
Myopathy-Sensitive G-Actin Segment 227-235 Is Involved in Salt-Induced Stabilization of Contacts within the Actin Filament

 ,
,
Abstract
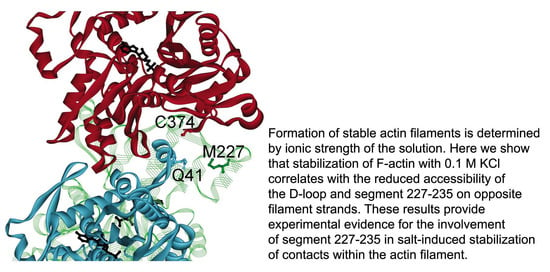
1. Introduction
2. Results
2.1. Effects of the Type of Cation Tightly Bound to Actin and of Solvent Conditions on ATP Hydrolysis in Steady-State F-Actin Solutions
2.2. Effects of the Type of Cation Tightly Bound to Actin and of Solvent Conditions on the Relative Filament Number
2.3. Probing Interprotomer Contacts by DED-F-Actin Fluorescence Measurements
2.4. Probing Interprotomer Contacts in F-Actin by Limited Proteolysis with Subtilisin
2.5. Probing the Effect of KCl on Susceptibility of G-Actin to Limited Proteolysis with Subtilisin
3. Discussion
4. Material and Methods
4.1. Reagents
4.2. Protein Preparations
4.3. Measurements of Cu2+ Binding
4.4. Steady-State ATPase Activity Measurements
4.5. Determination of the Relative Number Concentration of Actin Filaments
4.6. Critical Concentration for Actin Polymerization
4.7. Proteolytic Digestions
4.8. Fluorescence Anisotropy Measurements
4.9. Fluorescence and Light Scattering Measurements
4.10. Electron Microscopy
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holmes, K.C.; Popp, D.; Gebhard, W.; Kabsch, W. Atomic model of the actin filament. Nature 1990, 347, 44–49. [Google Scholar] [CrossRef]
- Holmes, K.C.; Angert, I.; Kull, F.J.; Jahn, W.; Schröder, R.R. Electron cryo-microscopy shows how strong binding of myosin to actin releases nucleotide. Nature 2003, 425, 423–427. [Google Scholar] [CrossRef]
- Oda, T.; Iwasa, M.; Aihara, T.; Maéda, Y.; Narita, A. The nature of the globular- to fibrous-actin transition. Nature 2009, 457, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Splettstoesser, T.; Holmes, K.C.; Noé, F.; Smith, J.C. Structural modeling and molecular dynamics simulation of the actin filament. Proteins Struct. Funct. Bioinform. 2011, 79, 2033–2043. [Google Scholar] [CrossRef]
- Fujii, T.; Iwane, A.H.; Yanagida, T.; Namba, K. Direct visualization of secondary structures of F-actin by electron cryomicroscopy. Nature 2010, 467, 724–728. [Google Scholar] [CrossRef]
- Galkin, V.E.; Orlova, A.; Vos, M.R.; Schröder, G.F.; Egelman, E.H. Near-Atomic Resolution for One State of F-Actin. Structure 2015, 23, 173–182. [Google Scholar] [CrossRef]
- Von der Ecken, J.; Müller, M.; Lehman, W.; Manstein, D.J.; Penczek, P.A.; Raunser, S. Structure of the F-actin-tropomyosin complex. Nature 2015, 19, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Reisler, E. Intermolecular coupling between loop 38–52 and the C-terminus in actin filaments. Biophys. J. 1996, 71, 1914–1919. [Google Scholar] [CrossRef][Green Version]
- Hegyi, G.; Mák, M.; Kim, E.; Elzinga, M.; Muhlrad, A.; Reisler, E. Intrastrand cross-linked actin between Gln-41 and Cys-374. I. Mapping of sites cross-linked in F-actin by N-(4-azido-2-nitrophenyl) putrescine. Biochemistry 1998, 37, 17784–17792. [Google Scholar] [CrossRef]
- Kim, E.; Wriggers, W.; Phillips, M.; Kokabi, K.; Rubenstein, P.A.; Reisler, E. Cross-linking constraints on F-actin structure. J. Mol. Biol. 2000, 299, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Orlova, A.; Galkin, V.E.; VanLoock, M.S.; Kim, E.; Shvetsov, A.; Reisler, E.; Egelman, E.H. Probing the structure of F-actin: Cross-links constrain atomic models and modify actin dynamics 1 1Edited by M. F. Moody. J. Mol. Biol. 2001, 312, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Durer, Z.A.O.; Diraviyam, K.; Sept, D.; Kudryashov, D.S.; Reisler, E. F-actin structure destabilization and DNase-I binding loop fluctuations. Mutational cross-linking and electron microscopy analysis of the loop states and effects on F-actin. J. Mol. Biol. 2010, 395, 544–557. [Google Scholar] [CrossRef]
- Kim, E.; Reisler, E. Intermolecular dynamics and function in actin filaments. Biophys. Chem. 2000, 86, 191–201. [Google Scholar] [CrossRef]
- Das, S.; Ge, P.; Durer, Z.A.O.; Grintsevich, E.E.; Zhou, Z.H.; Reisler, E. D-loop Dynamics and Near-Atomic-Resolution Cryo-EM Structure of Phalloidin-Bound F-Actin. Structure 2020, 28, 586–593.e3. [Google Scholar] [CrossRef] [PubMed]
- Kudryashov, D.S.; Reisler, E. ATP and ADP actin states. Biopolymers 2013, 99, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Kim, E.; Lee, W.-L.; Miller, C.J.; Kuang, B.; Reisler, E.; Rubenstein, P.A. Fluorescence probing of yeast actin subdomain 3/4 hydrophobic loop 262–274. Actin-actin and actin-myosin interactions in actin filaments. J. Biol. Chem. 1997, 272, 16829–16837. [Google Scholar] [CrossRef] [PubMed]
- Owen, C.; DeRosier, D. A 13 Å map of the actin-scruin filament from the limulus acrosomal process. J. Cell Biol. 1993, 123, 337–344. [Google Scholar] [CrossRef]
- Tirion, M.M.; Ben-Avraham, D.; Lorenz, M.; Holmes, K.C. Normal modes as refinement parameters for the F-actin model. Biophys. J. 1995, 68, 5–12. [Google Scholar] [CrossRef][Green Version]
- Moraczewska, J.; Gruszczynska-Biegala, J.; Redowicz, M.J.; Khaitlina, S.Y.; Strzelecka-Golaszewska, H. The DNase-I binding Loop of actin may play a role in the regulation of actin-myosin interaction by tropomyosin/troponin. J. Biol. Chem. 2004, 279, 31197–31204. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.-Q.; Takamoto, K.; Almo, S.C.; Reisler, E.; Chance, M.R. Structure and Dynamics of the Actin Filament. Biochemistry 2005, 44, 3166–3175. [Google Scholar] [CrossRef]
- Chen, W.; Wen, K.-K.; Sens, A.E.; Rubenstein, P.A. Differential Interaction of Cardiac, Skeletal Muscle, and Yeast Tropomyosins with Fluorescent (Pyrene235) Yeast Actin. Biophys. J. 2006, 90, 1308–1318. [Google Scholar] [CrossRef]
- Otterbein, L.R.; Graceffa, P.; Dominguez, R. The Crystal Structure of Uncomplexed Actin in the ADP State. Science 2001, 293, 708–711. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Maéda, Y. Multiple Conformations of F-actin. Structure 2010, 18, 761–767. [Google Scholar] [CrossRef][Green Version]
- Strzelecka-Gołaszewska, H. Divalent Cations, Nucleotides, and Actin Structure. In Results and Problems in Cell Differentiation, Volume 32: Molecular Interactions of Actin; dos Remedios, C.G., Thomas, D.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; Volume 32, pp. 23–41. [Google Scholar]
- Strzelecka-Golaszewska, H.; Wozniak, A.; Hult, T.; Lindberg, U. Effects of the type of divalent cation, Ca2+ or Mg2+, bound at the high-affinity site and of the ionic composition of the solution on the structure of F-actin. Biochem. J. 1996, 316, 713–721. [Google Scholar] [CrossRef]
- Shvetsov, A.; Stamm, J.D.; Phillips, M.; Warshaviak, D.C.; Altenbach, P.A.; Rubenstein, K.H.; Hubbell, W.L.; Reisler, E. Conformational dynamics of loop 262–274 in G- and F-actin. Biochemistry 2006, 45, 6541–6549. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, J.C.; Nowak, K.J.; Durling, H.J.; Beggs, A.H.; Wallgren-Pettersson, C.; Romero, N.; Nonaka, I.; Laing, N.G. Muscle disease caused by mutations in the skeletal muscle alpha-actin gene (ACTA1). Neuromuscul. Disord. 2003, 13, 519–531. [Google Scholar] [CrossRef]
- Wallgren-Pettersson, C.; Pelin, K.; Nowak, K.J.; Muntoni, F.; Romero, N.B.; Goebel, H.H.; North, K.N.; Beggs, A.H.; Laing, N.G. The ENMC International Consortium on Nemaline Myopathy. Genotype-phenotype correlations in nemaline myopathy caused by mutations in the genes for nebulin and skeletal muscle α-actin. Neuromuscul. Disord. 2004, 14, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Korn, E.D.; Carlier, M.F.; Pantaloni, D. Actin polymerization and ATP hydrolysis. Science 1987, 238, 638–644. [Google Scholar] [CrossRef]
- Fujiwara, I.; Takeda, S.; Oda, T.; Honda, H.; Narita, A.; Maéda, Y. Polymerization and depolymerization of actin with nucleotide states at filament ends. Biophys. Rev. 2018, 10, 1513–1519. [Google Scholar] [CrossRef]
- Chou, S.Z.; Pollard, T.D. Mechanism of actin polymerization revealed by cryo-EM structures of actin filaments with three different bound nucleotides. Proc. Natl. Acad. Sci. USA 2019, 116, 4265–4274. [Google Scholar] [CrossRef]
- Jepsen, L.; Sept, D. Effects of Nucleotide and End-Dependent Actin Conformations on Polymerization. Biophys. J. 2020, 119, 1800–1810. [Google Scholar] [CrossRef] [PubMed]
- Asakura, S.; Oosawa, F. Dephosphorylation of adenosine triphosphate in actin solutions at low concentrations of magnesium. Arch. Biochem. Biophys. 1960, 87, 273–280. [Google Scholar] [CrossRef]
- Brenner, S.L.; Korn, E.D. On the mechanism of actin monomer-polymer subunit exchange at steady state. J. Biol. Chem. 1983, 258, 5013–5020. [Google Scholar] [CrossRef]
- Khaitlina, S.Y.; Strzelecka-Gołaszewska, H. Role of the DNase-I-Binding Loop in Dynamic Properties of Actin Filament. Biophys. J. 2002, 82, 321–334. [Google Scholar] [CrossRef]
- Drewes, G.; Faulstich, H. The enhanced ATPase activity of glutathione-substituted actin provides a quantitative approach to filament stabilization. J. Biol. Chem. 1990, 256, 3017–3021. [Google Scholar] [CrossRef]
- Kasai, M.; Oosawa, F. The Exchangeability of actin-bound calcium with various divalent cations. Biochim. Biophys. Acta (BBA) Protein Struct. 1968, 154, 520–528. [Google Scholar] [CrossRef]
- Strzelecka-Gołaszewska, H.; Drabikowski, W. Studies on the exchange of G-actin-bound calcium with bivalent cations. Biochim. Biophys. Acta (BBA) Bioenerg. 1968, 162, 581–595. [Google Scholar] [CrossRef]
- Carlier, M.F. Actin: Protein structure and filament dynamics. J. Biol. Chem. 1991, 266, 1–4. [Google Scholar] [CrossRef]
- Estes, J.E.; Selden, L.A.; Kinosian, H.J.; Gershman, L.C. Tightly-bound divalent cation of actin. J. Muscle Res. Cell Motil. 1992, 13, 272–284. [Google Scholar] [CrossRef]
- Moraczewska, J.; Wawro, B.; Seguro, K.; Strzelecka-Gołaszewska, H. Divalent Cation-, Nucleotide-, and Polymerization-Dependent Changes in the Conformation of Subdomain 2 of Actin. Biophys. J. 1999, 77, 373–385. [Google Scholar] [CrossRef]
- Borovikov, Y.S.; Dedova, I.V.; dos Remedios, C.G.; Vikhoreva, N.N.; Vikhorev, P.G.; Avrova, S.V.; Hazlett, T.L.; Van Der Meer, B.W. Fluorescence depolarization of actin filaments in reconstructed myofibers: The effect of S1 or pPDM-S1 on movements of distinct areas of actin. Biophys. J. 2004, 86, 3020–3029. [Google Scholar] [CrossRef]
- Takashi, R.; Kasprzak, A.A. Measurement of interprotein distances in the acto-subfragment 1 rigor complex. Biochemistry 1987, 26, 7471–7477. [Google Scholar] [CrossRef]
- Schwyter, D.; Phillips, M.; Reisler, E. Subtilisin-cleaved actin: Polymerization and interaction with myosin subfragment 1. Biochemistry 1989, 28, 5889–5895. [Google Scholar] [CrossRef] [PubMed]
- Vahdat, A.; Miller, C.; Phillips, M.; Muhlrad, A.; Reisler, E. A novel 27/16 kDa form of subtilisin cleaved actin: Structural and functional consequences of cleavage between Ser234 and Ser235. FEBS Lett. 1995, 365, 149–151. [Google Scholar] [CrossRef]
- Cooper, J.A.; Buhle, E.L.; Walker, S.B.; Tsong, T.Y.; Pollard, T.D. Kinetic evidence for a monomer activation step in actin polymerization. Biochemistry 1983, 22, 2193–2202. [Google Scholar] [CrossRef] [PubMed]
- Khaitlina, S.Y.; Collins, J.H.; Kuznetsova, I.M.; Pershina, V.P.; Synakevich, I.G.; Turoverov, K.K.; Usmanova, A.M. Physico-chemical properties of actin cleaved with bacterial protease from E. coli A2 strain. FEBS Lett. 1991, 279, 49–51. [Google Scholar] [CrossRef]
- Page, R.; Lindberg, U.; Schutt, C.E. Domain motions in actin. J. Mol. Biol. 1998, 280, 463–474. [Google Scholar] [CrossRef]
- Klenchin, V.A.; Khaitlina, S.Y.; Rayment, I. Crystal Structure of Polymerization-Competent Actin. J. Mol. Biol. 2006, 362, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Egelman, E.H. A tale of two polymers: New insights into helical filaments. Nat. Rev. Mol. Cell Biol. 2003, 4, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Frieden, C.; Lieberman, D.; Gilbert, H.R. A fluorescent probe for coformational changes in skeletal muscle actin. J. Biol. Chem. 1980, 255, 8991–8993. [Google Scholar] [CrossRef]
- Frieden, C. The Mg2+-induced conformational change in rabbit skeletal muscle G-actin. J. Biol. Chem. 1982, 257, 2882–2886. [Google Scholar] [CrossRef]
- Carlier, M.F.; Pantaloni, D.; Korn, E.D. Fluorescence measurements of the binding of cations to high-affinity and low-affinity sites on ATP-G-actin. J. Biol. Chem. 1986, 261, 10778–10784. [Google Scholar] [CrossRef]
- Strzelecka-Golaszewska, H.; Moraczewska, J.; Khaitlina, S.Y.; Mossakowska, M. Localization of the tightly bound divalent-cation—Dependent and nucleotide—Dependent conformation changes in G-actin using limited proteolytic digestion. JBIC J. Biol. Inorg. Chem. 1993, 211, 731–742. [Google Scholar] [CrossRef]
- Guan, J.-Q.; Almo, S.C.; Reisler, E.; Chance, M.R. Structural Reorganization of Proteins Revealed by Radiolysis and Mass Spectrometry: G-Actin Solution Structure Is Divalent Cation Dependent. Biochemistry 2003, 42, 11992–12000. [Google Scholar] [CrossRef] [PubMed]
- Adams, S.B.; Reisler, E. Sequence 18–29 on Actin: Antibody and Spectroscopic Probing of Conformational Changes. Biochemistry 1994, 33, 14426–14433. [Google Scholar] [CrossRef]
- Mejean, C.; Hué, H.K.; Pons, F.; Roustan, C.; Benyamin, Y. Cation binding sites on actin: A structural relationship between antigenic epitopes and cation exchange. Biochem. Biophys. Res. Commun. 1988, 152, 368–375. [Google Scholar] [CrossRef]
- Takamoto, K.; Kamal, J.K.A.; Chance, M.R. Biochemical Implications of a Three-Dimensional Model of Monomeric Actin Bound to Magnesium-Chelated ATP. Structure 2007, 15, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Makino, K.; Yamashita, I.; Namba, K.; Maéda, Y. Distinct Structural Changes Detected by X-Ray Fiber Diffraction in Stabilization of F-Actin by Lowering pH and Increasing Ionic Strength. Biophys. J. 2001, 80, 841–851. [Google Scholar] [CrossRef]
- Kang, H.; Bradley, M.J.; McCullough, B.R.; Pierre, A.; Grintsevich, E.E.; Reisler, E.; De La Cruz, E.M. Identification of cation-binding sites on actin that drive polymerization and modulate bending stiffness. Proc. Natl. Acad. Sci. USA 2012, 109, 16923–16927. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.-J.; Marston, S. Genotype–phenotype correlations in ACTA1 mutations that cause congenital myopathies. Neuromuscul. Disord. 2009, 19, 6–16. [Google Scholar] [CrossRef]
- Parker, F.; Baboolal, T.G.; Peckham, M. Actin Mutations and Their Role in Disease. Int. J. Mol. Sci. 2020, 21, 3371. [Google Scholar] [CrossRef]
- Crosbie, R.H.; Miller, C.; Cheung, P.; Goodnight, T.; Muhlrad, A.; Reisler, E. Structural connectivity in actin: Effect of C-terminal modifications on the properties of actin. Biophys. J. 1994, 67, 1957–1964. [Google Scholar] [CrossRef]
- Strzelecka-Gołaszewska, H.; Mossakowska, M.; Woźniak, A.; Moraczewska, J.; Nakayama, H. Long-range conformational effects of proteolytic removal of the last three residues of actin. Biochem. J. 1995, 307, 527–534. [Google Scholar] [CrossRef]
- Moraczewska, J.; Strzelecka-Gołaszewska, H.; Moens, P.D.J.; Dos Remedios, C.G. Structural changes in subdomain 2 of G-actin observed by fluorescence spectroscopy. Biochem. J. 1996, 317, 605–611. [Google Scholar] [CrossRef]
- Kuznetsova, I.; Antropova, O.; Turoverov, K.; Khaitlina, S. Conformational changes in subdomain I of actin induced by proteolytic cleavage within the DNase I-binding loop: Energy transfer from tryptophan to AEDANS. FEBS Lett. 1996, 383, 105–108. [Google Scholar] [CrossRef]
- Eli-Berchoer, L.; Reisler, E.; Muhlrad, A. Structural Implications of the Chemical Modification of Cys10 on Actin. Biophys. J. 2000, 78, 1482–1489. [Google Scholar] [CrossRef][Green Version]
- Dominguez, R.; Holmes, K.C. Actin Structure and Function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef]
- Harris, A.R.; Jreij, P.; Belardi, B.; Joffe, A.M.; Bausch, A.R.; Fletcher, D.A. Biased localization of actin binding proteins by actin filament conformation. Nat. Commun. 2020, 11, 5973. [Google Scholar] [CrossRef]
- Ochala, J.; Ravenscroft, G.; Laing, N.G.; Nowak, K.J. Nemaline Myopathy-Related Skeletal Muscle α-Actin (ACTA1) Mutation, Asp286Gly, Prevents Proper Strong Myosin Binding and Triggers Muscle Weakness. PLoS ONE 2012, 7, e45923. [Google Scholar] [CrossRef]
- Fan, J.; Chan, C.; McNamara, E.L.; Nowak, K.J.; Iwamoto, H.; Ochala, J. Molecular Consequences of the Myopathy-Related D286G Mutation on Actin Function. Front. Physiol. 2018, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Spudich, J.A.; Watt, S. The regulation of rabbit skeletal muscle contraction. I. Biochemical studies of the interaction of the tropomyosin-troponin complex with actin and the proteolytic fragments of myosin. J. Biol. Chem. 1971, 246, 4866–4871. [Google Scholar] [CrossRef]
- Takashi, R. A novel actin label: A fluorescent probe at glutamine-41 and its consequences. Biochemistry 1988, 27, 938–943. [Google Scholar] [CrossRef]
- Cooper, J.A.; Walker, S.B.; Pollard, T.D. Pyrene actin: Documentation of the validity of a sensitive assay for actin polymerization. J. Muscle Res. Cell Motil. 1983, 4, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Houk, T.; Ue, K. The measurement of actin concentration in solution: A comparison of methods. Anal. Biochem. 1974, 62, 66–74. [Google Scholar] [CrossRef]
- Lehrer, S.S.; Nagy, B.; Gergely, J. The binding of Cu2+ to actin without loss of polymerizability: The involvement of the rapidly reacting –SH group. Arch. Biochem. Biophys. 1972, 150, 164–174. [Google Scholar] [CrossRef]
- Kodama, T.; Fukui, K.; Kometani, K. The Initial Phosphate Burst in ATP Hydrolysis by Myosin and Subfragment-1 as Studied by a Modified Malachite Green Method for Determination of Inorganic Phosphate. J. Biochem. 1986, 99, 1465–1472. [Google Scholar] [CrossRef]
- Pantaloni, D.; Carlier, M.F.; Coué, M.; Lal, A.A.; Brenner, S.L.; Korn, E.D. The critical concentration of actin in the presence of ATP increases with the number concentration of filaments and approaches the critical concentration of actin.ADP. J. Biol. Chem. 1984, 259, 6274–6283. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Fluorescence Anisotropy. In Principles of Fluorescence Spectroscopy, 2nd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999; pp. 291–319. [Google Scholar]
- Kouyama, T.; Mihashi, K. Fluorimetry study of N-(1-pyrenyl)iodoacetamide-labelled F-actin. Local structural change of actin protomer both on polymerization and on binding of heavy meromyosin. Eur.J. Biochem. 1981, 114, 33–38. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ATP Hydrolysis (mol/mol actin/h) | ||||
|---|---|---|---|---|
| Actin Species | Polymerizing Salt | N | Measured | Normalized to the Same N |
| Ca-F-actin | 0.1 M KCl | 1 | 0.66 | 0.66 |
| Ca-F-actin | 2 mM CaCl2 | 2.4 | 0.60 | 0.25 |
| Mg-F-actin | 0.1 M KCl | 2.2 | 0.61 | 0.28 |
| Mg-F-actin | 2 mM MgCl2 | 7.4 | 5.73 | 0.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruszczynska-Biegala, J.; Stefan, A.; Kasprzak, A.A.; Dobryszycki, P.; Khaitlina, S.; Strzelecka-Gołaszewska, H. Myopathy-Sensitive G-Actin Segment 227-235 Is Involved in Salt-Induced Stabilization of Contacts within the Actin Filament. Int. J. Mol. Sci. 2021, 22, 2327. https://doi.org/10.3390/ijms22052327
Gruszczynska-Biegala J, Stefan A, Kasprzak AA, Dobryszycki P, Khaitlina S, Strzelecka-Gołaszewska H. Myopathy-Sensitive G-Actin Segment 227-235 Is Involved in Salt-Induced Stabilization of Contacts within the Actin Filament. International Journal of Molecular Sciences. 2021; 22(5):2327. https://doi.org/10.3390/ijms22052327
Chicago/Turabian StyleGruszczynska-Biegala, Joanna, Andrzej Stefan, Andrzej A. Kasprzak, Piotr Dobryszycki, Sofia Khaitlina, and Hanna Strzelecka-Gołaszewska. 2021. "Myopathy-Sensitive G-Actin Segment 227-235 Is Involved in Salt-Induced Stabilization of Contacts within the Actin Filament" International Journal of Molecular Sciences 22, no. 5: 2327. https://doi.org/10.3390/ijms22052327
APA StyleGruszczynska-Biegala, J., Stefan, A., Kasprzak, A. A., Dobryszycki, P., Khaitlina, S., & Strzelecka-Gołaszewska, H. (2021). Myopathy-Sensitive G-Actin Segment 227-235 Is Involved in Salt-Induced Stabilization of Contacts within the Actin Filament. International Journal of Molecular Sciences, 22(5), 2327. https://doi.org/10.3390/ijms22052327