1. Introduction
Iron is a vital nutrient involved in a wide range of enzymatic functions and biological processes and is essential for bacterial growth and virulence. The paradox of this key nutrient is its low bioavailability. Fe
3+ is poorly soluble at neutral pH, with a limit of Fe
3+ concentration in aqueous solutions of 10
−18 M. Moreover, in mammals, iron is sequestered by proteins or heme to avoid Fe
2+ toxicity (Fenton reaction) and maintain Fe
3+ in a soluble form. Many bacteria overcome the problem of iron accessibility by producing siderophores, small organic compounds with an extremely high affinity for ferric iron [
1].
Enterobactin (ENT), a triscatechol derivative of a cyclic triserine lactone, is an archetype in the field of siderophores. The chemistry, biology, and iron uptake by this siderophore have been extensively investigated over the last several decades, especially in
E. coli [
2,
3]. ENT is produced by
Enterobacteriaceae such as
Escherichia coli,
Klebsiella pneumonia,
Shigella flexneri, and
Salmonella typhimurium [
4,
5,
6,
7] and was thought to be unique to Gram-negative bacteria. However, the isolation of ENT from Gram-positive Streptomyces species [
8] strongly suggests that the production of this siderophore may be wider than previously thought. ENT is synthesized in the bacterial cytoplasm by non-ribosomal peptide synthetases from chorismic acid [
2].
ENT scavenges ferric iron in the bacterial environment with a Ka of 10
52 M
−1 [
9]. Afterwards, ferric-ENT recapture by
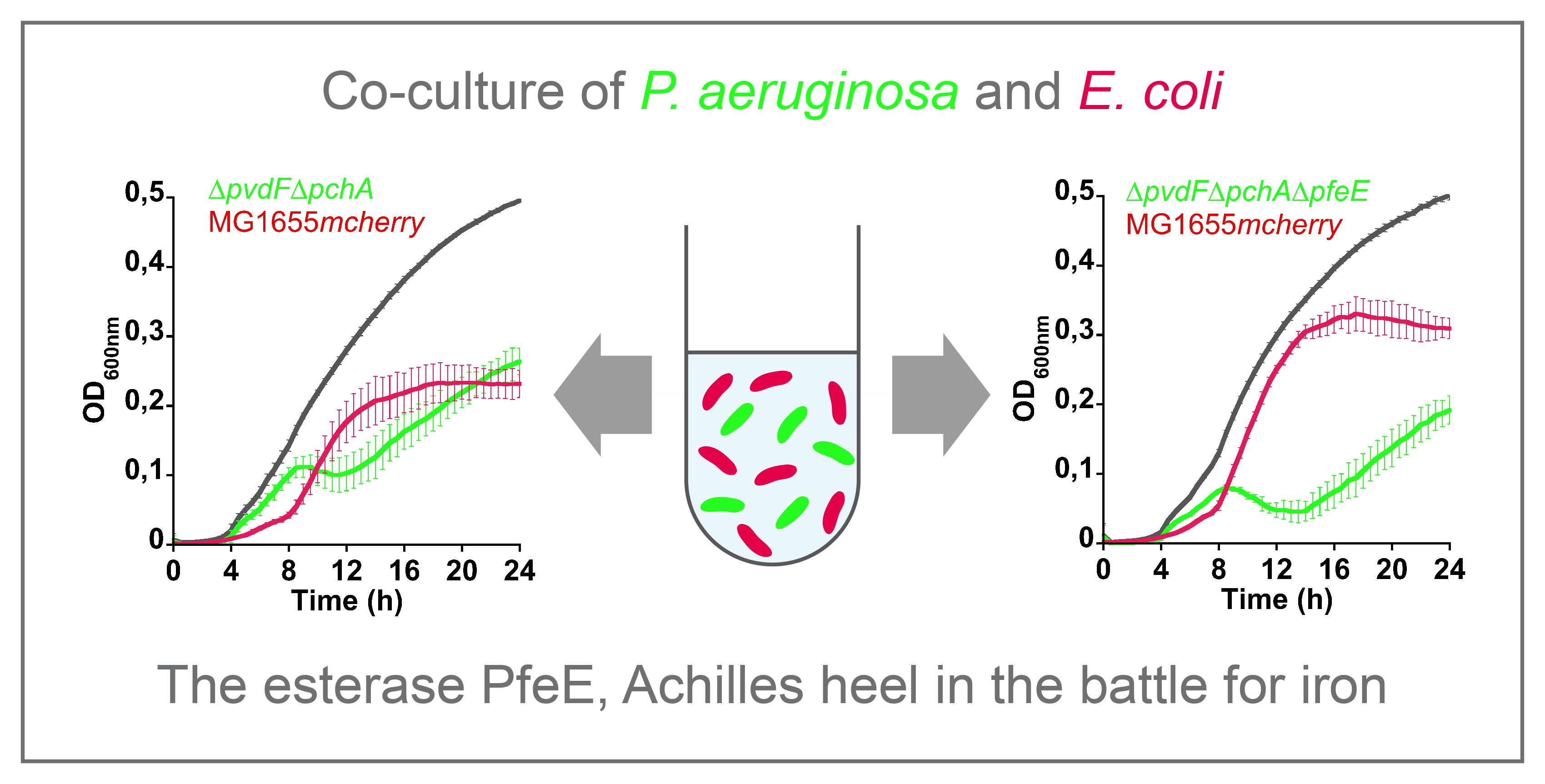
E. coli cells involves the TonB-dependent transporter (TBDT), FepA (
Figure 1) [
3,
10]. This uptake involves the three conserved inner membrane proteins—TonB, ExbB, and ExbD—which transduce energy generated by the proton motive force of the inner membrane to the outer-membrane transporter, FepA [
11]. In the periplasm, the ferri-ENT binds to the periplasmic binding protein, FepB [
12], and is transported into the cytoplasm by the ABC transporter FepDGC (FepD and FepG forming the permease and FepC being the ATPase) [
13,
14,
15]. In the cytoplasm, the esterase Fes hydrolyses ENT into three molecules of N-(2,3-dihydrobenzoyl) serine (DHBS), which are still able to chelate ferric iron [
16,
17,
18]. Iron needs to be reduced by the NADPH-dependent reductase, YdjH, to be released from DHBS and transfered to cytoplasmic iron-binding proteins [
19]. These gene sets (
fepA,
fepB,
fepDGC, and
fes) necessary to access to iron
via the siderophore ENT in
E. coli are present in many genomes (
Yersinia enterocolitica,
Y. pestis,
Y. pseudotuberculosis,
Salmonella enterica,
S. typhimurium, and
Shigella species), even if these bacteria are unable to produce ENT. Indeed, bacteria often use xenosiderophores (siderophores produced by other organisms) to access iron in a siderophore piracy strategy.
The opportunist pathogen
P. aeruginosa also uses ENT as a xenosiderophore, but the molecular mechanisms involved are clearly different from those described for
E. coli. Data from Poole
et al. showed, at the beginning of the 1990s, that there are at least two uptake systems for ferri-ENT in
P. aeruginosa: one of higher affinity, which is specifically inducible by ENT under iron-limiting conditions and involves the outer-membrane transporter, PfeA, and a second of lower affinity and independent of ENT for induction [
20,
21]. This notion was reinvestigated and the data clearly showed that the TBDT involved in high-affinity uptake is PfeA, that the transcription of
pfeA gene is induced by ENT [
21,
22,
23,
24], and that the low-affinity TBDT is PirA [
23]. Next to the
pfeA gene on the chromosome is the
pfeE gene, encoding a periplasmic esterase that acts by hydrolyzing ferri-ENT to promote iron release [
25]. In
P. aeruginosa, ENT releases iron in the bacterial periplasm and does not enter the cytoplasm at any time [
25]. A similar mechanism has also been described for
Campylobacter, in which a periplasmic trilactone esterase, Cee, also hydrolyses ferri-ENT in the bacterial periplasm [
26]. Moreover, PfeA and PfeE expression is induced by the presence of ENT in the bacterial environment and a two-component system, PfeS/PfeR, with PfeS being the inner membrane sensor that detects the presence of ENT-Fe in the bacterial periplasm and PfeR the transcriptional regulator [
22]. According to the
P. aeruginosa genome, a similar PirS/PirR two-component system also regulates the transcription of PirA [
27].
In addition to the ability to access iron
via ENT,
P. aeruginosa is able to produce two siderophores—pyoverdine (PVD) and pyochelin (PCH) [
28]—and is also able to use many other xenosiderophores. For each of these siderophores or xenosiderophores used by the pathogen,
P. aeruginosa possesses in its genome, genes encoding for a specific TBDT involved in the capture and uptake across the outer membrane of the ferric forms of these chelators and proteins involved in the mechanisms of iron release.
Table S1 in the
Supplementary Materials summarizes all the known iron uptake pathways used by
P. aeruginosa PAO1.
Here, we further investigated iron uptake by ENT in P. aeruginosa. We show that ENT-Fe apparently interacts with PfeS, the sensor of the PfeS/PfeR two-component system, before its hydrolysis by PfeE. Moreover, in the absence of the sensor, PfeS, the two proteins, PfeA and PfeE, are constitutively expressed. Grow assays under iron restricted conditions showed that P. aeruginosa is unable to grow in the presence of ENT if pfeE is deleted. Finally, two-species co-cultures between P. aeruginosa and E. coli highlight the key and unique role PfeE plays in the ability of P. aeruginosa to grow in the presence of E. coli producing ENT, especially when P. aeruginosa is unable to produce its own siderophores.
2. Results
2.1. Insights on the Variation of pfeA and pfeE Trancription
Transcription of the
pfeA and
pfeE genes is induced by the PfeS/PfeR two-component system in the presence of ENT [
29]. We used RT-qPCR to evaluate the importance of the proteins PfeA, PirA, PfeE, and PfeS in the regulation of the transcription of the
pfeA and
pfeE genes. Deletion mutants of the
pfeA,
pirA,
pfeE, and
pfeS genes (
Table S2) were grown under iron-restricted conditions in the absence of ENT and the transcription levels of the
pfeA,
pfeE, and
pirA genes followed (
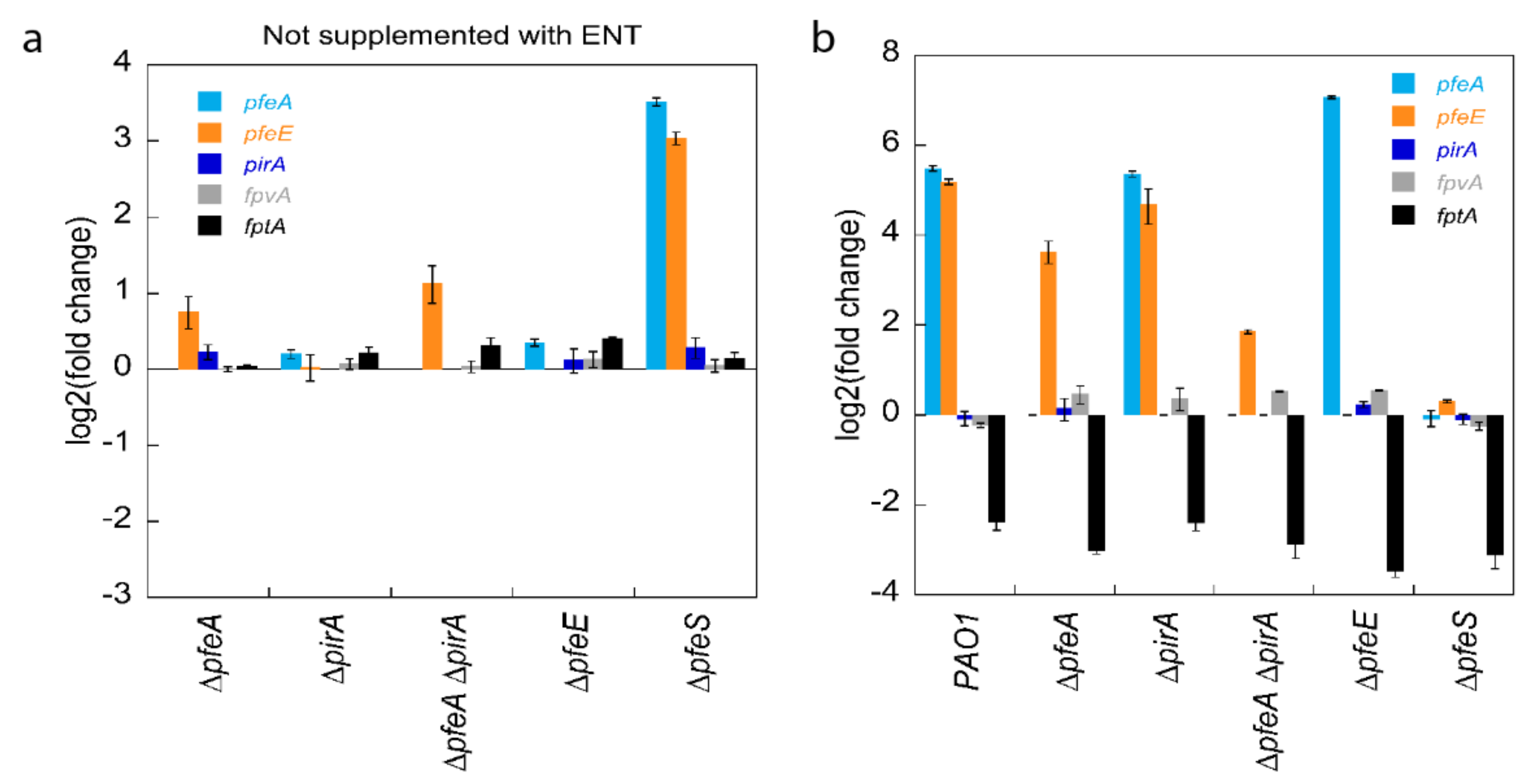
Figure 2a).
pfeS deletion had an effect on the transcription of
pfeA and
pfeE, leading to log
2 fold differences of 3.5 and 3.0, respectively, relative to their level of transcription in PAO1. In the absence of PfeS, PfeR probably becomes active because it is no longer associated with its inner-membrane sensor and, consequently, the regulator induces the transcription of the genes of the ENT pathway. The deletion of
pfeA and of both
pfeA and
pirA had a small effect on the transcription of
pfeE; in these two mutants, PfeR seems to be slightly more active via an unknown mechanism.
The experiment was repeated with 10 µM ENT in the growth media and we compared the level of transcription for each strain between cultures in the presence of ENT and those in its absence (
Figure 2b). In PAO1, as previously described [
24,
29,
32,
33], the presence of 10 µM ENT in the growth medium induced the transcription of
pfeA and
pfeE, with a log
2 fold change of 5.5 and 5.2, respectively. As discussed above, the absence of the sensor PfeS of the two-component system (∆
pfeS strain) resulted in the induction of
pfeA and
pfeE gene transcription in the absence of ENT (
Figure 2a). The addition of ENT to the growth media of this mutant had no additional effect on the transcription of these two genes (
Figure 2b). Growth of the ∆
pfeA strain in the presence of ENT resulted in the induction of
pfeE transcription that was of lower intensity, as in PAO1 (log2 fold change of 3.6), probably because ENT-Fe cannot enter anymore in the
P. aeruginosa periplasm by PfeA but only by PirA.
pfeA and
pfeE transcription in the ∆
pirA mutant was similar as in PAO1, probably because all ENT-Fe is transported efficiently by PfeA and the uptake
via PirA is less essential. The activation of
pfeE transcription was very low for the double
pfeA and
pirA deletion mutant (∆
pfeA∆
pirA) (log
2 fold change of 1.8), because there was no more uptake of ENT-Fe across the outer membrane. On the contrary, the presence of ENT in the growth media of ∆
pfeE led to the strong induction of
pfeA transcription (log2 of 7.1), indicating that ENT-Fe hydrolysis is not necessary for its interaction with PfeS of the two-component system. In the absence of PfeE, ferri-ENT was no longer hydrolyzed, probably accumulating in the bacterial periplasm and explaining the strong stimulation of
pfeA transcription observed in the ∆
pfeE strain. This observation is consistent with previous data showing that TCV-Fe (a non-hydrolysable tris-catechol iron chelator) is also able to induce
pfeA and
pfeE transcription with a higher efficiency than ENT-Fe [
24]. Finally,
pirA transcription was not activated by the presence of ENT in any of the strains or conditions tested, indicating that ENT-Fe is unable to interact with the sensor PirS of the PirS/PirR two-component system.
In conclusion, ENT-Fe needs to be transported across the P. aeruginosa outer membrane by either PfeA or PirA to activate the PfeS/PfeR two-component system. Once inside the bacterial periplasm, ENT-Fe interacts with the PfeS sensor to activate the transcription of the pfeA and pfeE genes before its hydrolysis by PfeE. PirA imports ENT-Fe across the outer membrane, but ENT-Fe is unable to activate the transcription of the pirA gene, which indicates that ENT-Fe is unable to interact with the two-component PirS/PirR system. Finally, PfeR is constitutively active in a pfeS deletion mutant, even in the absence of ENT.
2.2. Insights on the Mechanisms of Repression of PCH Pathway Transcription in the Presence of ENT
We previously showed that 10 µM ENT in the growth medium of
P. aeruginosa cells also represses transcription of the genes encoding for the proteins involved in iron acquisition by the siderophore PCH (PCH pathway) produced by
P. aeruginosa [
24,
32,
33]. Here, we observed the repression of
fptA transcription (TBDT of PCH-Fe) by RT-qPCR, with a log
2 fold change between 2.3 and 3.5 for PAO1 and all mutants tested (
Figure 2b), indicating that ENT-Fe does not need to enter
P. aeruginosa cells by PfeA or PirA or interact somehow with the PfeS/PfeR two-component system to repress the endogenous PCH pathway. Such repression occurred only when ENT was present in the growth media and was completely independent of the expression of PfeA, PfeE, PfeS, and PfeR. The observed decrease in transcription of
fptA and of the genes of the PCH locus [
24,
32,
33] is assuredly a consequence of the competition for iron between ENT and PCH in the bacterial environment, resulting in less PCH-Fe being imported by FptA to activate the positive transcriptional regulator, PchR [
34,
35]. There was no effect on the transcription of
fpvA, the TBDT of PVD-Fe, the other siderophore produced by
P. aeruginosa.
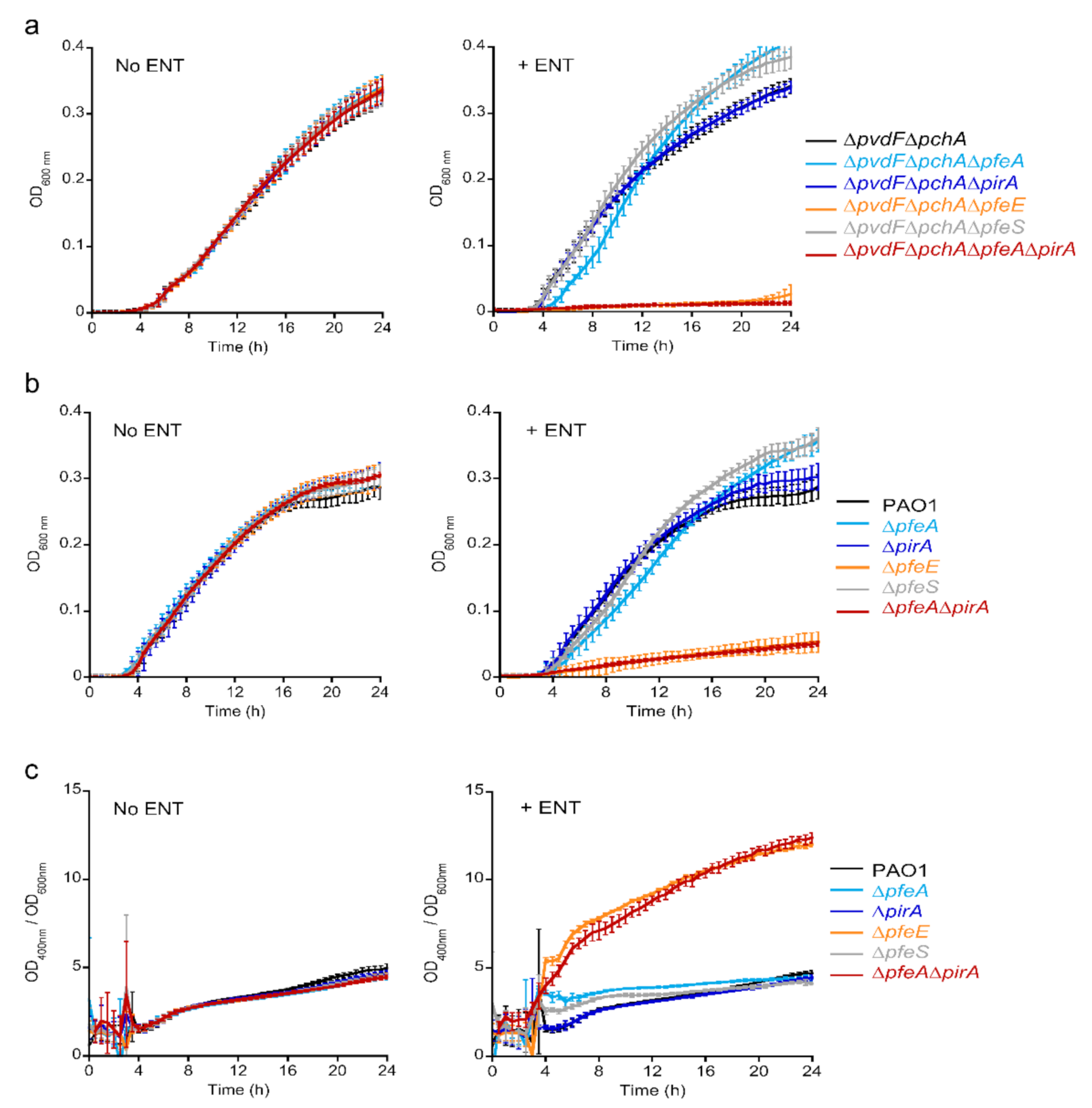
2.3. PfeE Is an Irreplaceable Enzyme to Access Iron via ENT
According to the literature, ENT-Fe complexes can be transported across the outer membrane by two TBDTs, PfeA and PirA [
21,
23,
33]. Then, the siderophore-Fe complex is hydrolyzed by PfeE to facilitate iron release [
25]. We evaluated the relative importance of the proteins encoded by the
pfeA,
pirA, and
pfeE genes in the ability of
P. aeruginosa cells to access iron in the presence of ENT by carrying out growth assays under iron-restricted conditions with
P. aeruginosa strains deleted for at least one of these genes in (i) a PVD and PCH-deficient background (∆
pvdF∆
pchA,
Table S2) to avoid any iron uptake by PVD and PCH, the two siderophores produced by
P. aeruginosa (
Figure 3a), and (ii) a wild-type background (
Figure 3b). The growth assays were carried out in CAA medium (containing approximately 20 nM iron, [
36]), with or without 10 µM ENT. As ENT is a very strong iron chelator (Ka = 10
52 M
−1, [
9]), all iron is chelated by ENT under these growth conditions, resulting in ENT-Fe being the only iron source.
As previously described, the addition of ENT did not inhibit the growth of the
pfeA and
pirA single mutants [
23] in either background (strains producing or not producing the siderophores PVD and PCH) (
Figure 3a,b). In the ∆
pvdF∆
pchA background (strain unable to produce PVD and PCH), we observed complete growth inhibition with the double
pirA and
pfeA mutant, indicating that both of these transporters are involved in iron acquisition by ENT and that one can replace the other if it is absent, as described previously [
23]. We also observed complete growth inhibition with the
pfeE mutant in the ∆
pvdF∆
pchA background (∆
pvdF∆
pchA∆
pfeE strain), confirming the key role of this esterase in iron acquisition by ENT: if ENT cannot be hydrolyzed by this enzyme, iron remains complexed with it in the bacterial periplasm and does not become accessible to the bacteria. In the PAO1 background, we observed strong growth inhibition with the
pfeE mutation and the double
pfeA and
pirA mutation (strains ∆
pfeE and ∆
pfeA∆
pirA), but with residual growth. Such residual growth is likely due to the high production of the siderophore PVD that is observed in these mutants (
Figure 3c). We were unable to monitor PCH production, even after extraction from the growth media, because of the overlap of the absorbance spectra of PCH and ENT.
Deletion of the inner membrane sensor
pfeS had no effect on bacterial growth (
Figure 3a,b), which is consistent with the RT-qPCR data above, showing that the
pfeA and
pfeE genes are transcribed and the corresponding proteins are certainly expressed in this mutant. There was no increase in PVD production by the ∆
pfeS strain compared to PAO1 (
Figure 3c), indicating that the PVD pathway is not involved in iron uptake in this mutant.
The levels of PVD production were highly similar between the PAO1, ∆pfeA, ∆pirA, and ∆pfeS strains in the absence or presence of ENT; the addition of ENT did not stimulate the production of PVD (panels c). Stimulation occurred only for the ∆pfeE and ∆pfeA∆pirA mutants, which are unable to access iron chelated by ENT. P. aeruginosa adapted by increasing its PVD production. This adaptation does not appear to be essential for the growth of any of the other mutants, the pathogen being able to access iron either via ENT or the lower levels of PVD produced.
In conclusion, ENT-Fe complexes can be imported across the outer membranes of P. aeruginosa by the two TBDTs, PfeA and PirA. If one is absent, the other is able to take over uptake. On the contrary, PfeE cannot be replaced; in its absence, bacteria are unable to access iron from ENT. Finally, a pfeS mutant is able to access iron via ENT like PAO1, because both PfeA and PfeE are expressed, as in the wild type strain.
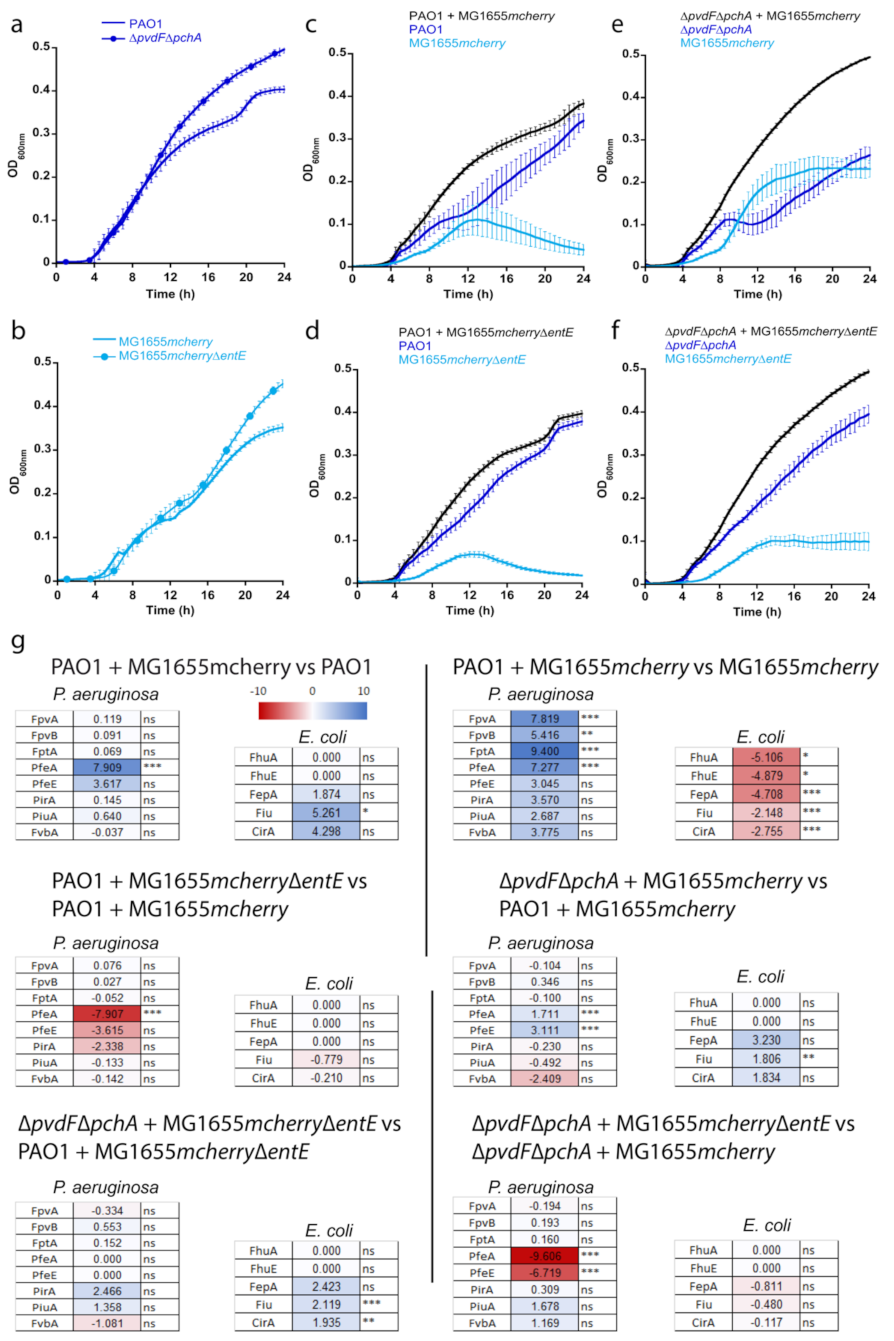
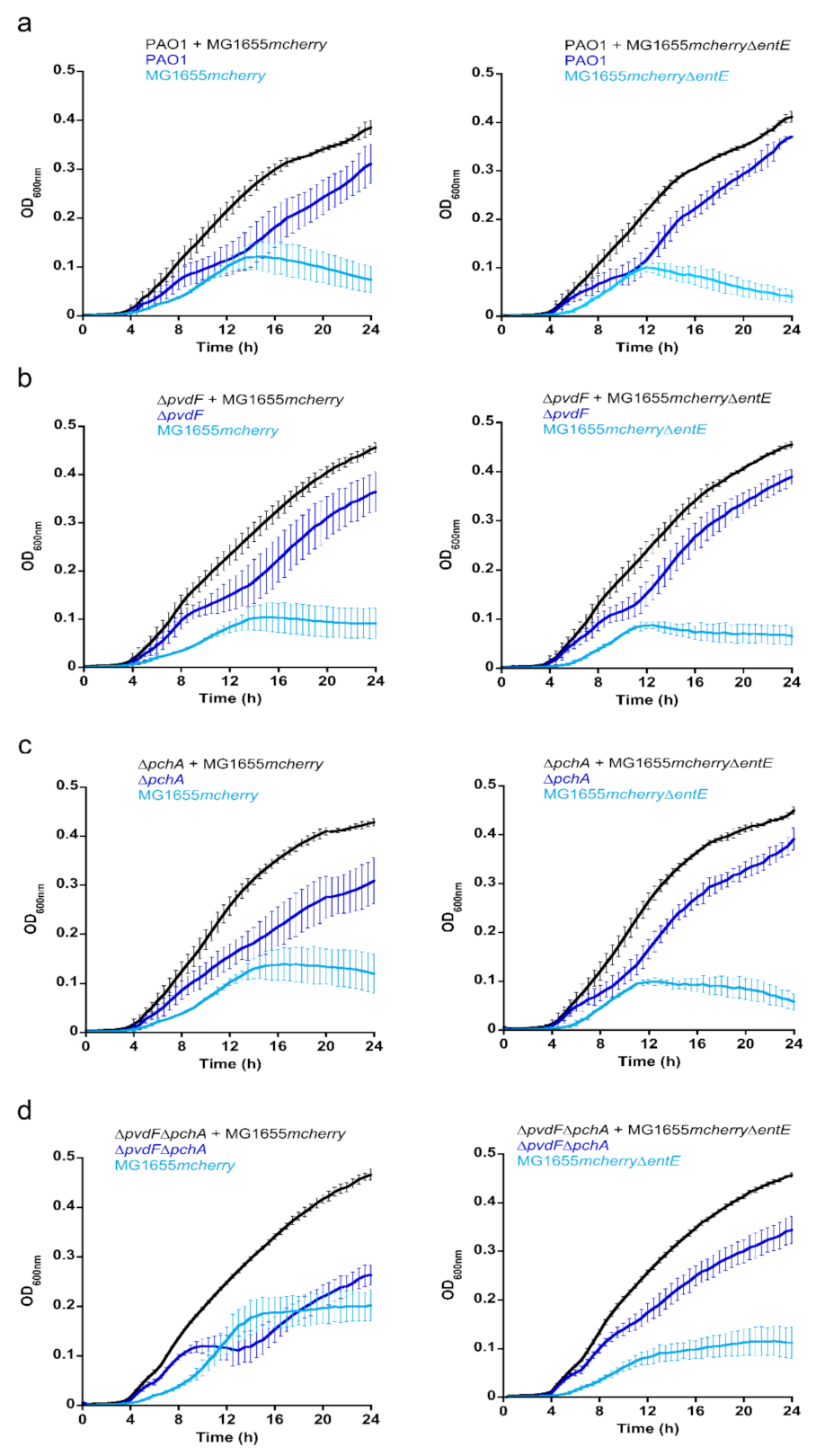
2.4. Co-Cultures between P. aeruginosa and E. coli: The Impact of ENT
We investigated the importance of ENT in co-cultures between
P. aeruginosa and
E. coli strains. We studied this bi-species system in planktonic culture using an iron-restricted medium in which both strains show very similar growth rates (
Figure 4a,b). We first grew
P. aeruginosa, able, or not, to produce its own siderophores PVD and PCH (PAO1 or ∆
pvdF∆
pchA strains) in the presence of
E. coli strains, able, or not, to produce ENT (MG1655
mcherry or MG1655
mcherry∆
entE strains), which resulted in four combinations of cultures (
Figure 4c–f). Both strains of
E. coli carry a plasmid harboring the gene encoding the fluorescent protein mCherry, used to differentiate
E. coli from
P. aeruginosa in the co-cultures. The OD was monitored at 600 nm and the mCherry fluorescence at 610 nm (excitation 570 nm) for each co-culture. The two strains of
E. coli were also grown alone to be able to relate the intensity of mCherry fluorescence to the OD at 600 nm of
E. coli throughout the co-culture (
Figure S1). These values were used to identify the portion of the OD signal at 600 nm in the co-cultures due to
E. coli and
P. aeruginosa. The production of the siderophore PVD by
P. aeruginosa strains in the various co-cultures was followed by monitoring the characteristic absorbance of PVD at 400 nm [
37] (
Figure S1). PCH production by
P. aeruginosa and ENT production by MG1655
mcherry could not be monitored directly in the bacterial cultures because the corresponding absorbances were too low and also slightly overlapping.
When P. aeruginosa was able to produce its own siderophores—PVD and PCH—it largely outgrew E. coli in co-cultures (E. coli producing or not producing ENT, MG1655mcherry or MG1655mcherry∆entE strains). However, when the ∆pvdF∆pchA strain was co-cultured with the strain of E. coli capable of producing ENT (MG1655mcherry), the two bacterial strains were able to co-exist, showing relatively similar ODs. Finally, when neither of the two strains produced siderophores (∆pvdF∆pchA and MG1655mcherry∆entE), P. aeruginosa once again outgrew E. coli.
We also investigated the importance of each siderophore produced by
P. aeruginosa, PVD and PCH, in the competition in the co-cultures (
Figure 5). Strains unable to produce either PVD or PCH were grown in the presence of MG1655
mcherry or MG1655
mcherry∆
entE. As long as one of the siderophores, PVD or PCH, was produced by
P. aeruginosa, it made up the majority in the co-cultures and
E. coli growth remained repressed. When
P. aeruginosa was unable to produce either of its siderophores, the growth of
E. coli was less repressed and both bacterial species were able to grow in similar proportions.
Overall, these data show that the ability to produce ENT is an advantage for E. coli only if P. aeruginosa is unable to produce both of its own siderophores—PVD and PCH. Under all the other conditions tested, P. aeruginosa repressed the growth of E. coli.
2.5. Phenotypic Adaptation of P. aeruginosa and E. coli When Grown Together
We used a differential proteomic approach to compare the proteomes of
P. aeruginosa and
E. coli in the various cultures and co-cultures presented in
Figure 4. The volcano plots of these analyses are presented in
Figure S2 for
P. aeruginosa and
Figure S3 for
E. coli. The heat maps of the outer membrane transporters of PVD (FpvA and FpvB, [
30,
38]), PCH (FptA, [
31]), ENT (PfeA and PirA, [
21,
23]), two other catechol-siderophore outer-membrane transporters (FvbA and PiuA, [
39,
40]), and the esterase PfeE in
P. aeruginosa [
25] are presented in
Figure 4g.
The presence of the ENT-producing MG1655
mcherry induced strong production of PfeA and PfeE in
P. aeruginosa PAO1 relative to the culture of
P. aeruginosa alone in CAA (
Figure 4g, PAO1 + MG1655
mcherry vs. PAO1). The expression of FpvA, FpvB, and FptA was not modified, nor the expression of the other proteins of the PVD and PCH pathways (
Figure S2). Differential proteomic analyses of the co-culture between MG1655
mcherry and
P. aeruginosa PAO1 compared to the culture of
E. coli alone clearly show that FpvA, FpvB, FptA, were highly expressed (
Figure 4g heat map of PAO1 + MG1655
mcherry vs. MG1655
mcherry) as well as all proteins of the PVD and PCH pathways (
Figure S2) and PirA, PiuA, and FvbA, the TBDTs involved in iron acquisition by catechol siderophores. In the co-culture,
P. aeruginosa PAO1 with MG1655
mcherry, the catechol siderophore TBDTs FepA, Fiu, and CirA are expressed in
E. coli, but not FhuA and FhuE, the ferrichrome and desferioxamine TBDTs.
When
E. coli did not produce ENT (MG1655
mcherry∆
entE), PfeA was no longer expressed in
P. aeruginosa PAO1 cells (
Figure 4g heat map of PAO1 + MG1655
mcherry∆
entE vs. PAO1 + MG1655
mcherry). There was slight repression of PirA and PfeE, but FpvA, FpvB, and FptA production were not modified. The expression of
E. coli TBDTs was not modified compared to the co-culture between MG1655
mcherry and PAO1 (
Figure 4g heat map PAO1 + MG1655
mcherry∆
entE vs. PAO1 + MG1655
mcherry).
When
P. aeruginosa was unable to produce PVD and PCH and in the presence of
E. coli producing ENT, the expression of the various proteins of the PVD and PCH pathways was not affected (
Figure S2); these proteins all showed approximately the same level of production as in the co-culture between PAO1 and MG1655
mcherry (
Figure 4, ∆
pvdF∆
pchA + MG1655
mcherry vs. PAO1 + MG1655
mcherry). Moreover, in these conditions, PfeA and PfeE expression was slightly induced, indicating that
P. aeruginosa probably uses more ENT, as in the condition of co-cultures presented in panel 4c.
E. coli, in these co-culture conditions, increased the expression of FepA, Fiu, and CirA (
Figure 4, ∆
pvdF∆
pchA + MG1655
mcherry vs. PAO1 + MG1655
mcherry).
Finally, when
P. aeruginosa was unable to produce PVD or PCH and
E. coli was unable to produce ENT, the expression of PfeA and PfeE dropped sharply relative to the culture between ∆
pvdF∆
pchA and MG1655
mcherry (
Figure 4g, ∆
pvdF∆
pchA + MG1655
mcherry∆
entE vs. ∆
pvdF∆
pchA + MG1655
mcherry).
In conclusion, P. aeruginosa adapts its phenotype to the ability of E. coli to produce, or not ENT, by expressing, or not, PfeA and PfeE; in the presence of ENT-producing E. coli, P. aeruginosa induces the expression of the PfeA and PfeE proteins.
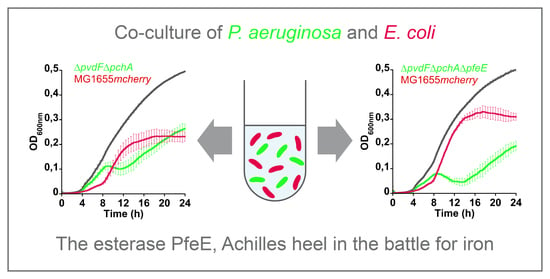
2.6. PfeE: The Achilles’ Heel of P. aeruginosa in Co-Cultures with E. coli
We then co-cultured MG1655
mcherry able to produce ENT with
P. aeruginosa strains unable to produce PVD and PCH (∆
pvdF∆
pchA) and mutated for one of the genes of the ENT pathway (
Figure 6). When
pfeA,
pirA, and
pfeS genes were deleted, both bacterial species were able to co-exist and grow, with equivalent ODs. In contrast, when PfeE was not expressed,
E. coli completely overtook
P. aeruginosa within the first 20 h of culture (
Figure 6d). When PfeE is not expressed,
P. aeruginosa cannot access the iron chelated by ENT, and ENT-Fe complexes certainly accumulate in the periplasm of
P. aeruginosa cells, as all the other proteins of the pathway (PfeA, PirA, PfeS, and PfeR) were expressed. When ∆
pvdF∆
pchA∆
pfeE was grown in the presence of
E. coli strain MG1655
mcherry∆
entE unable to produce ENT,
P. aeruginosa clearly grew better than
E. coli, confirming that sequestration of iron by ENT played a role in the shape of the growth curves of panel d in
Figure 6. As iron was no longer being scavenged by ENT,
P. aeruginosa could again access iron, even if PfeE was not expressed.
Differential proteomic analyses of
P. aeruginosa in co-cultures between MG1655
mchery with ∆
pvdF∆
pchA∆
pfeE (
Figure 6d) and MG1655
mcherry with ∆
pvdF∆
pchA (
Figure 6a) showed a strong difference in PfeE expression, which was due to the fact that
pfeE was deleted in one of the conditions (∆
pvdF∆
pchA∆
pfeE strain). There was no difference in PfeA expression, indicating that PfeA was expressed in equivalent proportions in
P. aeruginosa cells in both co-cultures, even if the growth curves were completely different. We also observed no significant differences in the expression of
E. coli outer-membrane transporters. These proteomic data indicate that differences in the shape of the growth curves of
P. aeruginosa or
E. coli do not necessarily involve phenotypic adaptation.
In conclusion, these data all show that PfeE plays a key and unique role in the ability of P. aeruginosa to acquire iron by ENT and therefore in its ability to grow in the presence of E. coli producing ENT, especially if P. aeruginosa is unable to produce its own siderophores, PVD and PCH.
3. Discussion
ENT is a siderophore produced by the
Enterobacteriaceae, like
E. coli and
Salmonella—[
4,
5,
6,
7], but can also be produced by certain Gram-positive
Streptomyces species [
8]. In addition, many bacteria that are unable to produce ENT, such as
P. aeruginosa, use it in a siderophore piracy strategy. As ENT is the siderophore with the highest known affinity for iron (Ka of 10
52 M
−1, [
9]), the ability of bacteria species to use this chelator to access iron is a key asset in many microbiota.
All bacteria using ENT as a siderophore, do not contain in their genome the same gene set for the access of iron
via this chelator. As described in the introduction, a major difference between the molecular mechanism involved in iron acquisition by ENT in
P. aeruginosa and
E. coli is that in
P. aeruginosa, ENT is hydrolyzed in the bacterial periplasm, whereas in
E. coli, it is hydrolyzed in the cytoplasm (
Figure 1).
P. aeruginosa imports ENT-Fe complexes
via two outer-membrane transporters, PfeA and PirA [
21,
22,
23,
24] (
Figure 1). If one is absent, the other can take over ENT-Fe uptake. In the periplasm, the process of iron release from ENT in
P. aeruginosa cells involves ENT-Fe hydrolyzes by the periplasmic esterase, PfeE, into three molecules of 2,3-DHBS and an iron reduction by a yet unidentified reductase [
25]. Nothing is currently known about how freed iron is imported across the inner membrane into the cytoplasm.
P. aeruginosa is able to detect any ENT-Fe complex in its environment using the PfeS/PfeR two-component system, and the consequence of this is an induction of the transcription of
pfeA and
pfeE genes [
25]. This system allows a controlled phenotypic adaptation of
P. aeruginosa to the presence of the siderophore ENT. However, in the absence of PfeS, transcription of
pfeA and
pfeE genes is induced, even when no ENT is present (
Figure 2), suggesting that PfeR becomes constitutively active because it is no longer associated with the inner membrane sensor PfeS. The regulation of PirA transcription is also associated with a two-component system, PirS/PirR (
Figure 2). However, even if ENT-Fe can be imported across the outer membrane by PirA, it is unable to activate the two-component system PirS/PirR.
In the presence of ENT, the activation of the transcription of the
pfeA and
pfeE genes
via PfeS/PfeR goes hand in hand with the repression of the transcription of the genes involved in iron acquisition by the siderophore PCH, as we have already described previously (
Figure 2 and [
32,
33]). This repression is independent of PfeS/PfeR or the expression of PfeA, PfeE or PirA. It is a consequence of the competition for iron between ENT and PCH in the bacterial environment. According to the affinities of ENT and PCH for iron (K
a of 10
52 M
−1 for ENT [
9] and 10
18 M
−2 for PCH, [
42]), most of the metal is chelated by ENT and fewer PCH-Fe complexes are formed, with the consequence that more ferric-ENT is transported into
P. aeruginosa cells, and are available to induce their corresponding pathway. Concerning the PVD-dependent iron uptake pathway, no repression of the different genes of the PVD operon has been observed in the presence of 10 µM ENT, even if PVD has a lower iron affinity than ENT (Ka of 10
32 M
−1 for PVD, [
37]) and ENT easily removes iron from PVD-Fe [
32]. In addition, no increase in PVD production by PAO1 was observed in the presence of ENT (
Figure 3) or when PAO1 was co-cultured with the
E. coli strain
MG1655mcherry. A large increase in PVD production was only observed for the ∆
pfeE and ∆
pfeA∆
pirA mutants in the presence of ENT (
Figure 3), two strains unable to access iron chelated by ENT.
Growth assays in the iron-deficient planktonic conditions of
P. aeruginosa, alone or in co-cultures with
E. coli, showed that the periplasmic esterase, PfeE, is a key protein in the acquisition of iron by ENT (
Figure 3 and
Figure 6). When absent,
P. aeruginosa can no longer access iron
via ENT and the presence of this siderophore in its environment becomes a major handicap. PfeA can be replaced by PirA and vice versa, but apparently no other enzyme can replace PfeE. PfeS is also irreplaceable, but its absence does not have a substantial effect on the acquisition of iron by ENT, as PfeR is constitutionally active when
pfeS is deleted, and both PfeA and PfeE are expressed.
In co-cultures between
P. aeruginosa and
E. coli,
P. aeruginosa dominates and represses the growth of
E. coli as long as it can produce its own siderophores—PVD and PCH—or
E. coli is unable to produce ENT.
E. coli can only grow and co-exist with
P. aeruginosa if two conditions are simultaneously met:
E. coli must be able to produce ENT and
P. aeruginosa must be deficient in the production of both of its siderophores—PVD and PCH (
Figure 4). Under these conditions, both strains co-exist and share ENT to access iron. In this fragile balance of coexistence between
P. aeruginosa and
E. coli, with
P. aeruginosa pirating the
E. coli siderophore, PfeE plays a key role. If
pfeE is deleted,
E. coli again has the upper hand in the culture, because
P. aeruginosa is no longer able to access the iron chelated by ENT. If
pfeA is mutated,
E. coli growth is less repressed than in the presence of a strain unable to express PfeE because PirA can take over the function of PfeA in iron acquisition
via the siderophore ENT. Surprisingly,
E. coli growth in the presence of ∆
pvdF∆
pchA∆
pfeA∆
pirA is less repressed as in the presence of a ∆
pvdF∆
pchA∆
pfeE mutant. The other genes involved in the ENT pathway in
P. aeruginosa (
pfeA,
pfeS, and
pfeR) are less essential than
pfeE for the ability of
P. aeruginosa to grow in the presence of
E. coli.
This battle for ferri-ENT between P. aeruginosa and E. coli involves PfeE expression but is also based on phenotypic adaptation of the expression of the various iron import pathways by P. aeruginosa. The presence of ENT induces the expression of PfeE and PfeA by P. aeruginosa, whether or not this pathogen produces PVD and/or PCH. The expression of these two proteins is absent when E. coli does not produce ENT. This phenotypic adaptation occurs only at the level of expression of the PfeA and PfeE proteins; the level of FpvA, FpvB, and FptA and all proteins of the PVD and PCH-dependent iron uptake pathways does not change in P. aeruginosa. PirA is expressed in P. aeruginosa in CAA medium, but its expression does not change depending on whether or not E. coli can produce ENT. These phenotypic adaptations are linked to the Fur regulator, which induces the expression of all proteins of the PVD- and PCH-dependent iron uptake pathways, resulting in the production of the siderophores PCH and PVD. It is also linked to the ability of ENT to induce the expression of PfeA and PfeE by P. aeruginosa via the PfeS/PfeR two-component system. In E. coli, the Fur regulator induces the transcription and expression of the ENT-dependent iron-uptake pathways under iron-restricted conditions and such activation occurs regardless of the P. aeruginosa mutant present in the co-culture.
Khare and Tavazoie also investigated the antagonism between
P. aeruginosa and
E. coli in two-species systems and highlighted the molecular complexity of the interactions [
43] but were unable to identify the importance of PfeE. They showed that the siderophores PVD and PCH produced by
P. aeruginosa, as well as the redox-active phenazines, are involved in the repression of
E. coli growth by
P. aeruginosa using genome-scale methods. However, they did not particularly dissect the mechanisms of the interactions concerning the iron-uptake strategies used by the two bacterial species in their battle for iron.
In conclusion, our results show that even bi-species microbial interactions are complex, including both exploitative and interference competition, for which the balance is very fragile, such that the mutation of a single gene can disrupt it. PfeE is the Achilles’ heel of P. aeruginosa in the presence of ENT or bacteria that can produce ENT. A strain of P. aeruginosa that can no longer produce PVD and PCH has the upper hand over the growth of E. coli as long as it can express PfeE. If pfeE is mutated, the growth of P. aeruginosa is strongly repressed by ENT-producing E. coli.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





