
Long Noncoding RNAs: Recent Insights into Their Role in Male Infertility and Their Potential as Biomarkers and Therapeutic Targets

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Classification and Regulatory Mechanisms Underlying the Action of Long Noncoding RNAs
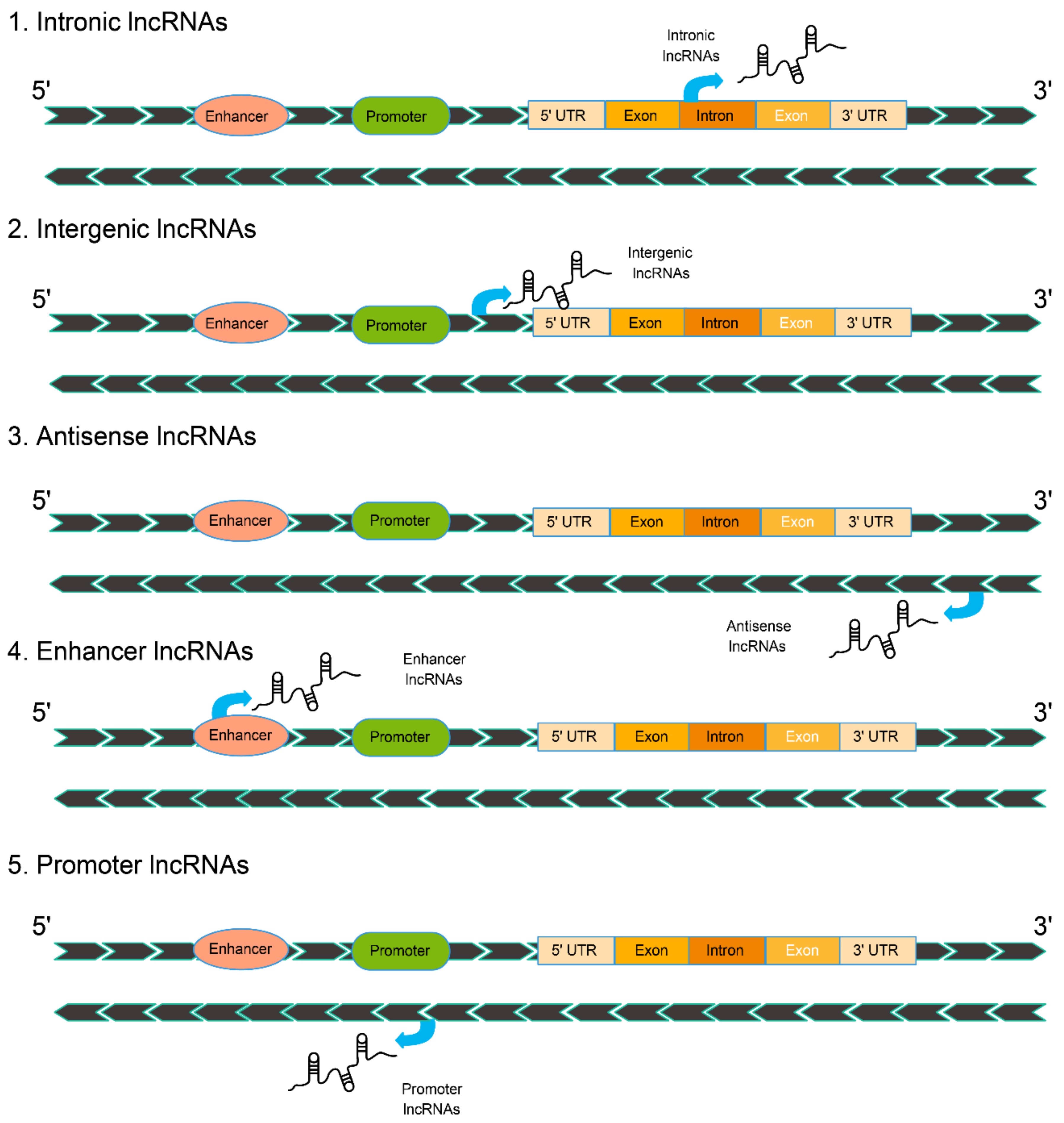
2.1. The Classification of Long Noncoding RNAs
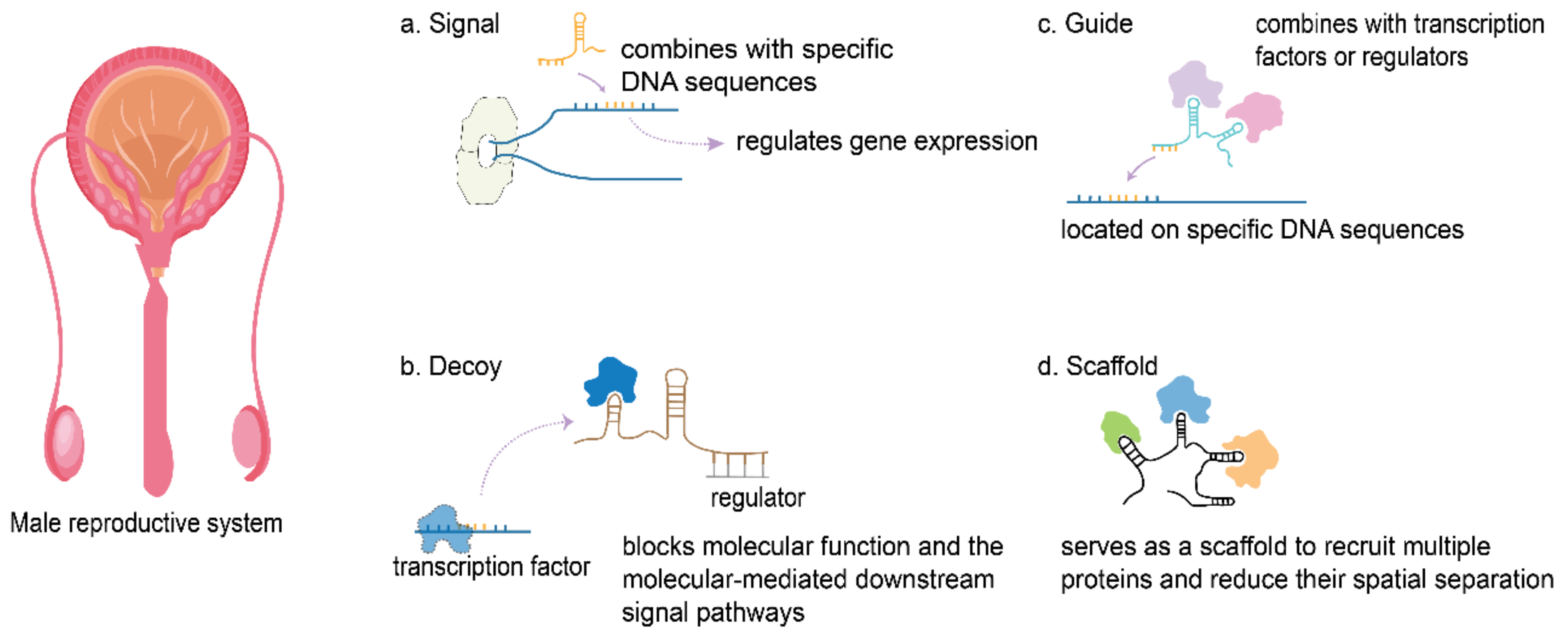
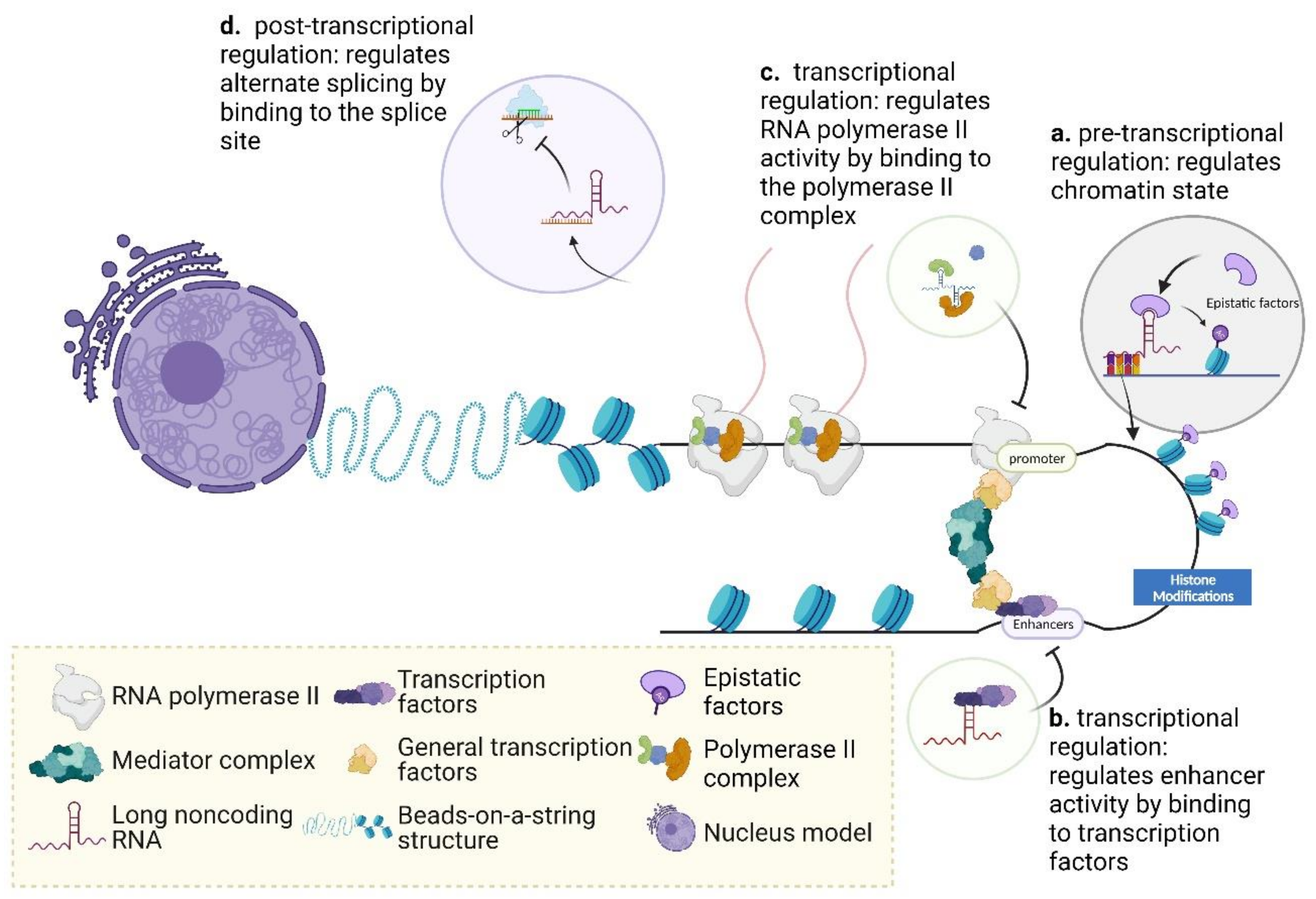
2.2. The Regulatory Mechanisms of Long Noncoding RNAs
3. The Role of LncRNAs in Spermatogenesis
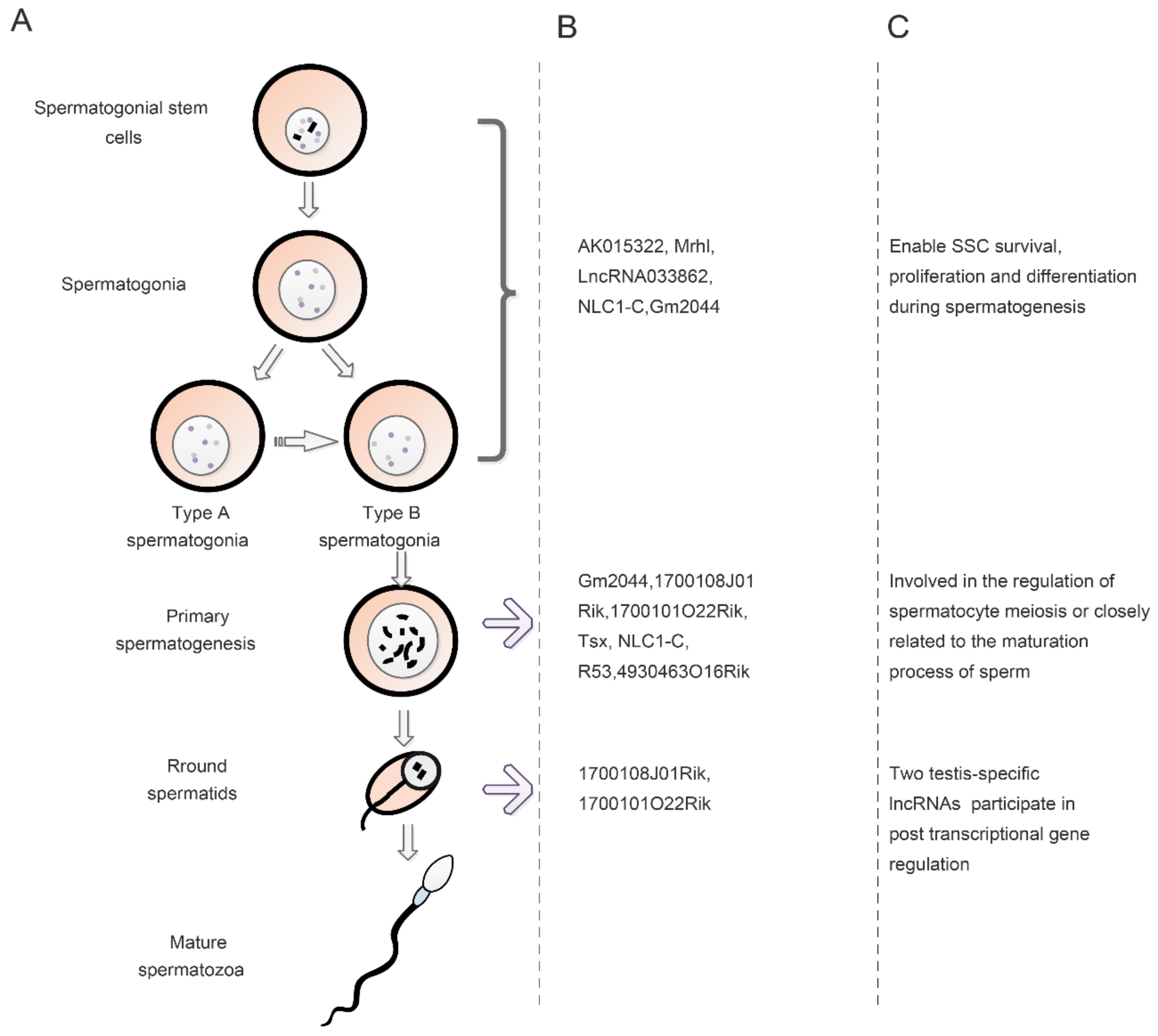
3.1. Regulation of LncRNAs in Spermatogonial Stem Cells
3.2. Regulation of LncRNAs in Meiosis and Spermatogenesis
3.3. Interactions between LncRNAs, miRNA, and mRNAs during Spermatogenesis
4. LncRNAs Are Promising Biomarkers for Dysfunction in the Male Reproductive System
4.1. LncRNAs in Livestock
4.2. The Role of LncRNAs in Model Animals
4.3. LncRNAs in Humans
5. Intergenerational Inheritance of LncRNAs
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hutchins, A.P.; Pei, D. Transposable elements at the center of the crossroads between embryogenesis, embryonic stem cells, reprogramming, and long non-coding RNAs. Sci. Bull. 2015, 60, 1722–1733. [Google Scholar] [CrossRef]
- Qu, Z.; Adelson, D.L. Evolutionary conservation and functional roles of ncRNA. Front. Genet. 2012, 3, 205. [Google Scholar] [CrossRef] [PubMed]
- Jathar, S.; Kumar, V.; Srivastava, J.; Tripathi, V. Technological Developments in lncRNA Biology. Adv. Exp. Med. Biol. 2017, 1008, 283–323. [Google Scholar] [CrossRef]
- Yoneda, R.; Ueda, N.; Uranishi, K.; Hirasaki, M.; Kurokawa, R. Long noncoding RNA pncRNA-D reduces cyclin D1 gene expression and arrests cell cycle through RNA m6A modification. J. Biol. Chem. 2020, 295, 5626–5639. [Google Scholar] [CrossRef]
- Li, D.; Yang, Y.; Li, Y.; Zhu, X.; Li, Z. Epigenetic regulation of gene expression in response to environmental exposures: From bench to model. Sci. Total. Environ. 2021, 776, 145998. [Google Scholar] [CrossRef]
- Klastrup, L.K.; Bak, S.T.; Nielsen, A.L. The influence of paternal diet on sncRNA-mediated epigenetic inheritance. Mol. Genet. Genom. 2019, 294, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Ma, Y.; Li, Y.; Li, P.; Cheng, Z.; Li, H.; Zhang, L.; Tang, Z. The comprehensive detection of miRNA, lncRNA, and circRNA in regulation of mouse melanocyte and skin development. Biol. Res. 2020, 53, 4. [Google Scholar] [CrossRef]
- Bie, B.; Wang, Y.; Li, L.; Fang, H.; Liu, L.; Sun, J. Noncoding RNAs: Potential players in the self-renewal of mammalian spermatogonial stem cells. Mol. Reprod. Dev. 2018, 85, 720–728. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, A.V.; Barciszewska, Z.M.; Szymanski, M.; Hochberg, A.; Groot, N.D.; Barciszewski, J. The non-coding RNAs as riboregulators. Nucleic Acids Res. 2001, 29, 189–193. [Google Scholar] [CrossRef]
- Taylor, D.H.; Chu, E.T.; Spektor, R.; Soloway, P.D. Long non-coding RNA regulation of reproduction and development. Mol. Reprod. Dev. 2015, 82, 932–956. [Google Scholar] [CrossRef] [PubMed]
- McSwiggin, H.M.; O’Doherty, A.M. Epigenetic reprogramming during spermatogenesis and male factor infertility. Reproduction 2018, 156, R9–R21. [Google Scholar] [CrossRef]
- Vra, B.; Dgv, A.; Mfr, A. Non-coding RNA regulation in reproduction: Their potential use as biomarkers. Non-Coding RNA Res. 2019, 4, 54–62. [Google Scholar] [CrossRef]
- Bourhis, D.L.; Idrissi, S.J.; Mermillod, P.; Carmen, A.; Dizier, M.S. 65 Effect of oviductal fluid extracellular vesicle supplementation during invitro culture on development and quality of bovine embryos. Reprod. Fertil. Dev. 2020, 32, 158. [Google Scholar] [CrossRef]
- Kamalidehghan, B.; Habibi, M.; Afjeh, S.S.; Shoai, M.; Alidoost, S.; Almasi Ghale, R.; Eshghifar, N.; Pouresmaeili, F. The Importance of Small Non-Coding RNAs in Human Reproduction: A Review Article. Appl. Clin. Genet. 2020, 13, 1–11. [Google Scholar] [CrossRef]
- Taborska, E.; Pasulka, J.; Malik, R.; Horvat, F.; Jenickova, I.; Jelic Matosevic, Z.; Svoboda, P. Restricted and non-essential redundancy of RNAi and piRNA pathways in mouse oocytes. PLoS Genet. 2019, 15, e1008261. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.G.; He, N.; Wang, F.; Van Iperen, L.; Eguizabal, C.; Matorras, R.; Roelen, B.A.J.; De Sousa Lopes, S.M.C. Human-specific subcellular compartmentalization of P-element induced wimpy testis-like (PIWIL) granules during germ cell development and spermatogenesis. Hum. Reprod. 2018, 33, 258–269. [Google Scholar] [CrossRef]
- Song, X.; Kyi-Tha-Thu, C.; Takizawa, T.; Naing, B.T.; Takizawa, T. 1700108J01Rik and 1700101O22Rik are mouse testis-specific long non-coding RNAs. Histochem. Cell Biol. 2018, 149, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, M.; Wang, M.; Wu, X.; Geng, L.; Xue, Y.; Wei, X.; Jia, Y.; Wu, X. LncRNA analysis of mouse spermatogonial stem cells following glial cell-derived neurotrophic factor treatment. Genom. Data 2015, 5, 275–278. [Google Scholar] [CrossRef][Green Version]
- Mäkelä, J.; Hobbs, R. Molecular regulation of spermatogonial stem cell renewal and differentiation. Reproduction 2019, 158, 169–187. [Google Scholar] [CrossRef]
- Sahlu, B.W.; Zhao, S.; Wang, X.; Umer, S.; Zou, H.; Huang, J.; Zhu, H. Long noncoding RNAs: New insights in modulating mammalian spermatogenesis. J. Anim. Sci. Biotechnol. 2020, 11, 16. [Google Scholar] [CrossRef]
- Gou, L.T.; Dai, P.; Liu, M.F. Small noncoding RNAs and male infertility. Wiley Interdiscip. Rev. RNA 2014, 5, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Wichman, L.; Somasundaram, S.; Breindel, C.; Valerio, D.M.; McCarrey, J.R.; Hodges, C.A.; Khalil, A.M. Dynamic expression of long noncoding RNAs reveals their potential roles in spermatogenesis and fertility. Biol. Reprod. 2017, 97, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.; Rajender, S. Long non-coding RNAs (lncRNAs) in spermatogenesis and male infertility. Reprod. Biol. Endocrinol. 2020, 18, 103. [Google Scholar] [CrossRef]
- Lee, C.; Kikyo, N. Strategies to identify long noncoding RNAs involved in gene regulation. Cell Biosci. 2012, 2, 37. [Google Scholar] [CrossRef]
- Liu, X.Q.; Li, B.X.; Zeng, G.R.; Liu, Q.Y.; Ai, D.M. Prediction of Long Non-Coding RNAs Based on Deep Learning. Genes 2019, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Laurent, G.S.; Wahlestedt, C.; Kapranov, P. The Landscape of long noncoding RNA classification. Trends Genet. 2015, 31, 239–251. [Google Scholar] [CrossRef]
- Hudson, W.H.; Ortlund, E.A. The structure, function and evolution of proteins that bind DNA and RNA. Nat. Rev. Mol. Cell Biol. 2014, 15, 749–760. [Google Scholar] [CrossRef]
- Sanchita; Trivedi, P.K.; Asif, M.H. Updates on plant long non-coding RNAs (lncRNAs): The regulatory components. Plant Cell Tissue Organ Cult. 2020, 140, 259–269. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Pei, W.; Zhang, M.; Yang, H.; Zhong, M.; Kong, X.; Xu, Y.; Zhu, X.; Chen, T.; et al. LncRNA Dnmt3aos regulates Dnmt3a expression leading to aberrant DNA methylation in macrophage polarization. FASEB J. 2020, 34, 5077–5091. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Lei, P.; Wen, T. LncRNA MALAT1 shuttled by bone marrow-derived mesenchymal stem cells-secreted exosomes alleviates osteoporosis through mediating microRNA-34c/SATB2 axis. Aging 2019, 11, 8777–8791. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Wang, X.; Youmans, D.T.; Cech, T.R. How do lncRNAs regulate transcription? Sci. Adv. 2017, 3, eaao2110. [Google Scholar] [CrossRef]
- Su, Z.D.; Yan, H.; Zhang, Z.Y.; Zhao, Y.W.; Dong, W.; Wei, C.; Kuo-Chen, C.; Hao, L. iLoc-lncRNA: Predict the subcellular location of lncRNAs by incorporating octamer composition into general PseKNC. Bioinformatics 2018, 34, 4196–4204. [Google Scholar] [CrossRef] [PubMed]
- Bo, H.; Liu, Z.; Zhu, F.; Zhou, D.; Tan, Y.; Zhu, W.; Fan, L. Long noncoding RNAs expression profile and long noncoding RNA-mediated competing endogenous RNA network in nonobstructive azoospermia patients. Epigenomics 2020, 12, 673–684. [Google Scholar] [CrossRef]
- Chadourne, M.; Poumerol, E.; Jouneau, L.; Passet, B.; Castille, J.; Sellem, E.; Pailhoux, E.; Mandon-Pepin, B. Structural and Functional Characterization of a Testicular Long Non-coding RNA (4930463O16Rik) Identified in the Meiotic Arrest of the Mouse Topaz1 (-/-) Testes. Front. Cell Dev. Biol. 2021, 9, 700290. [Google Scholar] [CrossRef]
- Dai, Y.B.; Lin, Y.; Song, N.; Sun, F. LncRNA4667 is dispensable for spermatogenesis and fertility in mice. Reprod. Dev. Med. 2019, 3, 18. [Google Scholar] [CrossRef]
- He, L.; He, Q.; Qiao, L.; Huang, S.; Dai, Z.; Yang, T.; Liu, L.; Zhao, Z. LncWNT3-IT affects the proliferation of Sertoli cells by regulating the expression of the WNT3 gene in goat testis. Reprod. Domest. Anim. 2020, 55, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Laiho, A.; Kotaja, N.; Gyenesei, A.; Sironen, A. Transcriptome profiling of the murine testis during the first wave of spermatogenesis. PLoS ONE 2013, 8, e61558. [Google Scholar] [CrossRef]
- Sun, J.; Wu, J. Expression profiling of long noncoding RNAs in neonatal and adult mouse testis. Data Brief 2015, 4, 322–327. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, X.; Yang, C.; Guo, F.; Zhang, Y.; Ju, Z.; Jiang, Q.; Zhao, X.; Liu, Y.; Zhao, H.; Wang, J.; et al. Integrated analysis of mRNAs and long noncoding RNAs in the semen from Holstein bulls with high and low sperm motility. Sci. Rep. 2019, 9, 2092. [Google Scholar] [CrossRef]
- Ing, N.H.; Konganti, K.; Ghaffari, N.; Johnson, C.D.; Forrest, D.W.; Love, C.C.; Varner, D.D. Identification and quantification of coding and long non-coding RNAs in stallion spermatozoa separated by density. Andrology 2020, 8, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Mao, X.; Chen, Y.; Li, T.; Ton, H. Regulatory role of long non-coding RNAs during reproductive disease. Am. J. Transl. Res. 2018, 10, 1–12. [Google Scholar] [PubMed]
- Elango, R.; Vishnubalaji, R.; Shaath, H.; Alajez, N.M. Transcriptional alterations of protein coding and noncoding RNAs in triple negative breast cancer in response to DNA methyltransferases inhibition. Cancer Cell Int. 2021, 21, 515. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Chen, L.; Yan, C.; Endo, Y.; Mi, B.; Liu, G. The lncRNA Rhno1/miR-6979-5p/BMP2 Axis Modulates Osteoblast Differentiation. Int. J. Biol. Sci. 2020, 16, 1604–1615. [Google Scholar] [CrossRef]
- Zhao, W.; Geng, D.; Li, S.; Chen, Z.; Sun, M. LncRNA HOTAIR influences cell growth, migration, invasion, and apoptosis via the miR-20a-5p/HMGA2 axis in breast cancer. Cancer Med. 2018, 7, 842–855. [Google Scholar] [CrossRef]
- Yan, P.; Lu, J.Y.; Niu, J.; Gao, J.; Zhang, M.Q.; Yin, Y.; Shen, X. LncRNA Platr22 promotes super-enhancer activity and stem cell pluripotency. J. Mol. Cell Biol. 2021, 13, 295–313. [Google Scholar] [CrossRef]
- Dossin, F.; Pinheiro, I.; Zylicz, J.J.; Roensch, J.; Collombet, S.; Le Saux, A.; Chelmicki, T.; Attia, M.; Kapoor, V.; Zhan, Y.; et al. SPEN integrates transcriptional and epigenetic control of X-inactivation. Nature 2020, 578, 455–460. [Google Scholar] [CrossRef]
- Wang, C.; Duan, Y.; Duan, G.; Wang, Q.; Zhang, K.; Deng, X.; Qian, B.; Gu, J.; Ma, Z.; Zhang, S.; et al. Stress Induces Dynamic, Cytotoxicity-Antagonizing TDP-43 Nuclear Bodies via Paraspeckle LncRNA NEAT1-Mediated Liquid-Liquid Phase Separation. Mol. Cell 2020, 79, 443–458.e7. [Google Scholar] [CrossRef]
- Ali, T.; Grote, P. Beyond the RNA-dependent function of LncRNA genes. eLife 2020, 9, e60583. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, X. The functions of LncRNA in the heart. Diabetes Res. Clin. Pract. 2020, 168, 108249. [Google Scholar] [CrossRef]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Gao, L.; Wang, Y.; Chiu, D.K.; Wang, T.; Deng, Y. Advances in long noncoding RNAs: Identification, structure prediction and function annotation. Brief Funct. Genom. 2016, 15, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tang, Y.; Xiong, F.; He, Y.; Wei, F.; Zhang, S.; Guo, C.; Xiang, B.; Zhou, M.; Xie, N.; et al. LncRNAs regulate cancer metastasis via binding to functional proteins. Oncotarget 2018, 9, 1426–1443. [Google Scholar] [CrossRef]
- Gao, N.; Li, Y.; Li, J.; Gao, Z.; Fan, T. Long Non-Coding RNAs: The Regulatory Mechanisms, Research Strategies, and Future Directions in Cancers. Front. Oncol. 2020, 10, 2903. [Google Scholar] [CrossRef] [PubMed]
- Juvvuna, P.K.; Mondal, T.; Marco, M.; Kosalai, S.T.; Kanduri, C. NBAT1/CASC15-003/USP36 control MYCN expression and its downstream pathway genes in neuroblastoma. Neuro-Oncol. Adv. 2021, 3, vdab056. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.J.; Hu, F.; He, S.Y.; Wang, Y. Epigenetic landscape analysis of lncRNAs in acute myeloid leukemia with DNMT3A mutations. Ann. Transl. Med. 2020, 8, 318. [Google Scholar] [CrossRef]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Bhattacharyya, N.; Pandey, V.; Bhattacharyya, M.; Dey, A. Regulatory role of long non coding RNAs (lncRNAs) in neurological disorders: From novel biomarkers to promising therapeutic strategies. Asian J. Pharm. Sci. 2021, 16, 533–550. [Google Scholar] [CrossRef] [PubMed]
- Bolha, L.; Ravnik-Glavac, M.; Glavac, D. Long Noncoding RNAs as Biomarkers in Cancer. Dis. Markers 2017, 2017, 7243968. [Google Scholar] [CrossRef] [PubMed]
- Mazar, J.; Wei, Z.; Khalil, A.M.; Lee, B.; Perera, R.J. The functional characterization of long noncoding RNA SPRY4-IT1 in human melanoma cells. Oncotarget 2014, 5, 8959–8969. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Cao, G.; Jing, L.; Lin, S.; Wang, X.; Zhang, J.; Wang, M.; Liu, W.; Lv, C. Analysing the relationship between lncRNA and protein-coding gene and the role of lncRNA as ceRNA in pulmonary fibrosis. J. Cell. Mol. Med. 2014, 18, 991–1003. [Google Scholar] [CrossRef]
- Wang, W.; Zhuang, Q.; Ji, K.; Wen, B.; Lin, P.; Zhao, Y.; Li, W.; Yan, C. Identification of miRNA, lncRNA and mRNA-associated ceRNA networks and potential biomarker for MELAS with mitochondrial DNA A3243G mutation. Sci. Rep. 2017, 7, 41639. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, H.; Wu, C.; Yan, M.; Wu, H.; Wang, J. Construction and investigation of lncRNA-associated ceRNA regulatory network in papillary thyroid cancer. Oncol. Rep. 2018, 39, 1197–1206. [Google Scholar] [CrossRef]
- Yu, G.; Li, S.; Liu, P.; Shi, Y.; Zhu, W. LncRNA TUG1 functions as a ceRNA for miR-6321 to promote endothelial progenitor cell migration and differentiation. Exp. Cell Res. 2020, 388, 111839. [Google Scholar] [CrossRef] [PubMed]
- Herriges, M.J.; Swarr, D.T.; Morley, M.P.; Rathi, K.S.; Peng, T.; Stewart, K.M.; Morrisey, E.E. Long noncoding RNAs are spatially correlated with transcription factors and regulate lung development. Genes Dev. 2014, 28, 1363. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Lee, E.S. Non-coding RNA: What is functional and what is junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef]
- Graham, D.B.; Xavier, R.J. From Genetics of Inflammatory Bowel Disease Towards Mechanistic Insights. Trends Immunol. 2013, 34, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Kazemzadeh, M.; Safaralizadeh, R.; Orang, A.V. LncRNAs: Emerging players in gene regulation and disease pathogenesis. J. Genet. 2015, 94, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Shahid, Z.; Simpson, B.; Miao, K.H.; Singh, G. Genetics, Histone Code. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Stahl, F.; Lepple-Wienhues, A.; Kuppinger, M.; Tamm, E.; Wiederholt, M. Electrogenic sodium-bicarbonate cotransport in human ciliary muscle cells. Am. J. Physiol. 1992, 262, C427–C435. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Histone modifications for human epigenome analysis. J. Hum. Genet. 2013, 58, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, C.T.; Sandstrom, R.S.; Huang, S.A.; Wang, Y.; Schotta, G.; Berger, M.S.; Lin, C.A. Cross-species Analyses Unravel the Complexity of H3K27me3 and H4K20me3 in the Context of Neural Stem Progenitor Cells. Neuroepigenetics 2016, 6, 10–25. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Cho, S.W.; Xu, J.; Sun, R.; Mumbach, M.R.; Carter, A.C.; Chen, Y.G.; Yost, K.E.; Kim, J.; He, J.; Nevins, S.A.; et al. Promoter of lncRNA Gene PVT1 Is a Tumor-Suppressor DNA Boundary Element. Cell 2018, 173, 1398–1412.e22. [Google Scholar] [CrossRef]
- Wang, F.W.; Cao, C.H.; Han, K.; Zhao, Y.X.; Cai, M.Y.; Xiang, Z.C.; Zhang, J.X.; Chen, J.W.; Zhong, L.P.; Huang, Y.; et al. APC-activated long noncoding RNA inhibits colorectal carcinoma pathogenesis through reduction of exosome production. J. Clin. Investig. 2019, 129, 727–743. [Google Scholar] [CrossRef]
- Feng, J.; Yang, G.; Liu, Y.; Gao, Y.; Zhao, M.; Bu, Y.; Yuan, H.; Yuan, Y.; Yun, H.; Sun, M.; et al. LncRNA PCNAP1 modulates hepatitis B virus replication and enhances tumor growth of liver cancer. Theranostics 2019, 9, 5227–5245. [Google Scholar] [CrossRef] [PubMed]
- Grixti, J.M.; Ayers, D. Long noncoding RNAs and their link to cancer. Noncoding RNA Res. 2020, 5, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Requena, J.M. Lights and shadows on gene organization and regulation of gene expression in Leishmania. Front. Biosci. 2011, 16, 2069–2085. [Google Scholar] [CrossRef] [PubMed]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Chinen, M.; Tani, T. Diverse functions of nuclear non-coding RNAs in eukaryotic gene expression. Front. Biosci. 2012, 17, 1402–1417. [Google Scholar] [CrossRef]
- Lee, S.; Kopp, F.; Chang, T.C.; Sataluri, A.; Chen, B.; Sivakumar, S.; Yu, H.; Xie, Y.; Mendell, J.T. Noncoding RNA NORAD Regulates Genomic Stability by Sequestering PUMILIO Proteins. Cell 2016, 164, 69–80. [Google Scholar] [CrossRef]
- Khawar, M.B.; Mehmood, R.; Roohi, N. MicroRNAs: Recent insights towards their role in male infertility and reproductive cancers. Bosn. J. Basic Med. Sci. 2019, 19, 31. [Google Scholar] [CrossRef]
- O’Donnell, L.; Robertson, K.M.; Jones, M.E.; Simpson, E.R. Estrogen and Spermatogenesis 1. Endocr. Rev. 2001, 22, 289–318. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, A.; Anand, S.; Bhartiya, D. Altered Biology of Testicular VSELs and SSCs by Neonatal Endocrine Disruption Results in Defective Spermatogenesis, Reduced Fertility and Tumor Initiation in Adult Mice. Stem Cell Rev. Rep. 2020, 16, 893–908. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, X.; Li, H.; Cai, G.; Wu, Z. Intratesticular injection of busulfan for producing recipient male pigs for spermatogonial stem cell transplantation. Livest. Sci. 2021, 245, 104448. [Google Scholar] [CrossRef]
- Gao, W.; Zhang, C.; Jin, K.; Zhang, Y.; Zuo, Q.; Li, B. Analysis of lncRNA Expression Profile during the Formation of Male Germ Cells in Chickens. Animals 2020, 10, 1850. [Google Scholar] [CrossRef]
- Liang, M.; Li, W.; Tian, H.; Hu, T.; Wang, L.; Lin, Y.; Li, Y.; Huang, H.; Sun, F. Sequential expression of long noncoding RNA as mRNA gene expression in specific stages of mouse spermatogenesis. Sci. Rep. 2014, 4, 5966. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Hu, K.; He, C.; Zhou, J.; Liao, Y. Upregulated lncRNA Gm2044 inhibits male germ cell development by acting as miR-202 host gene. Anim. Cells Syst. 2019, 23, 128–134. [Google Scholar] [CrossRef]
- Hu, K.; Li, L.; Liao, Y.; Liang, M. LncRNA Gm2044 highly expresses in spermatocyte and inhibits Utf1 translation by interacting with Utf1 mRNA. Genes Genom. 2018, 40, 781–787. [Google Scholar] [CrossRef]
- Hu, K.; Zhang, J.; Liang, M. LncRNA AK015322 promotes proliferation of spermatogonial stem cell C18-4 by acting as a decoy for microRNA-19b-3p. In Vitro Cell Dev. Biol. Anim. 2017, 53, 277–284. [Google Scholar] [CrossRef]
- Aiyama, Y.; Tsunekawa, N.; Kishi, K.; Kawasumi, M.; Suzuki, H.; Kanai-Azuma, M.; Kurohmaru, M.; Kanai, Y. A Niche for GFRalpha1-Positive Spermatogonia in the Terminal Segments of the Seminiferous Tubules in Hamster Testes. Stem Cells 2015, 33, 2811–2824. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, M.; Wu, X.; Geng, L.; Xue, Y.; Wei, X.; Jia, Y. A long non-coding RNA interacts with Gfra1 and maintains survival of mouse spermatogonial stem cells. Cell Death Dis. 2016, 7, e2140. [Google Scholar] [CrossRef]
- Nishant, K.T.; Ravishankar, H.; Rao, M.R. Characterization of a mouse recombination hot spot locus encoding a novel non-protein-coding RNA. Mol. Cell. Biol. 2004, 24, 5620–5634. [Google Scholar] [CrossRef] [PubMed]
- Kataruka, S.; Akhade, V.S.; Kayyar, B.; Rao, M. Mrhl lncRNA mediates meiotic commitment of mouse spermatogonial cells by regulating Sox8 expression. Mol. Cell. Biol. 2017, 37, e00632-16. [Google Scholar] [CrossRef] [PubMed]
- Suresh, A.V.; Nivrutti, D.S.; Shubhangini, K.; Manchanahalli, R.; Satyanarayana, R. Mechanism of Wnt signaling induced down regulation of mrhl long non-coding RNA in mouse spermatogonial cells. Nucleic Acids Res. 2016, 44, 387–401. [Google Scholar] [CrossRef]
- Lu, M.; Tian, H.; Cao, Y.X.; He, X.; Chen, L.; Song, X.; Ping, P.; Huang, H.; Sun, F. Downregulation of miR-320a/383-sponge-like long non-coding RNA NLC1-C (narcolepsy candidate-region 1 genes) is associated with male infertility and promotes testicular embryonal carcinoma cell proliferation. Cell Death Dis. 2015, 6, e1960. [Google Scholar] [CrossRef] [PubMed]
- Anguera, M.C.; Ma, W.; Clift, D.; Namekawa, S.; Lee, J.T. Tsx Produces a Long Noncoding RNA and Has General Functions in the Germline, Stem Cells, and Brain. PLoS Genet. 2011, 7, e1002248. [Google Scholar] [CrossRef]
- Zhang, C.; Gao, L.; Xu, E.Y. LncRNA, a new component of expanding RNA-protein regulatory network important for animal sperm development. Semin. Cell Dev. Biol. 2016, 59, 110–117. [Google Scholar] [CrossRef]
- Nakajima, R.; Sato, T.; Ogawa, T.; Okano, H.; Noce, T. A noncoding RNA containing a SINE-B1 motif associates with meiotic metaphase chromatin and has an indispensable function during spermatogenesis. PLoS ONE 2017, 12, e0179585. [Google Scholar] [CrossRef] [PubMed]
- Saberiyan, M.; Mirfakhraie, R.; Moghni, M.; Teimori, H. Study of Linc00574 Regulatory Effect on the TCTE3 Expression in Sperm Motility. Reprod. Sci. 2021, 28, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Karakas, D.; Ozpolat, B. The Role of LncRNAs in Translation. Noncoding RNA 2021, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Xing, C.; Sun, S.G.; Yue, Z.Q.; Bai, F. Role of lncRNA LUCAT1 in cancer. Biomed. Pharmacother. 2021, 134, 111158. [Google Scholar] [CrossRef]
- Ma, L.; Sun, X.; Kuai, W.; Hu, J.; Yuan, Y.; Feng, W.; Lu, X. LncRNA SOX2 overlapping transcript acts as a miRNA sponge to promote the proliferation and invasion of Ewing’s sarcoma. Am. J. Transl. Res. 2019, 11, 3841–3849. [Google Scholar]
- Liu, Q.; Ran, R.; Song, M.; Li, X.; Wu, Z.; Dai, G.; Xia, R. LncRNA HCP5 acts as a miR-128-3p sponge to promote the progression of multiple myeloma through activating Wnt/beta-catenin/cyclin D1 signaling via PLAGL2. Cell Biol. Toxicol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhu, Y.T.; Chu, Q.J.; Wan, P.C.; Ou, Y.; Quan, S. Effects and mechanism of lncRNA serving as ceRNA in non-obstructive azoospermia. Zhonghua Yi Xue Za Zhi 2019, 99, 2761–2767. [Google Scholar] [CrossRef]
- Sha, Y.W.; Wang, X.; Su, Z.Y.; Wang, C.; Ji, Z.Y.; Mei, L.B.; Zhang, L.; Deng, B.B.; Huang, X.J.; Yan, W.; et al. TDRD6 is associated with oligoasthenoteratozoospermia by sequencing the patient from a consanguineous family. Gene 2018, 659, 84–88. [Google Scholar] [CrossRef]
- Liu, Y.; DeBoer, K.; de Kretser, D.M.; O’Donnell, L.; O’Connor, A.E.; Merriner, D.J.; Okuda, H.; Whittle, B.; Jans, D.A.; Efthymiadis, A.; et al. LRGUK-1 is required for basal body and manchette function during spermatogenesis and male fertility. PLoS Genet. 2015, 11, e1005090. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, H.; Han, L.; Li, F.; Zhang, T.; Pang, J.; Feng, X.; Ren, C.; Mao, S.; Wang, F. Long noncoding RNA expression profile changes associated with dietary energy in the sheep testis during sexual maturation. Sci. Rep. 2017, 7, 5180. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.C.; Zhang, Y.; Yu, K.; Li, Y.; Yu, H.; Zhou, S.J.; Wang, Y.P.; Deng, S.L.; Tian, L. LncRNAs induce oxidative stress and spermatogenesis by regulating endoplasmic reticulum genes and pathways. Aging 2021, 13, 13764–13787. [Google Scholar] [CrossRef]
- Lei, Q.; Pan, Q.; Li, N.; Zhou, Z.; Zhang, J.; He, X.; Peng, S.; Li, G.; Sidhu, K.; Chen, S.; et al. H19 regulates the proliferation of bovine male germline stem cells via IGF-1 signaling pathway. J. Cell Physiol. 2018, 234, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Ghazal, S.; McKinnon, B.; Zhou, J.; Mueller, M.; Men, Y.; Yang, L.; Mueller, M.; Flannery, C.; Huang, Y.; Taylor, H.S. H19 lncRNA alters stromal cell growth via IGF signaling in the endometrium of women with endometriosis. EMBO Mol. Med. 2015, 7, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Eggenschwiler, J.; Ludwig, T.; Fisher, P.; Leighton, P.A.; Tilghman, S.M.; Efstratiadis, A. Mouse mutant embryos overexpressing IGF-II exhibit phenotypic features of the Beckwith-Wiedemann and Simpson-Golabi-Behmel syndromes. Genes Dev. 1997, 11, 3128–3142. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, Q.; Zhang, Q.; Qu, X.; Dong, L.; Yang-Zom, C.; Zhuang, X.; Liu, H. Analysis of IGF2 mRNA expression and its methylation status between cattle yaks and their parents. Prog. Nat. Sci. 2009, 19, 1063–1069. [Google Scholar] [CrossRef]
- Moreira, D.P.; Melo, R.M.C.; Weber, A.A.; Rizzo, E. Insulin-like growth factors 1 and 2 are associated with testicular germ cell proliferation and apoptosis during fish reproduction. Reprod. Fertil. Dev. 2020, 32, 988–998. [Google Scholar] [CrossRef] [PubMed]
- An, L.F.; Huang, J.W.; Han, X.; Wang, J. Downregulation of lncRNA H19 sensitizes melanoma cells to cisplatin by regulating the miR-18b/IGF1 axis. Anti-Cancer Drugs 2020, 31, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Huat, T.J.; Khan, A.A.; Pati, S.; Mustafa, Z.; Abdullah, J.M.; Jaafar, H. IGF-1 enhances cell proliferation and survival during early differentiation of mesenchymal stem cells to neural progenitor-like cells. BMC Neurosci. 2014, 15, 91. [Google Scholar] [CrossRef] [PubMed]
- Kerr, G.E.; Young, J.C.; Horvay, K.; Abud, H.E.; Loveland, K.L. Regulated Wnt/beta-catenin signaling sustains adult spermatogenesis in mice. Biol. Reprod. 2014, 90, 3. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.J.; Ni, J.H.; Wang, L.M.; Tan, F.Q.; Yang, W.X. KIF3A regulates the Wnt/beta-catenin pathway via transporting beta-catenin during spermatogenesis in Eriocheir sinensis. Cell Tissue Res. 2020, 381, 527–541. [Google Scholar] [CrossRef]
- Zhao, Z.; Qiao, L.; Dai, Z.; He, Q.; Lan, X.; Huang, S.; He, L. LncNONO-AS regulates AR expression by mediating NONO. Theriogenology 2020, 145, 198–206. [Google Scholar] [CrossRef]
- Ran, M.; Huang, H.; Hu, B.; Hu, S.; Hu, J.; Li, L.; He, H.; Liu, H.; Wang, J. Comparative Analysis of Testicular Histology and lncRNA-mRNA Expression Patterns Between Landes Geese (Anser anser) and Sichuan White Geese (Anser cygnoides). Front. Genet 2021, 12, 627384. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, Y.; Li, Y.; Bai, H.; Xue, F.; Xu, S.; Xu, H.; Shi, L.; Yang, N.; Chen, J. Analyses of Long Non-Coding RNA and mRNA profiling using RNA sequencing in chicken testis with extreme sperm motility. Sci. Rep. 2017, 7, 9055. [Google Scholar] [CrossRef] [PubMed]
- Vining, B.; Ming, Z.; Bagheri-Fam, S.; Harley, V. Diverse Regulation but Conserved Function: SOX9 in Vertebrate Sex Determination. Genes 2021, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Richardson, N.; Albina, A.; Chaboissier, M.C.; Perea-Gomez, A. Mouse Gonad Development in the Absence of the Pro-Ovary Factor WNT4 and the Pro-Testis Factor SOX9. Cells 2020, 9, 1103. [Google Scholar] [CrossRef]
- Li, N.; Cheng, C.Y. Mammalian target of rapamycin complex (mTOR) pathway modulates blood-testis barrier (BTB) function through F-actin organization and gap junction. Histol. Histopathol. 2016, 31, 961–968. [Google Scholar] [CrossRef]
- Xu, H.; Shen, L.; Chen, X.; Ding, Y.; He, J.; Zhu, J.; Wang, Y.; Liu, X. mTOR/P70S6K promotes spermatogonia proliferation and spermatogenesis in Sprague Dawley rats. Reprod. Biomed. Online 2016, 32, 207–217. [Google Scholar] [CrossRef]
- Yoneda, R.; Takahashi, T.; Matsui, H.; Takano, N.; Hasebe, Y.; Ogiwara, K.; Kimura, A.P. Three testis-specific paralogous serine proteases play different roles in murine spermatogenesis and are involved in germ cell survival during meiosis. Biol. Reprod. 2013, 88, 118. [Google Scholar] [CrossRef] [PubMed]
- Satoh, Y.; Takei, N.; Kawamura, S.; Takahashi, N.; Kotani, T.; Kimura, A.P. A novel testis-specific long noncoding RNA, Tesra, activates the Prss42/Tessp-2 gene during mouse spermatogenesisdagger. Biol. Reprod. 2019, 100, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Lin, Y.; He, Y.; Wang, H.; Sun, F. Deletion of lncRNA5512 has no effect on spermatogenesis and reproduction in mice. Reprod. Fertil. Dev. 2020, 32, 706–713. [Google Scholar] [CrossRef]
- Liang, M.; Wang, H.; He, C.; Zhang, K.; Hu, K. LncRNA-Gm2044 is transcriptionally activated by A-MYB and regulates Sycp1 expression as a miR-335-3p sponge in mouse spermatocyte-derived GC-2spd(ts) cells. Differentiation 2020, 114, 49–57. [Google Scholar] [CrossRef]
- Sage, J.; Martin, L.; Meuwissen, R.; Heyting, C.; Cuzin, F.; Rassoulzadegan, M. Temporal and spatial control of the Sycp1 gene transcription in the mouse meiosis: Regulatory elements active in the male are not sufficient for expression in the female gonad. Mech. Dev. 1999, 80, 29–39. [Google Scholar] [CrossRef]
- Vries, D.; At, F. Mouse Sycp1 functions in synaptonemal complex assembly, meiotic recombination, and XY body formation. Genes Dev. 2005, 19, 1376–1389. [Google Scholar] [CrossRef]
- Tang, H.; Goldberg, E. A-MYB (MYBL1) stimulates murine testis-specific Ldhc expression via the cAMP-responsive element (CRE) site. Biol. Reprod. 2012, 86, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Toscani, A.; Mettus, R.V.; Coupland, R.; Simpkins, H.; Litvin, J.; Orth, J.; Hatton, K.S.; Reddy, E.P. Arrest of spermatogenesis and defective breast development in mice lacking A-myb. Nature 1997, 386, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, P.; Song, D.; Xiong, S.; Zhang, H.; Fu, J.; Gao, F.; Chen, H.; Zeng, X. Expression profiles and characteristics of human lncRNA in normal and asthenozoospermia sperm. Biol. Reprod. 2019, 100, 982–993. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.Z.; Vormer, T.L.; Jongejan, A.; Roling, M.D.; Silber, S.J.; de Rooij, D.G.; Hamer, G.; Repping, S.; van Pelt, A.M.M. Unraveling transcriptome dynamics in human spermatogenesis. Development 2017, 144, 3659–3673. [Google Scholar] [CrossRef] [PubMed]
- Rolland, A.D.; Evrard, B.; Darde, T.A.; Le Beguec, C.; Le Bras, Y.; Bensalah, K.; Lavoue, S.; Jost, B.; Primig, M.; Dejucq-Rainsford, N.; et al. RNA profiling of human testicular cells identifies syntenic lncRNAs associated with spermatogenesis. Hum. Reprod. 2019, 34, 1278–1290. [Google Scholar] [CrossRef]
- Ms, A.; Rm, B.; Dg, A.; Ld, C.; Ht, A. Investigating the regulatory function of the ANO1-AS2 on the ANO1 gene in infertile men with asthenozoospermia and terato-asthenozoospermia. Exp. Mol. Pathol. 2020, 117, 104528. [Google Scholar] [CrossRef]
- Carkci, S.; Etem, E.O.; Ozaydin, S.; Karakeci, A.; Tektemur, A.; Ozan, T.; Orhan, I. Ion channel gene expressions in infertile men: A case-control study. Int. J. Reprod. Biomed. 2017, 15, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Fullston, T.; Ohlsson Teague, E.M.; Palmer, N.O.; DeBlasio, M.J.; Mitchell, M.; Corbett, M.; Print, C.G.; Owens, J.A.; Lane, M. Paternal obesity initiates metabolic disturbances in two generations of mice with incomplete penetrance to the F2 generation and alters the transcriptional profile of testis and sperm microRNA content. FASEB J. 2013, 27, 4226–4243. [Google Scholar] [CrossRef] [PubMed]
- Gapp, K.; Jawaid, A.; Sarkies, P.; Bohacek, J.; Pelczar, P.; Prados, J.; Farinelli, L.; Miska, E.; Mansuy, I.M. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nat. Neurosci. 2014, 17, 667–669. [Google Scholar] [CrossRef]
- Hsiao, C.H.; Ji, A.T.; Chang, C.C.; Chien, M.H.; Lee, L.M.; Ho, J.H. Mesenchymal stem cells restore the sperm motility from testicular torsion-detorsion injury by regulation of glucose metabolism in sperm. Stem Cell Res. Ther. 2019, 10, 270. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.H.; Li, Y.W.; Xie, H.L.; Li, Q.; Dong, H.B.; Sun, M.J.; Gao, W.Q.; Tan, J.H. Effects of glucose metabolism pathways on sperm motility and oxidative status during long-term liquid storage of goat semen. Theriogenology 2016, 86, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Gorus, F.K.; Pipeleers, D.G. Glucose metabolism in human spermatozoa: Lack of insulin effects and dissociation from alloxan handling. J. Cell Physiol. 1986, 127, 261–266. [Google Scholar] [CrossRef]
- Yan, W.; Yue, H.; Ji, X.; Li, G.; Sang, N. Prenatal NO2 exposure and neurodevelopmental disorders in offspring mice: Transcriptomics reveals sex-dependent changes in cerebral gene expression. Environ. Int. 2020, 138, 105659. [Google Scholar] [CrossRef]
- Li, J.; Li, K.; Gao, J.; Guo, X.; Lu, M.; Li, Z.; Li, D. Maternal exposure to an n-3 polyunsaturated fatty acid diet decreases mammary cancer risk of female offspring in adulthood. Food Funct. 2018, 9, 5768–5777. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zuo, X.; Deng, H.; Liu, X.; Liu, L.; Ji, A. Roles of long noncoding RNAs in brain development, functional diversification and neurodegenerative diseases. Brain Res. Bull. 2013, 97, 69–80. [Google Scholar] [CrossRef]
- Vance, J.E.; Hayashi, H. Formation and function of apolipoprotein E-containing lipoproteins in the nervous system. Biochim. Biophys. Acta 2010, 1801, 806–818. [Google Scholar] [CrossRef] [PubMed]
- Sadler-Riggleman, I.; Klukovich, R.; Nilsson, E.; Beck, D.; Xie, Y.; Yan, W.; Skinner, M.K. Epigenetic transgenerational inheritance of testis pathology and Sertoli cell epimutations: Generational origins of male infertility. Environ. Epigenetics 2019, 5, dvz013. [Google Scholar] [CrossRef]
- Wu, S.; Divall, S.; Nwaopara, A.; Radovick, S.; Wondisford, F.; Ko, C.; Wolfe, A. Obesity-induced infertility and hyperandrogenism are corrected by deletion of the insulin receptor in the ovarian theca cell. Diabetes 2014, 63, 1270–1282. [Google Scholar] [CrossRef] [PubMed]
- Zippl, A.L.; Seeber, B.; Wildt, L. Obesity and infertility: Are hyperlipidemia and hyperinsulinemia the bad guys? Fertil. Steril. 2021, 116, 365–366. [Google Scholar] [CrossRef]
- An, T.; Zhang, T.; Teng, F.; Zuo, J.C.; Pan, Y.Y.; Liu, Y.F.; Miao, J.N.; Gu, Y.J.; Yu, N.; Zhao, D.D.; et al. Long non-coding RNAs could act as vectors for paternal heredity of high fat diet-induced obesity. Oncotarget 2017, 8, 47876–47889. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Ip, J.Y.; Shioi, G.; Tripathi, V.; Zong, X.; Hirose, T.; Prasanth, K.V. Malat1 is not an essential component of nuclear speckles in mice. RNA 2012, 18, 1487–1499. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.C.; Diermeier, S.D.; Yu, A.T.; Brine, L.D.; Spector, D.L. MaTAR25 LncRNA Regulates the Tensin1 Gene to Impact Breast Cancer Progression. Nat. Commun. 2020, 11, 6438. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
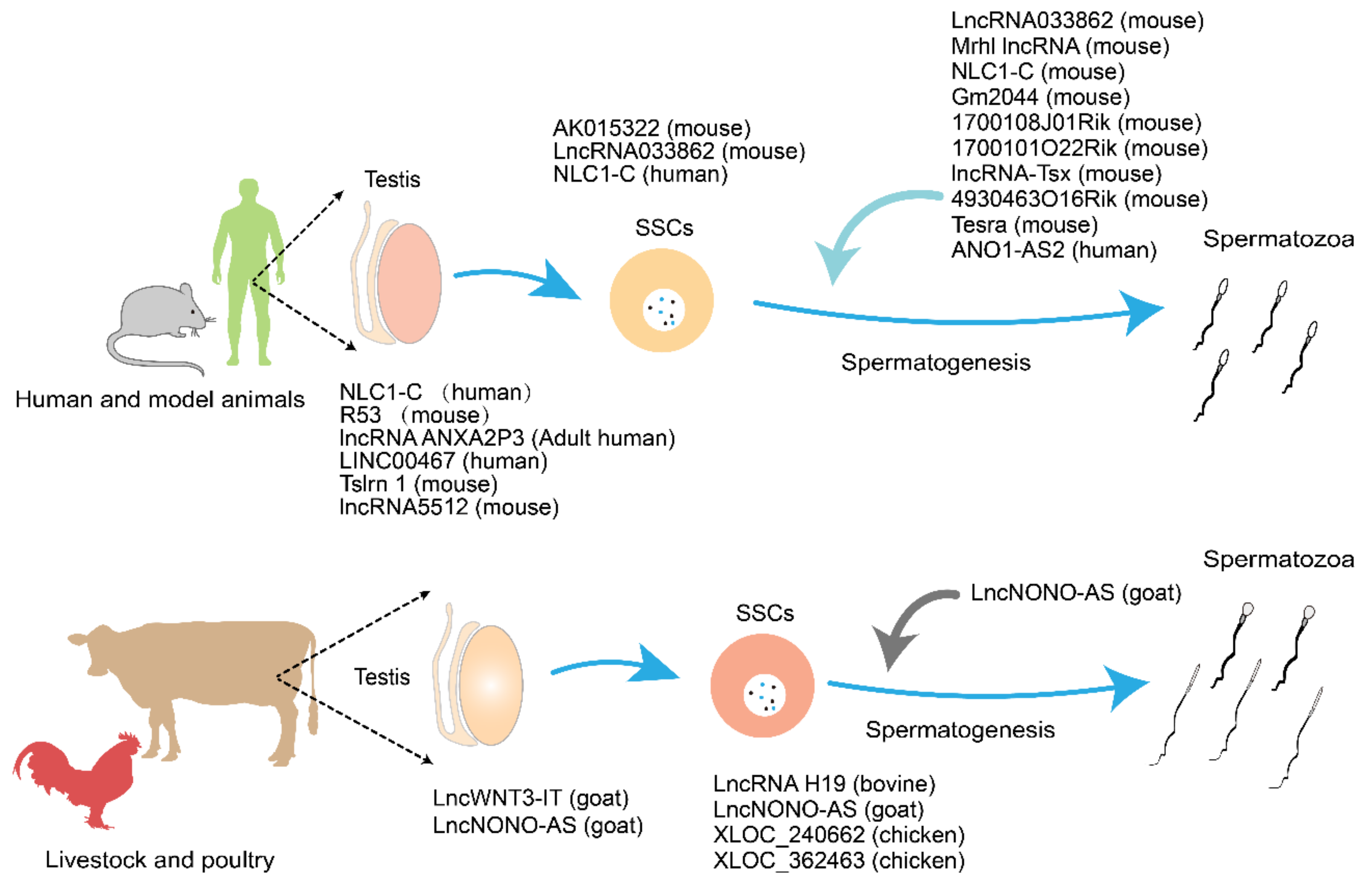
| LncRNA Name | Location | Cell Type | Description [Ref] |
|---|---|---|---|
| AK015322 | Chromosome 12, NC_000078.7 | Mouse SSCs line C18-4 | Highly expressed in spermatogonial stem cells; antagonizes the function of miR-19b-3 as a decoy; attenuates the repression of its endogenous target transcriptional factor, Ets-variant 5 (ETV5), which is a pivotal gene for SSC self-renewal [90]. |
| LncRNA033862 | Chromosome 19, NC_000085.7 | Mouse SSCs and early spermatogonial cells | An antisense transcript of the GDNF receptor alpha1 (Gfra1); highly expressed in SSCs and early spermatogonia; regulates Gfra1 expression level through interactions with Gfra1 chromatin; and maintains the self-renewal and survival of SSCs [92]. |
| Mrhl lncRNA | Chromosome 8 | Mouse spermatogonial cells | Important for meiotic progression and differentiation; and negatively regulates Wnt signaling. During differentiation of spermatogonial cells, it binds to SOX8 which directly regulates the expression of premeiotic and meiotic markers [94,95]. |
| NLC1-C | Chr 21 (NC_000021.9) | Human testicular spermatogonia and early spermatocytes | Knockout of the narcolepsy candidate 1 gene (NLC1-C) accelerates germ cell apoptosis, whereas its overexpression promotes germ cell proliferation [96]. |
| LncRNA Name | Location | Cell Type | Description [Ref] |
|---|---|---|---|
| AK015322 | Chr 12, NC_000078.7 | Mouse spermatogonial stem cells (SSCs) line C18-4 | Highly expressed in SSCs; antagonizes the function of miR-19b-3 as a decoy; attenuates the repression of its endogenous target transcriptional factor, Ets-variant 5 (ETV5), which is a pivotal gene for SSC self-renewal [90]. |
| LncRNA033862 | Chr 19, NC_000085.7 | Mouse SSCs and early spermatogonial cells | Antisense transcript of the GDNF receptor alpha1 (Gfra1); highly expressed in SSCs and early spermatogonia; regulates Gfra1 expression level through interactions with Gfra1 chromatin; maintains SSC self-renewal and survival [92]. |
| Mrhl lncRNA | Chr 8 | Mouse spermatogonial cells | Important for meiotic progression and differentiation; negatively regulates Wnt signaling. During differentiation of spermatogonial cells, it binds to SOX8, which directly regulates the expression of premeiotic and meiotic markers [94,95]. |
| NLC1-C | Chr 21 (NC_000021.9) | Human testicular spermatogonia and early spermatocytes | Knockout of the narcolepsy candidate 1 gene (NLC1-C) accelerates germ cell apoptosis, whereas its overexpression promotes germ cell proliferation [96]. |
| Gm2044 | Chr 7 | Mouse spermatocytes | Highly expressed in spermatogenesis; overexpression of lncRNA Gm2044 inhibits cell proliferation, and can be used as a miR-335-3p sponge to increase the expression levels of miR-335-3p direct target protein, Sycp1, which is expressed specifically in spermatocytes and regulates meiosis during spermatogenesis [88,89,129]. |
| 1700108J01Rik | Chr 14 (NC_000080.7) | Mouse round spermatids | A mouse testis-specific lncRNA; only expressed in testicular germ cells at the pre-meiotic and round sperm cell stages; involved in post-transcriptional gene regulation [17]. |
| 1700101O22Rik | Chr 12 (NC_000078.7) | Mouse round spermatids | A mouse testis-specific lncRNA; only expressed in testicular germ cells at the pre-meiotic and round sperm cell stages; involved in post-transcriptional gene regulation [17]. |
| lncRNA-Tsx | Chr X (NC_000086.8) | Spermatocytes | Located at the X-inactivation center and expressed in meiotic germ cells; knockout lncRNA-Tsx results in the apoptosis of pachytene spermatocytes [97]. |
| R53 | Chr 4 (NC_007090.3) | Mouse testis | Involved in regulation of the metaphase of meiosis; overexpression of lncRNA R53 inhibits the subsequent progress of meiosis [99]. |
| 4930463O16Rik | Chr 10 (NC_000076.7) | Mouse meiotic germ cells | Related to the protein expression of Topaz1, which is a germ cell-specific gene highly conserved in mammals; the absence of TOPAZ1 disturbs the expression of 4930463O16Rik, resulting in male infertility [34]. |
| lncRNA ANXA2P3 | Chromosome 10 (NC_000010.11) | Adult human testis | Binds with miR-613 and miR-206 to inhibit mRNA TKT expression [105]. |
| LINC00467 | Chr 1 | Human testis | A promising biomarker for male infertility; can positively regulate TDRD6 and LRGUK, both of which are key regulators of human spermatogenesis and maturation [33]. |
| Tslrn 1 | Chr X | Mouse testis | One of the X-linked lncRNAs, testis-specific long noncoding RNA 1 (Tslrn 1), knocking out Tslrn1 shows that males with deleted Tslrn1 show normal fertility, and have significantly reduced sperm count [22]. |
| lncRNA H19 | Chromosome 11 (NC_000011.10) | Bovine testes | Regulates the IGF-1 signaling pathway, which maintains the survival of a variety of stem cells, and participates in the proliferation and differentiation of male germ stem cells [110,111,115]. |
| lncWNT3-IT | Chr 17 | Goat testes | Overexpression of lncWNT3-IT increases the expression of WNT3, and up-regulates the proliferation of Sertoli cells [36]. |
| lncNONO-AS | Chr X (NC_000023.11) | Goat testis | Mainly expressed in the nucleus; regulates spermatogenesis and testicular development in goats by regulating androgen receptor (AR) expression [119]. |
| Tesra | Chr 1 | Mouse spermatogenesis | Nuclear Tesra regulates meiosis from secondary spermatocytes to round spermatids by increasing Prss42/Tessp-2 promoter activity through binding to the promoter [127]. |
| lncRNA5512 | - | Mouse testis | May be a useful biomarker for identifying spermatocytes and round sperm cells due to its specific location in these cells, although knockout does not affect spermatogenesis and fertility [128]. |
| ANO1-AS2 (linc02584) | Chr 11 | Human spermatogenesis | High expression of ANO1-AS2 down-regulates the ANO1 gene by interacting with ANO1 gene promoter, which influences sperm motility and morphology [137]. |
| Neat1 | Chr 11 (NC_000011.10) | Paternal line and offspring of mice | Located in the nucleosome; involved in spermatogenesis, and reduced expression of Neat1 which is associated with reduced semen quality and reduced fertility [151]. |
| Linc00574 | Chr 6 (NC_000006.12) | Human sperm | Transcribed from chr6q27 in the vicinity of TCTE3, regulated TCTE3 through a negative self-regulating mechanism [100,109]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Heng, N.; Weldegebriall Sahlu, B.; Wang, H.; Zhu, H. Long Noncoding RNAs: Recent Insights into Their Role in Male Infertility and Their Potential as Biomarkers and Therapeutic Targets. Int. J. Mol. Sci. 2021, 22, 13579. https://doi.org/10.3390/ijms222413579
Zhao S, Heng N, Weldegebriall Sahlu B, Wang H, Zhu H. Long Noncoding RNAs: Recent Insights into Their Role in Male Infertility and Their Potential as Biomarkers and Therapeutic Targets. International Journal of Molecular Sciences. 2021; 22(24):13579. https://doi.org/10.3390/ijms222413579
Chicago/Turabian StyleZhao, Shanjiang, Nuo Heng, Bahlibi Weldegebriall Sahlu, Huan Wang, and Huabin Zhu. 2021. "Long Noncoding RNAs: Recent Insights into Their Role in Male Infertility and Their Potential as Biomarkers and Therapeutic Targets" International Journal of Molecular Sciences 22, no. 24: 13579. https://doi.org/10.3390/ijms222413579
APA StyleZhao, S., Heng, N., Weldegebriall Sahlu, B., Wang, H., & Zhu, H. (2021). Long Noncoding RNAs: Recent Insights into Their Role in Male Infertility and Their Potential as Biomarkers and Therapeutic Targets. International Journal of Molecular Sciences, 22(24), 13579. https://doi.org/10.3390/ijms222413579