
Intracellular and Tissue Levels of Vitamin B12 in Hepatocytes Are Modulated by CD320 Receptor and TCN2 Transporter

 ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Intracellular and Tissue Levels of B12
2.1.1. Intracellular B12 Level in Hep G2 Cell Line
2.1.2. Intracellular B12 Level in Mice Liver Tissues
2.2. B12 Transcobalamin Receptor (CD320)
2.2.1. Hep G2 Cells
2.2.2. Mice Liver
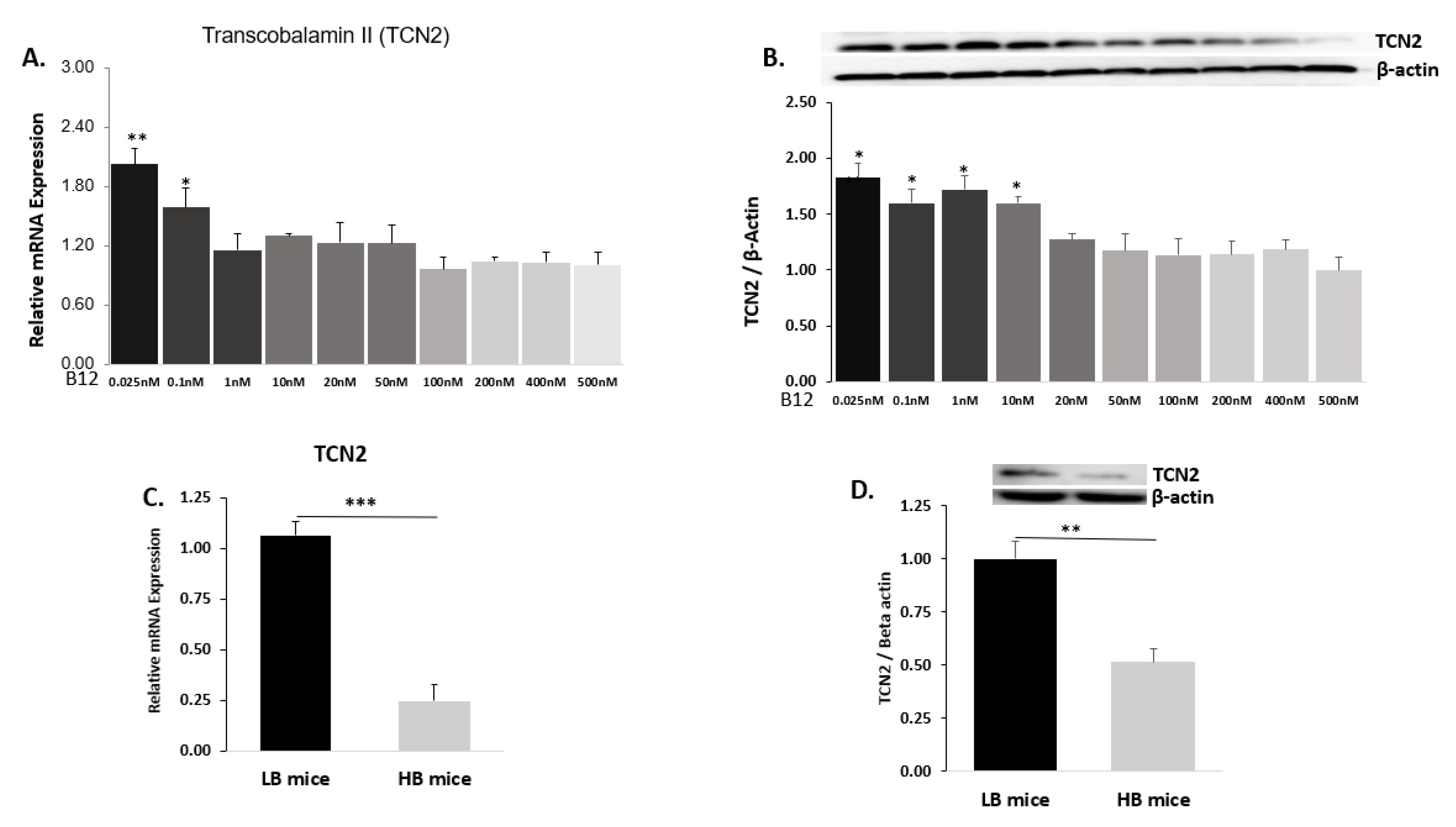
2.3. B12 Transporter Transcobalamin II (TCN2)
2.3.1. Hep G2 Cells
2.3.2. Mice Liver
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Animal Model
4.3. RNA Isolation and Quantitative Real Time-PCR (qRT-PCR)
4.4. Western Blot Analysis
4.5. Measurement of B12
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Smith, A.D.; Warren, M.J.; Refsum, H. Vitamin B12. Adv. Food Nutr. Res. 2018, 83, 215–279. [Google Scholar] [CrossRef] [PubMed]
- Andrès, E.; Fothergill, H.; Mecili, M. Efficacy of oral cobalamin (vitamin B12) therapy. Expert Opin. Pharmacother. 2009, 11, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.J.; Rasmussen, M.R.; Andersen, C.B.; Nexø, E.; Moestrup, S.K. Vitamin B 12 transport from food to the body’s cells—A sophisticated, multistep pathway. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Kozyraki, R.; Fyfe, J.; Kristiansen, M.; Gerdes, C.; Jacobsen, C.; Cui, S.; Christensen, E.I.; Aminoff, M.; De La Chapelle, A.; Krahe, R.; et al. The intrinsic factor–vitamin B12 receptor, cubilin, is a high-affinity apolipoprotein A-I receptor facilitating endocytosis of high-density lipoprotein. Nat. Med. 1999, 5, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Layden, A.J.; O’Brien, K.O.; Pressman, E.K.; Cooper, E.M.; Kent, T.R.; Finkelstein, J.L. Vitamin B12 and placental expression of transcobalamin in pregnant adolescents. Placenta 2016, 45, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Sequeira, J.M.; Nakayama, Y.; Lai, S.-C.; Quadros, E.V. Characterization of the promoter region of TCblR/CD320 gene, the receptor for cellular uptake of transcobalamin-bound cobalamin. Gene 2010, 466, 49–55. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Park, H.S.; Kim, J.O.; An, H.J.; Ryu, C.S.; Ko, E.J.; Kim, Y.R.; Ahn, E.H.; Lee, W.S.; Kim, J.H.; Kim, N.K. Genetic polymorphisms of the cobalamin transport system are associated with idiopathic recurrent implantation failure. J. Assist. Reprod. Genet. 2019, 36, 1513–1522. [Google Scholar] [CrossRef]
- Bernard, D.J.; Pangilinan, F.J.; Cheng, J.; Molloy, A.M.; Brody, L.C. Mice lacking the transcobalamin-vitamin B12 receptor, CD320, suffer from anemia and reproductive deficits when fed vitamin B12-deficient diet. Hum. Mol. Genet. 2018, 27, 3627–3640. [Google Scholar] [CrossRef]
- Greibe, E.; Nymark, O.; Fedosov, S.N.; Heegaard, C.W.; Nexo, E. Differences in Tissue Distribution of Cyano–B12 and Hydroxo–B12 One Week after Oral Intake: An Experimental Study in Male Wistar Rats. Nutrients 2018, 10, 1487. [Google Scholar] [CrossRef] [PubMed]
- Greibe, E.; Kornerup, L.S.; Juul, C.B.; Fedosov, S.N.; Heegaard, C.W.; Nexo, E. The tissue profile of metabolically active coenzyme forms of vitamin B12 differs in vitamin B12-depleted rats treated with hydroxo-B12 or cyano-B12. Br. J. Nutr. 2018, 120, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Greibe, E.; Nymark, O.; Fedosov, S.N.; Heegaard, C.W.; Nexo, E. Dietary Intake of Vitamin B12 is Better for Restoring a Low B12 Status Than a Daily High-Dose Vitamin Pill: An Experimental Study in Rats. Nutrients 2018, 10, 1096. [Google Scholar] [CrossRef]
- Greibe, E.; Nymark, O.; Fedosov, S.; Ostenfeld, M.S.; Heegaard, C.W.; Nexo, E. Absorption of Vitamin B12 Administered in Buffalo Milk: An Experimental Study in Rats. SciTz Nutr. Food Sci. 2017, 2, 1006. [Google Scholar]
- Kornerup, L.S.; Fedosov, S.N.; Juul, C.B.; Greibe, E.; Heegaard, C.W.; Nexo, E. Tissue distribution of oral vitamin B12 is influenced by B12 status and B12 form: An experimental study in rats. Eur. J. Nutr. 2018, 57, 1459–1469. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, L.; Juul, C.; Fedosov, S.; Heegaard, C.; Greibe, E.; Nexo, E. Absorption and retention of free and milk protein-bound cyano- and hydroxocobalamins. An experimental study in rats. Biochimie 2016, 126, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Thagaard, M.S.N.E.; Greibe, E. Vitamin B12 Levels in Rat Organs—No Change Following Prolonged Treatment with Metformin. Int. J. Diabetes Clin. Res. 2015, 2, 1–4. [Google Scholar]
- Nymark, O.; Nexo, E.; Greibe, E. Nutritional 1C Imbalance, B12 Tissue Accumulation, and Pregnancy Outcomes: An Experimental Study in Rats. Nutrients 2018, 10, 1579. [Google Scholar] [CrossRef] [PubMed]
- Golding, P.H. Holotranscobalamin (HoloTC, Active-B12) and Herbert’s model for the development of vitamin B12 deficiency: A review and alternative hypothesis. SpringerPlus 2016, 5, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Alhusaini, S.; McGee, K.; Schisano, B.; Harte, A.; McTernan, P.; Kumar, S.; Tripathi, G. Lipopolysaccharide, high glucose and saturated fatty acids induce endoplasmic reticulum stress in cultured primary human adipocytes: Salicylate alleviates this stress. Biochem. Biophys. Res. Commun. 2010, 397, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Schisano, B.; Tripathi, G.; McGee, K.; McTernan, P.G.; Ceriello, A. Glucose oscillations, more than constant high glucose, induce p53 activation and a metabolic memory in human endothelial cells. Diabetologia 2011, 54, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Andres, E.; Serraj, K.; Zhu, J.; Vermorken, A.J.M. The pathophysiology of elevated vitamin B12 in clinical practice. QJM Int. J. Med. 2013, 106, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Brito, A.; Habeych, E.; Silva-Zolezzi, I.; Galaffu, N.; Allen, L.H. Methods to assess vitamin B12 bioavailability and technologies to enhance its absorption. Nutr. Rev. 2018, 76, 778–792. [Google Scholar] [CrossRef] [PubMed]
- Yajnik, C.S.; Behere, R.V.; Bhat, D.S.; Memane, N.; Raut, D.; Ladkat, R.; Yajnik, P.C.; Kumaran, K.; Fall, C.H.D. A physiological dose of oral vitamin B-12 improves hematological, biochemical-metabolic indices and peripheral nerve function in B-12 deficient Indian adolescent women. PLoS ONE 2019, 14, e0223000. [Google Scholar] [CrossRef] [PubMed]
- Lweno, O.N.; Sudfeld, C.R.; Hertzmark, E.; Manji, K.P.; Aboud, S.; Noor, R.A.; Masanja, H.; Salim, N.; Shahab-Ferdows, S.; Allen, L.H.; et al. Vitamin B12 is Low in Milk of Early Postpartum Women in Urban Tanzania, and was not Significantly Increased by High dose Supplementation. Nutrients 2020, 12, 963. [Google Scholar] [CrossRef]
- Kendrick, N. A Gene’s mRNA Level does not Usually Predict Its Protein Level; Kendricklabscom Inc.: Madison, WI, USA, 2014; pp. 1–8. [Google Scholar]
- Hall, C.A.; Colligan, P.D.; Begley, J.A. Cyclic activity of the receptors of cobalamin bound to transcobalamin II. J. Cell. Physiol. 1987, 133, 187–191. [Google Scholar] [CrossRef]
- Herbert, V. Staging vitamin B-12 (cobalamin) status in vegetarians. Am. J. Clin. Nutr. 1994, 59, 1213S–1222S. [Google Scholar] [CrossRef] [PubMed]
- Zittoun, J.J.H. Cobalamins: An update of fundamental and clinical data. Hématologie 1996, 2, 119–129. [Google Scholar]
- Ermens, A.; Vlasveld, L.; Lindemans, J. Significance of elevated cobalamin (vitamin B12) levels in blood. Clin. Biochem. 2003, 36, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Cooksley, W.G.E.; England, J.M.; Louis, L.; Down, M.C.; Tavill, A.S. Hepatic Vitamin B12 Release and Transcobalamin II Synthesis in the Rat. Clin. Sci. Mol. Med. 1974, 47, 531–545. [Google Scholar] [CrossRef] [PubMed]
- England, J.M.; Clarke, H.G.M.; Down, M.C.; Chanarin, I. Studies on the Transcobalamins. Br. J. Haematol. 1973, 25, 737–749. [Google Scholar] [CrossRef]
- Refsum, H.; Nurk, E.; Smith, A.D.; Ueland, P.M.; Gjesdal, C.G.; Bjelland, I.; Tverdal, A.; Tell, G.S.; Nygaård, O.; Vollset, S.E. The Hordaland Homocysteine Study: A Community-Based Study of Homocysteine, Its Determinants, and Associations with Disease. J. Nutr. 2006, 136, 1731S–1740S. [Google Scholar] [CrossRef]
- Baik, H.; Russell, R. Vitamin B12deficiency in the Elderly. Annu. Rev. Nutr. 1999, 19, 357–377. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, A.; Sapehia, D.; Thakur, S.; Mohanraj, P.S.; Bagga, R.; Kaur, J. Effect of imbalance in folate and vitamin B12 in maternal/parental diet on global methylation and regulatory miRNAs. Sci. Rep. 2019, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Molloy, A.M.; Kirke, P.N.; Brody, L.C.; Scott, J.M.; Mills, J.L. Effects of Folate and Vitamin B12Deficiencies During Pregnancy on Fetal, Infant, and Child Development. Food Nutr. Bull. 2008, 29, S101–S111. [Google Scholar] [CrossRef] [PubMed]
- Abuyaman, O.; Andreasen, B.H.; Kronborg, C.; Vittinghus, E.; Nexo, E. The Soluble Receptor for Vitamin B12 Uptake (sCD320) Increases during Pregnancy and Occurs in Higher Concentration in Urine than in Serum. PLoS ONE 2013, 8, e73110. [Google Scholar] [CrossRef] [PubMed]
- Wurie, H.R.; Buckett, L.; Zammit, V.A. Diacylglycerol acyltransferase 2 acts upstream of diacylglycerol acyltransferase 1 and utilizes nascent diglycerides and de novo synthesized fatty acids in HepG2 cells. FEBS J. 2012, 279, 3033–3047. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boachie, J.; Adaikalakoteswari, A.; Goljan, I.; Samavat, J.; Cagampang, F.R.; Saravanan, P. Intracellular and Tissue Levels of Vitamin B12 in Hepatocytes Are Modulated by CD320 Receptor and TCN2 Transporter. Int. J. Mol. Sci. 2021, 22, 3089. https://doi.org/10.3390/ijms22063089
Boachie J, Adaikalakoteswari A, Goljan I, Samavat J, Cagampang FR, Saravanan P. Intracellular and Tissue Levels of Vitamin B12 in Hepatocytes Are Modulated by CD320 Receptor and TCN2 Transporter. International Journal of Molecular Sciences. 2021; 22(6):3089. https://doi.org/10.3390/ijms22063089
Chicago/Turabian StyleBoachie, Joseph, Antonysunil Adaikalakoteswari, Ilona Goljan, Jinous Samavat, Felino R. Cagampang, and Ponnusamy Saravanan. 2021. "Intracellular and Tissue Levels of Vitamin B12 in Hepatocytes Are Modulated by CD320 Receptor and TCN2 Transporter" International Journal of Molecular Sciences 22, no. 6: 3089. https://doi.org/10.3390/ijms22063089
APA StyleBoachie, J., Adaikalakoteswari, A., Goljan, I., Samavat, J., Cagampang, F. R., & Saravanan, P. (2021). Intracellular and Tissue Levels of Vitamin B12 in Hepatocytes Are Modulated by CD320 Receptor and TCN2 Transporter. International Journal of Molecular Sciences, 22(6), 3089. https://doi.org/10.3390/ijms22063089