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Tsror, L. Biology, epidemiology and management of Rhizoctonia solani on potato. J. Phytopathol. 2010, 158, 649–658. [Google Scholar] [CrossRef]
- Woodhall, J.W.; Lees, A.K.; Edwards, S.G.; Jenkinson, P. Characterization of Rhizoctonia solani from potato in Great Britain. Plant Pathol. 2007, 56, 286–295. [Google Scholar] [CrossRef]
- Lehtonen, M.J.; Wilson, P.S.; Ahvenniemi, P. Formation of canker lesions on stems and black scurf on tubers in experimentally inoculated potato plants by isolates of AG2-1, AG3 and AG5 of Rhizoctonia solani: A pilot study and literature review. Agric. Food Sci. 2009, 18, 223–233. [Google Scholar] [CrossRef]
- Djébali, N.; Belhassen, T. Field study of the relative susceptibility of eleven potato (Solanum tuberosum L.) varieties and the efficacy of two fungicides against Rhizoctonia solani attack. Crop Prot. 2010, 29, 998–1002. [Google Scholar] [CrossRef]
- Kushalappa, A.C.; Yogendra, K.N.; Karre, S. Plant innate immune response: Qualitative and quantitative resistance. Crit. Rev. Plant Sci. 2016, 35, 38–55. [Google Scholar] [CrossRef]
- Genzel, F.; Franken, P.; Witzel, K.; Grosch, R. Systemic induction of salicylic acid-related plant defences in potato in response to Rhizoctonia solani AG3PT. Plant Pathol. 2018, 67, 337–348. [Google Scholar] [CrossRef]
- Lehtonen, M.J.; Somervuo, P.; Valkonen, J.P. Infection with Rhizoctonia solani induces defense genes and systemic resistance in potato sprouts grown without light. Phytopathology 2008, 98, 1190–1198. [Google Scholar] [CrossRef]
- Westermann, A.J.; Barquist, L.; Vogel, J. Resolving host-pathogen interactions by dual RNA-seq. PLoS Pathog. 2017, 13, e1006033. [Google Scholar] [CrossRef]
- Zrenner, R.; Genzel, F.; Verwaaijen, B.; Wibberg, D.; Grosch, R. Necrotrophic lifestyle of Rhizoctonia solani AG3-PT during interaction with its host plant potato as revealed by transcriptome analysis. Sci. Rep. 2020, 10, 12574. [Google Scholar] [CrossRef] [PubMed]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinform. 2009, 10, 48. [Google Scholar] [CrossRef]
- Usadel, B.; Poree, F.; Nagel, A.; Lohse, M.; Czedik-Eysenberg, A.; Stitt, M. A guide to using MapMan to visualize and compare Omics data in plants: A case study in the crop species Maize. Plant Cell Environ. 2009, 32, 1211–1229. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.-J. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: Do we understand what they are whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.-L.; Du, L.; Huang, Y.; Gao, S.-M.; Yu, M. Origin and diversification of leucine-rich repeat receptor-like protein kinase (LRR-RLK) genes in plants. BMC Evol. Biol. 2017, 17, 47. [Google Scholar] [CrossRef]
- Dievart, A.; Gottin, C.; Périn, C.; Ranwez, V.; Chantret, N. Origin and diversity of plant receptor-like kinases. Annu. Rev. Plant Biol. 2020, 71, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.D.; Fiers, M.; Ellendorff, U.; Wang, Z.Z.; de Wit, P.; Angenent, G.C.; Thomma, B. The diverse roles of extracellular leucine-rich repeat-containing receptor-like proteins in plants. Crit. Rev. Plant Sci. 2010, 29, 285–299. [Google Scholar] [CrossRef]
- Pan, L.; Lv, S.; Yang, N.; Lv, Y.; Liu, Z.; Wu, J.; Wang, G. The multifunction of CLAVATA2 in plant development and immunity. Front. Plant Sci. 2016, 7, 1573. [Google Scholar] [CrossRef]
- Wan, J.; Zhang, X.-C.; Neece, D.; Ramonell, K.M.; Clough, S.; Kim, S.-Y.; Stacey, M.G.; Stacey, G. A LysM receptor-like kinase plays a critical role in chitin signaling and fungal resistance in Arabidopsis. Plant Cell 2008, 20, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Gan, L.; Shang, K.; Wang, C.; Song, Y.; Liu, H.; Zhou, S.; Zhu, C. Global transcriptome analyses reveal the molecular signatures in the early response of potato (Solanum tuberosum L.) to Phytophthora infestans, Ralstonia solanacearum, and Potato virus Y infection. Planta 2020, 252, 57. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Tanaka, K.; Zhang, X.-C.; Son, G.H.; Brechenmacher, L.; Nguyen, T.H.N.; Stacey, G. LYK4, a lysin motif receptor-like kinase, is important for chitin signaling and plant innate immunity in Arabidopsis. Plant Physiol. 2012, 160, 396–406. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Halane, M.K.; Kim, S.H.; Gassmann, W. Pathogen effectors target Arabidopsis EDS1 and alter its interactions with immune regulators. Science 2011, 334, 1405–1408. [Google Scholar] [CrossRef]
- Ray, S.K.; Macoy, D.M.; Kim, W.-Y.; Lee, S.Y.; Kim, M.G. Role of RIN4 in regulating PAMP-triggered immunity and effector-triggered immunity: Current status and future perspectives. Mol. Cells 2019, 42, 503–511. [Google Scholar] [CrossRef]
- Day, B.; Dahlbeck, D.; Staskawicz, B.J. NDR1 interaction with RIN4 mediates the differential activation of multiple disease resistance pathways in Arabidopsis. Plant Cell 2006, 18, 2782–2791. [Google Scholar] [CrossRef]
- Stanford, A.; Bevan, M.; Northcote, D. Differential expression within a family of novel wound-induced genes in potato. Mol. Gen. Genet. 1989, 215, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Peng Yuan, D.; Xu, X.F.; Hong, W.-J.; Wang, S.T.; Jia, X.T.; Liu, Y.; Li, S.; Li, Z.M.; Sun, Q.; Mei, Q.; et al. Transcriptome analysis of rice leaves in response to Rhizoctonia solani infection and reveals a novel regulatory mechanism. Plant Biotechnol. Rep. 2020, 14, 559–573. [Google Scholar] [CrossRef]
- Zhu, C.; Ai, L.; Wang, L.; Yin, P.; Liu, C.; Li, S.; Zeng, H. De novo transcriptome analysis of Rhizoctonia solani AG1 IA strain early invasion in Zoysia japonica root. Front. Microbiol. 2016, 7, 708. [Google Scholar] [CrossRef]
- Hu, C.-H.; Wang, P.-Q.; Zhang, P.-P.; Nie, X.-M.; Li, B.-B.; Tai, L.; Liu, W.-T.; Li, W.-Q.; Chen, K.-M. NADPH oxidases: The vital performers and center hubs during plant growth and signaling. Cells 2020, 9, 437. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, H.; Sugie, K.; Park, H.-J.; Maeda, H.; Tsuda, N.; Kawakita, K.; Doke, N. Induction of plant gp91 phox homolog by fungal cell wall, arachidonic acid, and salicylic acid in potato. Mol. Plant Microbe Interact. 2001, 14, 725–736. [Google Scholar] [CrossRef]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-transferase enzymes in plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1836. [Google Scholar] [CrossRef] [PubMed]
- Samsatly, J.; Copley, T.R.; Jabaji, S.H. Antioxidant genes of plants and fungal pathogens are distinctly regulated during disease development in different Rhizoctonia solani pathosystems. PLoS ONE 2018, 13, e0192682. [Google Scholar] [CrossRef]
- Perl-Treves, R.; Foley, R.C.; Chen, W.; Singh, K.B. Early induction of the Arabidopsis GSTF8 promoter by specific strains of the fungal pathogen Rhizoctonia solani. Mol. Plant Microbe Interact. 2004, 17, 70–80. [Google Scholar] [CrossRef]
- Goodwin, P.H.; Xie, W.; Valliani, M. Three genes of miraculin-like proteins from Nicotiana benthamiana with dissimilar putative structures show highly similar patterns of induction following bacterial and fungal infections. Eur. J. Plant Pathol. 2012, 134, 795–810. [Google Scholar] [CrossRef]
- Huang, H.; Qi, S.-D.; Qi, F.; Wu, C.-A.; Yang, G.-D.; Zheng, C.-C. NtKTI1, a Kunitz trypsin inhibitor with antifungal activity from Nicotiana tabacum, plays an important role in tobacco’s defense response. FEBS J. 2010, 277, 4076–4088. [Google Scholar] [CrossRef]
- Stumpe, M.; Kandzia, R.; Göbel, C.; Rosahl, S.; Feussner, I. A pathogen-inducible divinyl ether synthase (CYP74D) from elicitor-treated potato suspension cells. FEBS Lett. 2001, 507, 371–376. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.-y.; Guo, W.-z. The cytochrome P450 superfamily: Key players in plant development and defense. J. Integr. Agric. 2015, 14, 1673–1686. [Google Scholar] [CrossRef]
- Pandian, B.A.; Sathishraj, R.; Djanaguiraman, M.; Prasad, P.V.V.; Jugulam, M. Role of cytochrome P450 enzymes in plant stress response. Antioxidants 2020, 9, 454. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, L.; Fu, C.; Wang, L.; Liu, H.; Cheng, Y.; Li, S.; Deng, Q.; Wang, S.; Zhu, J.; et al. Comparative transcriptome analyses of gene expression changes triggered by Rhizoctonia solani AG1 IA infection in resistant and susceptible rice varieties. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Duan, S.; Armstrong, M.R.; Xu, J.; Zheng, J.; Hu, J.; Chen, X.; Hein, I.; Li, G.; Jin, L. Comparative transcriptome profiling reveals compatible and incompatible patterns of potato toward Phytophthora infestans. G3 GenesGenomesGenet. 2020, 10, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Kouzai, Y.; Kimura, M.; Watanabe, M.; Kusunoki, K.; Osaka, D.; Suzuki, T.; Matsui, H.; Yamamoto, M.; Ichinose, Y.; Toyoda, K.; et al. Salicylic acid-dependent immunity contributes to resistance against Rhizoctonia solani, a necrotrophic fungal agent of sheath blight, in rice and Brachypodium distachyon. New Phytol. 2018, 217, 771–783. [Google Scholar] [CrossRef]
- Koornneef, A.; Pieterse, C.M.J. Cross talk in defense signaling. Plant Physiol. 2008, 146, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef]
- Al-Whaibi, M.H. Plant heat-shock proteins: A mini review. J. King Saud Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- Kanzaki, H.; Saitoh, H.; Ito, A.; Fujisawa, S.; Kamoun, S.; Katou, S.; Yoshioka, H.; Terauchi, R. Cytosolic HSP90 and HSP70 are essential components of INF1-mediated hypersensitive response and non-host resistance to Pseudomonas cichorii in Nicotiana benthamiana. Mol. Plant Pathol. 2003, 4, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Shibata, Y.; Kawakita, K.; Takemoto, D. SGT1 and HSP90 are essential for age-related non-host resistance of Nicotiana benthamiana against the oomycete pathogen Phytophthora infestans. Physiol. Mol. Plant Pathol. 2011, 75, 120–128. [Google Scholar] [CrossRef]
- Jelenska, J.; van Hal, J.A.; Greenberg, J.T. Pseudomonas syringae hijacks plant stress chaperone machinery for virulence. Proc. Natl. Acad. Sci. USA 2010, 107, 13177–13182. [Google Scholar] [CrossRef] [PubMed]
- Bricchi, I.; Bertea, C.M.; Occhipinti, A.; Paponov, I.A.; Maffei, M.E. Dynamics of membrane potential variation and gene expression induced by Spodoptera littoralis, Myzus persicae, and Pseudomonas syringae in Arabidopsis. PLoS ONE 2012, 7, e46673. [Google Scholar] [CrossRef]
- Chen, L.; Hamada, S.; Fujiwara, M.; Zhu, T.; Thao, N.P.; Wong, H.L.; Krishna, P.; Ueda, T.; Kaku, H.; Shibuya, N.; et al. The Hop/Sti1-Hsp90 chaperone complex facilitates the maturation and transport of a PAMP receptor in rice innate immunity. Cell Host Microbe 2010, 7, 185–196. [Google Scholar] [CrossRef]
- Yogendra, K.N.; Kumar, A.; Sarkar, K.; Li, Y.; Pushpa, D.; Mosa, K.A.; Duggavathi, R.; Kushalappa, A.C. Transcription factor StWRKY1 regulates phenylpropanoid metabolites conferring late blight resistance in potato. J. Exp Bot. 2015, 66, 7377–7389. [Google Scholar] [CrossRef]
- Verwaaijen, B.; Wibberg, D.; Winkler, A.; Zrenner, R.; Bednarz, H.; Niehaus, K.; Grosch, R.; Pühler, A.; Schlüter, A. A comprehensive analysis of the Lactuca sativa, L. transcriptome during different stages of the compatible interaction with Rhizoctonia solani. Sci. Rep. 2019, 9, 7221. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Bolser, D.; de Boer, J.; Sonderkaer, M.; Amoros, W.; Carboni, M.F.; D’Ambrosio, J.M.; de la Cruz, G.; Di Genova, A.; Douches, D.S.; et al. Construction of reference chromosome-scale pseudomolecules for potato: Integrating the potato genome with genetic and physical maps. G3 (Bethesda) 2013, 3, 2031–2047. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Pan, S.; Cheng, S.; Zhang, B.; Mu, D.; Ni, P.; Zhang, G.; Yang, S.; Li, R.; Wang, J.; et al. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Rupp, O.; Becker, J.; Brinkrolf, K.; Timmermann, C.; Borth, N.; Pühler, A.; Noll, T.; Goesmann, A. Construction of a public CHO cell line transcript database using versatile bioinformatics analysis pipelines. PLoS ONE 2014, 9, e85568. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Simao, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Hilker, R.; Stadermann, K.B.; Schwengers, O.; Anisiforov, E.; Jaenicke, S.; Weisshaar, B.; Zimmermann, T.; Goesmann, A. ReadXplorer 2-detailed read mapping analysis and visualization from one single source. Bioinformatics 2016, 32, 3702–3708. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Wagner, G.P.; Kin, K.; Lynch, V.J. Measurement of mRNA abundance using RNA-seq data: RPKM measure is inconsistent among samples. Theory Biosci. 2012, 131, 281–285. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Boettiger, C. An introduction to Docker for reproducible research. SIGOPS Oper Syst. Rev. 2015, 49, 71–79. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Lohse, M.; Nagel, A.; Herter, T.; May, P.; Schroda, M.; Zrenner, R.; Tohge, T.; Fernie, A.R.; Stitt, M.; Usadel, B. Mercator: A fast and simple web server for genome scale functional annotation of plant sequence data. Plant Cell Environ. 2014, 37, 1250–1258. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zentgraf, U. The Antagonist function of Arabidopsis WRKY53 and ESR/ESP in leaf senescence is modulated by the jasmonic and salicylic acid equilibrium. Plant Cell 2007, 19, 819–830. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Gene Product | −Rs | +Rs | Average |
|---|---|---|---|---|
| TCONS_00022201_m.32854 | Abscisic stress-ripening protein 1 | 6687 | 5280 | 5983 |
| TCONS_00049449_m.67948 | Non-specific lipid-transfer protein 2 | 7349 | 3723 | 5536 |
| TCONS_00046088_m.64962 | Proteinase inhibitor 1 | 3979 | 3740 | 3860 |
| TCONS_00022130_m.32684 | Histidine-rich glycoprotein | 2305 | 3558 | 2931 |
| TCONS_00058960_m.82079 | Catalase isozyme 2 | 2607 | 3138 | 2872 |
| TCONS_00046852_m.60760 | Metallothionein-like protein type 2 B | 3083 | 2618 | 2851 |
| TCONS_00046087_m.64961 | Proteinase inhibitor 1 | 3029 | 2596 | 2813 |
| TCONS_00046864_m.60797 | Heat shock cognate 70 kDa protein 2 | 2546 | 2401 | 2474 |
| TCONS_00002891_m.7661 | Translationally-controlled tumour protein | 2356 | 2144 | 2250 |
| TCONS_00039696_m.54216 | Heat shock cognate protein 80 | 1988 | 1723 | 1855 |
| Loci | Base Mean | Log2 Fold Change | p-Adjusted | Gene Name (Predicted) | Gene Product (Predicted) |
|---|---|---|---|---|---|
| TCONS_00004194_m.10757 | 563 | 6.8 | 1.0 × 10−4 | DES | 9-Divinyl ether synthase |
| TCONS_00041095_m.57761 | 1959 | 6.4 | 1.1 × 10−7 | CYP71D7 | Cytochrome P450 71D7 |
| TCONS_00047931_m.63689 | 42 | 6.3 | 4.9 × 10−6 | ERF098 | Ethylene-responsive transcription factor |
| TCONS_00040978_m.57527 | 1091 | 5.9 | 3.0 × 10−5 | AIM1 | Peroxisomal fatty acid β-oxidation protein |
| TCONS_00008898_m.1264 | 696 | 5.8 | 2.6 × 10−7 | CYP71D55 | Premnaspirodiene oxygenase |
| TCONS_00053685_m.73894 | 2282 | 5.7 | 6.2 × 10−109 | CBP | Citrate-binding protein |
| TCONS_00032279_m.47383 | 97 | 5.6 | 1.9 × 10−10 | PUB24 | E3 ubiquitin-protein ligase PUB24 |
| TCONS_00038986_m.52164 | 437 | 5.5 | 2.9 × 10−30 | GDS | (-)-Germacrene D synthase |
| TCONS_00048229_m.64462 | 371 | 5.3 | 8.8 × 10−9 | HIDM | 2-Hydroxyisoflavanone dehydratase |
| TCONS_00021788_m.31824 | 2715 | 5.1 | 4.3 × 10−8 | CYP71D7 | Cytochrome P450 71D7 |
| TCONS_00038015_m.49462 | 3156 | −1.2 | 8.7 × 10−3 | At4g36180 | Probable LRR receptor-like ser/thr-PK |
| TCONS_00010359_m.16650 | 723 | −1.3 | 2.4 × 10−3 | STR15 | Rhodanese-like domain protein 15; chloro |
| TCONS_00047559_m.62592 | 403 | −1.3 | 5.0 × 10−4 | HSP17.8 | 17.8 kDa class I heat shock protein |
| TCONS_00058679_m.81355 | 148 | −1.3 | 3.0 × 10−2 | DMG400005010 | Uncharacterized protein |
| TCONS_00048318_m.64746 | 19,682 | −1.3 | 1.9 × 10−3 | ELIP1 | Early light-induced protein; chloroplastic |
| TCONS_00037799_m.48946 | 221 | −1.5 | 2.7 × 10−2 | NCS1 | S-norcoclaurine synthase 1 |
| TCONS_00008336_m.11190 | 2494 | −1.5 | 2.3 × 10−31 | SN2 | Snakin-2 |
| TCONS_00019937_m.26919 | 658 | −1.5 | 1.9 × 10−4 | DMG400014234 | Uncharacterized protein |
| TCONS_00002388_m.6328 | 1515 | −1.7 | 2.7 × 10−6 | WSD1 | O-acyltransferase WSD1 |
| TCONS_00035423_m.48440 | 244 | −1.9 | 1.6 × 10−2 | At5g33370 | GDSL esterase/lipase |
| Loci | Base Mean | Log2 Fold Change | p-Adjusted | Gene Name (Predicted) | Gene Product (Predicted) |
|---|---|---|---|---|---|
| TCONS_00021878_m.32075 | 22 | 6.8 | 4. × 10−2 | TPS31 | Viridiflorene synthase |
| TCONS_00016163_m.24776 | 16 | 6.3 | 3.3 × 10−2 | MCL | Miraculin |
| TCONS_00047931_m.63689 | 22 | 4.3 | 1.3 × 10−3 | ERF098 | Ethylene-responsive transcription factor |
| TCONS_00004194_m.10757 | 126 | 3.9 | 6.8 × 10−6 | DES | 9-Divinyl ether synthase |
| TCONS_00053685_m.73894 | 735 | 3.7 | 5.1 × 10−19 | CBP | Citrate-binding protein |
| TCONS_00009061_m.13072 | 1210 | 3.6 | 1.5 × 10−14 | PLP1 | Patatin-like protein 1 |
| TCONS_00054756_m.76933 | 52 | 3.5 | 2.0 × 10−2 | Kwl1 | Kiwellin |
| TCONS_00050984_m.71786 | 119 | 3.3 | 3.6 × 10−13 | CYP98A2 | Cytochrome P450 98A2 |
| TCONS_00018583_m.23294 | 490 | 3.3 | 2.7 × 10−5 | DREB1A | Dehydration-response element-binding 1A |
| TCONS_00041095_m.57761 | 207 | 3.3 | 1.2 × 10−8 | CYP71D7 | Cytochrome P450 71D7 |
| TCONS_00050284_m.70004 | 31 | 3.3 | 1.7 × 10−2 | UGT73D1 | UDP-glycosyltransferase 73D1 |
| TCONS_00059966_m.79472 | 425 | −1.8 | 3.1 × 10−5 | BBX22 | B-box zinc finger protein 22 |
| TCONS_00019878_m.26786 | 9475 | −1.9 | 7.6 × 10−8 | HSP70 | Heat shock cognate 70 kDa protein |
| TCONS_00025911_m.35167 | 84 | −1.9 | 2.2 × 10−2 | PER64 | Peroxidase 64 |
| TCONS_00047789_m.63274 | 364 | −1.9 | 1.8 × 10−2 | At4g02900 | CSC1-like protein |
| TCONS_00047789_m.63275 | 364 | −1.9 | 1.8 × 10−2 | - | Hypothetical protein |
| TCONS_00048318_m.64746 | 15,884 | −2.0 | 5.0 × 10−5 | ELIP1 | Early light-induced protein; chloroplastic |
| TCONS_00059918_m.79395 | 402 | −2.2 | 3.3 × 10−2 | PER27 | Peroxidase 27 |
| TCONS_00053904_m.74476 | 6317 | −2.4 | 3.2 × 10−2 | HSP22.7 | 22.7 kDa class IV heat shock protein |
| TCONS_00059319_m.82977 | 56 | −2.9 | 1.3 × 10−2 | EXT2 | Extensin-2 |
| Up/Down | GO Term | GO Term Name | FDR 1 q-Value | Enrichment |
|---|---|---|---|---|
| + | GO:0006952 | defence response | 1.8 × 10−14 | 1.9 |
| + | GO:0050896 | response to stimulus | 3.9 × 10−13 | 1.5 |
| + | GO:0006468 | protein phosphorylation | 9.5 × 10−11 | 1.6 |
| + | GO:0043207 | response to external biotic stimulus | 2.0 × 10−10 | 1.8 |
| + | GO:0009607 | response to biotic stimulus | 1.6 × 10−10 | 1.8 |
| + | GO:0016310 | phosphorylation | 1.4 × 10−10 | 1.5 |
| + | GO:0051707 | response to other organisms | 1.8 × 10−10 | 2.1 |
| + | GO:0051704 | multi-organism process | 3.2 × 10−10 | 1.6 |
| + | GO:0010200 | response to chitin | 3.9 × 10−10 | 11.3 |
| + | GO:0006950 | response to stress | 2.9 × 10−9 | 1.5 |
| − | GO:1901700 | response to oxygen-containing compound | 4.2 × 10−8 | 2.3 |
| − | GO:0022900 | electron transport chain | 3.9 × 10−8 | 3.3 |
| − | GO:0009699 | phenylpropanoid biosynthetic process | 1.4 × 10−8 | 5.1 |
| − | GO:0009698 | phenylpropanoid metabolic process | 6.7 × 10−9 | 4.8 |
| − | GO:0009314 | response to radiation | 5.9 × 10−9 | 1.7 |
| − | GO:0009628 | response to abiotic stimulus | 3.8 × 10−9 | 1.4 |
| − | GO:0009416 | response to light stimulus | 3.1 × 10−10 | 1.8 |
| − | GO:0055114 | oxidation-reduction process | 2.3 × 10−10 | 1.8 |
| − | GO:0006091 | generation of precursor metabolites and energy | 1.6 × 10−10 | 3.0 |
| − | GO:0009767 | photosynthetic electron transport chain | 1.9 × 10−12 | 5.1 |
| Up/Down | GO Term | GO Term Name | FDR 1 q-Value | Enrichment |
|---|---|---|---|---|
| + | GO:0006952 | defence response | 5.3 × 10−40 | 3.0 |
| + | GO:0050896 | response to stimulus | 2.6 × 10−23 | 1.6 |
| + | GO:0010200 | response to chitin | 3.6 × 10−23 | 10.6 |
| + | GO:0006950 | response to stress | 2.0 × 10−22 | 1.8 |
| + | GO:0051707 | response to other organisms | 1.4 × 10−21 | 2.4 |
| + | GO:0009607 | response to biotic stimulus | 4.2 × 10−21 | 2.4 |
| + | GO:0043207 | response to external biotic stimulus | 4.9 × 10−21 | 2.4 |
| + | GO:0051704 | multi-organism process | 2.2 × 10−20 | 1.9 |
| + | GO:0006468 | protein phosphorylation | 1.3 × 10−15 | 2.0 |
| + | GO:0042493 | response to drug | 2.3 × 10−15 | 4.4 |
| − | GO:0055114 | oxidation-reduction process | 1.7 × 10−7 | 1.9 |
| − | GO:0006979 | response to oxidative stress | 1.9 × 10−8 | 9.1 |
| − | GO:0042542 | response to hydrogen peroxide | 1.6 × 10−8 | 28.4 |
| − | GO:0009266 | response to temperature stimulus | 9.6 × 10−10 | 7.9 |
| − | GO:0043603 | cellular amide metabolic process | 5.2 × 10−10 | 1.7 |
| − | GO:0009408 | response to heat | 3.4 × 10−12 | 16.1 |
| − | GO:0006518 | peptide metabolic process | 1.5 × 10−12 | 2.0 |
| − | GO:0043604 | amide biosynthetic process | 4.7 × 10−13 | 2.0 |
| − | GO:0043043 | peptide biosynthetic process | 1.2 × 10−15 | 2.2 |
| − | GO:0006412 | translation | 4.2 × 10−16 | 2.2 |
| Bin 1 | MapMan Bin Name | Elements 2 | p-Value 3 |
|---|---|---|---|
| 20.1.7 | Stress.biotic.PR-proteins | 365 | 0.0 |
| 30 | Signalling | 1222 | 0.0 |
| 20 | Stress | 894 | 0.0 |
| 20.1 | Stress.biotic | 543 | 0.0 |
| 30.2 | Signalling.receptor kinases | 534 | 0.0 |
| 1 | PS | 170 | 7.4 × 10−54 |
| 1.1 | PS.lightreaction | 113 | 1.2 × 10−43 |
| 1.1.1 | PS.lightreaction.photosystem II | 39 | 5.6 × 10−16 |
| 1.1.1.2 | PS.lightreaction.photosystem II.PSII polypeptide subunit | 30 | 2.8 × 10−13 |
| 29.2.1.1.1 | Protein.synthesis.ribosomal protein.prokaryotic.chloro | 51 | 4.4 × 10−11 |
| 29.2.1.1 | Protein.synthesis.ribosomal protein.prokaryotic | 99 | 3.0 × 10−10 |
| 30.2.11 | Signalling.receptor kinases.leucine rich repeat XI | 63 | 5.2 × 10−10 |
| 1.1.2 | PS.lightreaction.photosystem I | 22 | 3.1 × 10−9 |
| 29.2.1 | Protein.synthesis.ribosomal protein | 216 | 4.1 × 10−9 |
| 30.2.24 | Signalling.receptor kinases.S-locus glycoprotein like | 56 | 2.7 × 10−8 |
| 1.1.2.2 | PS.lightreaction.photosystem I.PSI polypeptide subunits | 15 | 2.0 × 10−7 |
| 30.2.20 | Signalling.receptor kinases.wheat LRK10 like | 25 | 3.0 × 10−7 |
| 26.9 | Misc.glutathione S transferases | 50 | 3.0 × 10−7 |
| 29.2 | Protein.synthesis | 412 | 3.0 × 10−7 |
| 26.2 | Misc.UDP glucosyl and glucoronyl transferases | 204 | 3.0 × 10−7 |
| Bin 1 | MapMan Bin Name | Elements 2 | p-Value 3 |
|---|---|---|---|
| 20.1.7 | Stress.biotic.PR-proteins | 366 | 0.0 |
| 30 | Signalling | 1222 | 0.0 |
| 20 | Stress | 895 | 0.0 |
| 20.1 | Stress.biotic | 544 | 0.0 |
| 30.2 | Signalling.receptor kinases | 534 | 0.0 |
| 29.2.1 | Protein.synthesis.ribosomal protein | 216 | 2.7 × 10−30 |
| 29.2 | Protein.synthesis | 412 | 1.2 × 10−29 |
| 1 | PS | 170 | 1.4 × 10−22 |
| 1.1 | PS.lightreaction | 113 | 6.5 × 10−18 |
| 35 | Not assigned | 11290 | 1.4 × 10−17 |
| 29.2.1.1 | Protein.synthesis.ribosomal protein.prokaryotic | 99 | 8.0 × 10−16 |
| 29.2.1.2 | Protein.synthesis.ribosomal protein.eukaryotic | 106 | 3.3 × 10−14 |
| 27.3.32 | RNA.regulation transcription.WRKY transcription factor | 62 | 1.6 × 10−13 |
| 35.1.5 | Not assigned. pentatricopeptide (PPR) repeat-containing | 422 | 1.6 × 10−13 |
| 30.2.24 | Signalling.receptor kinases.S-locus glycoprotein like | 56 | 2.6 × 10−13 |
| 20.1.2 | Stress.biotic.receptors | 33 | 1.3 × 10−11 |
| 30.2.20 | Signalling.receptor kinases.wheat LRK10 like | 25 | 5.4 × 10−10 |
| 30.2.8 | Signalling.receptor kinases.leucine rich repeat VIII | 36 | 5.1 × 10−9 |
| 29.4 | Protein.postranslational modification | 607 | 5.3 × 10−9 |
| 29.2.1.1.1 | Protein.synthesis.ribosomal protein.prokaryotic.chloro | 51 | 8.7 × 10−9 |
| SeqName | ∆Cq (a) 1 | ∆Cq (b) 1 | ∆∆Cq 2 | BaseMean 3 | Log2fold Change 3 | Comparison (a):(b) 4 |
|---|---|---|---|---|---|---|
| TCONS_00019878_m.26787; HSP70 | 6.9 ± 0.12 | 7.8 ± 0.56 | −1.86 * | 10605 | −0.6 | (3 dpi):(−Rs) |
| TCONS_00019878_m.26787; HSP70 | 6.3 ± 0.80 | 7.1 ± 2.11 | −0.76 | 9475 | −1.9 | (8 dpi):(−Rs) |
| TCONS_00004194_m.10757; DES | 4.0 ± 2.23 | −0.8 ± 0.83 | 4.01 | 563 | 6.8 | (3 dpi):(−Rs) |
| TCONS_00004194_m.10757; DES | 1.4 ± 0.29 | −3.5 ± 2.09 | 4.87 * | 126 | 3.9 | (8 dpi):(−Rs) |
| TCONS_00008898_m.12640; CYP71D55 | 2.4 ± 2.00 | n.d. | - | 1959 | 5.8 | (3 dpi):(−Rs) |
| TCONS_00008898_m.12640; CYP71D55 | 0.02 ± 0.4 | n.d. | - | 120 | 3.0 | (8 dpi):(−Rs) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zrenner, R.; Verwaaijen, B.; Genzel, F.; Flemer, B.; Grosch, R. Transcriptional Changes in Potato Sprouts upon Interaction with Rhizoctonia solani Indicate Pathogen-Induced Interference in the Defence Pathways of Potato. Int. J. Mol. Sci. 2021, 22, 3094. https://doi.org/10.3390/ijms22063094
Zrenner R, Verwaaijen B, Genzel F, Flemer B, Grosch R. Transcriptional Changes in Potato Sprouts upon Interaction with Rhizoctonia solani Indicate Pathogen-Induced Interference in the Defence Pathways of Potato. International Journal of Molecular Sciences. 2021; 22(6):3094. https://doi.org/10.3390/ijms22063094
Chicago/Turabian StyleZrenner, Rita, Bart Verwaaijen, Franziska Genzel, Burkhardt Flemer, and Rita Grosch. 2021. "Transcriptional Changes in Potato Sprouts upon Interaction with Rhizoctonia solani Indicate Pathogen-Induced Interference in the Defence Pathways of Potato" International Journal of Molecular Sciences 22, no. 6: 3094. https://doi.org/10.3390/ijms22063094
APA StyleZrenner, R., Verwaaijen, B., Genzel, F., Flemer, B., & Grosch, R. (2021). Transcriptional Changes in Potato Sprouts upon Interaction with Rhizoctonia solani Indicate Pathogen-Induced Interference in the Defence Pathways of Potato. International Journal of Molecular Sciences, 22(6), 3094. https://doi.org/10.3390/ijms22063094