
Replication-Coupled Chromatin Remodeling: An Overview of Disassembly and Assembly of Chromatin during Replication



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
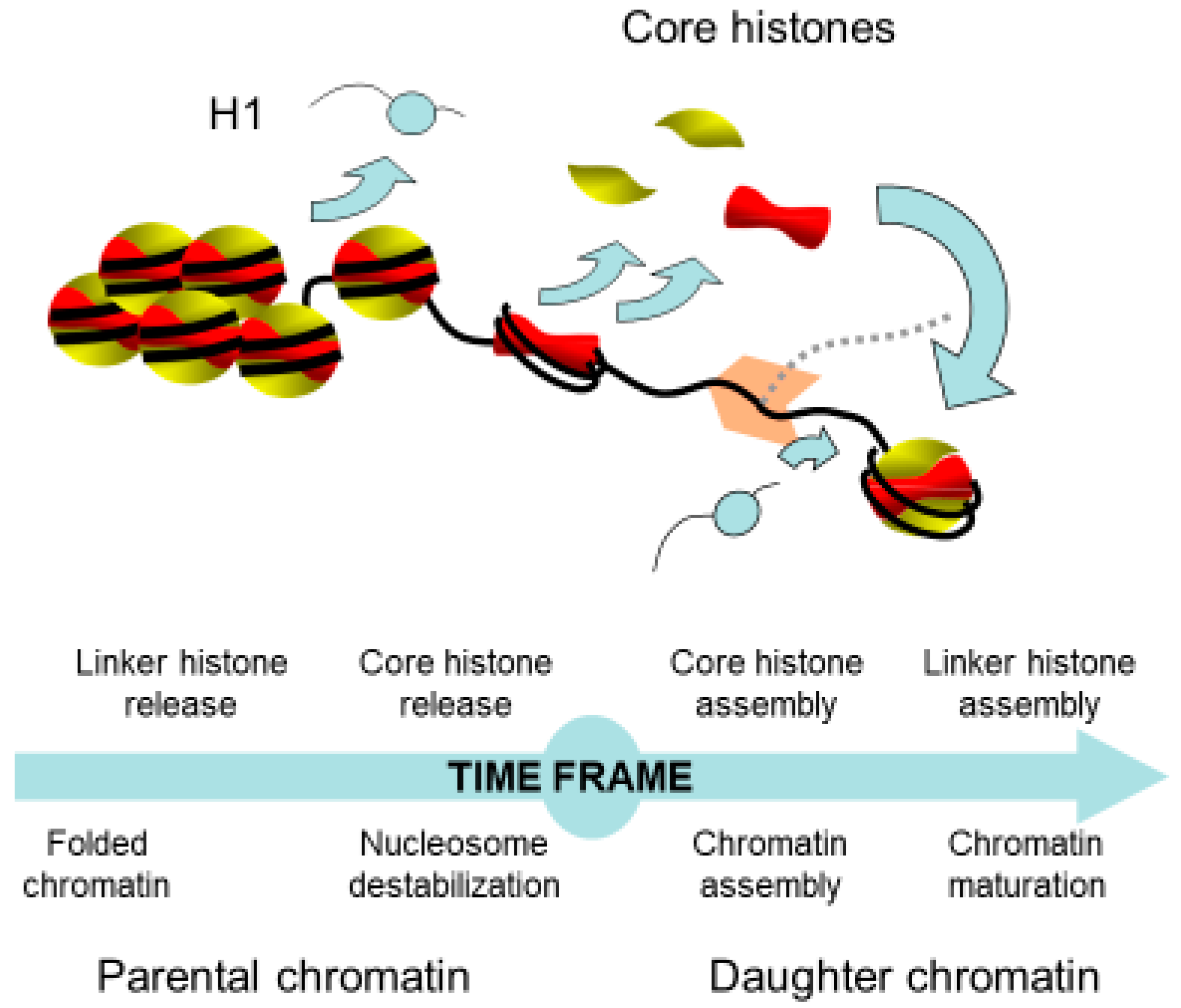
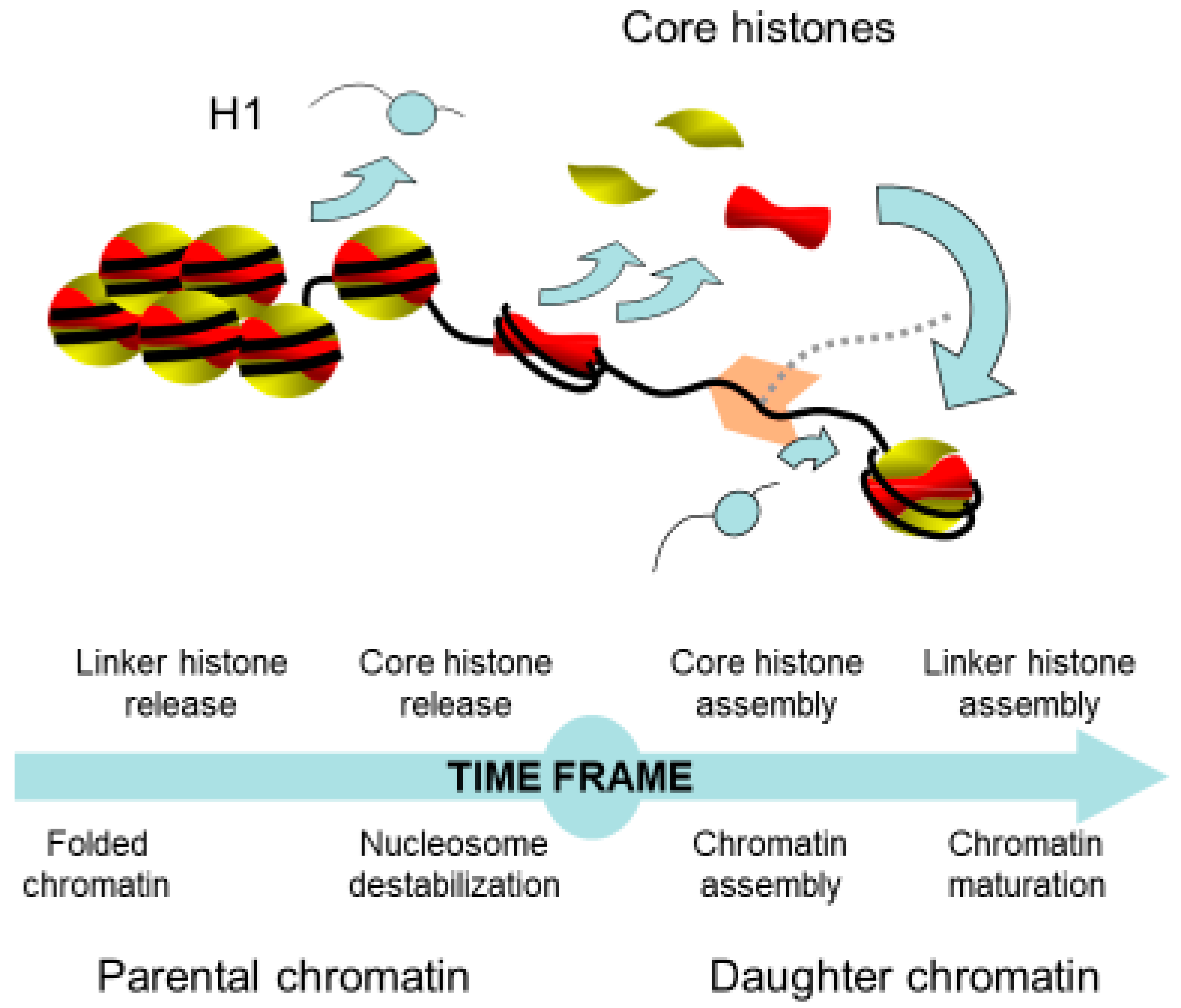
2. Chromatin Remodeling in Front of the Replication Fork
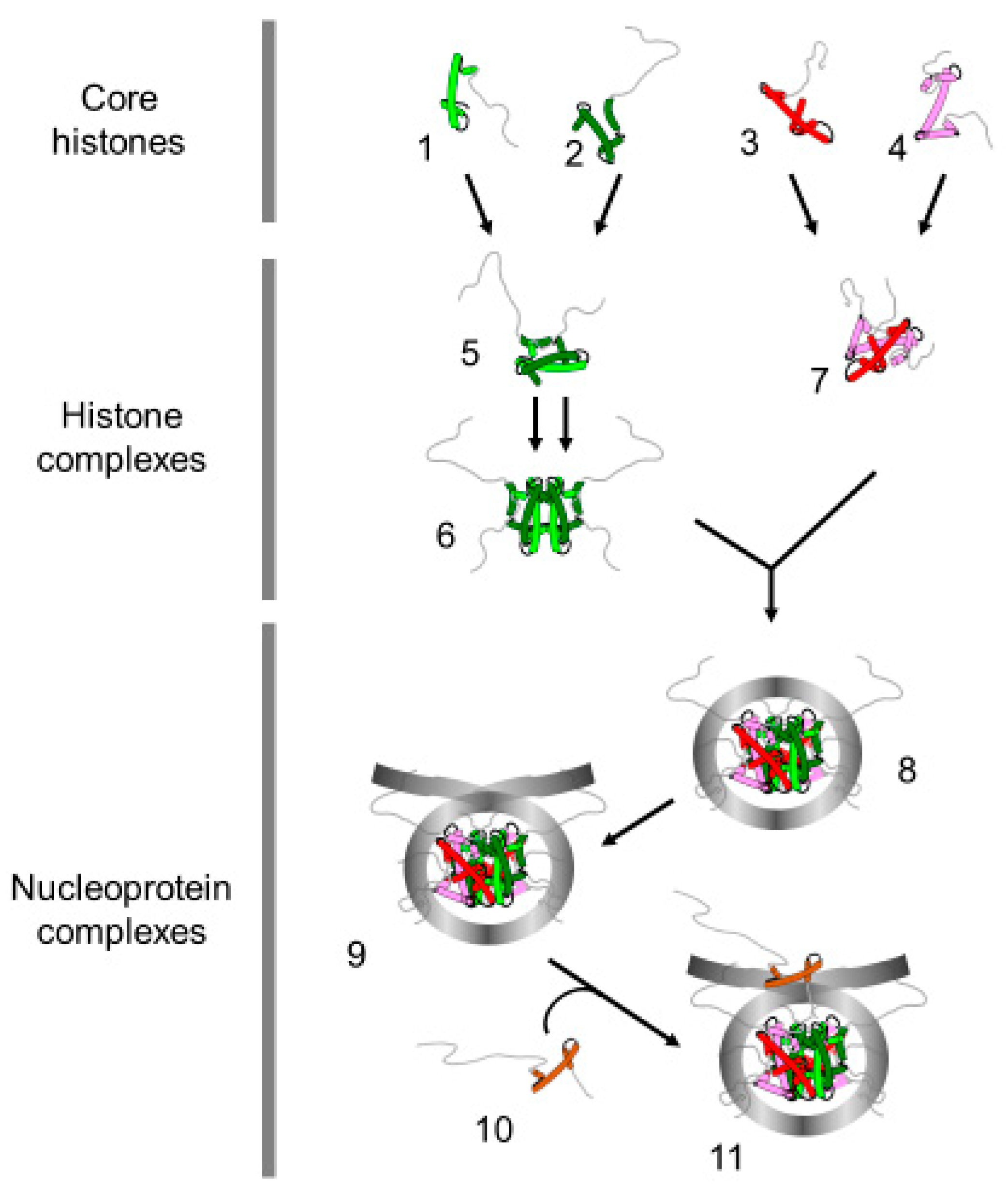
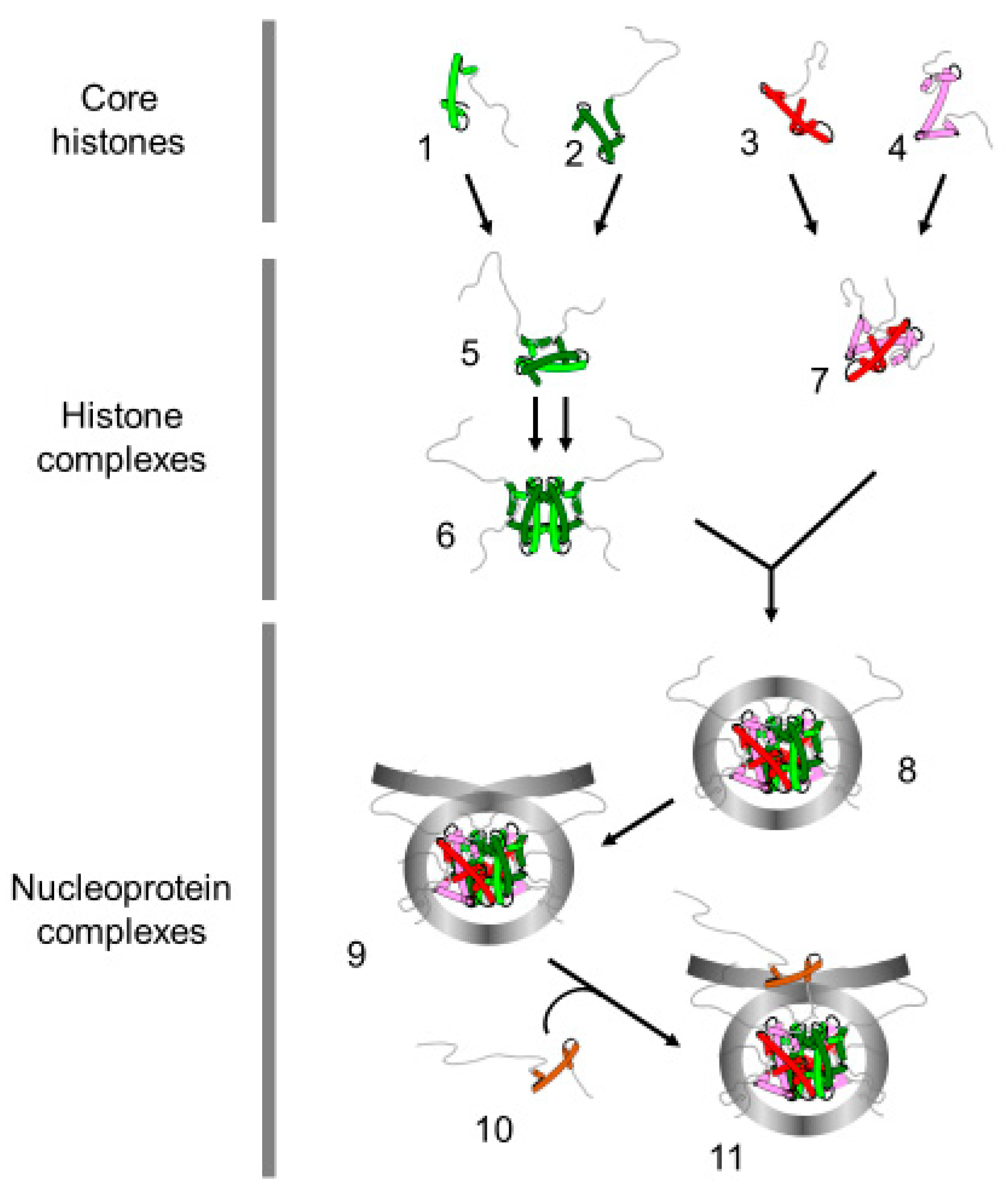
3. Nuclear Transportation of New Histones
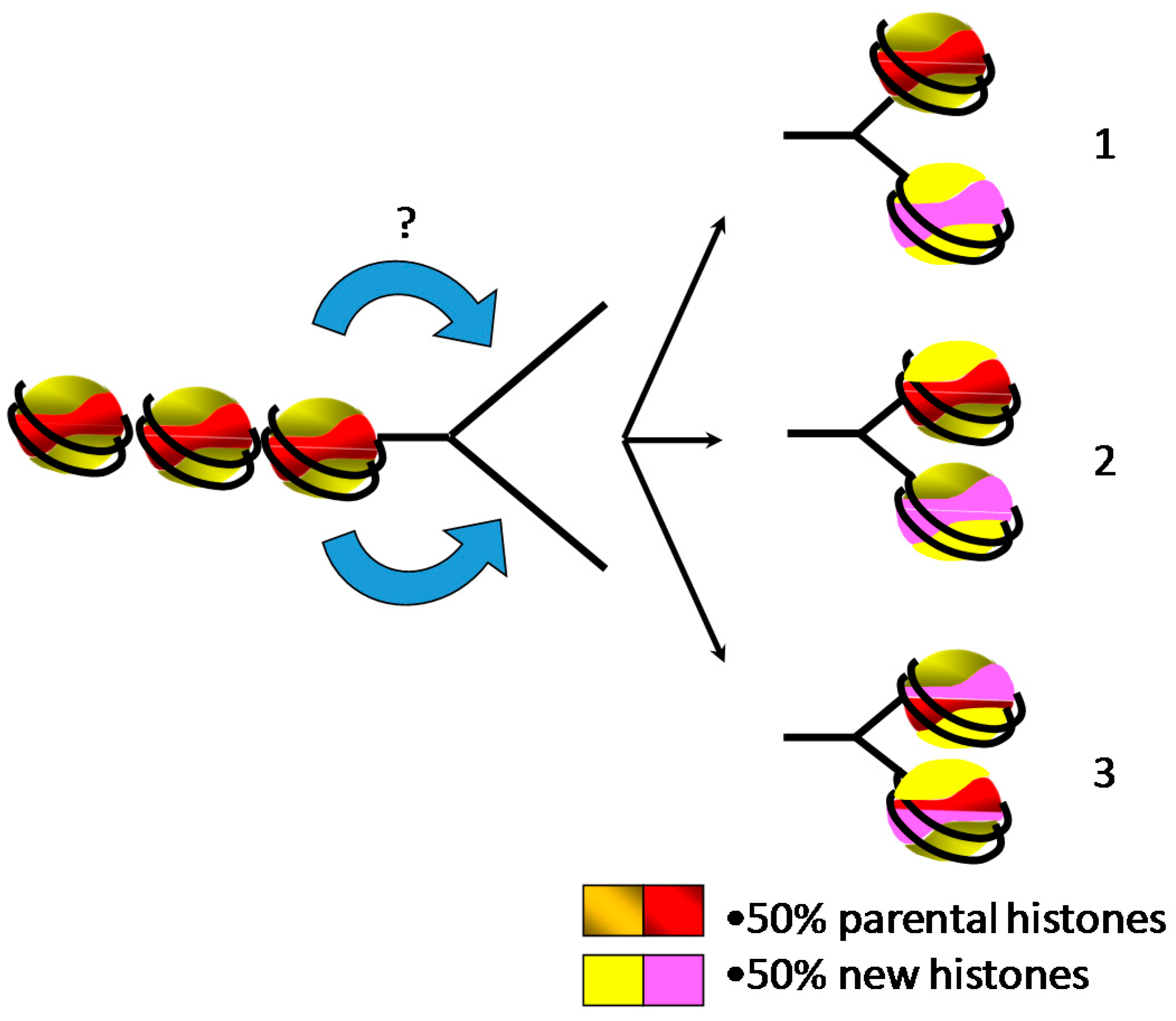
4. Assembly of Nucleosome behind the Replication Fork
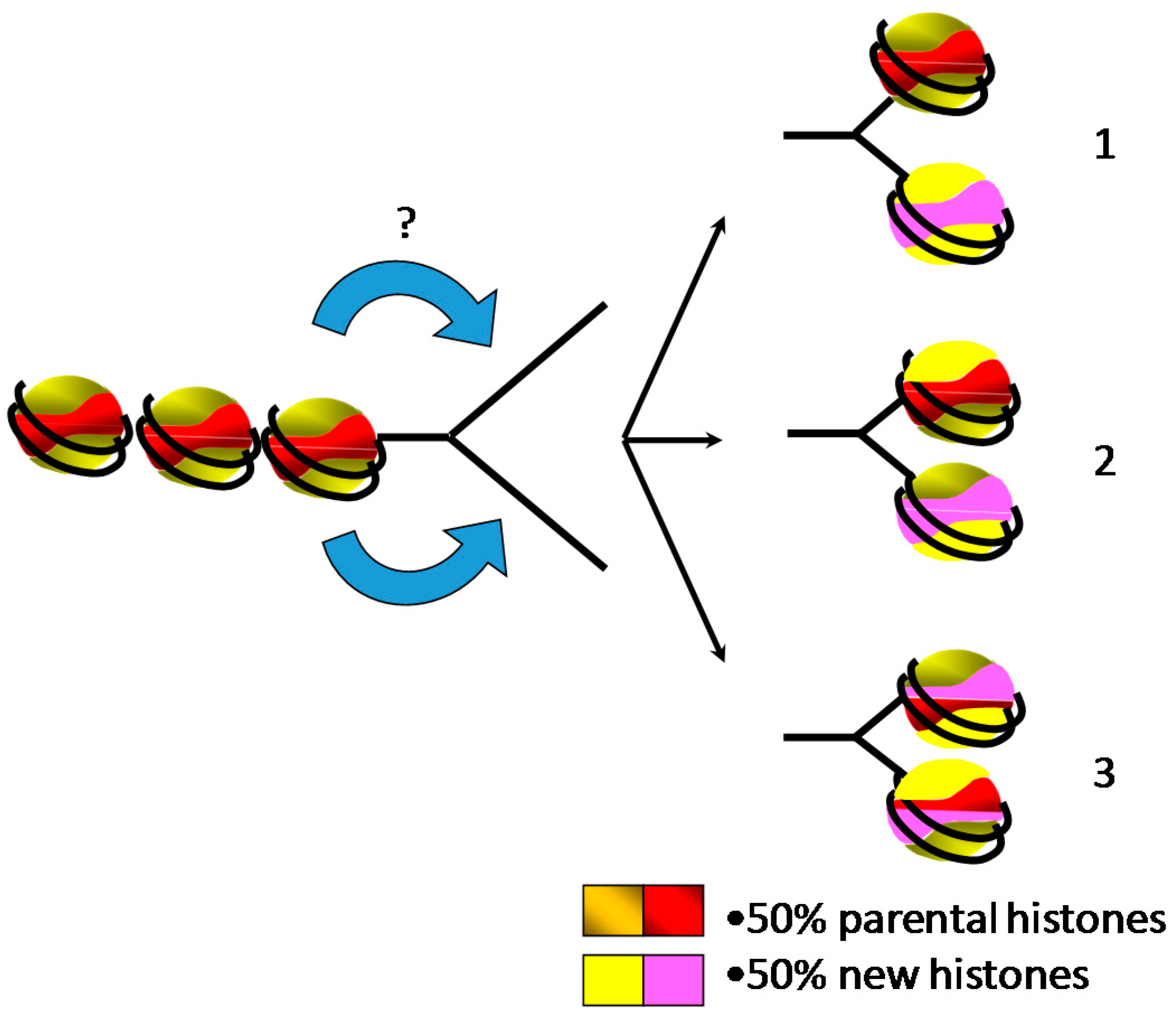
5. Inheritance of Epigenetic Marks during Replication
6. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Méchali, M. Eukaryotic DNA replication origins: Many choices for appropriate answers. Nat. Rev. Mol. Cell Biol. 2010, 11, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Groth, A.; Rocha, W.; Verreault, A.; Almouzni, G. Chromatin challenges during DNA replication and repair. Cell 2007, 128, 721–733. [Google Scholar] [CrossRef] [Green Version]
- Wolffe, A.P. Chromatin: Structure and Function; Academic Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Van Holde, K.E. Chromatin; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Arents, G.; Burlingame, R.W.; Wang, B.I.C.C.; Love, W.E.; Moudrianakis, E.N. The nucleosomal core histone octamer at 3.1 A resolution: A tripartite protein assembly and a left-handed superhelix. Proc. Natl. Acad. Sci. USA 1991, 88, 10148–10152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luger, K.; Mader, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Bednar, J.; Garcia-Saez, I.; Boopathi, R.; Cutter, A.R.; Papai, G.; Reymer, A.; Syed, S.H.; Lone, I.N.; Tonchev, O.; Crucifix, C.; et al. Structure and Dynamics of a 197 bp Nucleosome in Complex with Linker Histone H1. Mol. Cell 2017, 66, 384–397.e8. [Google Scholar] [CrossRef] [Green Version]
- Bednar, J.; Hamiche, A.; Dimitrov, S. H1-nucleosome interactions and their functional implications. Biochim. Biophys. Acta 2016, 1859, 436–443. [Google Scholar] [CrossRef]
- Wolffe, A.P.; Khochbin, S.; Dimitrov, S. What do linker histones do in chromatin? Bioessays 1997, 19, 249–255. [Google Scholar] [CrossRef]
- Harshman, S.W.; Young, N.L.; Parthun, M.R.; Freitas, M.A. H1 histones: Current perspectives and challenges. Nucleic Acids Res. 2013, 41, 9593–9609. [Google Scholar] [CrossRef] [Green Version]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Wolffe, A.P.; Hayes, J.J. Chromatin disruption and modification. Nucleic Acids Res. 1999, 27, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Jenuwein, T.; Allis, C.D. Translating the histone code. Science 2001, 293, 1074–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothbart, S.B.; Strahl, B.D. Interpreting the language of histone and DNA modifications. Biochim. Biophys. Acta 2014, 1839, 627–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, K.; Jeon, J.; Park, K.; Kim, J. Writing, erasing and reading histone lysine methylations. Exp. Mol. Med. 2017, 49, e324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Probst, A.V.; Dunleavy, E.; Almouzni, G. Epigenetic inheritance during the cell cycle. Nat. Rev. Mol. Cell Biol. 2009, 10, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Zweidler, A. Core Histone Variants of the Mouse: Primary Structure and Differential Expression. In Histone Genes, Structure, Organization and Differential Expression; John Wiley & Sons: Hoboken, NJ, USA, 1984. [Google Scholar]
- Talbert, P.B.; Henikoff, S. Histone variants on the move: Substrates for chromatin dynamics. Nat. Rev. Mol. Cell Biol. 2017, 18, 115–126. [Google Scholar] [CrossRef]
- Redon, C.; Pilch, D.; Rogakou, E.; Sedelnikova, O.; Newrock, K.; Bonner, W. Histone H2A variants H2AX and H2AZ. Curr. Opin. Genet. Dev. 2002, 12, 162–169. [Google Scholar] [CrossRef]
- Long, M.; Sun, X.; Shi, W.; Yanru, A.; Leung, S.T.; Ding, D.; Cheema, M.S.; MacPherson, N.; Nelson, C.J.; Ausio, J.; et al. A novel histone H4 variant H4G regulates rDNA transcription in breast cancer. Nucleic Acids Res. 2019, 47, 8399–8409. [Google Scholar] [CrossRef] [Green Version]
- Boulard, M.; Bouvet, P.; Kundu, T.K.; Dimitrov, S. Histone variant nucleosomes: Structure, function and implication in disease. Subcell. Biochem. 2007, 41, 71–89. [Google Scholar]
- Luger, K.; Dechassa, M.L.; Tremethick, D.J. New insights into nucleosome and chromatin structure: An ordered state or a disordered affair? Nat. Rev. Mol. Cell Biol. 2012, 13, 436–447. [Google Scholar] [CrossRef] [Green Version]
- Ferrand, J.; Rondinelli, B.; Polo, S.E. Histone Variants: Guardians of Genome Integrity. Cells 2020, 9, 2424. [Google Scholar] [CrossRef] [PubMed]
- Maxson, R.; Cohn, R.; Kedes, L.; Mohun, T. Expression and Organization of Histone Genes. Annu. Rev. Genet. 1983, 17, 239–277. [Google Scholar] [CrossRef]
- Wu, R.S.; Bonner, W.M. Separation of basal histone synthesis from S-phase histone synthesis in dividing cells. Cell 1981, 27, 321–330. [Google Scholar] [CrossRef]
- Hammond, C.M.; Strømme, C.B.; Huang, H.; Patel, D.J.; Groth, A. Histone chaperone networks shaping chromatin function. Nat. Rev. Mol. Cell Biol. 2017, 18, 141–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gontijo, A.M.; Green, C.M.; Almouzni, G. Repairing DNA damage in chromatin. Biochimie 2003, 85, 1133–1147. [Google Scholar] [CrossRef]
- Gurard-Levin, Z.A.; Quivy, J.-P.; Almouzni, G. Histone Chaperones: Assisting Histone Traffic and Nucleosome Dynamics. Annu. Rev. Biochem. 2014, 83, 487–517. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, A.T. Assembling chromatin: The long and winding road. Biochim. Biophys. Acta Gene Regul. Mech. 2011, 1819, 196–210. [Google Scholar] [CrossRef]
- Shen, X.; Yu, L.; Weir, J.W.; Gorovsky, M.A. Linker histones are not essential and affect chromatin condensation in vivo. Cell 1995, 82, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Thiriet, C.; Hayes, J.J. Linker histone phosphorylation regulates global timing of replication origin firing. J. Biol. Chem. 2009, 284, 2823–2829. [Google Scholar] [CrossRef] [Green Version]
- Rutowicz, K.; Lirski, M.; Mermaz, B.; Schubert, J.; Teano, G.; Mestiri, I.; Kroteń, M.A.; Fabrice, T.N.; Fritz, S.; Grob, S.; et al. Linker histones regulate fine-scale chromatin organization and modulate developmental decisions in Arabidopsis. Genome Biol. 2018. [Google Scholar] [CrossRef]
- Dou, Y.; Gorovsky, M.A. Phosphorylation of linker histone H1 regulates gene expression in vivo by creating a charge patch. Mol. Cell 2000, 6, 225–231. [Google Scholar] [CrossRef]
- Bayona-Feliu, A.; Casas-Lamesa, A.; Carbonell, A.; Climent-Cantó, P.; Tatarski, M.; Pérez-Montero, S.; Azorín, F.; Bernués, J. Histone H1: Lessons from Drosophila. Biochim. Biophys. Acta Gene Regul. Mech. 2016, 1859, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Fyodorov, D.V.; Zhou, B.R.; Skoultchi, A.I.; Bai, Y. Emerging roles of linker histones in regulating chromatin structure and function. Nat. Rev. Mol. Cell Biol. 2018, 19, 192. [Google Scholar] [CrossRef] [PubMed]
- Willcockson, M.A.; Healton, S.E.; Weiss, C.N.; Bartholdy, B.A.; Botbol, Y.; Mishra, L.N.; Sidhwani, D.S.; Wilson, T.J.; Pinto, H.B.; Maron, M.I.; et al. H1 histones control the epigenetic landscape by local chromatin compaction. Nature 2020, 589, 293–298. [Google Scholar] [CrossRef]
- DePamphilis, M.L.; Wassarman, P.M. Replication of eukaryotic chromosomes: A close-up of the replication fork. Annu. Rev. Biochem. 1980, 49, 627–666. [Google Scholar] [CrossRef]
- Gruss, C.; Wu, J.; Koller, T.; Sogo, J.M. Disruption of the nucleosomes at the replication fork. EMBO J. 1993, 12, 4533–4545. [Google Scholar] [CrossRef]
- Gasser, R.; Koller, T.; Sogo, J.M. The stability of nucleosomes at the replication fork. J. Mol. Biol. 1996, 258, 224–239. [Google Scholar] [CrossRef]
- Nightingale, K.P.; Becker, P.B. Structural and functional analysis of chromatin assembled from defined histones. Methods 1998, 15, 343–353. [Google Scholar] [CrossRef]
- Gruss, C.; Sogo, J.M. Chromatin replication. Bioessays 1992, 14, 1–8. [Google Scholar] [CrossRef]
- DePamphilis, M.L.; Wassarman, P.M. Regulation of chromosomal replication and transcription during early mammalian development. Bioessays 1987, 7, 265–271. [Google Scholar] [CrossRef]
- Weintraub, H.; Worcel, A.; Alberts, B. A model for chromatin based upon two symmetrically paired half-nucleosomes. Cell 1976, 9, 409–417. [Google Scholar] [CrossRef]
- Riley, D.; Weintraub, H. Conservative segregation of parental histones during replication in the presence of cycloheximide. Proc. Natl. Acad. Sci. USA 1979, 76, 328–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alabert, C.; Barth, T.K.; Reverón-Gómez, N.; Sidoli, S.; Schmidt, A.; Jensen, O.N.; Imhof, A.; Groth, A. Two distinct modes for propagation of histone PTMs across the cell cycle. Genes Dev. 2015, 29, 585–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlatanova, J.; Bishop, T.C.; Victor, J.M.; Jackson, V.; van Holde, K. The nucleosome family: Dynamic and growing. Structure 2009, 17, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.F.; Jackson, V. Measurement of the frequency of histone displacement during the in vitro transcription of nucleosomes: RNA is a competitor for these histones. Biochemistry 1997, 36, 12371–12382. [Google Scholar] [CrossRef]
- Chabouté, M.E.; Chaubet, N.; Gigot, C.; Philipps, G. Histones and histone genes in higher plants: Structure and genomic organization. Biochimie 1993, 75, 523–531. [Google Scholar] [CrossRef]
- Chaboute, M.E.; Chaubet, N.; Clement, B.; Gigot, C.; Philipps, G. Polyadenylation of histone H3 and H4 mRNAs in dicotyledonous plants. Gene 1988, 71, 217–223. [Google Scholar] [CrossRef]
- Old, R.W.; Woodland, H.R. Histone genes: Not so simple after all. Cell 1984, 38, 624–626. [Google Scholar] [CrossRef]
- Campos, E.I.; Fillingham, J.; Li, G.; Zheng, H.; Voigt, P.; Kuo, W.H.; Seepany, H.; Gao, Z.; Day, L.A.; Greenblatt, J.F.; et al. The program for processing newly synthesized histones H3.1 and H4. Nat. Struct. Mol. Biol. 2010, 17, 1343–1351. [Google Scholar] [CrossRef] [Green Version]
- Pardal, A.J.; Fernandes-Duarte, F.; Bowman, A.J. The histone chaperoning pathway: From ribosome to nucleosome. Essays Biochem. 2019, 63, 29–43. [Google Scholar] [CrossRef] [Green Version]
- Apta-Smith, M.J.; Hernandez-Fernaud, J.R.; Bowman, A.J. Evidence for the nuclear import of histones H3.1 and H4 as monomers. EMBO J. 2018, 37, e98714. [Google Scholar] [CrossRef] [PubMed]
- Ejlassi, A.; Menil-Philippot, V.; Galvani, A.; Thiriet, C. Histones H3 and H4 require their relevant amino-tails for efficient nuclear import and replication-coupled chromatin assembly in vivo. Sci. Rep. 2017, 7, 3050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Carrillo, A.; Wangh, L.J.; Allfrey, V.G. Processing of newly synthesized histone molecules. Science 1975, 190, 117–128. [Google Scholar] [CrossRef]
- Sobel, R.E.; Cook, R.G.; Perry, C.A.; Annunziato, A.T.; Allis, C.D. Conservation of deposition-related acetylation sites in newly synthesized histones H3 and H4. Proc. Natl. Acad. Sci. USA 1995, 92, 1237–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verreault, A.; Kaufman, P.D.; Kobayashi, R.; Stillman, B. Nucleosome assembly by a complex of CAF-1 and acetylated histones H3/H4. Cell 1996, 87, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Mersfelder, E.L.; Parthun, M.R. Involvement of Hat1p (Kat1p) catalytic activity and subcellular localization in telomeric silencing. J. Biol. Chem. 2008, 283, 29060–29068. [Google Scholar] [CrossRef] [Green Version]
- Ai, X.; Parthun, M.R. The nuclear Hat1p/Hat2p complex: A molecular link between type B histone acetyltransferases and chromatin assembly. Mol. Cell 2004, 14, 195–205. [Google Scholar] [CrossRef]
- Ejlassi-Lassallette, A.; Mocquard, E.; Arnaud, M.C.; Thiriet, C. H4 replication-dependent diacetylation and Hat1 promote s-phase chromatin assembly in vivo. Mol. Biol. Cell 2011, 22, 245–255. [Google Scholar] [CrossRef]
- Chang, L.; Loranger, S.S.; Mizzen, C.; Ernst, S.G.; Allis, C.D.; Annunziato, A.T. Histones in transit: Cytosolic histone complexes and diacetylation of H4 during nucleosome assembly in human cells. Biochemistry 1997, 36, 469–480. [Google Scholar] [CrossRef]
- Ling, X.; Harkness, T.A.; Schultz, M.C.; Fisher-Adams, G.; Grunstein, M. Yeast histone H3 and H4 amino termini are important for nucleosome assembly in vivo and in vitro: Redundant and position-independent functions in assembly but not in gene regulation. Genes Dev. 1996, 10, 686–699. [Google Scholar] [CrossRef] [Green Version]
- Thiriet, C.; Hayes, J.J. Histone proteins in vivo: Cell-cycle-dependent physiological effects of exogenous linker histones incorporated into Physarum polycephalum. Methods A Companion Methods Enzymol. 1999, 17, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Thiriet, C. Analysis of chromatin assembled in vivo using exogenous histones in Physarum polycephalum. Methods 2004, 33, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Thiriet, C.; Hayes, J.J. A novel labeling technique reveals a function for histone H2A/H2B dimer tail domains in chromatin assembly in vivo. Genes Dev. 2001, 15, 2048–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardes, N.E.; Chook, Y.M. Nuclear import of histones. Biochem. Soc. Trans. 2020, 48, 2753–2767. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, A.T. The fork in the road: Histone partitioning during DNA replication. Genes 2015, 6, 353–371. [Google Scholar] [CrossRef] [Green Version]
- Jackson, V.; Chalkley, R. Histone segregation on replicating chromatin. Biochemistry 1985, 24, 6930–6938. [Google Scholar] [CrossRef]
- Jackson, V.; Granner, D.; Chalkley, R. Deposition of histone onto the replicating chromosome: Newly synthesized histone is not found near the replication fork. Proc. Natl. Acad. Sci. USA 1976, 73, 2266–2269. [Google Scholar] [CrossRef] [Green Version]
- Jackson, V.; Chalkley, R. A new method for the isolation of replicative chromatin: Selective deposition of histone on both new and old DNA. Cell 1981, 23, 121–134. [Google Scholar] [CrossRef]
- Petryk, N.; Dalby, M.; Wenger, A.; Stromme, C.B.; Strandsby, A.; Andersson, R.; Groth, A. MCM2 promotes symmetric inheritance of modified histones during DNA replication. Science 2018, 361, 1389–1392. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, S.; Wolffe, A.P. Chromatin and nuclear assembly: Experimental approaches towards the reconstitution of transcriptionally active and silent states. Biochim. Biophys. Acta 1995, 1260, 1–13. [Google Scholar] [CrossRef]
- Eickbush, T.H.; Moudrianakis, E.N. The histone core complex: An octamer assembled by two sets of protein-protein interactions. Biochemistry 1978, 17, 4955–4963. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.J. In vitro analysis of chromatin structure in model systems. Methods 2004, 33, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Stillman, B. Stepwise assembly of chromatin during DNA replication in vitro. EMBO J. 1991, 10, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Stillman, B. Purification and characterization of CAF-I, a human cell factor required for chromatin assembly during DNA replication in vitro. Cell 1989, 58, 15–25. [Google Scholar] [CrossRef]
- Annunziato, A.T.; Seale, R.L. Presence of nucleosomes within irregularly cleaved fragments of newly replicated chromatin. Nucleic Acids Res. 1984, 12, 6179–6196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worcel, A.; Han, S.; Wong, M.L. Assembly of newly replicated chromatin. Cell 1978, 15, 969–977. [Google Scholar] [CrossRef]
- Jackson, V. In vivo studies on the dynamics of histone-DNA interaction: Evidence for nucleosome dissolution during replication and transcription and a low level of dissolution independent of both. Biochemistry 1990, 29, 719–731. [Google Scholar] [CrossRef]
- Zhang, W.; Feng, J.; Li, Q. The replisome guides nucleosome assembly during DNA replication. Cell Biosci. 2020, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Escobar, T.M.; Oksuz, O.; Saldaña-Meyer, R.; Descostes, N.; Bonasio, R.; Reinberg, D. Active and Repressed Chromatin Domains Exhibit Distinct Nucleosome Segregation during DNA Replication. Cell 2019, 179, 953–963.e11. [Google Scholar] [CrossRef]
- Schlissel, G.; Rine, J. The nucleosome core particle remembers its position through DNA replication and RNA transcription. Proc. Natl. Acad. Sci. USA 2019, 116, 20605–20611. [Google Scholar] [CrossRef] [Green Version]
- Gruszka, D.T.; Xie, S.; Kimura, H.; Yardimci, H. Single-molecule imaging reveals control of parental histone recycling by free histones during DNA replication. Sci. Adv. 2020, 6, eabc0330. [Google Scholar] [CrossRef] [PubMed]
- Elliott, G.O.; Murphy, K.J.; Hayes, J.J.; Thiriet, C. Replication-independent nucleosome exchange is enhanced by local and specific acetylation of histone H4. Nucleic Acids Res. 2013, 41, 2228–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mocquard-Bucher, E.; Galvani, A.; Thiriet, C. Histone H4 acetylation links nucleosome turnover and nucleosome assembly: Lessons from the slime mold Physarum polycephalum. Front. Life Sci. 2013, 7, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Henikoff, S.; Smith, M.M. Histone variants and epigenetics. Cold Spring Harb. Perspect. Biol. 2015, 7, a019364. [Google Scholar] [CrossRef] [Green Version]
- Clément, C.; Orsi, G.A.; Gatto, A.; Boyarchuk, E.; Forest, A.; Hajj, B.; Miné-Hattab, J.; Garnier, M.; Gurard-Levin, Z.A.; Quivy, J.P.; et al. High-resolution visualization of H3 variants during replication reveals their controlled recycling. Nat. Commun. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Pierron, G.; Sauer, H.W. More evidence for replication-transcription-coupling in Physarum polycephalum. J. Cell Sci. 1980, 41, 105–113. [Google Scholar]
- Vasseur, P.; Tonazzini, S.; Ziane, R.; Camasses, A.; Rando, O.J.; Radman-Livaja, M. Dynamics of Nucleosome Positioning Maturation following Genomic Replication. Cell Rep. 2016, 16, 2651–2665. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, S.; Henikoff, S. Transcriptional Regulators Compete with Nucleosomes Post-replication. Cell 2016, 165, 580–592. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duc, C.; Thiriet, C. Replication-Coupled Chromatin Remodeling: An Overview of Disassembly and Assembly of Chromatin during Replication. Int. J. Mol. Sci. 2021, 22, 1113. https://doi.org/10.3390/ijms22031113
Duc C, Thiriet C. Replication-Coupled Chromatin Remodeling: An Overview of Disassembly and Assembly of Chromatin during Replication. International Journal of Molecular Sciences. 2021; 22(3):1113. https://doi.org/10.3390/ijms22031113
Chicago/Turabian StyleDuc, Céline, and Christophe Thiriet. 2021. "Replication-Coupled Chromatin Remodeling: An Overview of Disassembly and Assembly of Chromatin during Replication" International Journal of Molecular Sciences 22, no. 3: 1113. https://doi.org/10.3390/ijms22031113
APA StyleDuc, C., & Thiriet, C. (2021). Replication-Coupled Chromatin Remodeling: An Overview of Disassembly and Assembly of Chromatin during Replication. International Journal of Molecular Sciences, 22(3), 1113. https://doi.org/10.3390/ijms22031113