
Hopping and Flipping of RNA Polymerase on DNA during Recycling for Reinitiation after Intrinsic Termination in Bacterial Transcription


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
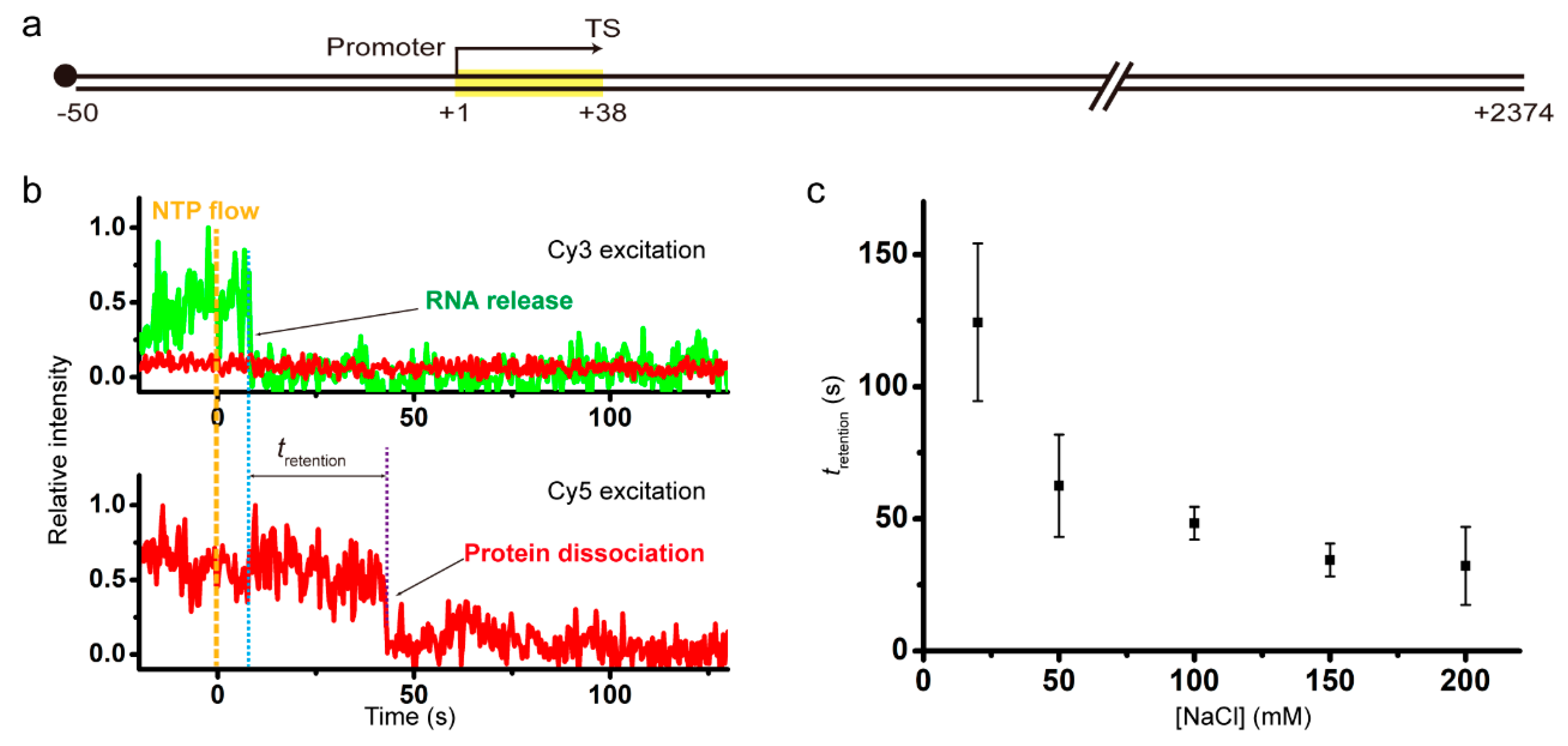
2.1. Salt Dependent Retention of Recycling RNAP on DNA
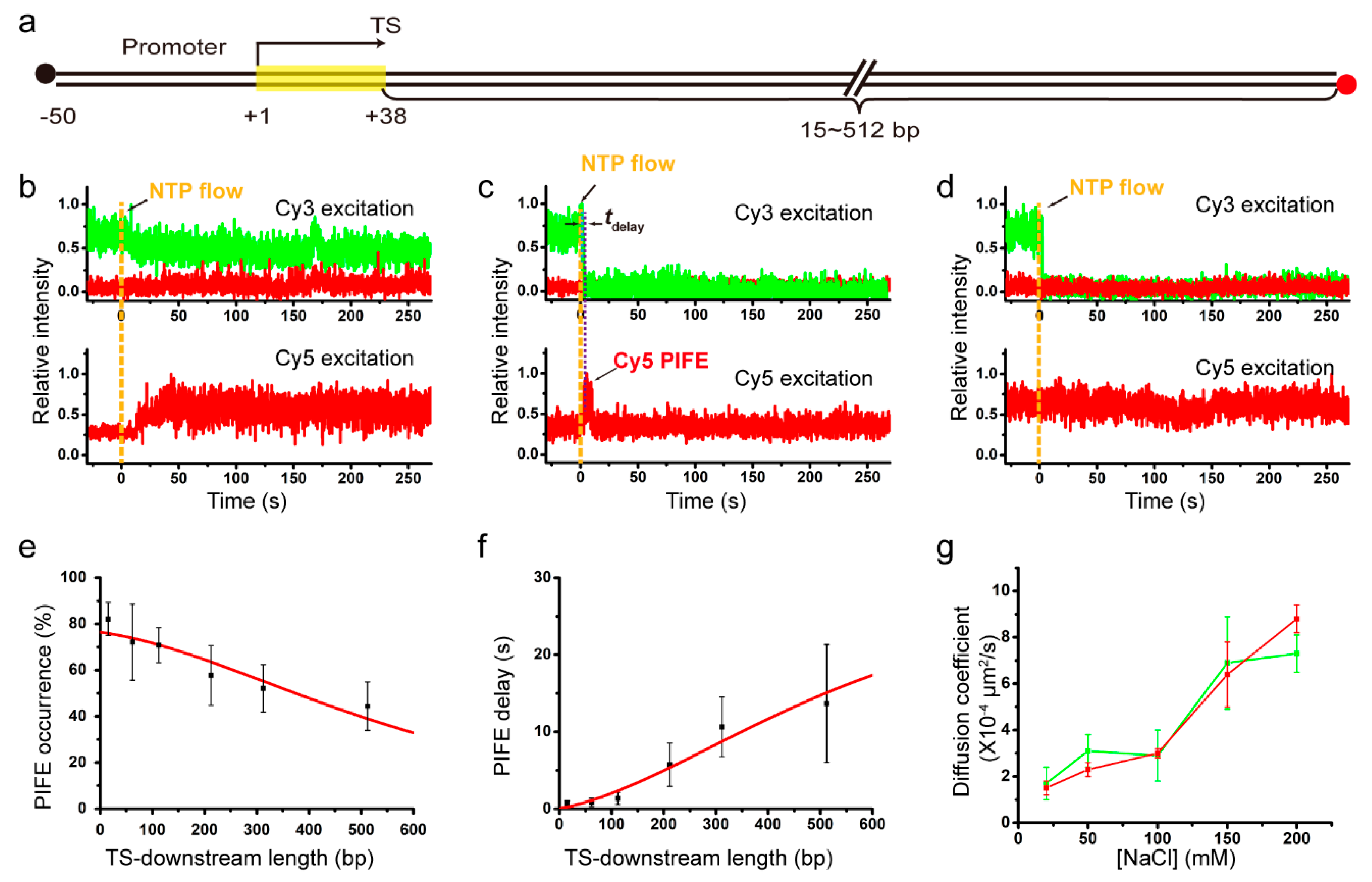
2.2. Salt Dependent 1D Diffusion Coefficient of Recycling RNAP
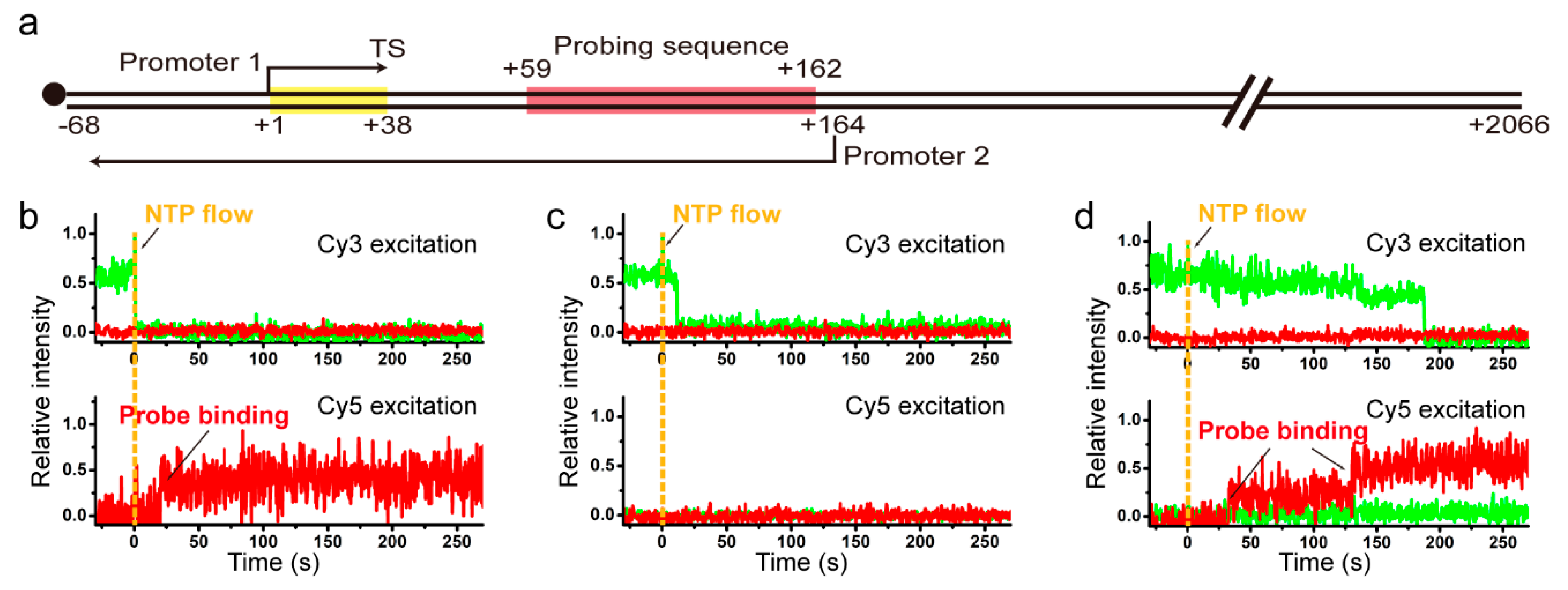
2.3. Sense and Antisense Reinitiation Efficiencies of Recycling RNAP
3. Discussion
4. Materials and Methods
4.1. Single-Molecule Experiments of Transcription Termination
4.2. A 1D Diffusion Model for RNAP Recycling on DNA after Termination
4.3. Calculation of Transcription Reinitiation Efficiencies
4.4. Cy5 Labeling of E. coli Core RNAP
4.5. Preparation of Transcription Templates
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1D | one-dimensional |
| 3D | three-dimensional |
| NEB | New England Biolabs |
| NTP | ribonucleoside triphosphate or ribonucleotide |
| RNAP | RNA polymerase |
| PIFE | protein-induced fluorescence enhancement |
References
- Riggs, A.D.; Bourgeois, S. The lac repressor-operator interaction. 3. Kinetic studies. J. Mol. Biol. 1970, 53, 401–417. [Google Scholar] [CrossRef]
- Berg, O.G.; Winter, R.B.; von Hippel, P.H. Diffusion-driven mechanisms of protein translocation on nucleic acids. 1. Models and theory. Biochemistry 1984, 20, 6929–6948. [Google Scholar] [CrossRef]
- Hedglin, M.; O’Brien, P.J. Hopping enables a DNA repair glycosylase to search both strands and bypass a bound protein. ACS Chem. Biol. 2010, 5, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Gorman, J.; Plys, A.J.; Visnapuu, M.L.; Alani, E.; Greene, E.C. Visualizing one-dimensional diffusion of eukaryotic DNA repair factors along a chromatin lattice. Nat. Struct. Mol. Biol. 2010, 17, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.W.; Kim, Y.; Williams, G.M.; Huck, J.D.; Surtees, J.A.; Finkelstein, I.J. Dynamic DNA binding licenses a repair factor to bypass roadblocks in search of DNA lesions. Nat. Commun. 2016, 7, 10607. [Google Scholar] [CrossRef]
- Cheon, N.Y.; Kim, H.S.; Yeo, J.E.; Schärer, O.D.; Lee, J.Y. Single-molecule visualization reveals the damage search mechanism for the human NER protein XPC-RAD23B. Nucleic Acids Res. 2019, 47, 8337–8347. [Google Scholar] [CrossRef] [PubMed]
- Winter, R.B.; Berg, O.G.; von Hippel, P.H. Diffusion-driven mechanisms of protein translocation on nucleic acids. 3. The Escherichia coli lac repressor-operator interaction: Kinetic measurements and conclusions. Biochemistry 1981, 20, 6961–6977. [Google Scholar] [CrossRef]
- Blainey, P.C.; van Oijen, A.M.; Banerjee, A.; Verdine, G.L.; Xie, X.S. A base-excision DNA-repair protein finds intrahelical lesion bases by fast sliding in contact with DNA. Proc. Natl. Acad. Sci. USA 2006, 103, 5752–5757. [Google Scholar] [CrossRef]
- Gorman, J.; Chowdhury, A.; Surtees, J.A.; Shimada, J.; Reichman, D.R.; Alani, E.; Greene, E.C. Dynamic basis for one-dimensional DNA scanning by the mismatch repair complex Msh2-Msh6. Mol. Cell 2007, 28, 359–370. [Google Scholar] [CrossRef]
- Lin, J.; Countryman, P.; Buncher, N.; Kaur, P.; E, L.; Zhang, Y.; Gibson, G.; You, C.; Watkins, S.C.; Piehler, J.; et al. TRF1 and TRF2 use different mechanisms to find telomeric DNA but share a novel mechanism to search for protein partners at telomeres. Nucleic Acids Res. 2014, 42, 2493–2504. [Google Scholar] [CrossRef]
- Ricchetti, M.; Metzger, W.; Heumann, H. One-dimensional diffusion of Escherichia coli DNA-dependent RNA polymerase: A mechanism to facilitate promoter location. Proc. Natl. Acad. Sci. USA 1988, 85, 4610–4614. [Google Scholar] [CrossRef] [PubMed]
- Kabata, H.; Kurosawa, O.; Arai, I.; Washizu, M.; Margarson, S.A.; Glass, R.E.; Shimamoto, N. Visualization of single molecules of RNA polymerase sliding along DNA. Science 1993, 262, 1561–1563. [Google Scholar] [CrossRef] [PubMed]
- Guthold, M.; Zhu, X.; Rivetti, C.; Yang, G.; Thomson, N.H.; Kasas, S.; Hansma, H.G.; Smith, B.; Hansma, P.K.; Bustamante, C. Direct observation of one-dimensional diffusion and transcription by Escherichia coli RNA polymerase. Biophys. J. 1999, 77, 2284–2294. [Google Scholar] [CrossRef]
- Harada, Y.; Funatsu, T.; Murakami, K.; Nonoyama, Y.; Ishihama, A.; Yanagida, T. Single-molecule imaging of RNA polymerase-DNA interactions in real time. Biophys. J. 1999, 76, 709–715. [Google Scholar] [CrossRef]
- Wang, F.; Redding, S.; Finkelstein, I.J.; Gorman, J.; Reichman, D.R.; Greene, E.C. The promoter-search mechanism of Escherichia coli RNA polymerase is dominated by three-dimensional diffusion. Nat. Struct. Mol. Biol. 2013, 20, 174–181. [Google Scholar] [CrossRef]
- Friedman, L.J.; Mumm, J.P.; Gelles, J. RNA polymerase approaches its promoter without long-range sliding along DNA. Proc. Natl. Acad. Sci. USA 2013, 110, 9740–9745. [Google Scholar] [CrossRef]
- Kang, W.; Ha, K.S.; Uhm, H.; Park, K.; Lee, J.Y.; Hohng, S.; Kang, C. Transcription reinitiation by recycling RNA polymerase that diffuses on DNA after releasing terminated RNA. Nat. Commun. 2020, 11, 450. [Google Scholar] [CrossRef]
- Harden, T.T.; Herlambang, K.S.; Chamberlain, M.; Lalanne, J.B.; Wells, C.D.; Li, G.W.; Landick, R.; Hochschild, A.; Kondev, J.; Gelles, J. Alternative transcription cycle for bacterial RNA polymerase. Nat. Commun. 2020, 11, 448. [Google Scholar] [CrossRef]
- Marklund, E.; van Oosten, B.; Mao, G.; Amselem, E.; Kipper, K.; Sabantsev, A.; Emmerich, A.; Globisch, D.; Zheng, X.; Lehmann, L.C.; et al. DNA surface exploration and operator bypassing during target search. Nature 2020, 583, 858–861. [Google Scholar] [CrossRef]
- Georg, J.; Hess, W.R. Cis-antisense RNA, another level of gene regulation in bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 286–300. [Google Scholar] [CrossRef]
- Thomason, M.K.; Bischler, T.; Eisenbart, S.K.; Förstner, K.U.; Zhang, A.; Herbig, A.; Nieselt, K.; Sharma, C.M.; Storz, G. Global transcriptional start site mapping using differential RNA sequencing reveals novel antisense RNAs in Escherichia coli. J. Bacteriol. 2015, 197, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Pelechano, V.; Steinmetz, L.M. Gene regulation by antisense transcription. Nat. Rev. Genet. 2013, 14, 880–893. [Google Scholar] [CrossRef] [PubMed]
- Lenstra, T.L.; Coulon, A.; Chow, C.C.; Larson, D.R. Single-molecule imaging reveals a switch between spurious and functional ncRNA transcription. Mol. Cell 2015, 60, 597–610. [Google Scholar] [CrossRef]
- Brophy, J.A.; Voigt, C.A. Antisense transcription as a tool to tune gene expression. Mol. Syst. Biol. 2016, 12, 854. [Google Scholar] [CrossRef] [PubMed]
- Tetone, L.E.; Friedman, L.J.; Osborne, M.L.; Ravi, H.; Kyzer, S.; Stumper, S.K.; Mooney, R.A.; Landick, R.; Gelles, J. Dynamics of GreB-RNA polymerase interaction allow a proofreading accessory protein to patrol for transcription complexes needing rescue. Proc. Natl. Acad. Sci. USA 2017, 114, 1081–1090. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, W.; Hwang, S.; Kang, J.Y.; Kang, C.; Hohng, S. Hopping and Flipping of RNA Polymerase on DNA during Recycling for Reinitiation after Intrinsic Termination in Bacterial Transcription. Int. J. Mol. Sci. 2021, 22, 2398. https://doi.org/10.3390/ijms22052398
Kang W, Hwang S, Kang JY, Kang C, Hohng S. Hopping and Flipping of RNA Polymerase on DNA during Recycling for Reinitiation after Intrinsic Termination in Bacterial Transcription. International Journal of Molecular Sciences. 2021; 22(5):2398. https://doi.org/10.3390/ijms22052398
Chicago/Turabian StyleKang, Wooyoung, Seungha Hwang, Jin Young Kang, Changwon Kang, and Sungchul Hohng. 2021. "Hopping and Flipping of RNA Polymerase on DNA during Recycling for Reinitiation after Intrinsic Termination in Bacterial Transcription" International Journal of Molecular Sciences 22, no. 5: 2398. https://doi.org/10.3390/ijms22052398
APA StyleKang, W., Hwang, S., Kang, J. Y., Kang, C., & Hohng, S. (2021). Hopping and Flipping of RNA Polymerase on DNA during Recycling for Reinitiation after Intrinsic Termination in Bacterial Transcription. International Journal of Molecular Sciences, 22(5), 2398. https://doi.org/10.3390/ijms22052398