1. Introduction
Photosynthetic organisms are often challenged by biotic and abiotic stresses resulting in a redox imbalance that must be counteracted by the organism to survive. The redox status of chloroplast proteins is mainly controlled and influenced by photosynthetic light reactions, which can lead to an increased generation of reactive oxygen species (ROS) during stress [
1,
2]. Generation of ROS can occur in response to an imbalance between the excess of excited-state electron populations and the electron carriers. As a result, the excited electrons cannot be dissipated by their electron carriers, eventually leading to the generation of ROS and oxidative damage [
2,
3]. Proteins with cysteine residues and disulfide bridges are sensitive to redox changes that affect the redox status of these bridges leading to structural changes, impaired ability to function and to interact with partner proteins. Thus, the redox status of proteins plays an essential role in cell signaling and antioxidizing defense [
4,
5].
Here we describe a novel DnaJ-like protein, ZnJ6 (Cre06.g251716.t1.2), from
C. reinhardtii. This protein has four cysteine-rich CXXCX(G)X(G) motifs that form two C4-type zinc fingers (ZFs) that can bind two zinc atoms [
6] similar to those of DnaJ and can be categorized as a DnaJ-like protein (DnaJE1). DnaJE1 proteins lack all the other motifs of DnaJ, such as the J and G/F domains, and show no C-terminal homology to DnaJ [
7]. The proteins of this group act independently of DnaK by direct substrate interaction and stabilization [
8]. Furthermore, they do not require ATP fueling to assist in the prevention of aggregation as they bind their substrates directly. This was previously shown for ZnJ2 [
9]. To date, 20 proteins of this family have been identified in
Arabidopsis [
7], but their orthologs in
Chlamydomonas have not yet been determined, mainly due to their limited sequence similarities. Many DnaJ-like proteins function as chaperones and in the assembly of photosynthetic complexes. One example is the Bundle Sheath Defective Gene2 (BSD2) initially identified in maize, which is required for RuBisCO biogenesis [
10,
11]. Other examples are the thylakoid-associated DnaJ-like PSA2 and LQY1 proteins, which interact with components of the PSI [
12] and PSII [
13] complexes, respectively. Many DnaJEs reside in the chloroplast and have a chloroplast associated function [
7,
10,
12,
13].
Here we report that ZnJ6 is found in the chloroplast thylakoid membranes and explore its potential function as a chaperone and its response to changes in the redox environment. The localization of ZnJ6 was confirmed by biochemical subfractionation, followed by western analysis of different fractions using the organelle-specific protein markers. In addition to substrate-binding, we show that the ZF domain is required for protein stability at elevated temperatures. We analyzed the ZnJ6 response to the changing redox environment in the chloroplast using a ZnJ6 insertional mutant with reduced expression. Under heat and reductive stress, the WT strain grew better, indicating that ZnJ6 assists in resistance to these conditions. However, under oxidizing conditions, the mutant grew better than its background WT strain, compatible with our observation that ZnJ6 possesses an oxidizing activity. Furthermore, we show that under normal conditions, ZnJ6 is required for maintaining photosynthetic efficiency.
3. Discussion
We report that the
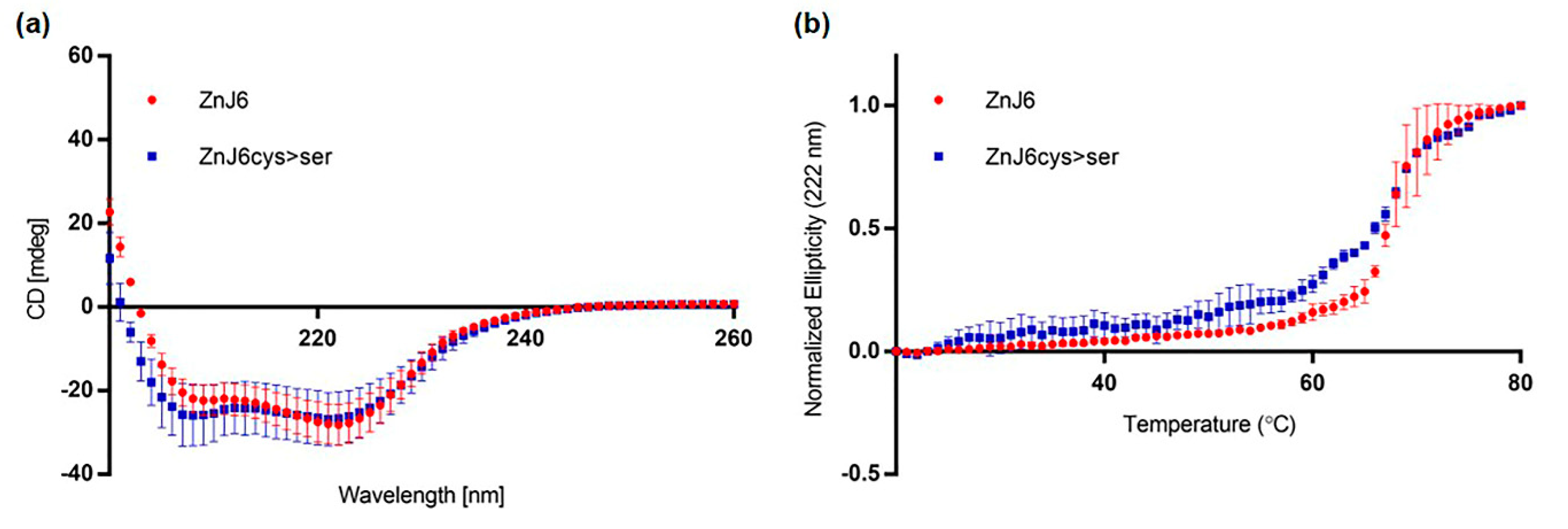
C. reinhardtii DnaJ-like protein ZnJ6 (Cre06.g251716) is associated with the chloroplast thylakoid membrane. We established this using biochemical fractionation assays and western analysis. These experimental results are supported by our primary sequence analysis indicating that ZnJ6 has both a putative chloroplast transit peptide and a transmembrane domain. ZnJ6 is a ZF oxidase that contains four cysteine-rich CXXCX(G)X(G) motifs that form two C4-type ZFs that bind two zinc atoms. These ZFs confer a stable, coordinated structure to the protein at elevated temperatures, as we show by circular dichroism (
Figure 2a,b). However, ZnJ6 has distinct properties and cannot be considered to be the algal ortholog of the DnaJE chaperones of higher plants such as maize BSD2 (GRMZM2G062788_T01) [
9] that is a chloroplast protein but lacks a transmembrane domain [
29,
30].
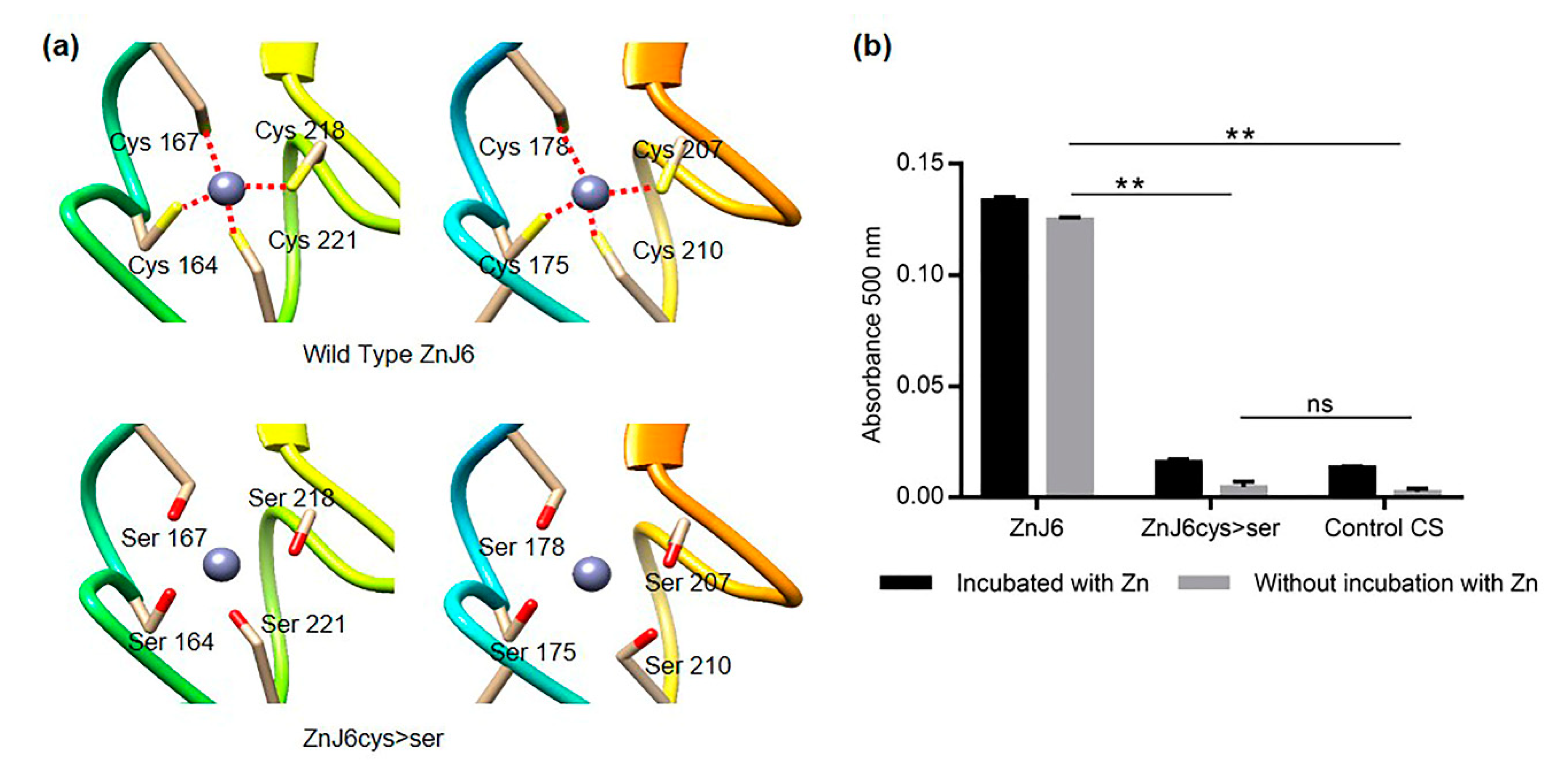
ZF domains are known to be involved in protein–protein interactions and can even contribute to the ability of chaperones to identify denatured substrate proteins [
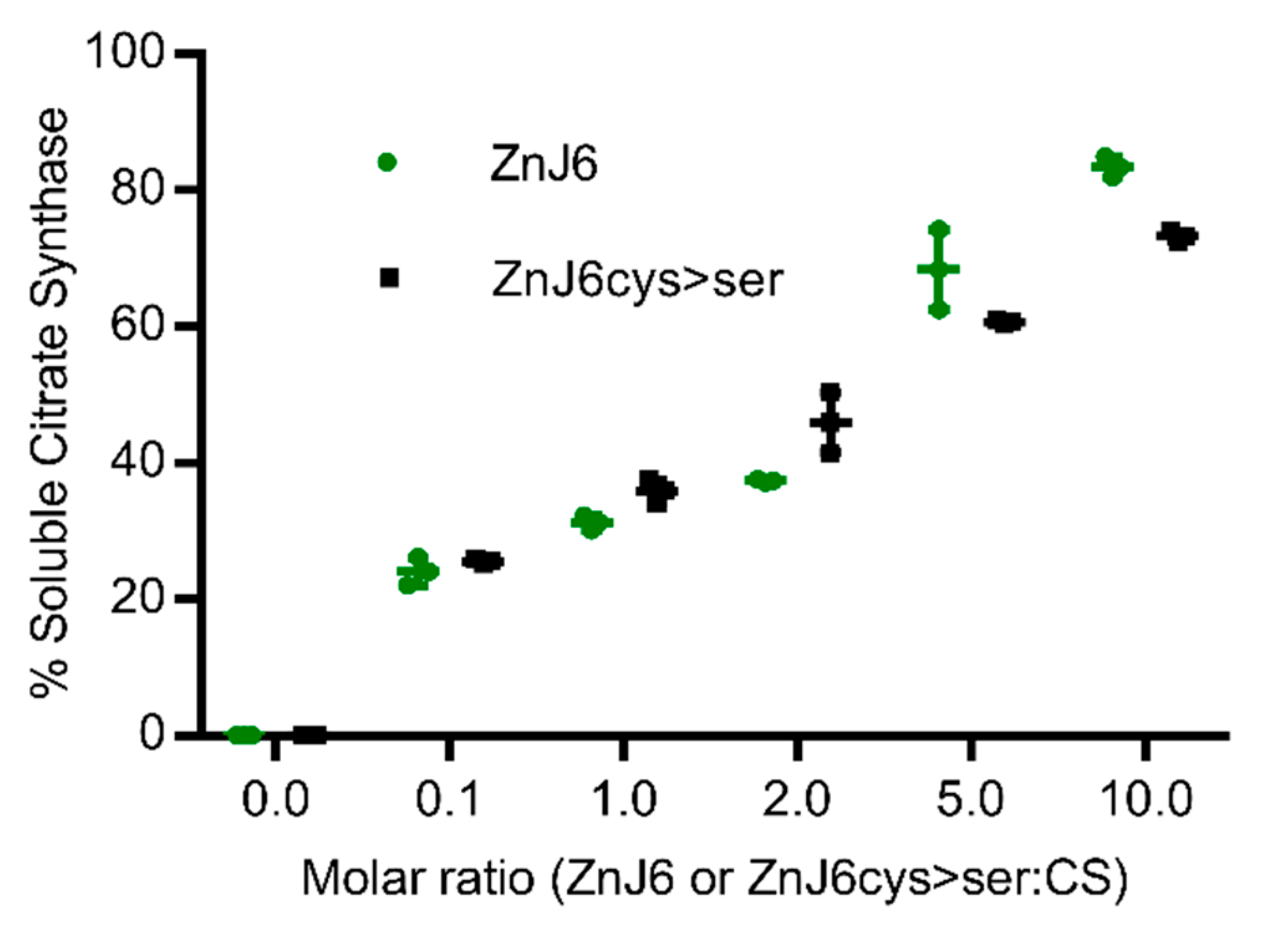
8]. We, therefore, explored the role of the cysteine-rich motif in chaperone activities of ZnJ6 using recombinant proteins (WT ZnJ6 and its ZnJ6cys > Ser-mutant) in classical in vitro chaperone assays. Based on the citrate synthase prevention of aggregation assay, we show that the chaperone “holding activity” of ZnJ6 does not require its Cys-rich domain. However, this domain is required for the redox activity of ZnJ6.
ZnJ6 activity varies from other redox-related chaperones. For example, whereas thioredoxin can induce precipitation of the insulin β-chain [
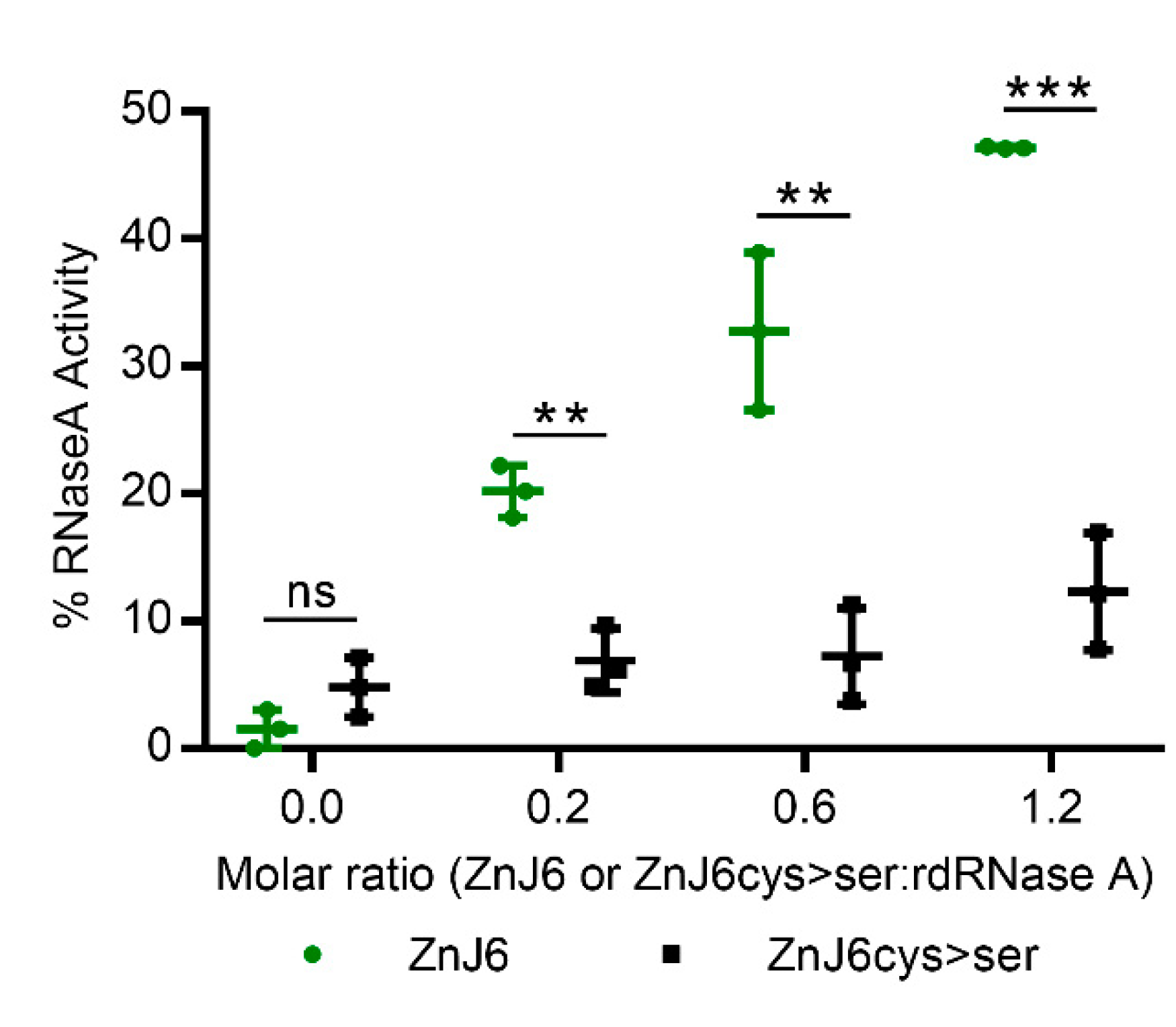
21], ZnJ6 did not have this reducing activity. However, it could protect these chains from precipitation in the presence of a reducing agent such as DTT. This activity required the Cys-rich domain. We, therefore, conclude that ZnJ6 can not reduce disulfide bonds in its target proteins. Overall, this also excludes the possibility that ZnJ6 has protein disulfide isomerization activity (PDI). Based on the RNaseA assay, we show that ZnJ6 has oxidizing activity and assists in stabilizing the spontaneous native folding of rdRNaseA as we do not observe the restoration of activity in the absence of ZnJ6. This activity was dependent on the presence of a functional ZFs. Thus, ZnJ6 is a chaperone that can hold its target to prevent aggregation; it lacks reducing activity but can promote disulfide-bridge formation in its target proteins.
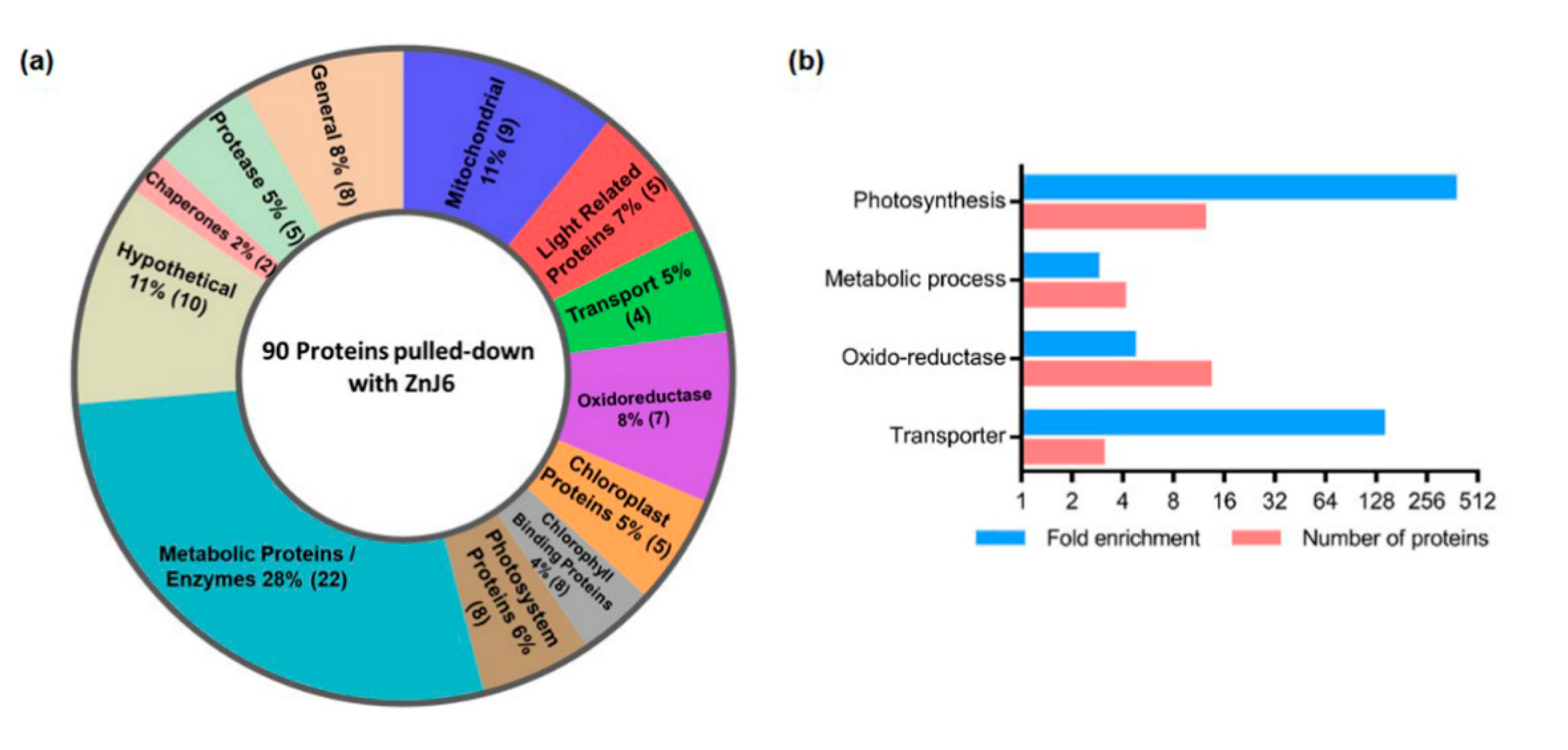
To further expand our general understanding of ZnJ6 protein interactions and in which context they occur, we performed a pull-down assay in which we affinity-purified chloroplast extracts of
Chlamydomonas over immobilized recombinant ZnJ6. ZnJ6 co-purified with the majority of photosynthetic proteins (12), oxidoreductases (13), proteases (5), transporters (4) and chaperones (2), suggesting that it could be responsible for chaperoning a multitude of substrate proteins. However, at this stage, we cannot relate these findings to a direct interaction between these proteins and ZnJ6. The association of ZnJ6 with oxidoreductases could indicate its involvement in maintaining a subcellular redox balance. Co-purification of ZnJ6 with photosynthetic proteins (the majority of photosystem I proteins) could indicate a role in the assembly of the photosynthetic complex, as shown for other DnaJ-like proteins such as PSA2 [
12] and LQY1 [
13]. ZnJ6 localization in the thylakoid membranes could restrict its function to complexes that are formed in the membranes. ZnJ6 could also affect the biosynthesis of thylakoid membrane components as these are also coordinated with the photosynthetic machinery.
We have observed the enrichment of metabolic enzymes in the BLAST2GO analysis of ZnJ6-interacting proteins. This observation is in agreement with growing evidence for dual functions observed for metabolic enzymes, among them RNA-binding activities. Such activities could be related to their moonlighting activities, although other explanations are possible. We previously reported that RuBisCO LSU possesses RNA-binding activity, which is related to its regulation under oxidizing conditions [
31]. The RNA-binding activity of metabolic enzymes was recently expanded to a multitude of enzymes [
32], raising intriguing possibilities for such activity on the part of ZnJ6 [
33].
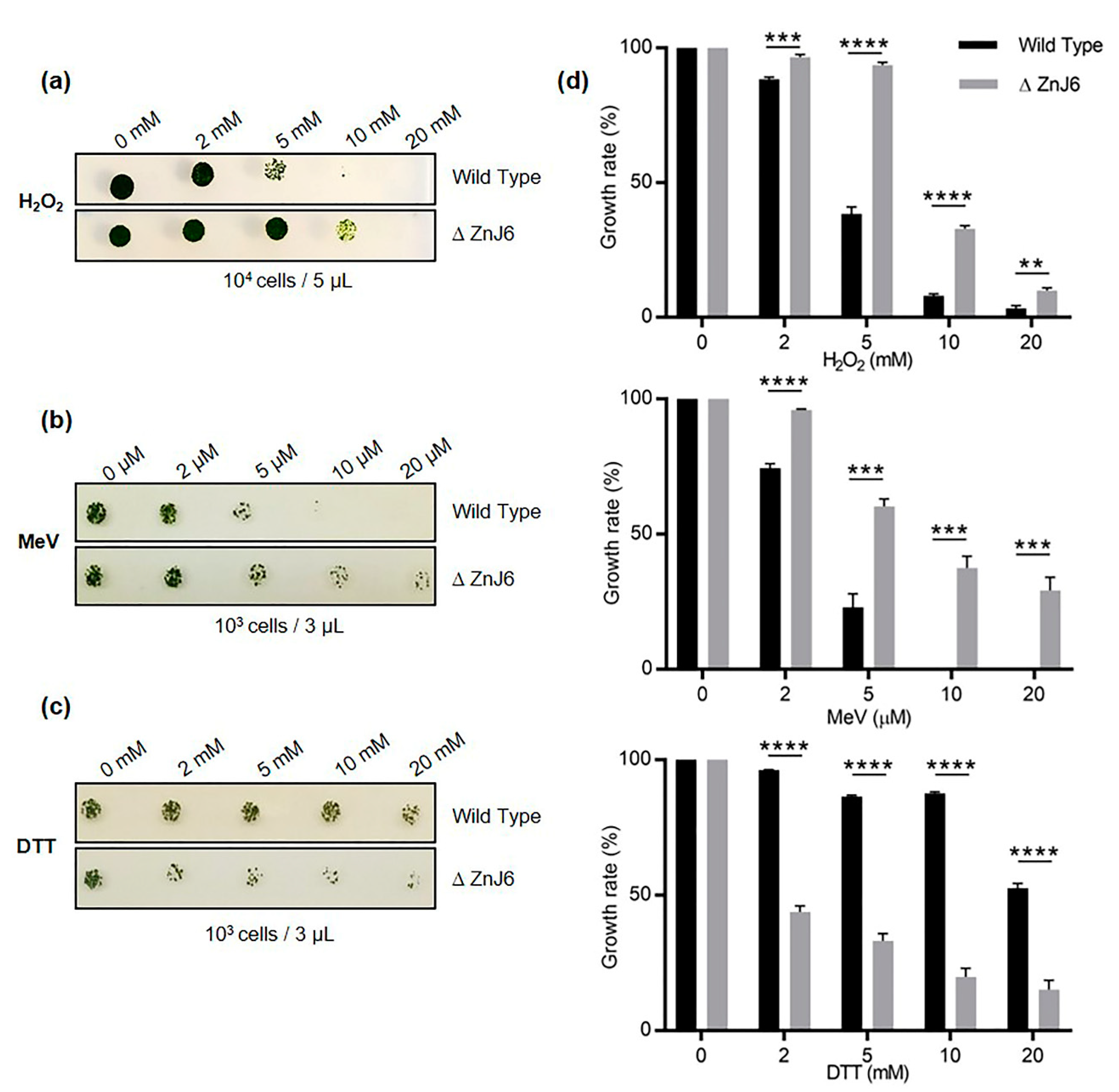
To elaborate on the role of ZnJ6 in redox responses, we performed in vivo experiments monitoring the growth of
C. reinhardtii ZnJ6 knock-down mutant cells that expressed a low level of ZnJ6 in different redox environments. These assays indicate that the ΔZnJ6 mutants are more tolerant to oxidative stress caused by short incubations with H
2O
2 or MeV compared to WT cc-4533 cells. The mechanism behind this activity is not fully resolved, but it could be related to the oxidizing features of ZnJ6. A similar physiological effect was reported for the FtsH5-interacting protein (FIP, At5g02160) in
Arabidopsis. This protein has been found in mosses and higher plants and is another DnaJ-like protein that lacks the typical J-domains [
34]. ZnJ6 and FIP have four and two cysteine-rich motifs, respectively; mutants of both proteins show tolerance to oxidative stress. ZnJ6 associates with an FtsH-like protease as well as with members of the ClpP protease complex. ClpP proteases are involved in the maintenance of chloroplast protein homeostasis [
35,
36]. Furthermore, chloroplast chaperones are known to regulate protease activities and function in synergy to maintain protein quality control [
37]. Therefore, ZnJ6 could be involved in protein quality control and in regulating protein homeostasis.
In contrast to the improved growth of the ΔZnJ6 mutant cells under oxidizing conditions, these cells were more sensitive to a reducing force induced by DTT, showing impaired growth as compared to WT cells. DTT can easily cross the membrane and cause disulfide bond reduction, leading to reductive stress [
22]. We assume that in the presence of DTT, protein structures of the chloroplast proteins could be affected by the reduction of disulfide bridges. Thus, the oxidizing activity of ZnJ6 could restore disulfide bridge formation, thus protecting protein structures in the WT cells. In the absence of ZnJ6, the reductive force of DTT could impair protein structures and possibly lead to their aggregation. Thus, ZnJ6 could provide resistance to the WT cells against DTT induced stress. This hypothesis is supported by the results of the in vitro insulin aggregation assay.
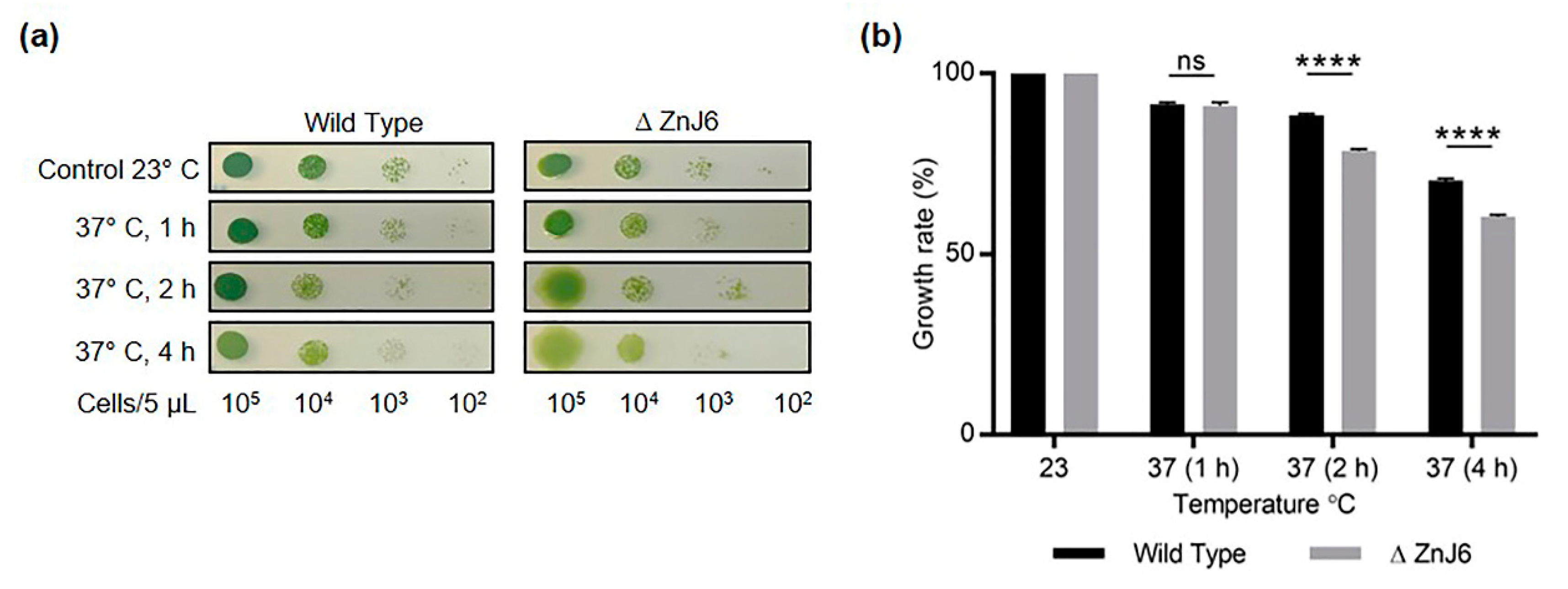
Shifting
Chlamydomonas cells from 25 °C to 37 °C induces a heat stress response [
38]. Our data suggest that ZnJ6 is involved in protection against heat stress since the exposure of the ΔZnJ6 mutant cells to elevated temperatures for 2–4 h resulted in the degradation of chlorophyll and impaired growth. Thus, the mutant cells appear to be much more sensitive to heat stress (37 °C) than WT cells. This could possibly be attributed to the chaperone activity of ZnJ6 under heat stress and is further supported by our finding that ZnJ6 interacts with ClpP proteases involved in the chloroplast unfolded protein response, UPR, and proteostatic processes [
39]. The ability of ZnJ6 to protect a substrate protein from heat-induced aggregation was shown in vitro in the CS assays. ZnJ6 structure is stable up to 65 °C, which would enable such activity. ZnJ6 also interacts with the temperature-sensitive catalytic chaperone RA in our pull-down assays. Unfolding and aggregation of RA occur at 37 °C [
20]. RA is a chloroplast protein responsible for RuBisCO activation and the primary cause of reduced photosynthesis at elevated temperatures due to its temperature sensitivity [
40,
41]. In our in vitro assays, we showed that ZnJ6 prevents the aggregation of RA at 42 °C. This could suggest a role for ZnJ6 in preventing the aggregation of temperature-sensitive proteins in the chloroplast, thus assisting the growth and survival of cells at elevated temperatures.
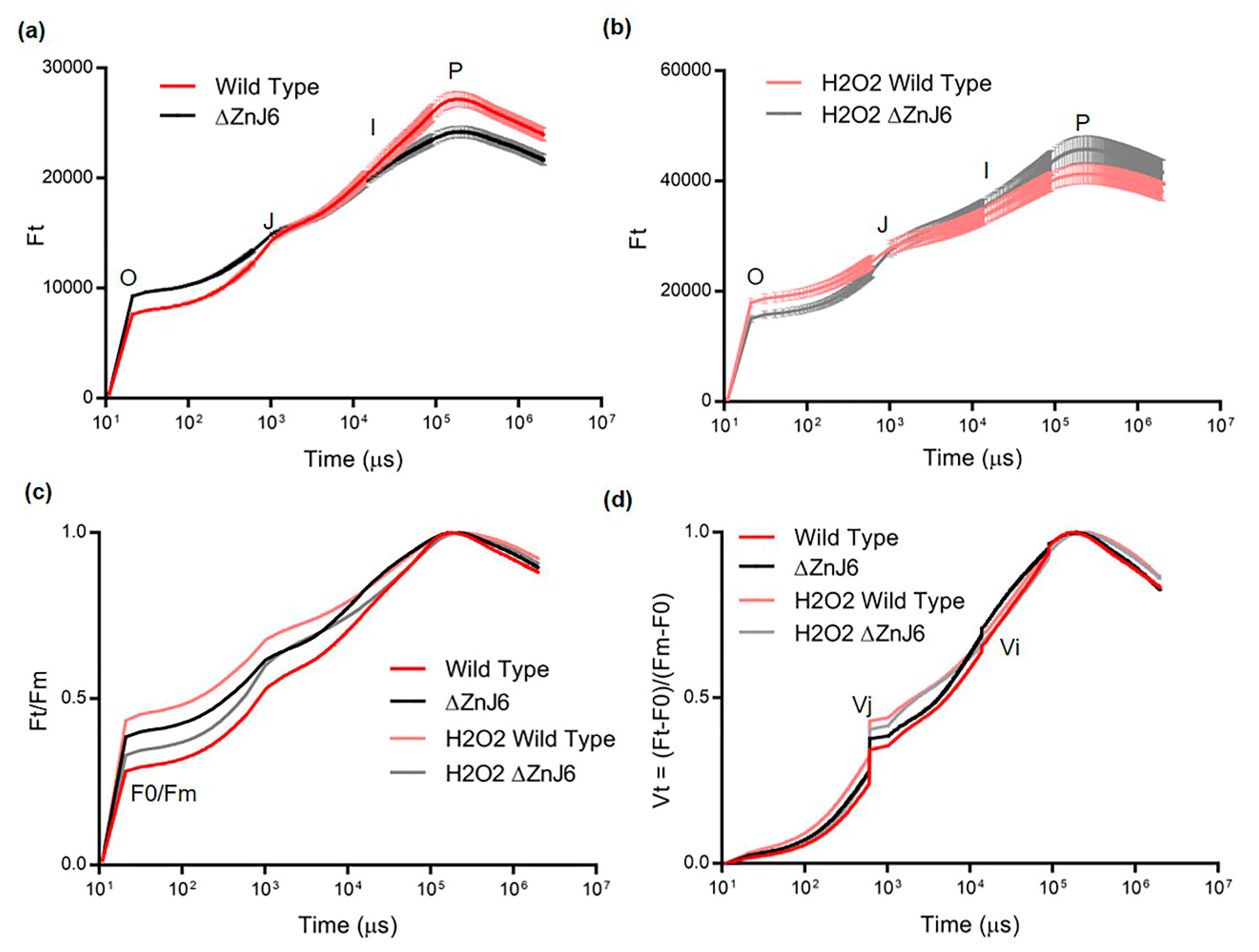
From the perspective of stress studies, the OJIP fluorescence transient measurements may serve as an indication of the physiological condition of plants [
24]. The OJIP experiments using the ΔZnJ6 mutant and its WT control cells under optimal environmental conditions showed a higher F0 and lower Fm values for the ΔZnJ6 mutant (
Figure 10a). This could indicate a lower photosynthetic activity of the ΔZnJ6 mutant in an optimal environment and could suggest that the ZnJ6 protein plays an important role in maintaining the functional state of the protein components in the photosynthetic machinery, as indicated elsewhere [
23,
26]. This is also supported by the observation that the recombinant ZnJ6 protein interacts with the Chlorophyll-binding proteins a and b along with PSI components in pull-down assays. An increase in F0/Fm (
Figure 10c) and Vj values (
Figure 10d) for the ΔZnJ6 cells under normal conditions also suggest that the ZnJ6 protein could help in maintaining the plastoquinol pool oxidized, thereby improving the photosynthetic efficiency of WT. However, under oxidizing conditions (2 mM H
2O
2), we observed that the ΔZnJ6 mutant displayed a better photosynthetic performance as compared to the WT cells (
Figure 10b–d) indicating that in an oxidizing environment, ZnJ6 was not required as under normal conditions. The higher photosynthetic efficiency of WT cells under optimal conditions could also be explained by the oxidizing activity of ZnJ6 that enables the electron flow from PSII to PSI. This is also supported by the co-purification of PSI components with ZnJ6 in the pull-down assays (
Figure 7 and
Table S1). This association could also be explained by the putative involvement of ZnJ6 in the PSI assembly.
In conclusion, ZnJ6 is a DnaJ-like oxidizing chaperone that has no reducing power. It was shown to localize in the thylakoid membrane of
Chlamydomonas. ZnJ6 increased the photosynthetic activity of WT cells under normal conditions, and its reduced expression in the ΔZnJ6 mutant decreased this activity. However, the ΔZnJ6 mutant cells were more tolerant to oxidizing conditions as compared to WT cells, indicating that the oxidizing activity of ZnJ6 was not required under oxidizing conditions and even increased the sensitivity of the WT cells to grow under these conditions. This is also supported by the results obtained under reducing conditions (DTT), where the ΔZnJ6 mutant growth was inferior to WT cells. ZnJ6 was also shown to be required for growth at elevated temperatures, in line with its ability to prevent protein aggregation at elevated temperatures using in vitro assays (
Figure 4 and
Figure S6). ZnJ6 also assists in maintaining a photosynthetic balance in an optimal environment as it interacts with PSI complexes and may contribute to the oxidative status of PSI electron acceptors. We also hypothesize that ZnJ6 could be involved in the assembly of photosynthetic complexes, as observed for other DnaJ-like proteins [
12,
13].
4. Materials and Methods
4.1. Isolation of RNA, cDNA Synthesis, and Cloning
Early log-phase cells of
Chlamydomonas reinhardtii cc-125 were measured by taking absorbance at 750 nm and counting (OD
750 = 0.25 − 0.35; 2 × 10
6 cells/mL). The cells were used to isolate total RNA by the TRI Reagent (Sigma-Aldrich, Rehovot, Israel) protocol. The cDNA was synthesized using the high capacity cDNA reverse transcription protocol (Applied Biosystems, Foster City, CA, USA) with 1 µg RNA as a template. Bacterial clones for recombinant protein expression were generated as described in the Supporting methods S1, S2 and, S3 section and the primers used are listed in
Tables S2 and S3.
4.2. Chlamydomonas Strains and Growth Conditions
Chlamydomonas strains ((cc-125 and cc-4533) were grown and maintained on Tris-acetate-phosphate (TAP) plates (1 L of TAP media contains 50 mM of 2-amino-2-(hydroxymethyl)-1,3-propanediol (Tris) base, 1 mL Hunter’s trace elements (50 gr EDTA, 22 gr ZnSO
4∙7H
2O, 11.4 gr H
3BO
3, 5.06 gr MnCl
2 ∙4H
2O, 4.99 FeSO
4∙7H
2O, 1.61 gr CoCl
2∙6H
2O, 1.57 gr CuSO
4∙5H
2O, 1.10 gr Mo
7O
24(NH
4)
6∙4H
2O in 800 mL), 0.1% acetic acid, 1 mL concentrated phosphate buffer (10.8 gr K
2HPO
4, and 5.6 gr KH
2PO4 in 100 mL ddH
2O) and 25 mL TAP salt solution (16 gr NH
4Cl, 4 gr MgSO
4∙7H
2O, 2 gr CaCl
2∙2H
2O in 1 L ddH
2O) at 23 °C with light intensity of 100 µmol/s/m
2. The knock-down ZnJ6 CLiP mutant LMJ.RY0402.048147 (∆ZnJ6) with insertion in the 3′ UTR along with the background strain cc-4533 was obtained from the
Chlamydomonas Resource Center [
42]. The knock-down mutant was maintained on 10 µg/mL paromomycin on TAP plates and verified by colony PCR followed by western analysis. A fresh colony was inoculated in 10 mL TAP media with required antibiotics followed by large-scale culturing in high salt (HS, 1 L of HS media contains 5 mL of concentrated Beijerink’s (100 gr NH
4Cl, 4 gr MgSO
4∙7H
2O, 2 gr CaCl
2∙2H
2O in 1 L), 5 mL of concentrated phosphate buffer and 1 mL of Hunter’s trace metals) or TAP media (as per requirement), with 12 h dark/light cycles (at 100–150 µmol/s/m
2) and constant rotary shaking at 100 rpm.
4.3. Recombinant Protein Purification
An overnight bacterial starter culture (10 mL) in LB medium supplemented with 100 μg/mL ampicillin and 25 μg/mL chloramphenicol (for Rosetta strain only) was inoculated into 1 L of LB supplemented with required antibiotics and 1% glucose. Expression of the SBP-tagged pMBP-GB1-ZnJ6 (see method S1) was induced by the addition of 0.2 mM IPTG when cells reached OD
600 = 0.5–0.7, at 20 °C for 16 h. The culture was harvested and resuspended in lysis buffer (20 mM Tris-HCl, pH 7.4, 200 mM NaCl, 1 mM EDTA) containing 0.1% Brij 58 (Sigma-Aldrich, Rehovot, Israel), a protease inhibitor (PI) cocktail, and 5 μg/mL DNaseI. The cells were disrupted in a French press at 1500 PSI and centrifuged at 45,000 rpm (Beckman 70 Ti rotor). The supernatant was loaded onto an amylose column (NEB). After washing the column with 5 column volumes of lysis buffer, ZnJ6 was eluted with 10 mM maltose in the same buffer. Next, the protein was cleaved with TEV protease to remove the MBP tag. The SBP tagged cleaved protein was purified again over a streptavidin–Sepharose (A2S) column. Protein concentration was estimated using a BCA protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA). The SBP-tagged ZnJ6cys > Ser-mutant and MBP proteins were purified similarly. Elution fractions were analyzed on 15% SDS–PAGE (
Figure S4).
4.4. Subcellular Fractionation of Cytoplasmic and Chloroplast Fractions
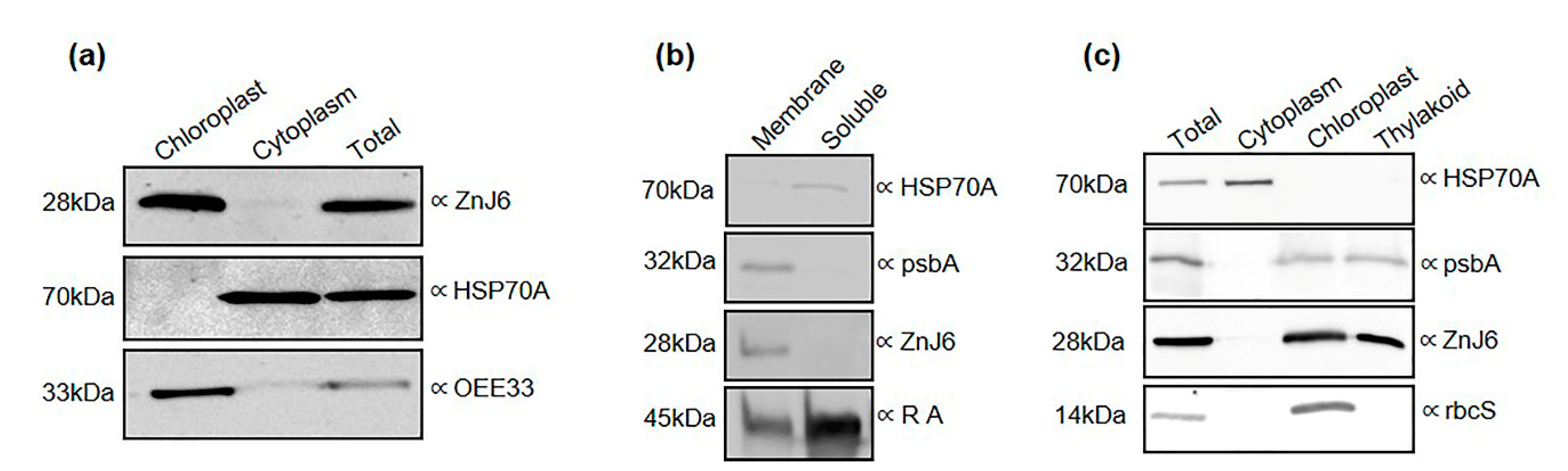
Mid-log cells of Chlamydomonas (cc-125, 1 L) were harvested and disrupted by nitrogen cavitation in a Yeda press cell disruptor at 100 PSI, which was slowly released. The disrupted cells were centrifuged at 2000× g in Corex glass tubes. The supernatant contained the cytoplasmic fraction. The pellets were resuspended in 6 mL of Percoll buffer (330 mM sorbitol, 1 mM MgCl2, 20 mM NaCl, 2 mM EDTA, 1 mM MnCl2, 2 mM NaNO3, 5 mM Na-ascorbate and 50 mM HEPES, pH 7.6). A sample of 5 mL was loaded over a 45/70% Percoll step gradient, which was centrifuged at 20,000× g for 10 min at 4 °C in an SW40 rotor. The intact chloroplasts were collected from the interphase of the step gradient. The efficiency of the chloroplast isolation was determined by measuring the chlorophyll content of the purified fraction.
The subcellular fractions were verified by Western blot analysis using gels loaded with equal amounts of protein, the protein concentration of each fraction was estimated using the bicinchoninic acid (BCA) protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA) and the ponceau stains of the Western blots were analyzed using the ImageJ software, to confirm the equal protein loads in the different fractions. Antibodies against organelle-specific proteins were used to verify the subcellular fractions [
15]. The RuBisCO small subunit (rbcS, previously raised in our lab [
43]) and the 33-kDa oxygen-evolving enzyme (OEE33, obtained from Prof. Zach Adam from the Hebrew University) as chloroplast markers, HSP70A as a cytoplasmic marker obtained from the lab of Prof. Rick Morimoto from Northwestern University. Antibodies against the 32 kDa psbA (Agrisera, Vännäs, Sweden), which encodes the D1 protein of photosystem II served as a marker for the chloroplast and its thylakoids. Antibodies against RA were from Agrisera, Vännäs, Sweden. Antibodies against the recombinant ZnJ6 fragment were generated against amino acids 1–165 (AdarBiotech, Rehovot, Israel), and the interaction is described in
Figure S1.
4.5. Separation of the Membrane and Soluble Protein Fractions
Chlamydomonas cells (cc-125, 250 mL) were grown, pelleted as above, and resuspended in 10 mL, 25 mM HEPES–KOH, pH 7.5, 5 mM MgCl
2, 0.3 M (10.2%) sucrose with PI. The resuspended pellet was then disrupted with the Yeda Press apparatus at 500 PSI. Membrane and soluble fractions were separated by centrifugation at 100,000×
g for 1 h at 4 °C. The soluble proteins were precipitated using 20% TCA (final concentration) for 1 h at 4 °C and washed twice with 100% acetone. Pellets of both soluble and membrane proteins were dissolved in 40 mM Tris-HCl pH 7.4, 5 mM EDTA, 4% SDS [
44].
4.6. Isolation of Thylakoid Membranes
Thylakoid membranes were isolated by a three-step sucrose gradient. 250 mL culture (cc-125) of mid-log cells were grown and harvested as described above. Cells were pelleted at 4500 g for 10 min at 4 °C and resuspended in 25 mM HEPES–KOH, pH 7.5, 5 mM MgCl
2, and 0.3 M sucrose supplemented with a cocktail of PIs. Cells were disrupted using nitrogen cavitation in the Yeda press cell disruptor at 500 PSI and centrifuged at 2316×
g (10 min, 4 °C) to separate between the membrane and soluble fractions. The pellet was resuspended in 5 mM HEPES–KOH, pH 7.5, 10 mM EDTA, 0.3 M sucrose, and a cocktail of PIs, followed by centrifugation at 68,600×
g for 20 min at 4 °C. The resulting pellet was resuspended in 5 mL 5 mM HEPES–KOH, pH 7.5, 10 mM EDTA, 1.8 M sucrose, and a mix of protease inhibitors. The resuspended sample (at the bottom of the tube) was carefully overlaid with 2 mL 5 mM HEPES–KOH, pH 7.5, 1.3 M sucrose, 10 mM EDTA, and then 5 mL, 5 mM HEPES–KOH, pH 7.5, 0.5 M sucrose. The thylakoid membranes were then centrifuged at 247,605×
g for 1 h at 4 °C. The thylakoid membranes were collected from the interface between the fractions containing 1.8 M and 1.3 M sucrose in the above-mentioned gradient. The thylakoid membranes were spun down at 68,600×
g for 20 min at 4 °C and washed twice with buffer containing 5 mM HEPES–KOH, pH 7.5, 10 mM EDTA, and a cocktail of PIs. The thylakoid pellet was resuspended in 200 µL of the same buffer [
44].
4.7. Circular Dichroism and Melting Curves
Circular dichroism measurements were done using a spectropolarimeter (JASCO J-815, Easton, MD, USA) with a 1 mm optical pass cuvette (Hellma, Mullhheim, Germany). The purified recombinant proteins (100 µL) were at a concentration ≥ of 100 µg/mL in Tris buffer, pH 7.5, and loaded into the clean cuvette. Single accumulation spectra ranging between 200 and 260 nm were recorded at RT. Scanning speed was set to 5 nm/min, with 6 sec response time and 1 nm bandwidth. Buffer blank (20 mM Tris, 10 mM NaCl, pH 7.5) without protein served as a control for the experiment. Spectra were baseline corrected by subtracting a blank spectrum.
Melting curves were monitored using the same conditions and buffer. The single wavelength melting curve was generated at a constant wavelength of 222 nm with a temperature range (20–80 °C). The CD Tool software was used to produce principal component analyses (PCA) for each sample. The two main components in the PCA analyses corresponded to spectra of folded and unfolded structures, and their magnitudes were plotted as a function of temperature, providing an overall indication of the thermal stability of the protein.
4.8. The PAR-PCMB Zn-Binding Assay
Zn-binding by ZnJ6 was determined using the PAR-PCMB assay, except that the thiol bound zinc was released with para-chloromercuribenzoic acid (PCMB) [
17]. Zinc release was measured by its interaction with 4-(2-pyridylazo) resorcinol (PAR) at 500 nm and compared to a ZnCl
2 standard curve. Metal-free buffers were used throughout the assay following treatment with Chelex 100 resin (5 gr in 40 mM KH
2PO
4, pH 7.5) for 1 h at 37 °C. ZnJ6 (3 μM) was mixed with 0.1 mM PAR in 40 mM KH
2PO
4 buffer to measure any free or loosely bound zinc in the solution. The addition of 30 μM PCMB to the protein solution (1 mL) caused immediate zinc release and allowed the determination and calculation of the total amount of bound zinc per ZnJ6 molecule. PAR in buffer KH
2PO
4 was used as a blank.
4.9. Citrate Synthase Assay
The ability to prevent aggregation of heat-sensitive proteins was tested using the citrate synthase (CS) assay, which monitors the holding activity of potential chaperones. ZnJ6 was added in increasing molar ratios (CS: ZnJ6, 1:0.1, 1:1, 1:2, 1:5, 1:10) to CS from porcine heart (Sigma-Aldrich, Rehovot, Israel) in 50 mM Tris pH 8.0 and 2 mM EDTA. CS, 12 μM was denatured by exposure to thermal stress (42 °C) in a 96-well plate containing 200 µL reaction volume in each well. The activity of ZnJ6 was measured by monitoring OD360 for 1 h in a plate reader (BioTek Instruments, Winooski, VT, USA). A similar experiment was done using RA as a substrate (Method S3).
4.10. Insulin (β-Chain) Aggregation Assay
The thiol-dependent activity of ZnJ6 was examined using the insulin turbidity assay [
21]. ZnJ6 and its ZnJ6cys > Ser-mutant were added to insulin in increasing molar ratios (ZnJ6: insulin, 0.2:1, 0.5:1, 1:1) in a solution of 32 μM bovine insulin (Sigma-Aldrich, Rehovot, Israel) (diluted from a stock of 1.7 mM) in a freshly prepared buffer containing 0.1 M potassium phosphate (pH 7.0) and 2 mM EDTA (100 µL). The reaction was initiated by the addition of freshly prepared DTT to a final concentration of 1 mM at 25 °C. A reaction mix containing insulin alone served as control. Precipitation of the insulin β-chain was measured at 650 nm for 2 h in a 96-well plate.
4.11. Ellman’s Test for Determination of Protein-Bound Sulfhydryl (PB-SH) Groups
To calculate the sulfhydryl (-SH) group bound to a protein in the reaction mix, ZnJ6 was added to bovine insulin in an equimolar ratio; ZnJ6 alone served as control. One mM DTT was added to the solution to reduce the insulin, as described above. The amount of protein bound-SH (PB-SH) in the reaction mix was calculated before and after incubation for 2 h. To calculate the PB-SH, DTNB was first added to the mixture, and total -SH groups (T-SH) were measured by taking absorbance at 412 nm [before precipitation with trichloroacetic acid (TCA)]. Next, a parallel mixture was TCA precipitated, centrifuged to remove all the proteins, and the DTNB was added to the protein-free supernatant. This step eliminated the effect of DTT on binding to DTNB. The value obtained was nonprotein bound-SH (NP-SH). The difference between the T-SH and the NP-SH values gave the PB-SH groups [
45]. Total-SH groups were quantified by adding 50 µL of reaction sample in 950 µL DTNB reagent (0.1 mM DTNB, 2.5 mM sodium acetate, and 100 mM Tris, pH 8). For each measurement of DTNB binding before and after TCA precipitation was measured. The mixture was incubated for 5 min at RT before monitoring the absorbance at 412 nm.
4.12. Reduced and Denatured RNaseA Reactivation Assay
Reduced and denatured RNaseA (rdRNaseA) was prepared by overnight incubation of the native RNaseA enzyme (Sigma-Aldrich, Rehovot, Israel) (20 mg/mL) in 500 μL of 0.1 M Tris-HCl pH 8.6 containing 150 mM DTT and 6 M GnHCl. Excess DTT and GnHCl were separated from the rdRNaseA using a Sephadex G-25 buffer replacement column equilibrated with 10 mM HCl. RNaseA aliquots (10 mg/mL stock) were stored at −80 °C. Reactivation of rdRNaseA was initiated by 200-fold dilution of the protein (to a final concentration of 50 μg/mL (3.8 μM) in 1 mL of reactivation buffer (0.1 M Tris-HCl pH 7.0, 0.1 M NaCl and 1 mM EDTA). The reactivation was performed in the absence or presence of increasing molar ratios (ZnJ6:rdRNaseA, 0.2:1, 0.6:1, and 1.2:1). Aliquots (50 μL) were removed at various intervals and mixed with 50 μL assay mix containing 0.1 M Tris- HCl pH 7.2, 0.1 M NaCl, and 0.3 mg/mL cytidine 2′,3′-cyclic monophosphate. RNaseA activity was measured by monitoring the hydrolysis of cytidine 2′,3′-cyclic monophosphate at 284 nm. The hydrolysis was calculated as the difference between OD
284 at t = 0 min and t = 10 min. Reactivation was presented as a percentage of hydrolysis of treated samples compared to the hydrolysis of native RNaseA [
9].
4.13. Analysis of Proteins that Associate with ZnJ6 by Pull-Down Experiments
Recombinant ZnJ6 fused to a 6 kDa streptavidin-binding peptide (SBP) tag (100 μL, 10 μM) was incubated with streptavidin–Sepharose resin (A2S). Chloroplasts (4 mL, 0.1 μg/μL) were isolated and solubilized on ice for 5 min using 1% β-DDM (n-dodecyl β-D-maltoside, Sigma-Aldrich, Rehovot, Israel) in a buffer containing 0.7 M sucrose, 0.1 M Tris-HCl, 0.3 M NaCl, pH 7.5 and a cocktail of PIs (Sigma-Aldrich, Rehovot, Israel). The sample was centrifuged for 40 min at 40,000× g, and the soluble protein fraction was collected, diluted 1:10 in the buffer (20 mM Tris-HCl, pH 7.4, 200 mM NaCl, 1 mM EDTA), and loaded onto the streptavidin–Sepharose beads (200 μL) following their incubation with the recombinant the ZnJ6 protein. The mixture was incubated at 4 °C for 2 h. The beads were then washed with 5 column volumes of the buffer (pH 7.4) to remove nonspecific proteins. Finally, the bound protein with its associated complex was eluted using 2 mM biotin. SBP-tagged MBP treated similarly served as a control for nonspecific binding of proteins to the beads.
4.14. Mass Spectrometry (MS)
The gel lane containing the proteins that were co-eluted from the streptavidin–Sepharose column were extracted from the gel and further reduced using 3 mM DTT (60 °C for 30 min), followed by modification with 10 mM iodoacetamide in 100 mM ammonium bicarbonate for 30 min at 25 °C. The sample was subsequently treated with trypsin (Promega, Madison, WI, USA) and digested overnight at 37 °C in 10 mM ammonium bicarbonate. Digested peptides were desalted, dried, resuspended in formic acid (0.1%) and resolved by reverse phase chromatography over a 30 min linear gradient with 5% to 35% acetonitrile and 0.1% formic acid in the water, a 15 min gradient with 35% to 95% acetonitrile and 0.1% formic acid in water and a 15 min gradient at 95% acetonitrile and 0.1% formic acid in water at a flow rate of 0.15 µL/min. Mass spectrometry was performed using Q-Exactive Plus mass spectrometer (Thermo Fisher Scientific, Whitbym, ON, Canada) in the positive mode set to conduct a repetitively full MS scan along with high-energy collision dissociation of the 10 dominant ions selected from the first MS scan. A mass tolerance of 10 ppm for precursor masses and 20 ppm for fragment ions was set. All analyses were performed in triplicate. The MS analyses were performed at the Smoler Center of the Technion Institute of Technology.
4.15. Statistical Analysis
Raw mass spectrometric data were analyzed using MaxQuant software version 1.5.2.8. The data were searched against C. reinhardtii proteins listed in the Phytozome database. Protein identification was set at less than a 1% false discovery rate. The MaxQuant settings selected were a minimum of 1 razor/unique peptide for identification, a minimum peptide length of six amino acids, and a maximum of two mis-cleavages. For protein quantification, summed peptide intensities were used. Missing intensities from the analyses were substituted with values close to baseline only if the values were present in the corresponding analyzed sample. LFQ intensities were compared between the three SBP-ZnJ6 biological repeats and the three SBP-MBP repeats on the Perseus software platform using a Student’s t-test.
The enrichment threshold (LFQ intensity of SBP-ZnJ6 subtracted from SBP-MBP control) was set to a log2-fold change ≤ −3 (8-fold enrichment compared to control) and p-value < 0.05. The filtered proteins were categorized both manually, based on their function in the Phytozome database, and using BLAST2GO software based on the biological process (with a minimum of 2-fold enrichment) as selected criteria. The minor categories (sub-branches) with the BLAST2GO software were merged into four broader classes: photosynthesis, metabolic process, oxidoreductases, and transporters.
4.16. Redox Sensitivity of Chlamydomonas Cells by Exposure to H2O2, MeV, or DTT
To verify the in vivo function of ZnJ6, the insertional knock-down mutant ΔZnJ6, with the paromomycin resistance, was used (LMJ.RY0402.048147).
C. reinhardtii cells (ΔZnJ6 and the WT background strain cc-4533) were grown to mid-log phase in HS medium with a light intensity of 150 µmol/s/m
2. The mutant was confirmed by PCR and western analysis using anti-ZnJ6 antibodies (
Figure S8). For all treatments, ~1 × 10
7 cells mL
−1 (1 mL) were exposed to different concentrations of H
2O
2 (2, 5, 10, and 20 mM), MeV (2, 5, 10, and 20 µM), and DTT (2, 5, 10 and 20 mM). After exposure of 2 h, the cells were washed twice and resuspended in HS media with required dilution. A total of 10
3 cells (3 µL) were seeded over HS plates and allowed to grow at 23 °C. Pictures were taken on day 5 of the growth and analyzed using MultiGauge software. Paired
t-test was performed on at least 3 replicates to compare the response of ΔZnJ6 and the WT background strain to different treatments.
4.17. Exposure of Chlamydomonas Cells to Heat Stress
C. reinhardtii cells (ΔZnJ6, WT background cells) as described above. 5 µL (serially diluted from 105 to 102 cells/5 µL) aliquots were spotted on HS plates and incubated at 23 °C (control) or exposed to heat stress by incubating the plates at 37 °C for 1, 2, and 4 h. Pictures were taken on day 5 of the growth, and the density was measured using MultiGauge software followed by t-test analysis.
4.18. Analysis of Chlorophyll Fluorescence Transient/Fast OJIP Analysis
Fluorescence of Chlorophyll a was taken using AquaPen-C AP 110-C. Mid-log
Chlamydomonas cells (2 mL) with a constant chlorophyll concentration of 2 mg/L for each sample (WT cc-4533, the ∆ZnJ6, with or without H
2O
2 treatment). Measurements were taken after turning off the actinic light for at least 5 min prior to the measurements. The fast chlorophyll fluorescence induction kinetics were done with saturating flash intensity of ca. 3000 µmol m
−2 s
−1, and measurement time was 1 sec. The plotted curve (
Figure 10a,b) is a mean of three independent repeats. In order to compare the F0/Fm and mid-log transition, the curve was normalized to Fm and relative variable fluorescence (Vt) curves (
Figure 10c,d). The Vt curve was obtained by Vt = (Ft − F0)/(Fm − F0).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









