
Increased Serum Levels of Brain-Derived Neurotrophic Factor Contribute to Inflammatory Responses in Patients with Rheumatoid Arthritis



Abstract
1. Introduction
2. Results
2.1. Serum BDNF, proBDNF Levels, and proBDNF/BDNF Ratio in Patients with RA and Controls
2.2. Correlation of Clinical Parameters with Serum BDNF Levels in Patients with RA
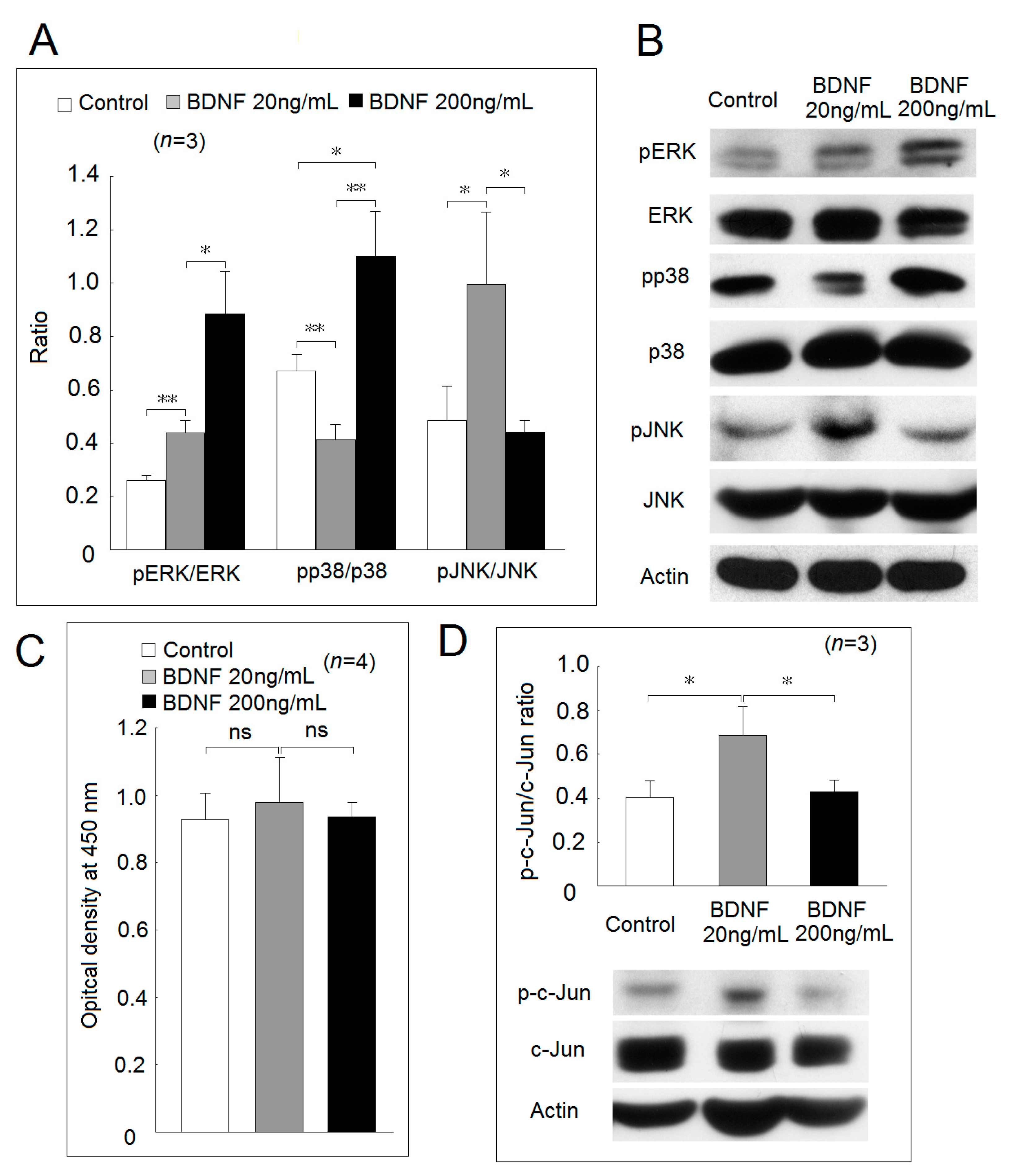
2.3. Expression of BDNF Receptors and Effects of BDNF in Mitogen-Activated Protein Kinase Phosphorylation and Cell Proliferation
2.4. Investigation of the BDNF-Regulated Gene Focusing on Inflammation-Related Proteins and Their Expression in Patients with RA
2.5. Correlation of Clinical Parameters with Expression Level of BDNF-Regulated Genes or BDNF Receptor-NGFR in T Cells from Patients with RA
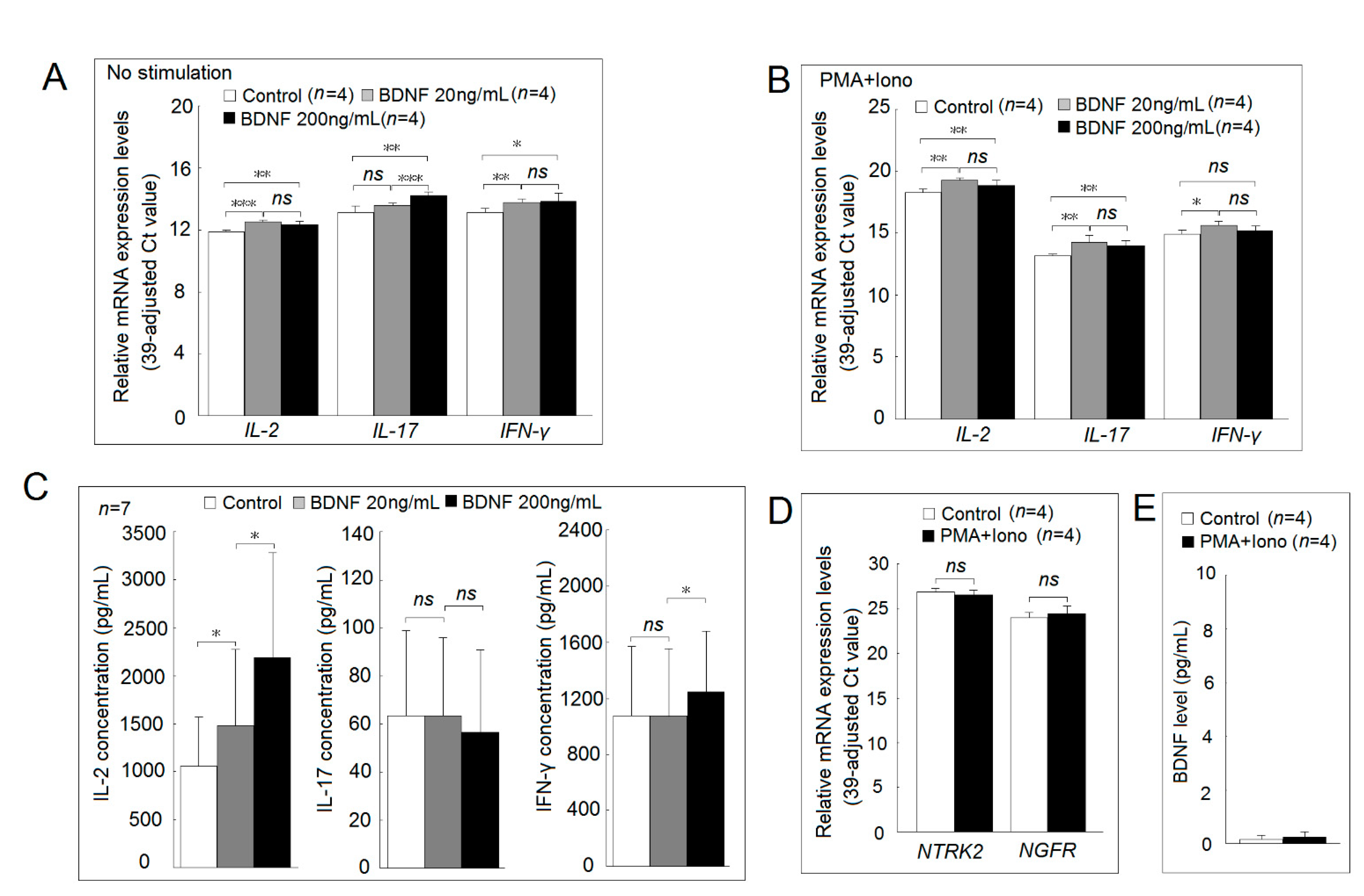
2.6. Functional Studies of BDNF and Its Receptor in Jurkat Cells and Normal PBMCs
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Flow Cytometry Analysis
4.3. Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. Western Blot Analysis
4.5. Cell Viability and Proliferation Using the Mitochondrial Dehydrogenase Cleavage Assay
4.6. Microarray Analysis
4.7. Measurement of mRNA Expression Levels by Real-Time Reverse Transcription–Polymerase Chain Reaction (RT–PCR)
4.8. Effect of BDNF on Proinflammatory Cytokines Secretion in Activated Peripheral Blood Mononuclear Cells (PBMCs) from Healthy Individuals
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BDNF | Brain-derived neurotrophic factor |
| CAMK2A | Calcium/calmodulin-dependent protein kinase II alpha |
| CD40 | Cluster of differentiation 40 |
| ELISA | Enzyme-linked immunosorbent assay |
| ERK | Extracellular signal-regulated kinase |
| GNG13 | G protein subunit gamma 13 |
| IFN-γ | Interferon-gamma |
| IL | Interleukin |
| Iono | Ionomycin |
| JNK | c-Jun N-terminal kinase |
| MASP2 | Mannan-binding lectin serine protease 2 |
| MUC5AC | Oligomeric mucus/gel-forming |
| NGFR | Nerve growth factor receptor |
| NTRK2 | Neurotrophic receptor tyrosine kinase 2 |
| PBMCs | Peripheral blood mononuclear cells |
| PMA | Phorbol 12-myristate 13-acetate |
| PVDF | Polyvinylidene difluoride |
| RA | Rheumatoid arthritis |
| RT–PCR | Real-time reverse transcription–polymerase chain reaction |
| STARD13 | Star-related lipid transfer domain-containing 13 |
References
- Lu, M.-C.; Guo, H.-R.; Lin, M.-C.; Livneh, H.; Lai, N.-S.; Tsai, T.-Y. Bidirectional associations between rheumatoid arthritis and depression: A nationwide longitudinal study. Sci. Rep. 2016, 6, 20647. [Google Scholar] [CrossRef]
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- Yang, J.; Siao, C.-J.; Nagappan, G.; Marinic, T.; Jing, D.; McGrath, K.; Chen, Z.-Y.; Mark, W.; Tessarollo, L.; Lee, F.-S.; et al. Neuronal release of proBDNF. Nat. Neurosci. 2009, 12, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Zhang, C.; Chen, J.; Su, Y.; Zhou, R.; Wang, F.; Xia, W.; Huang, J.; Wang, Z.; Hu, Y.; et al. Ratio of mBDNF to proBDNF for differential diagnosis of major depressive disorder and bipolar depression. Mol. Neurobiol. 2017, 54, 5573–5582. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xiong, J.; Lim, Y.; Ruan, Y.; Huang, C.; Zhu, Y.; Zhong, J.-H.; Xiao, Z.; Zhou, X.F. Upregulation of blood proBDNF and its receptors in major depression. J. Affect. Disord. 2013, 150, 776–784. [Google Scholar] [CrossRef]
- Cheon, Y.-H.; Lee, S.-G.; Kim, M.; Kim, H.-O.; Suh, Y.S.; Park, K.-S.; Kim, R.-B.; Yang, H.-S.; Kim, J.-M.; Son, C.-N.; et al. The association of disease activity, pro-inflammatory cytokines, and neurotrophic factors with depression in patients with rheumatoid arthritis. Brain Behav. Immun. 2018, 73, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Grimsholm, O.; Rantapää-Dahlqvist, S.; Dalén, T.; Forsgren, S. BDNF in RA: Downregulated in plasma following anti-TNF treatment but no correlation with inflammatory parameters. Clin. Rheumatol. 2008, 27, 1289–1297. [Google Scholar] [CrossRef]
- Huang, C.-C.; Chiou, C.-H.; Liu, S.-C.; Hu, S.-L.; Su, C.-M.; Tsai, C.-H.; Tang, C.-H. Melatonin attenuates TNF-α and IL-1β expression in synovial fibroblasts and diminishes cartilage degradation: Implications for the treatment of rheumatoid arthritis. J. Pineal Res. 2019, 66, e12560. [Google Scholar] [CrossRef]
- MacDonald, I.J.; Huang, C.-C.; Liu, S.-C.; Tang, C.-H. Reconsidering the role of melatonin in rheumatoid arthritis. Int. J. Mol. Sci. 2020, 21, 2877. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.-J.; Huang, K.-Y.; Tung, C.-H.; Hsu, B.-B.; Wu, C.-H.; Lu, M.-C.; Lai, N.-S. Risk factors, including different biologics, associated with depression and anxiety in patients with rheumatoid arthritis: A cross-sectional observational study. Clin. Rheumatol. 2020, 39, 737–746. [Google Scholar] [CrossRef]
- Suliman, S.; Hemmings, S.M.; Seedat, S. Brain-Derived neurotrophic factor (BDNF) protein levels in anxiety disorders: Systematic review and meta-regression analysis. Front. Integr. Neurosci. 2013, 7, 55. [Google Scholar] [CrossRef]
- Pluchino, N.; Russo, M.; Santoro, A.N.; Litta, P.; Cela, V.; Genazzani, A.R. Steroid hormones and BDNF. Neuroscience 2013, 239, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lombès, M.; Le Menuet, D. Glucocorticoid receptor represses brain-derived neurotrophic factor expression in neuron-like cells. Mol. Brain 2017, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Alten, R.; Wiebe, E. Hypothalamic-pituitary-adrenal axis function in patients with rheumatoid arthritis treated with different glucocorticoid approaches. Neuroimmunomodulation 2015, 22, 83–88. [Google Scholar] [CrossRef]
- Katoh-Semba, R.; Wakako, R.; Komori, T.; Shigemi, H.; Miyazaki, N.; Ito, H.; Kumagai, T.; Tsuzuki, M.; Shigemi, K.; Yoshida, F.; et al. Age-Related changes in BDNF protein levels in human serum: Differences between autism cases and normal controls. Int. J. Dev. Neurosci. 2007, 25, 367–372. [Google Scholar] [CrossRef]
- Raychaudhuri, S.P.; Raychaudhuri, S.K.; Atkuri, K.R.; Herzenberg, L.A. Nerve growth factor: A key local regulator in the pathogenesis of inflammatory arthritis. Arthritis Rheum. 2011, 63, 3243–3252. [Google Scholar] [CrossRef]
- Aloe, L.; Tuveri, M.A.; Carcassi, U.; Levi-Montalcini, R. Nerve growth factor in the synovial fluid of patients with chronic arthritis. Arthritis Rheum. 1992, 35, 351–355. [Google Scholar] [CrossRef]
- Manni, L.; Lundeberg, T.; Fiorito, S.; Bonini, S.; Vigneti, E.; Aloe, L. Nerve growth factor release by human synovial fibroblasts prior to and following exposure to tumor necrosis factor-alpha, interleukin-1 beta and cholecystokinin-8: The possible role of NGF in the inflammatory response. Clin. Exp. Rheumatol. 2003, 21, 617–624. [Google Scholar]
- Hashimoto, K.; Shimizu, E.; Iyo, M. Critical role of brain-derived neurotrophic factor in mood disorders. Brain Res. Rev. 2004, 45, 104–114. [Google Scholar] [CrossRef]
- Kerschensteiner, M.; Gallmeier, E.; Behrens, L.; Leal, V.V.; Misgeld, T.; Klinkert, W.E.; Kolbeck, R.; Hoppe, E.; Oropeza-Wekerle, R.L.; Bartke, I.; et al. Activated human T cells, B cells, and monocytes produce brain-derived neurotrophic factor in vitro and in inflammatory brain lesions: A neuroprotective role of inflammation? J. Exp. Med. 1999, 189, 865–870. [Google Scholar] [CrossRef] [PubMed]
- De Santi, L.; Annunziata, P.; Sessa, E.; Bramanti, P. Brain-Derived neurotrophic factor and TrkB receptor in experimental autoimmune encephalomyelitis and multiple sclerosis. J. Neurol. Sci. 2009, 287, 17–26. [Google Scholar] [CrossRef]
- Barthel, C.; Yeremenko, N.; Jacobs, R.; Schmidt, R.E.; Bernateck, M.; Zeidler, H.; Tak, P.P.; Baeten, D.; Rihl, M. Nerve growth factor and receptor expression in rheumatoid arthritis and spondyloarthritis. Arthritis Res. Ther. 2009, 11, R82. [Google Scholar] [CrossRef] [PubMed]
- Tejeda, G.S.; Díaz-Guerra, M. Integral characterization of defective BDNF/TrkB signalling in neurological and psychiatric disorders leads the way to new therapies. Int. J. Mol. Sci. 2017, 18, 268. [Google Scholar] [CrossRef]
- Liang, J.; Deng, G.; Huang, H. The activation of BDNF reduced inflammation in a spinal cord injury model by TrkB/p38 MAPK signaling. Exp. Ther. Med. 2019, 17, 1688–1696. [Google Scholar] [CrossRef]
- Numakawa, T.; Suzuki, S.; Kumamaru, E.; Adachi, N.; Richards, M.; Kunugi, H. BDNF function and intracellular signaling in neurons. Histol. Histopathol. 2010, 25, 237–258. [Google Scholar] [PubMed]
- Bothwell, M. Recent advances in understanding context-dependent mechanisms controlling neurotrophin signaling and function. F1000Research 2019, 8. [Google Scholar] [CrossRef]
- Raychaudhuri, S.P.; Raychaudhuri, S.K. The regulatory role of nerve growth factor and its receptor system in fibroblast-like synovial cells. Scand. J. Rheumatol. 2009, 38, 207–215. [Google Scholar] [CrossRef]
- del Porto, F.; Aloe, L.; Laganà, B.; Triaca, V.; Nofroni, I.; D’Amelio, R. Nerve growth factor and brain-derived neurotrophic factor levels in patients with rheumatoid arthritis treated with TNF-alpha blockers. Ann. N. Y. Acad. Sci. 2006, 1069, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.R.; Maiti, R.; Nath, S.; Sahoo, P.; Jena, M.; Mishra, A. Effect of sertraline, dosulepin, and venlafaxine on Non-BDNF neurotrophins in patients with depression: A cohort study. J. Clin. Psychopharmacol. 2019, 39, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Sheng, W.; Han, L.; He, Q.; Ji, X.; Liu, K. Activation of BDNF-TrkB signaling pathway-regulated brain inflammation in pentylenetetrazole-induced seizures in zebrafish. Fish Shellfish Immunol. 2018, 83, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Mattingly, A.; Lin, A.; Sacramento, J.; Mannent, L.; Castel, M.N.; Canolle, B.; Delbary-Gossart, S.; Ferzaz, B.; Morganti, J.M.; et al. A novel antagonist of p75NTR reduces peripheral expansion and CNS trafficking of pro-inflammatory monocytes and spares function after traumatic brain injury. J. Neuroinflamm. 2016, 13, 88. [Google Scholar] [CrossRef]
- Jin, Y.; Sun, L.-H.; Yang, W.; Cui, R.-J.; Xu, S.-B. The role of BDNF in the neuroimmune axis regulation of mood disorders. Front. Neurol. 2019, 10, 515. [Google Scholar] [CrossRef]
- Fauchais, A.L.; Boumediene, A.; Lalloue, F.; Gondran, G.; Loustaud-Ratti, V.; Vidal, E.; Jauberteau, M.O. Brain-Derived neurotrophic factor and nerve growth factor correlate with T-cell activation in primary Sjogren’s syndrome. Scand. J. Rheumatol. 2009, 38, 50–57. [Google Scholar] [CrossRef]
- Hsieh, M.-C.; Hsu, C.-W.; Lu, M.-C.; Koo, M. Increased risks of psychiatric disorders in patients with primary Sjogren’s syndrome-a secondary cohort analysis of nationwide, population-based health claim data. Clin. Rheumatol. 2019, 38, 3195–3203. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Chen, Z.-Y. The role of BDNF in depression on the basis of its location in the neural circuitry. Acta Pharmacol. Sin. 2011, 32, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 rheumatoid arthritis classification criteria: An American College of Rheumatology/European league against rheumatism collaborative initiative. Ann. Rheum. Dis. 2010, 69, 1580–1588. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.-J.; Huang, K.-Y.; Tung, C.-H.; Hsu, B.-B.; Wu, C.-H.; Koo, M.; Hsu, C.-W.; Lu, M.-C.; Lai, N.-S. Modified rheumatoid arthritis impact of disease (RAID) score, a potential tool for depression and anxiety screening for rheumatoid arthritis. Jt. Bone Spine 2019, 86, 805–807. [Google Scholar] [CrossRef]
- Zigmond, A.S.; Snaith, R.P. The hospital anxiety and depression scale. Acta Psychiatr. Scand. 1983, 67, 361–370. [Google Scholar] [CrossRef]
- Lu, M.-C.; Lai, N.-S.; Chen, H.-C.; Yu, H.-C.; Huang, K.-Y.; Tung, C.-H.; Huang, H.-B.; Yu, C.-L. Decreased microRNA(miR)-145 and increased miR-224 expression in T cells from patients with systemic lupus erythematosus involved in lupus immunopathogenesis. Clin. Exp. Immunol. 2013, 171, 91–99. [Google Scholar] [CrossRef]
- Lu, M.-C.; Lai, N.-S.; Yin, W.-Y.; Yu, H.-C.; Huang, H.-B.; Tung, C.-H.; Huang, K.-Y.; Yu, C.-L. Anti-Citrullinated protein antibodies activated ERK1/2 and JNK mitogen-activated protein kinases via binding to surface-expressed citrullinated GRP78 on mononuclear cells. J. Clin. Immunol. 2013, 33, 558–566. [Google Scholar] [CrossRef]
- Lai, N.-S.; Yu, C.-L.; Yin, W.-Y.; Yu, H.-C.; Huang, H.-B.; Tung, C.-H.; Lu, M.-C. Combination of nifedipine and subtherapeutic dose of cyclosporin additively suppresses mononuclear cells activation of patients with rheumatoid arthritis and normal individuals via Ca(2+)-calcineurin-nuclear factor of activated T cells pathway. Clin. Exp. Immunol. 2012, 168, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Lai, N.-S.; Yu, H.-C.; Huang, K.-Y.; Tung, C.-H.; Huang, H.-B.; Lu, M.-C. Decreased T cell expression of H/ACA box small nucleolar RNA 12 promotes lupus pathogenesis in patients with systemic lupus erythematosus. Lupus 2018, 27, 1499–1508. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Univariate Regression Analysis | Multiple Regression Analysis | ||||
|---|---|---|---|---|---|---|
| B | (95% CI) | p | B | (95% CI) | p | |
| Depression (yes/no) | 167.23 | (−743.59, 1078.05) | 0.719 | |||
| Anxiety (yes/no) | −1633.44 | (−2701.60, −565.28) | 0.003 | −1660.31 | (−2719.44, −601.18) | 0.002 |
| DAS-28 | −222.15 | (−467.34, 23.04) | 0.076 | |||
| TJC 28 | −22.19 | (−74.37, 29.98) | 0.404 | |||
| SJC 28 | −91.32 | (−169.83, −12.82) | 0.023 | −52.64 | (−131.60, 30.25) | 0.220 |
| ESR (mm per h) | −3.66 | (−24.65, 17.32) | 0.732 | |||
| PGA | −8.56 | (−21.28, 4.15) | 0.187 | |||
| CRP (mg/dL) | −158.45 | (−426.45, 109.56) | 0.247 | |||
| Biologics (yes/no) | −920.07 | (−1620.78, −219.36) | 0.010 | −827.25 | (−1548.76, −147.37) | 0.018 |
| csDMARD (yes/no) | 12.08 | (−1299.53, 1323.70) | 0.986 | |||
| Steroid (yes/no) | −312.66 | (−1088.67, 463.34) | 0.430 | −241.91 | (−1014.94, 531.11)) | 0.540 |
| Female | −84.11 | (−880.21, 712.00) | 0.836 | |||
| Age (year) | −30.08 | (−54.31, −5.86) | 0.015 | −12.34 | (−47.50, 21.04) | 0.449 |
| Educational level | ||||||
| Below high school | Ref | |||||
| High school or above | −33.33 | (−713.61, 646.94) | 0.923 | |||
| Marital status | ||||||
| Single | Ref | |||||
| Married | −728.57 | (−1864.78, 407.64) | 0.209 | |||
| Widowed, divorced or separated | −1144.96 | (−2430.36, 140.44) | 0.081 | |||
| Employment status | ||||||
| Being employed | Ref | |||||
| Unemployed | −601.33 | (−1522.66, 320.01) | 0.201 | −310.42 | (−1214.69, 647.55) | 0.551 |
| Retired | −1072.76 | (−1830.40, −315.12) | 0.006 | −564.53 | (−1554.66, 515.62) | 0.325 |
| Income | ||||||
| High | Ref | |||||
| Median | 309.26 | (−413.22, 1031.73) | 0.401 | |||
| Low | −147.90 | (−1170.72, 874.92) | 0.777 | |||
| Religious belief (yes/no) | −78.16 | (−1568.03, 1411.70) | 0.918 | |||
| Disease duration ≥5 years (yes/no) | −939.35 | (−1767.40, −111.29) | 0.026 | −626.75 | (−1469.76, 218.40) | 0.146 |
| Comorbidities | −453.24 | (−1153.14, 246.66) | 0.204 | |||
| Variable | Healthy Volunteers (N = 20) | Patients with Rheumatoid Arthritis (N = 40) | p |
|---|---|---|---|
| Age (mean years ± SD) | 48.0 ± 6.8 | 50.6 ± 10.2 | 0.144 |
| Sex (F:M) | 15:5 | 31:9 | >0.999 |
| RF positivity | - | 65.7% (26/40) | |
| ACPA positivity | - | 65.7% (23/35) | |
| DAS28-ESR | 3.19 ± 1.05 | ||
| CRP (mg/dL) | - | 0.56 ± 0.79 | |
| Medication | |||
| Corticosteroids | - | 87.5% (35/40) | |
| Salazopyrine | - | 72.5% (29/40) | |
| MTX | - | 80.0% (32/40) | |
| Leflunomide | - | 10.0% (4/40) | |
| Biologics | - | 67.5% (27/40) |
| CAMK2A | MASP2 | GNG13 | MUC5A | NGFR | |
|---|---|---|---|---|---|
| Sex (F/M) | 0.144 (0.791) | 0.581 (0.414) | −0.380 (0.458) | 0.081 (0.841) | 0.493 (0.299) |
| Age (per 10 years) | 0.409 (0.064) | 0.427 (0.144) | 0.549 (0.007) * | 0.029 (0.077) | 0.504 (0.012) * |
| Positivity of RF (yes/no) | 0.293 (0.538) | 0.648 (0.297) | 0.616 (0.165) | −0.077 (0.827) | 0.013 (0.977) |
| CRP (per 1 mg/dL) | −0.071 (0.807) | 0.037 (0.922) | −0.140 (0.610) | 0.082 (0.703) | −0.114 (0.672) |
| DAS28-ESR | −0.031 (0.887) | 0.035 (0.902) | 0.109 (0.597) | 0.011 (0.946) | 0.064 (0.753) |
| Biologic (yes/no) | −0.209 (0.666) | −0.248 (0.696) | 0.236 (0.606) | −0.143 (0.690) | −0.012 (0.979) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, N.-S.; Yu, H.-C.; Huang Tseng, H.-Y.; Hsu, C.-W.; Huang, H.-B.; Lu, M.-C. Increased Serum Levels of Brain-Derived Neurotrophic Factor Contribute to Inflammatory Responses in Patients with Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 1841. https://doi.org/10.3390/ijms22041841
Lai N-S, Yu H-C, Huang Tseng H-Y, Hsu C-W, Huang H-B, Lu M-C. Increased Serum Levels of Brain-Derived Neurotrophic Factor Contribute to Inflammatory Responses in Patients with Rheumatoid Arthritis. International Journal of Molecular Sciences. 2021; 22(4):1841. https://doi.org/10.3390/ijms22041841
Chicago/Turabian StyleLai, Ning-Sheng, Hui-Chun Yu, Hsien-Yu Huang Tseng, Chia-Wen Hsu, Hsien-Bin Huang, and Ming-Chi Lu. 2021. "Increased Serum Levels of Brain-Derived Neurotrophic Factor Contribute to Inflammatory Responses in Patients with Rheumatoid Arthritis" International Journal of Molecular Sciences 22, no. 4: 1841. https://doi.org/10.3390/ijms22041841
APA StyleLai, N.-S., Yu, H.-C., Huang Tseng, H.-Y., Hsu, C.-W., Huang, H.-B., & Lu, M.-C. (2021). Increased Serum Levels of Brain-Derived Neurotrophic Factor Contribute to Inflammatory Responses in Patients with Rheumatoid Arthritis. International Journal of Molecular Sciences, 22(4), 1841. https://doi.org/10.3390/ijms22041841