
Selection and Validation of Reference Genes for qRT-PCR Analysis in the Oil-Rich Tuber Crop Tiger Nut (Cyperus esculentus) Based on Transcriptome Data



Abstract
1. Introduction
2. Results
2.1. Primer Specificity and PCR Amplification Efficiency
2.2. Threshold Cycle (Ct) Values of Candidate RGs
2.3. Expression Stability of Candidate RGs
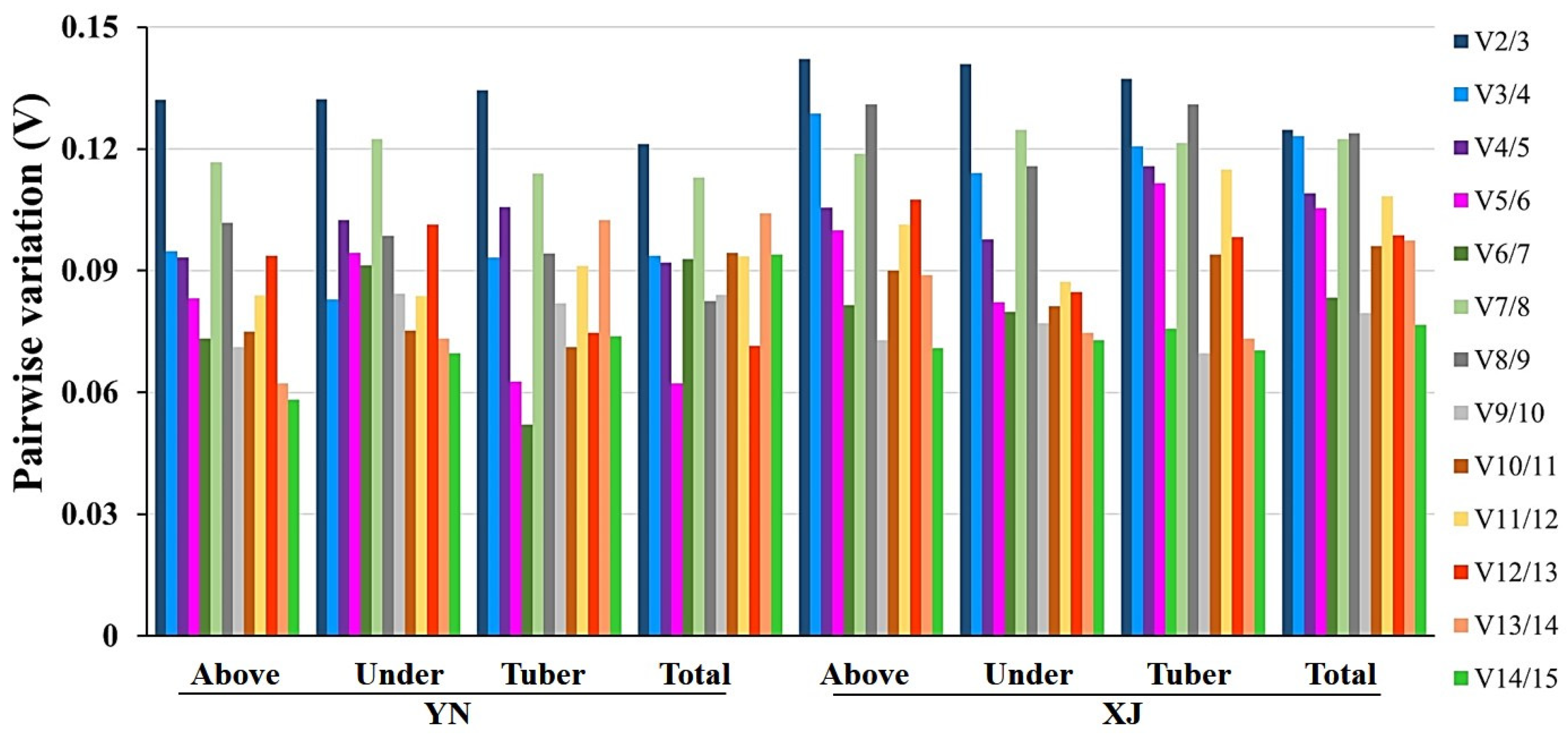
2.3.1. GeNorm analysis
2.3.2. NormFinder Analysis
2.3.3. BestKeeper Analysis
2.4. Unified Rank Lists by RankAggreg
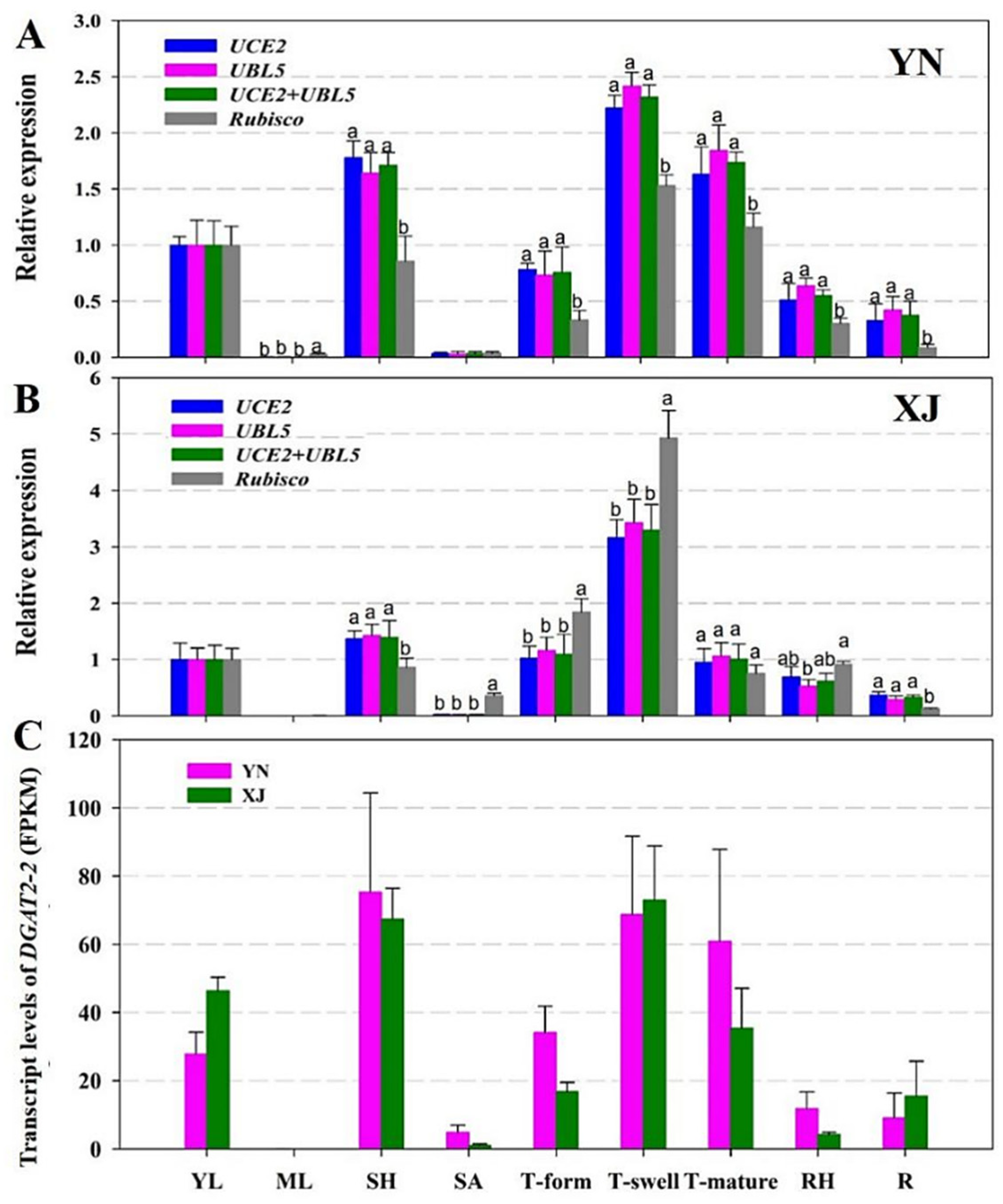
2.5. Validation of Candidate RGs
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Selection of Candidate RGs and Primer Design
4.3. RNA Isolation, cDNA Synthesis, and qRT-PCR
4.4. Data Analysis
4.5. Statistical Method for Rank Aggregation
4.6. Validation of the Candidate RGs
4.7. Library Construction, Sequencing, and Assembly of Transcriptomes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| cDNA | Complementary DNA |
| CLO | Caleosin |
| Ct | Cycle threshold |
| DAS | Day after sowing |
| DGAT | Diacylglycerol acyltransferase |
| FPKM | Fragments per kilobase of exon model per million mapped reads |
| MUFA | Monounsaturated fatty acid |
| qRT-PCR | Quantitative real-time reverse transcription-polymerase chain reaction |
| RG | Reference gene |
| RT | Room temperature |
| Tm | Melting temperature |
References
- Vanhercke, T.; Dyer, J.M.; Mullen, R.T.; Kilaru, A.; Rahman, M.M.; Petrie, J.R.; Green, A.G.; Yurchenko, O.; Singh, S.P. Metabolic engineering for enhanced oil in biomass. Prog. Lipid Res. 2019, 74, 103–129. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Shanklin, J. Triacylglycerol Metabolism, Function, and Accumulation in Plant Vegetative Tissues. Annu. Rev. Plant Biol. 2016, 67, 179–206. [Google Scholar] [CrossRef]
- Zhang, H.; Yasmin, F.; Song, B.H. Neglected treasures in the wild—Legume wild relatives in food security and human health. Curr. Opin. Plant Biol. 2019, 49, 17–26. [Google Scholar] [CrossRef]
- Chang, T.; Cheng, Q.; Zhang, Z.; Guan, C.; Zhao, W.; Wang, L. Advance in Breeding of New Rapeseed Varieties with High Oil Content. Mol. Plant Breed. 2019, 17, 4424–4430. [Google Scholar]
- Ojiewo, C.O.; Janila, P.; Bhatnagar-Mathur, P.; Pandey, M.K.; Desmae, H.; Okori, P.; Mwololo, J.; Ajeigbe, H.; Njuguna-Mungai, E.; Muricho, G.; et al. Advances in Crop Improvement and Delivery Research for Nutritional Quality and Health Benefits of Groundnut (Arachis hypogaea L.). Front. Plant Sci. 2020, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Fobes, J.F.; Mudd, J.B.; Marsden, M.P. Epicuticular lipid accumulation on the leaves of Lycopersicon pennellii (Corr.) D’Arcy and Lycopersicon esculentum Mill. Plant Physiol. 1985, 77, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Lin, Q.; Xu, Y. Tetraena mongolica Maxim can accumulate large amounts of triacylglycerol in phloem cells and xylem parenchyma of stems. Phytochemistry 2007, 68, 2112–2117. [Google Scholar] [CrossRef]
- Defelice, M.S. Yellow nutsedge Cyperus esculentus L.—Snack food of the Gods. Weed Technol. 2002, 16, 901–907. [Google Scholar] [CrossRef]
- Stoller, E.W.; Weber, E.J. Differential cold tolerance, starch, sugar, protein, and lipid of yellow and purple nutsedge tubers. Plant Physiol. 1975, 55, 859–863. [Google Scholar] [CrossRef][Green Version]
- Linssen, J.P.H.; Kielman, G.M.; Cozijnsen, J.L.; Pilnik, W. Comparison of chufa and olive oils. Food Chem. 1988, 28, 279–285. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Hoffmann, G. Monounsaturated fatty acids and risk of cardiovascular disease: Synopsis of the evidence available from systematic reviews and meta-analyses. Nutrients 2012, 4, 1989–2007. [Google Scholar] [CrossRef]
- Terés, S.; Barceló-Coblijn, G.; Benet, M.; Alvarez, R.; Bressani, R.; Halver, J.; Escribá, P. Oleic acid content is responsible for the reduction in blood pressure induced by olive oil. Proc. Natl. Acad. Sci. USA 2008, 105, 13811–13816. [Google Scholar] [CrossRef]
- Palomer, X.; Pizarro-Delgado, J.; Barroso, E.; Vazquez-Carrera, M. Palmitic and oleic acid: The yin and yang of fatty acids in type 2 diabetes mellitus. Trends Endocrinol. Metab. 2018, 29, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Bo, L.; Shibuya, T.; Yogo, Y.; Hara, T.; Matsuo, K. Effects of light quantity and quality on growth and reproduction of a clonal sedge, Cyperus esculentus. Plant Species Biol. 2001, 16, 69–81. [Google Scholar]
- Aljuhaimi, F.; Ghafoor, K.; Ozcan, M.M.; Miseckaite, O.; Babiker, E.E.; Hussain, S. The Effect of solvent type and roasting processes on physico-chemical properties of tigernut (Cyperus esculentus L.) tuber oil. J. Oleo Sci. 2018, 67, 823–828. [Google Scholar] [CrossRef]
- Hu, B.; Li, Y.; Song, J.; Li, H.; Zhou, Q.; Li, C.; Zhang, Z.; Liu, Y.; Liu, A.; Zhang, Q.; et al. Oil extraction from tiger nut (Cyperus esculentus L.) using the combination of microwave-ultrasonic assisted aqueous enzymatic method—Design, optimization and quality evaluation. J. Chromatogr. A 2020, 1627, 461380. [Google Scholar] [CrossRef] [PubMed]
- Turesson, H.; Marttila, S.; Gustavsson, K.E.; Hofvander, P.; Olsson, M.E.; Bulow, L.; Stymne, S.; Carlsson, A.S. Characterization of oil and starch accumulation in tubers of Cyperus esculentus var. sativus (Cyperaceae): A novel model system to study oil reserves in nonseed tissues. Am. J. Bot. 2010, 97, 1884–1893. [Google Scholar] [CrossRef]
- Moral-Anter, D.; Campo-Sabariz, J.; Ferrer, R.; Martin-Venegas, R. Cyperus esculentus L. tubers (tiger nuts) protect epithelial barrier function in Caco-2 cells infected by Salmonella enteritidis and promote Lactobacillus plantarum growth. Nutrients 2021, 13, 71. [Google Scholar] [CrossRef]
- Rubert, J.; Monforte, A.; Hurkova, K.; Pérez-Martínez, G.; Blesa, J.; Navarro, J.L.; Stranka, M.; Soriano, J.M.; Hajslova, J. Untargeted metabolomics of fresh and heat treatment Tiger nut (Cyperus esculentus L.) milks reveals further insight into food quality and nutrition. J. Chromatogr. A 2017, 1514, 80–87. [Google Scholar] [CrossRef]
- Ezeh, O.; Gordon, M.; Niranjan, K. Enhancing the recovery of tiger nut (Cyperus esculentus) oil by mechanical pressing: Moisture content, particle size, high pressure and enzymatic pre-treatment effects. Food Chem. 2016, 194, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ji, H.; Liu, D. Oil biosynthesis in underground oil-rich storage vegetative tissue: Comparison of Cyperus esculentus tuber with oil seeds and fruits. Plant Cell Physiol. 2016, 57, 2519–2540. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, D.; Ji, H. Sucrose metabolism in developing oil-rich tubers of Cyperus esculentus: Comparative transcriptome analysis. BMC Plant Biol. 2018, 18, 151. [Google Scholar] [CrossRef] [PubMed]
- De Castro, O.; Gargiulo, R.; Del Guacchio, E.; Caputo, P.; De Luca, P. A molecular survey concerning the origin of Cyperus esculentus (Cyperaceae, Poales): Two sides of the same coin (weed vs. crop). Ann. Bot. 2015, 115, 733–745. [Google Scholar] [CrossRef]
- Agarwal, P.; Parida, S.K.; Mahto, A.; Das, S.; Mathew, I.E.; Malik, N.; Tyagi, A.K. Expanding frontiers in plant transcriptomics in aid of functional genomics and molecular breeding. Biotechnol. J. 2014, 9, 1480–1492. [Google Scholar] [CrossRef]
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 2010, 50, 227–230. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef]
- Zhang, L.; He, L.-L.; Fu, Q.-T.; Xu, Z.-F. Selection of Reliable Reference Genes for Gene Expression Studies in the Biofuel Plant Jatropha curcas Using Real-Time Quantitative PCR. Int. J. Mol. Sci. 2013, 14, 24338–24354. [Google Scholar] [CrossRef]
- Tang, M.Y.; Bai, X.; Niu, L.J.; Chai, X.; Chen, M.S.; Xu, Z.F. miR172 Regulates both Vegetative and Reproductive Development in the Perennial Woody Plant Jatropha curcas. Plant Cell Physiol. 2018, 59, 2549–2563. [Google Scholar] [CrossRef] [PubMed]
- Llera-Herrera, R.; García-Gasca, A.; Huvet, A.; Ibarra, A.M. Identification of a tubulin-α gene specifically expressed in testis and adductor muscle during stable reference gene selection in the hermaphrodite gonad of the lion’s paw scallop Nodipecten subnodosus. Mar. Genom. 2012, 6, 33–44. [Google Scholar] [CrossRef]
- Almas, D.E.; Kamrodi, A.R. Validation of Appropriate Reference Genes for Real-Time Quantitative PCR Gene Expression Analysis in Rice Plants Exposed to Metal Stresses. Russ. J. Plant Physiol. 2018, 65, 890–897. [Google Scholar] [CrossRef]
- Morga, B.; Arzul, I.; Faury, N.; Renault, T. Identification of genes from flat oyster Ostrea edulis as suitable housekeeping genes for quantitative real time PCR. Fish. Shellfish Immunol. 2010, 29, 945. [Google Scholar] [CrossRef] [PubMed]
- Brattelid, T.; Winer, L.H.; Levy, F.O.; Liestøl, K.; Andersson, K.B. Reference gene alternatives to Gapdh in rodent and human heart failure gene expression studies. Bmc Mol. Biol. 2010, 11, 22. [Google Scholar] [CrossRef]
- Pei, D.S.; Sun, Y.H.; Chen, S.P.; Wang, Y.P.; Hu, W.; Zhu, Z.Y. Zebrafish GAPDH can be used as a reference gene for expression analysis in cross-subfamily cloned embryos. Anal. Biochem. 2007, 363, 291–293. [Google Scholar] [CrossRef]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.R. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Nam, H.Y.; Kim, S.U.; Kim, S.I.; Chang, Y.J. Normalization of reverse transcription quantitative-PCR with housekeeping genes in rice. Biotechnol. Lett. 2003, 25, 1869–1872. [Google Scholar] [CrossRef]
- Wittwer, C.T. High-Resolution DNA Melting Analysis: Advancements and Limitations. Hum. Mutat. 2009, 30, 857–859. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Pihur, V.; Datta, S.; Datta, S. RankAggreg, an R package for weighted rank aggregation. BMC Bioinform. 2009, 10, 62. [Google Scholar] [CrossRef]
- Etschmann, B.; Wilcken, B.; Stoevesand, K.; Von Der Schulenburg, A.; Sterner-Kock, A. Selection of reference genes for quantitative real-time PCR analysis in canine mammary tumors using the GeNorm algorithm. Vet. Pathol. 2006, 43, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Liu, M.; Wang, L.; Li, Z.; Taylor, D.; Li, Z.; Zhang, M. Identification, duplication, evolution and expression analyses of caleosins in Brassica plants and Arabidopsis subspecies. Mol. Genet. Genom. 2016, 291, 971–988. [Google Scholar] [CrossRef]
- Li, C.; Zheng, L.; Wang, X.; Hu, Z.; Zheng, Y.; Chen, Q.; Hao, X.; Xiao, X.; Wang, X.; Wang, G.; et al. Comprehensive expression analysis of Arabidopsis GA2-oxidase genes and their functional insights. Plant Sci. 2019, 285, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shockey, J.M.; Gidda, S.K.; Chapital, D.C.; Kuan, J.-C.; Dhanoa, P.K.; Bland, J.M.; Rothstein, S.J.; Mullen, R.T.; Dyer, J.M. Tung tree DGAT1 and DGAT2 have nonredundant functions in triacylglycerol biosynthesis and are localized to different subdomains of the endoplasmic reticulum. Plant Cell 2006, 18, 2294–2313. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.; et al. The need for transparency and good practices in the qPCR literature. Nat. Methods 2013, 10, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Ménard, R.; Berr, A.; Fuchs, J.; Shen, W.H. The E2 ubiquitin-conjugating enzymes, AtUBC1 and AtUBC2, play redundant roles and are involved in activation of FLC expression and repression of flowering in Arabidopsis Thaliana. Plant J. 2008, 57, 279–288. [Google Scholar] [CrossRef]
- Han, X.; Lu, M.; Chen, Y.; Zhan, Z.; Cui, Q.; Wang, Y. Selection of reliable reference genes for gene expression studies using real-time PCR in tung tree during seed development. PLoS ONE 2012, 7, e43084. [Google Scholar] [CrossRef]
- Chen, C.; Wu, J.; Hua, Q.; Tel-Zur, N.; Xie, F.; Zhang, Z.; Chen, J.; Zhang, R.; Hu, G.; Zhao, J.; et al. Identification of reliable reference genes for quantitative real-time PCR normalization in pitaya. Plant Methods 2019, 15, 70. [Google Scholar] [CrossRef]
- Hong, S.Y.; Seo, P.J.; Yang, M.S.; Xiang, F.; Park, C.M. Exploring valid reference genes for gene expression studies in Brachypodium distachyon by real-time PCR. BMC Plant Biol. 2008, 8, 11. [Google Scholar] [CrossRef]
- Schmidt, G.W.; Delaney, S.K. Stable internal reference genes for normalization of real-time RT-PCR in tobacco (Nicotiana tabacum) during development and abiotic stress. Mol. Genet. Genom. 2010, 283, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Khanlou, K.M.; Van Bockstaele, E. A critique of widely used normalization software tools and an alternative method to identify reliable reference genes in red clover (Trifolium pratense L.). Planta 2012, 236, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Xu, S.; Zhao, Y.; Xia, B.; Wang, R. Selection and validation of appropriate reference genes for quantitative real-time PCR analysis of gene expression in Lycoris aurea. Front. Plant Sci. 2016, 7, 7. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Z.; Huang, H.; Wei, S.; Zhang, C. Selection of relatively exact reference genes for gene expression studies in goosegrass (Eleusine indica) under herbicide stress. Sci. Rep. 2017, 7, 46494. [Google Scholar] [CrossRef]
- Basa, B.; Solti, A.; Sarvari, E.; Tamas, L. Housekeeping gene selection in poplar plants under Cd-stress: Comparative study for real-time PCR normalisation. Funct. Plant Biol. 2009, 36, 1079–1087. [Google Scholar] [CrossRef]
- Chang, E.; Shi, S.; Liu, J.; Cheng, T.; Xue, L.; Yang, X.; Yang, W.; Lan, Q.; Jiang, Z. Selection of reference genes for quantitative gene expression studies in Platycladus orientalis (Cupressaceae) using real-time PCR. PLoS ONE 2012, 7, e33278. [Google Scholar] [CrossRef]
- Zhang, C.; Fu, J.; Wang, Y.; Bao, Z.; Zhao, H. Identification of Suitable Reference Genes for Gene Expression Normalization in the Quantitative Real-Time PCR Analysis of Sweet Osmanthus (Osmanthus fragrans Lour.). PLoS ONE 2015, 10, e0136355. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.J.; Tao, Y.B.; Chen, M.S.; Fu, Q.T.; Li, C.Q.; Dong, Y.L.; Wang, X.L.; He, H.Y.; Xu, Z.F. Selection of reliable reference genes for gene expression studies of a promising oilseed crop, Plukenetia volubilis, by real-time quantitative PCR. Int. J. Mol. Sci. 2015, 16, 12513–12530. [Google Scholar] [CrossRef]
- Hurkan, K.; Sezer, F.; Ozbilen, A.; Taskin, K.M. Identification of reference genes for real-time quantitative polymerase chain reaction based gene expression studies on various Olive (Olea europaea L.) tissues. J. Hortic. Sci. Biotechnol. 2018, 93, 644–651. [Google Scholar] [CrossRef]
- Long, X.; Lu, J.; Kav, N.N.V.; Qin, Y.; Fang, Y. Identification and evaluation of suitable reference genes for gene expression analysis in rubber tree leaf. Mol. Biol. Rep. 2020, 47, 1921–1933. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.S.; Koop, B.F.; Raymond, V.; Walter, M.A. Isolation of a ubiquitin-like (UBL5) gene from a screen identifying highly expressed and conserved iris genes. Genomics 2001, 71, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Ammon, T.; Popowicz, G.M.; Krajewski, M.; Nagel, R.J.; Ares, M., Jr.; Holak, T.A.; Jentsch, S. Role of the ubiquitin-like protein Hub1 in splice-site usage and alternative splicing. Nature 2011, 474, U173–U205. [Google Scholar] [CrossRef]
- Wilkinson, C.R.M.; Dittmar, G.A.G.; Ohi, M.D.; Uetz, P.; Jones, N.; Finley, D. Ubiquitin-like protein Hub1 Is required for pre-mRNA splicing and localization of an essential splicing factor in fission yeast. Curr. Biol. 2006, 16, 2283–2288. [Google Scholar] [CrossRef][Green Version]
- Dittmar, G.A.G.; Wilkinson, C.R.M.; Jedrzejewski, P.T.; Finley, D. Role of a ubiquitin-like modification in polarized morphogenesis. Science 2002, 295, 2442–2446. [Google Scholar] [CrossRef]
- Benedetti, C.; Haynes, C.M.; Yang, Y.; Harding, H.P.; Ron, D. Ubiquitin-like protein 5 positively regulates chaperone gene expression in the mitochondrial unfolded protein response. Genetics 2006, 174, 229–239. [Google Scholar] [CrossRef]
- Kurepa, J.; Walker, J.M.; Smalle, J.; Gosink, M.M.; Davis, S.J.; Durham, T.L.; Sung, D.Y.; Vierstra, R.D. The small ubiquitin-like modifier (SUMO) protein modification system in Arabidopsis. J. Biol. Chem. 2003, 278, 6862–6872. [Google Scholar] [CrossRef] [PubMed]
- Olbrich, M.; Gerstner, E.; Welzl, G.; Fleischmann, F.; Oßwald, W.; Bahnweg, G.; Ernst, D. Quantification of mRNAs and housekeeping gene selection for quantitative real-time RT-PCR normalization in European beech (Fagus sylvatica L.) during abiotic and biotic stress. Z. Fur Nat. C J. Biosci. 2008, 63, 574–582. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, Y.; Liu, J.; Xu, L.; Lai, H.; Chen, Y.; Yang, Z.; Huang, B. Identification and validation of reference genes for Seashore Paspalum response to abiotic stresses. Int. J. Mol. Sci. 2017, 18, 1322. [Google Scholar] [CrossRef]
- Oakley, C.E.; Oakley, B.R. Identification of gamma-tubulin, a new member of the tubulin superfamily encoded by mipA gene of Aspergillus nidulans. Nature 1989, 338, 662–664. [Google Scholar] [CrossRef]
- Cleveland, D.W.; Lopata, M.A.; Macdonald, R.J.; Cowan, N.J.; Rutter, W.J.; Kirschner, M.W. Number and evolutionary conservation of alpha-tubulin and beta-tubulin and cytoplasmic beta-actin and gamma-actin genes using specific cloned cDNA probes. Cell 1980, 20, 95–105. [Google Scholar] [CrossRef]
- Dutcher, S.K. The tubulin fraternity: Alpha to eta. Curr. Opin. Cell Biol. 2001, 13, 49–54. [Google Scholar] [CrossRef]
- Stoppin-Mellet, V.; Peter, C.; Lambert, A.M. Distribution of gamma-tubulin in higher plant cells: Cytosolic gamma-tubulin is part of high molecular weight complexes. Plant Biol. 2000, 2, 290–296. [Google Scholar] [CrossRef]
- Vale, R.D. The molecular motor toolbox for intracellular transport. Cell 2003, 112, 467–480. [Google Scholar] [CrossRef]
- Shi, Y.H.; Zhu, S.W.; Mao, X.Z.; Feng, J.X.; Qin, Y.M.; Zhang, L.; Cheng, J.; Wei, L.P.; Wang, Z.Y.; Zhu, Y.X. Transcriptome profiling, molecular biological, and physiological studies reveal a major role for ethylene in cotton fiber cell elongation. Plant Cell 2006, 18, 651–664. [Google Scholar] [CrossRef]
- Kilmartin, J.V.; Adams, A.E.M. Structural rearrangements of tubulin and actin during the cell-cycle of the yeast saccharomyces. J. Cell Biol. 1984, 98, 922–933. [Google Scholar] [CrossRef]
- Karsenti, E.; Vernos, I. The mitotic spindle: A self-made machine. Science 2001, 294, 543–547. [Google Scholar] [CrossRef]
- Eckardt, N.A. Function of gamma-tubulin in plants. Plant Cell 2006, 18, 1327–1329. [Google Scholar] [CrossRef][Green Version]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.-W.; Sun, Q.-Y.; Wang, Z.-Y.; Sun, Y.-B.; Xu, Z.-F. Using silica particles to isolate total RNA from plant tissues recalcitrant to extraction in guanidine thiocyanate. Anal. Biochem 2008, 374, 426–428. [Google Scholar] [CrossRef]
- Shao, Q.; Liu, X.; Su, T.; Ma, C.; Wang, P. New Insights into the Role of Seed Oil Body Proteins in Metabolism and Plant Development. Front. Plant Sci. 2019, 10, 1568. [Google Scholar] [CrossRef]
- Lardizabal, K.D.; Mai, J.T.; Wagner, N.W.; Wyrick, A.; Voelker, T.; Hawkins, D.J. DGAT2 is a new diacylglycerol acyltransferase gene family—Purification, cloning, and expression in insect cells of two polypeptides from Mortierella ramanniana with diacylglycerol acyltransferase activity. J. Biol. Chem. 2001, 276, 38862–38869. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Dewey, C.N.; Bo, L. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 1–16. [Google Scholar]
- Trapnell, C.; Williams, B.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.; Salzberg, S.; Wold, B.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Description | Primer Sequence (5′-3′; F, forward; R, Reverse) | Amplicon Length (bp) | Primers Tm (°C) | E (%) | R2 |
|---|---|---|---|---|---|---|
| 18S | 18S Ribosomal RNA | F: GGAAGTTTGAGGCAATAACAGG | 140 | 53.71/55.18 | 97.29 | 0.9931 |
| R: TATCCCCATCACGATGAAATTTCTC | ||||||
| ACT | Actin | F: CTCAACCCCAAGGCCAACA | 146 | 53.52/53.81 | 115.7 | 0.9923 |
| R: CCATCACCAGAGTCAAGAACAATA | ||||||
| ADF7 | Actin-depolymerizing factor 7 | F: GACACCGCAAGGGTAAGG | 118 | 52.84/52.25 | 103.59 | 0.9942 |
| R: CAAGCCCCATCTCAGTAGG | ||||||
| CYC | Cyclophilin | F: GGTGAAAAGGGTATCGGTC | 140 | 51.73/52.41 | 105.16 | 0.9998 |
| R: TTTCCGTAGATGGACTCGC | ||||||
| EF1α | Elongation factor | F: CTGGTATGCTTGTGACATTTGG | 175 | 54.07/53.96 | 98.64 | 0.9931 |
| 1-alpha | R: TCGTCCTTGGAGTTGGAGG | |||||
| EF2 | Elongation factor2 | F: TGTCCTTCCGTGAGACCGTA | 203 | 55.85/53.17 | 103.93 | 0.9959 |
| R: TCCTTGTCCCATCCGAACT | ||||||
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | F: ATTCCCAGCAGCACTGGTG | 93 | 52.75/51.93 | 110.61 | 0.9962 |
| R: AGTTGGCACACGGAAAGCC | ||||||
| MDH | Malate | F: ACCCTCTTGTGTCGGTTCTT | 189 | 54.44/52.28 | 95.78 | 0.9944 |
| Dehydrogenase | R: TTGTCATGCCTGGTTTACG | |||||
| PGK | Phosphoglycerate | F: AGAAACCAAGGCTTCGTCA | 168 | 52.24/52.98 | 92.54 | 0.992 |
| Kinase | R: AAGGGAGTCACAACCATCATT | |||||
| RPL11 | Ribosomal protein | F: CTGGATGCTTTGGATTCGG | 174 | 51.82/53.5 | 96.39 | 0.9965 |
| L11 | R: CCTTGGTAACTCTGTGCTGGA | |||||
| Rubisco | Ribulose bisphosphate carboxylase | F: ATGTCTACGTGGTGGACTTGAT | 124 | 52.23/51.92 | 108.65 | 0.9973 |
| R: TGTTTCGGCTTGTGCTTTAT | ||||||
| TUB4 | Tubulin beta-4 | F: CAGGAAGGAGGCTGAAAAT | 156 | 52.06/53.6 | 118.22 | 0.9931 |
| R: GAGGGGAAGACAGAGAAGGT | ||||||
| UCE2 | Ubiquitin-conjugating enzyme 2 | F: ATCATCAAGGAGACCCAGCG | 183 | 55.14/53.39 | 98.21 | 0.9998 |
| R: CTTAGGGGCAGCCATAGGA | ||||||
| UBL5 | Ubiquitin-like | F: ATAATCCCCGTATTTCCACTGC | 96 | 54.34/55.54 | 109.54 | 0.9952 |
| protein 5 | R: GAATCTATCCTATCCACGCTCTCT | |||||
| CLO | Caleosin | F: ACGGCATTGTTTATCCCTGG | 178 | 53.86/54.32 | 117.02 | 0.9914 |
| R: TGTTTGGCTCTGTGTATGTTGTGT | ||||||
| DGAT2-2 | Diacylglycerol | F: CAGGTGGTGTTCAAGAGATGCT | 126 | 54.85/53.26 | 102.45 | 0.9953 |
| O-acyltransferase 2-2 | R: CAAAGGAGAAAACAGGGACAAGT |
| geNorm Rank | Tissues of YN | Tissues of XJ | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aboveground | Underground | Tuber | Total | Aboveground | Underground | Tuber | Total | |||||||||
| Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | |
| 1 | UCE2 | 0.316 | UCE2 | 0.048 | UCE2 | 0.041 | UCE2 | 0.403 | UCE2 | 0.292 | UCE2 | 0.215 | TUB4 | 0.220 | UCE2 | 0.267 |
| 2 | MDH | 0.320 | TUB4 | 0.062 | TUB4 | 0.053 | MDH | 0.409 | UBL5 | 0.292 | TUB4 | 0.233 | UBL5 | 0.241 | ADF7 | 0.296 |
| 3 | UBL5 | 0.398 | ACT | 0.120 | ACT | 0.115 | TUB4 | 0.422 | EF1α | 0.303 | ACT | 0.270 | UCE2 | 0.529 | UBL5 | 0.382 |
| 4 | ADF7 | 0.414 | EF2 | 0.380 | EF2 | 0.275 | UBL5 | 0.593 | CYC | 0.341 | UBL5 | 0.285 | PGK | 0.589 | 18S | 0.474 |
| 5 | ACT | 0.463 | MDH | 0.514 | MDH | 0.373 | CYC | 0.665 | GAPDH | 0.345 | ADF7 | 0.310 | ADF7 | 0.591 | CYC | 0.510 |
| 6 | CYC | 0.560 | RPL11 | 0.598 | GAPDH | 0.461 | EF2 | 0.722 | ACT | 0.346 | 18S | 0.348 | CYC | 0.591 | EF2 | 0.534 |
| 7 | TUB4 | 0.591 | CYC | 0.657 | CYC | 0.542 | RPL11 | 0.756 | MDH | 0.367 | EF1α | 0.375 | RPL11 | 0.679 | GAPDH | 0.551 |
| 8 | EF2 | 0.593 | EF1α | 0.724 | RPL11 | 0.608 | GAPDH | 0.818 | ADF7 | 0.382 | PGK | 0.386 | MDH | 0.711 | TUB4 | 0.574 |
| 9 | EF1α | 0.623 | GAPDH | 0.775 | Rubisco | 0.655 | 18S | 0.869 | TUB4 | 0.384 | RPL11 | 0.397 | EF1α | 0.770 | EF1α | 0.576 |
| 10 | RPL11 | 0.641 | UBL5 | 0.814 | EF1α | 0.684 | ACT | 0.969 | 18S | 0.394 | CYC | 0.419 | EF2 | 0.831 | MDH | 0.612 |
| 11 | GAPDH | 0.654 | 18S | 0.878 | UBL5 | 0.727 | ADF7 | 1.014 | EF2 | 0.446 | EF2 | 0.438 | ACT | 0.855 | RPL11 | 0.643 |
| 12 | 18S | 0.740 | PGK | 0.940 | 18S | 0.791 | EF1α | 1.089 | RPL11 | 0.485 | MDH | 0.448 | 18S | 0.975 | PGK | 0.777 |
| 13 | PGK | 1.163 | ADF7 | 0.995 | PGK | 0.865 | PGK | 1.365 | PGK | 0.513 | GAPDH | 0.485 | GAPDH | 1.320 | ACT | 0.790 |
| 14 | Rubisco | 1.304 | Rubisco | 1.182 | ADF7 | 0.954 | Rubisco | 1.552 | Rubisco | 0.956 | Rubisco | 0.660 | Rubisco | 1.409 | Rubisco | 0.963 |
| Norm- Finder Rank | Tissues of YN | Tissues of XJ | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aboveground | Underground | Tuber | Total | Aboveground | Underground | Tuber | Total | |||||||||
| Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | Gene | M | |
| 1 | UCE2 | 0.152 | TUB4 | 0.211 | TUB4 | 0.221 | UCE2 | 0.359 | UCE2 | 0.233 | TUB4 | 0.190 | TUB4 | 0.191 | UBL5 | 0.231 |
| 2 | UBL5 | 0.320 | RPL11 | 0.290 | RPL11 | 0.236 | UBL5 | 0.409 | EF1α | 0.292 | UBL5 | 0.233 | PGK | 0.241 | UCE2 | 0.345 |
| 3 | MDH | 0.225 | MDH | 0.335 | UCE2 | 0.264 | EF2 | 0.494 | CYC | 0.475 | UCE2 | 0.409 | ADF7 | 0.772 | ADF7 | 0.358 |
| 4 | ADF7 | 0.227 | UCE2 | 0.438 | MDH | 0.308 | MDH | 0.545 | PGK | 0.500 | CYC | 0.476 | CYC | 0.837 | CYC | 0.413 |
| 5 | EF1α | 0.444 | ACT | 0.454 | EF2 | 0.324 | ACT | 0.557 | ACT | 0.550 | EF1α | 0.493 | UBL5 | 0.862 | EF2 | 0.430 |
| 6 | ACT | 0.450 | EF2 | 0.458 | EF1α | 0.350 | ADF7 | 0.592 | MDH | 0.606 | RPL11 | 0.549 | UCE2 | 0.865 | 18S | 0.442 |
| 7 | GAPDH | 0.591 | GAPDH | 0.473 | GAPDH | 0.355 | GAPDH | 0.606 | RPL11 | 0.632 | 18S | 0.598 | RPL11 | 0.884 | ACT | 0.455 |
| 8 | CYC | 0.635 | CYC | 0.476 | ACT | 0.473 | CYC | 0.627 | TUB4 | 0.636 | ACT | 0.604 | MDH | 1.003 | GAPDH | 0.479 |
| 9 | EF2 | 0.639 | EF1α | 0.531 | UBL5 | 0.479 | EF1α | 0.643 | EF2 | 0.667 | EF2 | 0.646 | EF1α | 1.003 | EF1α | 0.485 |
| 10 | TUB4 | 0.691 | UBL5 | 0.563 | CYC | 0.496 | TUB4 | 0.698 | GAPDH | 0.724 | MDH | 0.773 | ACT | 1.037 | TUB4 | 0.714 |
| 11 | RPL11 | 0.694 | PGK | 0.679 | Rubisco | 0.527 | RPL11 | 0.722 | UBL5 | 0.817 | PGK | 0.787 | EF2 | 1.154 | MDH | 0.951 |
| 12 | 18S | 0.833 | 18S | 0.725 | PGK | 0.753 | 18S | 0.754 | 18S | 0.982 | GAPDH | 1.181 | 18S | 1.163 | RPL11 | 1.054 |
| 13 | PGK | 1.419 | ADF7 | 0.800 | 18S | 0.807 | PGK | 1.753 | Rubisco | 1.327 | ADF7 | 1.194 | GAPDH | 1.260 | Rubisco | 1.277 |
| 14 | Rubisco | 1.702 | Rubisco | 1.316 | ADF7 | 0.989 | Rubisco | 1.936 | ADF7 | 1.619 | Rubisco | 1.818 | Rubisco | 2.045 | PGK | 1.494 |
| Best- Keeper Rank | Tissues of YN | Tissues of XJ | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aboveground | Underground | Tuber | Total | Aboveground | Underground | Tuber | Total | |||||||||||||||||
| Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | |
| 1 | MDH | 0.27 | 1.05 | UCE2 | 0.42 | 1.35 | TUB4 | 0.10 | 1.16 | UCE2 | 0.44 | 1.94 | MDH | 0.09 | 0.96 | UCE2 | 0.55 | 1.31 | UCE2 | 0.23 | 1.13 | UCE2 | 0.57 | 1.31 |
| 2 | CYC | 0.30 | 1.36 | MDH | 0.43 | 1.93 | UCE2 | 0.28 | 1.54 | MDH | 0.48 | 2.23 | EF2 | 0.23 | 0.96 | MDH | 0.63 | 1.52 | TUB4 | 0.29 | 1.24 | 18S | 0.65 | 1.70 |
| 3 | UCE2 | 0.39 | 1.85 | ACT | 0.45 | 5.27 | EF2 | 0.40 | 1.80 | EF2 | 0.50 | 2.23 | UCE2 | 0.27 | 1.37 | UBL5 | 0.78 | 3.04 | PGK | 0.45 | 2.12 | UBL5 | 0.67 | 2.06 |
| 4 | ADF7 | 0.42 | 1.82 | TUB4 | 0.48 | 2.20 | MDH | 0.41 | 1.92 | 18S | 0.55 | 3.07 | GAPDH | 0.31 | 1.28 | TUB4 | 0.84 | 3.02 | ACT | 0.51 | 2.37 | ADF7 | 0.68 | 2.55 |
| 5 | UBL5 | 0.60 | 3.28 | ADF7 | 0.49 | 2.16 | 18S | 0.52 | 2.29 | UBL5 | 0.61 | 2.23 | CYC | 0.36 | 1.72 | 18S | 0.84 | 3.23 | MDH | 0.53 | 2.86 | EF2 | 0.72 | 2.70 |
| 6 | EF2 | 0.60 | 3.15 | EF2 | 0.56 | 3.10 | GAPDH | 0.55 | 2.38 | CYC | 0.64 | 2.52 | 18S | 0.36 | 1.38 | ADF7 | 0.88 | 3.54 | CYC | 0.55 | 2.20 | EF1α | 0.83 | 3.20 |
| 7 | 18S | 0.66 | 3.73 | GAPDH | 0.62 | 2.70 | CYC | 0.59 | 3.29 | RPL11 | 0.74 | 3.96 | ADF7 | 0.40 | 2.04 | EF1α | 0.92 | 3.44 | RPL11 | 0.56 | 2.19 | MDH | 0.85 | 3.17 |
| 8 | RPL11 | 0.69 | 4.46 | PGK | 0.63 | 3.21 | RPL11 | 0.70 | 3.56 | GAPDH | 0.77 | 4.24 | EF1α | 0.52 | 2.61 | ACT | 0.96 | 4.00 | GAPDH | 0.62 | 3.08 | CYC | 0.90 | 2.42 |
| 9 | ACT | 0.78 | 5.09 | UBL5 | 0.67 | 3.65 | Rubisco | 0.74 | 2.90 | ADF7 | 0.86 | 4.54 | UBL5 | 0.52 | 2.70 | RPL11 | 1.00 | 3.88 | UBL5 | 0.65 | 4.07 | GAPDH | 0.97 | 3.60 |
| 10 | GAPDH | 0.96 | 5.26 | RPL11 | 0.81 | 3.22 | EF1α | 0.79 | 4.57 | ACT | 0.88 | 5.67 | RPL11 | 0.75 | 3.40 | PGK | 1.07 | 4.01 | ADF7 | 0.80 | 4.24 | RPL11 | 1.22 | 4.76 |
| 11 | EF1α | 1.06 | 5.99 | CYC | 0.88 | 5.54 | UBL5 | 0.81 | 4.37 | TUB4 | 1.26 | 6.17 | TUB4 | 0.83 | 4.31 | EF2 | 1.09 | 4.47 | 18S | 0.89 | 4.66 | PGK | 1.46 | 6.17 |
| 12 | TUB4 | 1.10 | 5.59 | 18S | 0.98 | 4.96 | ACT | 0.92 | 4.55 | EF1α | 1.40 | 7.43 | PGK | 1.08 | 4.85 | CYC | 1.14 | 3.37 | EF1α | 0.90 | 4.63 | TUB4 | 1.69 | 7.09 |
| 13 | Rubisco | 2.31 | 7.59 | EF1α | 1.05 | 4.98 | PGK | 0.92 | 4.37 | Rubisco | 2.34 | 7.50 | ACT | 2.35 | 5.72 | GAPDH | 1.25 | 4.98 | EF2 | 1.01 | 4.83 | ACT | 1.81 | 7.79 |
| 14 | PGK | 2.35 | 8.03 | Rubisco | 1.58 | 8.53 | ADF7 | 0.96 | 4.75 | PGK | 2.41 | 8.22 | Rubisco | 2.35 | 6.30 | Rubisco | 2.74 | 6.33 | Rubisco | 1.21 | 6.15 | Rubisco | 2.60 | 7.83 |
| Rank | Tissues of YN | Tissues of XJ | Tissues of Two Cultivars | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Above | Under | Tuber | Total | Above | Under | Tuber | Total | Above | Under | Tuber | Total | |
| 1 | UCE2 | UCE2 | TUB4 | UCE2 | UCE2 | UCE2 | TUB4 | UCE2 | UCE2 | UCE2 | TUB4 | UCE2 |
| 2 | MDH | TUB4 | UCE2 | MDH | EF1α | UBL5 | PGK | UBL5 | MDH | TUB4 | UCE2 | UBL5 |
| 3 | UBL5 | MDH | EF2 | UBL5 | CYC | TUB4 | UCE2 | ADF7 | EF1α | ACT | MDH | EF2 |
| 4 | ADF7 | ACT | MDH | EF2 | MDH | ADF7 | CYC | 18S | UBL5 | UBL5 | CYC | CYC |
| 5 | ACT | EF2 | RPL11 | CYC | EF2 | 18S | UBL5 | EF2 | CYC | MDH | RPL11 | MDH |
| 6 | EF2 | RPL11 | GAPDH | GAPDH | UBL5 | ACT | ADF7 | CYC | EF2 | RPL11 | EF2 | ADF7 |
| 7 | CYC | GAPDH | ACT | TUB4 | GAPDH | EF1α | MDH | EF1α | ACT | ADF7 | PGK | 18S |
| 8 | EF1α | CYC | CYC | ACT | ACT | RPL11 | ACT | GAPDH | ADF7 | 18S | ACT | GAPDH |
| 9 | TUB4 | UBL5 | EF1α | RPL11 | 18S | MDH | RPL11 | MDH | GAPDH | EF1α | UBL5 | TUB4 |
| 10 | GAPDH | EF1α | Rubisco | 18S | PGK | CYC | EF1α | TUB4 | 18S | EF2 | GAPDH | EF1α |
| 11 | RPL11 | PGK | UBL5 | ADF7 | RPL11 | PGK | EF2 | ACT | RPL11 | CYC | EF1α | ACT |
| 12 | 18S | 18S | 18S | EF1α | ADF7 | EF2 | GAPDH | RPL11 | TUB4 | GAPDH | ADF7 | RPL11 |
| 13 | PGK | ADF7 | PGK | PGK | TUB4 | GAPDH | 18S | PGK | PGK | PGK | 18S | PGK |
| 14 | Rubisco | Rubisco | ADF7 | Rubisco | Rubisco | Rubisco | Rubisco | Rubisco | Rubisco | Rubisco | Rubisco | Rubisco |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, X.; Chen, T.; Wu, Y.; Tang, M.; Xu, Z.-F. Selection and Validation of Reference Genes for qRT-PCR Analysis in the Oil-Rich Tuber Crop Tiger Nut (Cyperus esculentus) Based on Transcriptome Data. Int. J. Mol. Sci. 2021, 22, 2569. https://doi.org/10.3390/ijms22052569
Bai X, Chen T, Wu Y, Tang M, Xu Z-F. Selection and Validation of Reference Genes for qRT-PCR Analysis in the Oil-Rich Tuber Crop Tiger Nut (Cyperus esculentus) Based on Transcriptome Data. International Journal of Molecular Sciences. 2021; 22(5):2569. https://doi.org/10.3390/ijms22052569
Chicago/Turabian StyleBai, Xue, Tao Chen, Yuan Wu, Mingyong Tang, and Zeng-Fu Xu. 2021. "Selection and Validation of Reference Genes for qRT-PCR Analysis in the Oil-Rich Tuber Crop Tiger Nut (Cyperus esculentus) Based on Transcriptome Data" International Journal of Molecular Sciences 22, no. 5: 2569. https://doi.org/10.3390/ijms22052569
APA StyleBai, X., Chen, T., Wu, Y., Tang, M., & Xu, Z.-F. (2021). Selection and Validation of Reference Genes for qRT-PCR Analysis in the Oil-Rich Tuber Crop Tiger Nut (Cyperus esculentus) Based on Transcriptome Data. International Journal of Molecular Sciences, 22(5), 2569. https://doi.org/10.3390/ijms22052569