
Aromadendrin Protects Neuronal Cells from Methamphetamine-Induced Neurotoxicity by Regulating Endoplasmic Reticulum Stress and PI3K/Akt/mTOR Signaling Pathway

Abstract
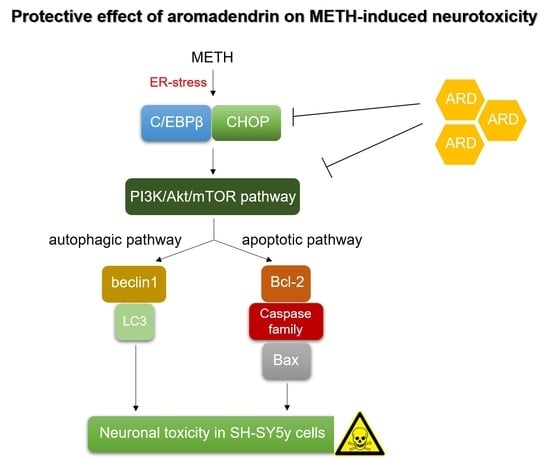
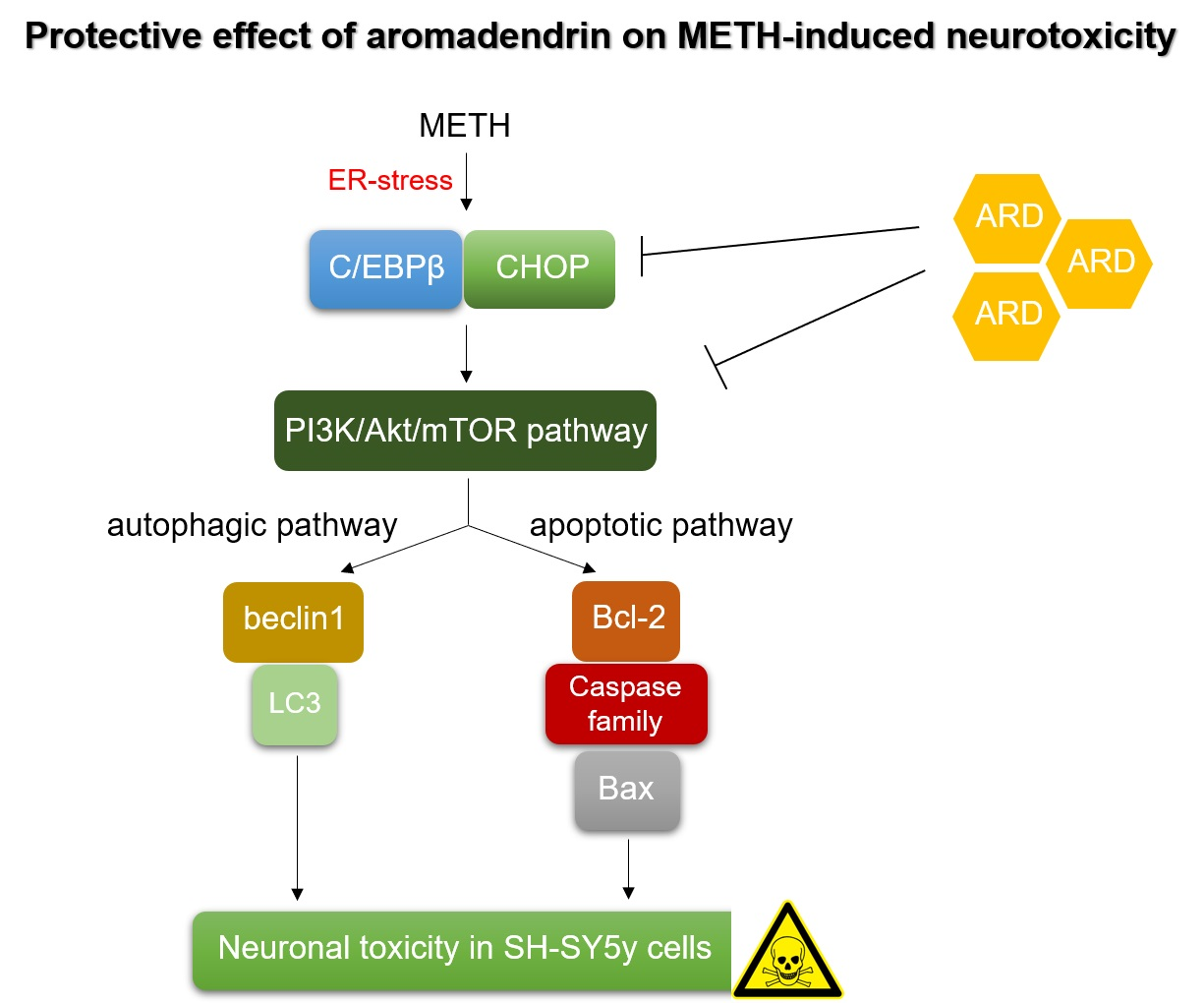{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Aromadendrin Does Not Lead to Neuronal Toxicity in SH-SY5y Cells
2.2. Aromadendrin Suppresses METH-Induced Cell Death in SH-SY5y Cells
2.3. Aromadendrin Regulates METH-Induced ER Stress in SH-SY5y Cells
2.4. Aromadendrin Preserves the Phosphorylation of PI3K/Akt/mTOR Signaling Pathway in METH-Exposed SH-SY5y Cells
2.5. Aromadendrin Mitigates the METH-Induced Autophagy in Neuronal Cells
2.6. Pre-Treatment with Aromadendrin Has an Anti-Apoptotic Effect on METH-Exposed SH-SY5y Cells
2.7. Pre-Treatment with Aromadendrin Modulates the Expression of Apoptosis-Related Proteins in METH-Exposed Condition
2.8. Restored mTOR Phosphorylation by Aromadendrin Protects Cells from Autophagy and Apoptosis
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Isolation of Aromadendrin from C. retusus Flowers
4.3. Reagents and Antibodies
4.4. MTT Assay
4.5. Assessment of Cell Confluency
4.6. AnnexinV/PI Apoptosis Assay by Flow Cytometry
4.7. Real-Time Quantitative PCR
4.8. Western Blot Analysis
4.9. Determination of AnnexinV and Caspase3/7 Expression by IncuCyte® Imaging System
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ARD | Aromadendrin |
| METH | Methamphetamine |
| mTOR | Mammalian target of rapamycin |
| PI | Propidium iodide |
References
- Cadet, J.L.; Krasnova, I.N. Molecular Bases of Methamphetamine-Induced Neurodegeneration. Int. Rev. Neurobiol. 2009, 88, 101–119. [Google Scholar] [PubMed]
- Radfar, S.R.; Rawson, R.A. Current research on methamphetamine: Epidemiology, medical and psychiatric effects, treatment, and harm reduction efforts. Addict. Heal. 2014, 6, 146–154. [Google Scholar]
- Kanthasamy, A.; Anantharam, V.; Ali, S.F.; Kathasamy, A.G. Methamphetamine induces autophagy and apoptosis in a mesencephalic dopaminergic neuronal culture model: Role of cathepsin-D in methamphetamine- induced apoptotic cell death. Ann. NY Acad. Sci. 2006, 1074, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Shukla, M.; Vincent, B. The multi-faceted impact of methamphetamine on Alzheimer’s disease: From a triggering role to a possible therapeutic use. Ageing Res. Rev. 2020, 60. [Google Scholar] [CrossRef]
- Granado, N.; Ares-Santos, S.; Moratalla, R. Methamphetamine and parkinson’s disease. Parkinsons. Dis. 2013, 2013, 308052. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy pathway: Cellular and molecular mechanisms. Autophagy 2018, 14, 207–215. [Google Scholar] [CrossRef]
- Barbosa, D.J.; Capela, J.P.; Feio-Azevedo, R.; Teixeira-Gomes, A.; Bastos, M.d.L.; Carvalho, F. Mitochondria: Key players in the neurotoxic effects of amphetamines. Arch. Toxicol. 2015, 89, 1695–1725. [Google Scholar] [CrossRef]
- Jayanthi, S.; McCoy, M.T.; Beauvais, G.; Ladenheim, B.; Gilmore, K.; Wood, W.; Becker, K.; Cadet, J.L. Methamphetamine Induces Dopamine D1 Receptor-Dependent Endoplasmic Reticulum Stress-Related Molecular Events in the Rat Striatum. PLoS ONE 2009, 4, e6092. [Google Scholar] [CrossRef]
- Beauvais, G.; Atwell, K.; Jayanthi, S.; Ladenheim, B.; Cadet, J.L. Involvement of dopamine receptors in binge Methamphetamine-Induced activation of endoplasmic reticulum and mitochondrial stress pathways. PLoS ONE 2011, 6, e28946. [Google Scholar] [CrossRef]
- Xu, X.; Huang, E.; Luo, B.; Cai, D.; Zhao, X.; Luo, Q.; Jin, Y.; Chen, L.; Wang, Q.; Liu, C.; et al. Methamphetamine exposure triggers apoptosis and autophagy in neuronal cells by activating the C/EBPb-related signaling pathway. FASEB J. 2018, 32, 6737–6759. [Google Scholar] [CrossRef]
- Xu, X.; Huang, E.; Tai, Y.; Zhao, X.; Chen, X.; Chen, C.; Chen, R.; Liu, C.; Lin, Z.; Wang, H.; et al. Nupr1 modulates methamphetamine-induced dopaminergic neuronal apoptosis and autophagy through CHOP-Trib3-mediated endoplasmic reticulum stress signaling pathway. Front. Mol. Neurosci. 2017, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Lutskii, V.I.; Gromova, A.S.; Tyukavkina, N.A. Aromadendrin, apigenin, and kaempferol from the wood of Pinus sibirica. Chem. Nat. Compd. 1971, 7, 197–198. [Google Scholar] [CrossRef]
- Binutu, O.A.; Cordell, G.A. Constituents of Afzelia bella stem bark. Phytochemistry 2001, 56, 827–830. [Google Scholar] [CrossRef]
- Kwak, J.H.; Kang, M.W.; Roh, J.H.; Choi, S.U.; Zee, O.P. Cytotoxic phenolic compounds from Chionanthus retusus. Arch. Pharm. Res. 2009, 32, 1681–1687. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Kim, S.; Lee, S.J.; Ham, I.; Whang, W.K. Antioxidant activities of new flavonoids from Cudrania tricuspidata root bark. Arch. Pharm. Res. 2009, 32, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Tran, M.H.; Phuong, T.P.; Tran, M.N.; Min, B.S.; Song, K.S.; Yeon, H.S.; Bae, K.H. Anti-inflammatory activity of flavonoids from Populus davidiana. Arch. Pharm. Res. 2006, 29, 1102–1108. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Lee, J.J.; Kim, I.S.; Kim, Y.; Myung, C.S. Stimulation of glucose uptake and improvement of insulin resistance by aromadendrin. Pharmacology 2011, 88, 266–274. [Google Scholar] [CrossRef]
- Lee, H.-S.; Jeong, G.-S. Aromadendrin Inhibits T Cell Activation via Regulation of Calcium Influx and NFAT Activity. Molecules 2020, 25, 4590. [Google Scholar] [CrossRef]
- Yang, G.; Zeng, X.; Li, J.; Leung, C.-K.; Zhang, D.; Hong, S.; He, Y.; Huang, J.; Li, L.; Li, Z. Protective effect of gastrodin against methamphetamine-induced autophagy in human dopaminergic neuroblastoma SH-SY5Y cells via the AKT/mTOR signaling pathway. Neurosci. Lett. 2019, 707, 134287. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kim, E.-N.; Jeong, G.-S. Lupenone Protects Neuroblastoma SH-SY5y Cells Against Methamphetamine-Induced Apoptotic Cell Death via PI3K/Akt/mTOR Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 1617. [Google Scholar] [CrossRef]
- Wongprayoon, P.; Govitrapong, P. Melatonin Protects SH-SY5Y Neuronal Cells Against Methamphetamine-Induced Endoplasmic Reticulum Stress and Apoptotic Cell Death. Neurotox. Res. 2017, 31, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hossini, A.M.; Quast, A.S.; Plötz, M.; Grauel, K.; Exner, T.; Küchler, J.; Stachelscheid, H.; Eberle, J.; Rabien, A.; Makrantonaki, E.; et al. PI3K/AKT signaling pathway is essential for survival of induced pluripotent stem cells. PLoS ONE 2016, 11, e0154770. [Google Scholar] [CrossRef] [PubMed]
- Ersahin, T.; Tuncbag, N.; Cetin-Atalay, R. The PI3K/AKT/mTOR interactive pathway. Mol. Biosyst. 2015, 11, 1946–1954. [Google Scholar] [CrossRef]
- Liu, H.Q.; An, Y.W.; Hu, A.Z.; Li, M.H.; Wu, J.L.; Liu, L.; Shi, Y.; Cui, G.H.; Chen, Y. Critical roles of the PI3K-Akt-mTOR signaling pathway in apoptosis and autophagy of astrocytes induced by methamphetamine. Open Chem. 2019, 17, 96–104. [Google Scholar] [CrossRef]
- Pan, A.L.; Hasalliu, E.; Hasalliu, M.; Angulo, J.A. Epigallocatechin Gallate Mitigates the Methamphetamine-Induced Striatal Dopamine Terminal Toxicity by Preventing Oxidative Stress in the Mouse Brain. Neurotox. Res. 2020, 37, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.H.; Zhang, K.K.; Xu, J.T.; Qu, D.; Chen, L.J.; Li, J.H.; Wang, Q.; Wang, H.J.; Xie, X.L. Luteolin alleviates methamphetamine-induced neurotoxicity by suppressing PI3K/Akt pathway-modulated apoptosis and autophagy in rats. Food Chem. Toxicol. 2020, 137, 111179. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Lee, J.H.; Seo, Y.H.; Jang, J.H.; Jeong, C.H.; Lee, S.; Jeong, G.S.; Park, B. Epicatechin prevents methamphetamine-induced neuronal cell death via inhibition of er stress. Biomol. Ther. 2019, 27, 145–151. [Google Scholar] [CrossRef]
- Hetz, C.; Chevet, E.; Harding, H.P. Targeting the unfolded protein response in disease. Nat. Rev. Drug Discov. 2013, 12, 703–719. [Google Scholar] [CrossRef]
- Jayanthi, S.; Deng, X.H.; Noailles, P.; Ladenheim, B.; Lud Cadet, J. Methamphetamine induces neuronal apoptosis via cross-talks between endoplasmic reticulum and mitochondria-dependent death cascades. FASEB J. 2004, 18, 238–251. [Google Scholar] [CrossRef]
- Hayashi, T.; Justinova, Z.; Hayashi, E.; Cormaci, G.; Mori, T.; Tsai, S.Y.; Barnes, C.; Goldberg, S.R.; Su, T.P. Regulation of σ-1 receptors and endoplasmic reticulum chaperones in the brain of methamphetamine self-administering rats. J. Pharmacol. Exp. Ther. 2010, 332, 1054–1063. [Google Scholar] [CrossRef]
- Qie, X.; Wen, D.; Guo, H.; Xu, G.; Liu, S.; Shen, Q.; Liu, Y.; Zhang, W.; Cong, B.; Ma, C. Endoplasmic Reticulum Stress Mediates Methamphetamine-Induced Blood–Brain Barrier Damage. Front. Pharmacol. 2017, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.G.; Jung, J.W.; Lee, H.; Seo, K.H.; Lee, D.Y.; Kim, H.G.; Ko, J.H.; Lee, D.S.; Baek, N.I. Flavonoids from Chionanthus retusus (Oleaceae) Flowers and Their Protective Effects against Glutamate-Induced Cell Toxicity in HT22 Cells. Int. J. Mol. Sci. 2019, 20, 3517. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Yu, J.S.; Kim, K.H.; Jeong, G.-S. Diketoacetonylphenalenone, Derived from Hawaiian Volcanic Soil-Associated Fungus Penicillium herquei FT729, Regulates T Cell Activation via Nuclear Factor-κB and Mitogen-Activated Protein Kinase Pathway. Molecules 2020, 25, 5374. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-S.; Kim, E.-N.; Jeong, G.-S. Aromadendrin Protects Neuronal Cells from Methamphetamine-Induced Neurotoxicity by Regulating Endoplasmic Reticulum Stress and PI3K/Akt/mTOR Signaling Pathway. Int. J. Mol. Sci. 2021, 22, 2274. https://doi.org/10.3390/ijms22052274
Lee H-S, Kim E-N, Jeong G-S. Aromadendrin Protects Neuronal Cells from Methamphetamine-Induced Neurotoxicity by Regulating Endoplasmic Reticulum Stress and PI3K/Akt/mTOR Signaling Pathway. International Journal of Molecular Sciences. 2021; 22(5):2274. https://doi.org/10.3390/ijms22052274
Chicago/Turabian StyleLee, Hyun-Su, Eun-Nam Kim, and Gil-Saeng Jeong. 2021. "Aromadendrin Protects Neuronal Cells from Methamphetamine-Induced Neurotoxicity by Regulating Endoplasmic Reticulum Stress and PI3K/Akt/mTOR Signaling Pathway" International Journal of Molecular Sciences 22, no. 5: 2274. https://doi.org/10.3390/ijms22052274
APA StyleLee, H.-S., Kim, E.-N., & Jeong, G.-S. (2021). Aromadendrin Protects Neuronal Cells from Methamphetamine-Induced Neurotoxicity by Regulating Endoplasmic Reticulum Stress and PI3K/Akt/mTOR Signaling Pathway. International Journal of Molecular Sciences, 22(5), 2274. https://doi.org/10.3390/ijms22052274