
Long-Term Sex- and Genotype-Specific Effects of 56Fe Irradiation on Wild-Type and APPswe/PS1dE9 Transgenic Mice

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. 56Fe Irradiation Decreased the Survival Rate of APP/PS1 Females but Did Not Affect Other Health Measures
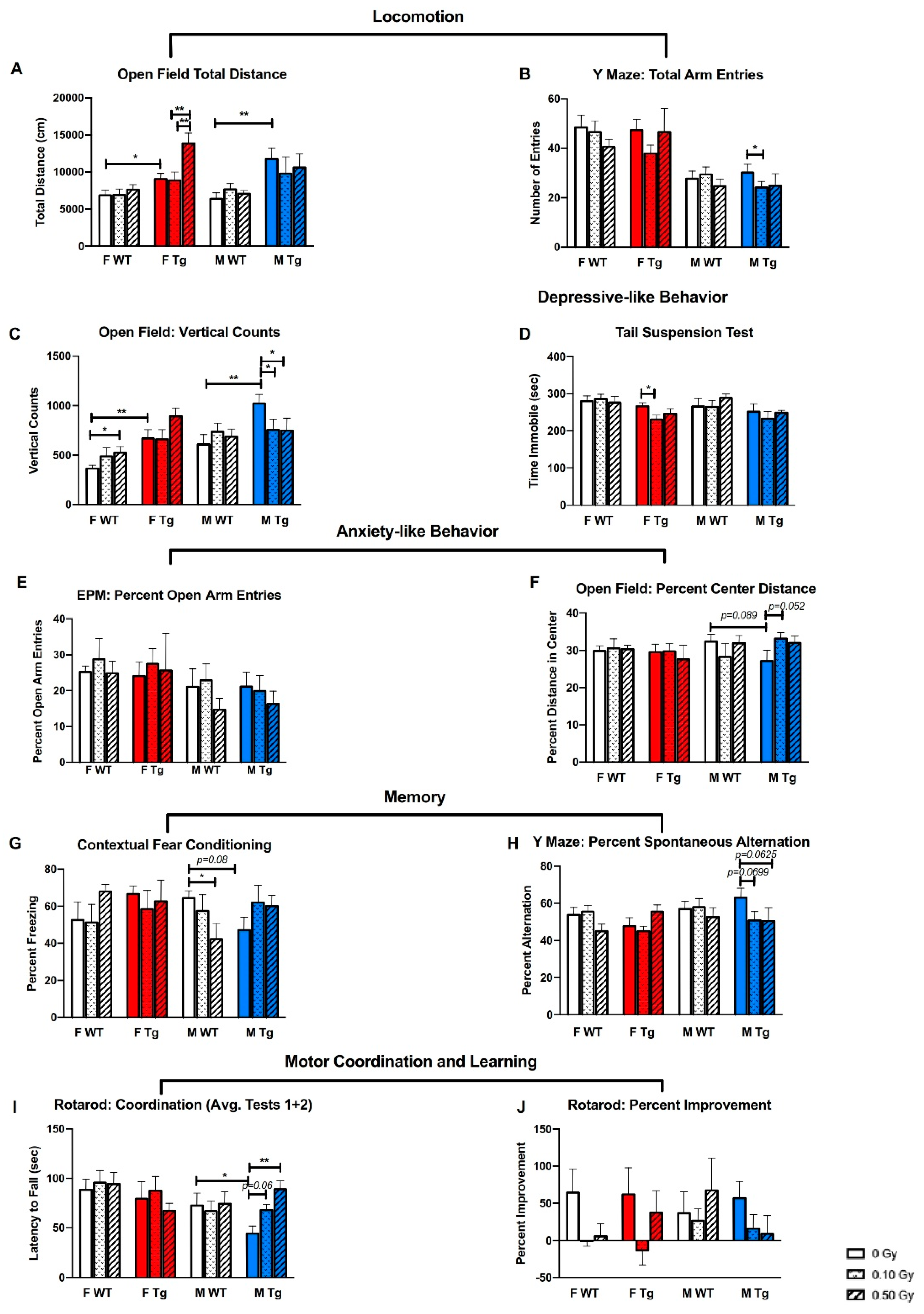
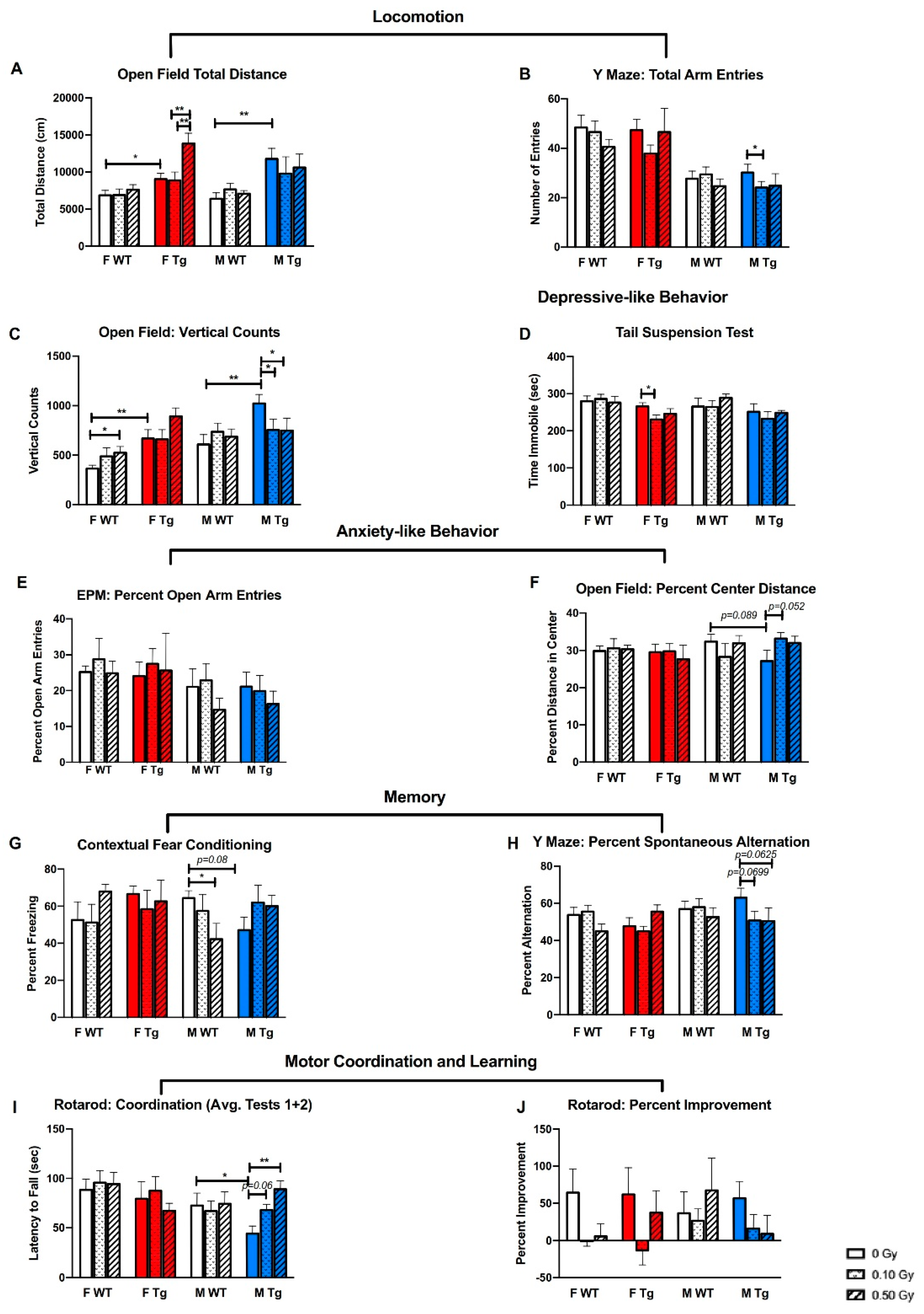
2.2. 56Fe Irradiation Caused More Behavioral Changes in Male Than in Female Mice
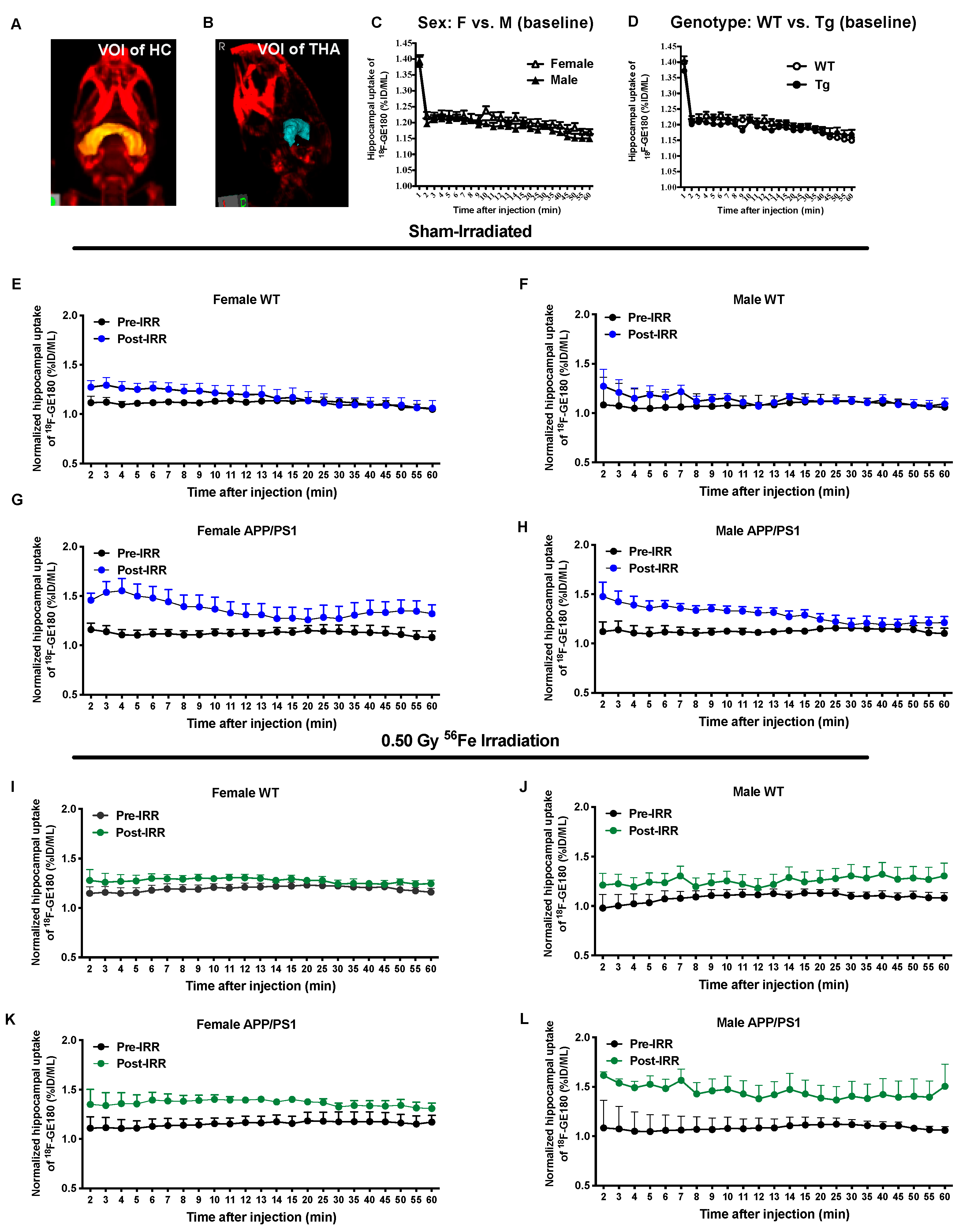
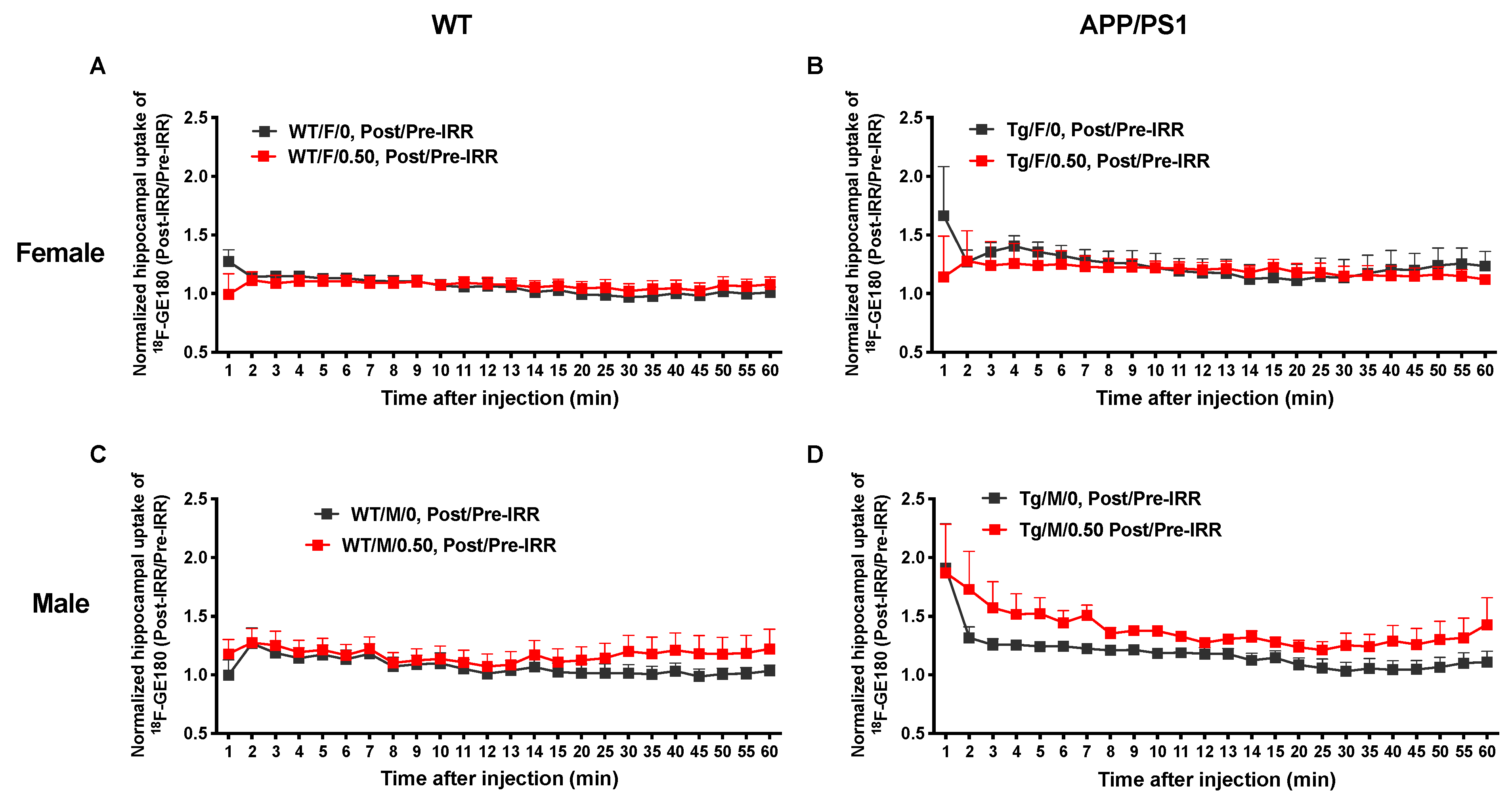
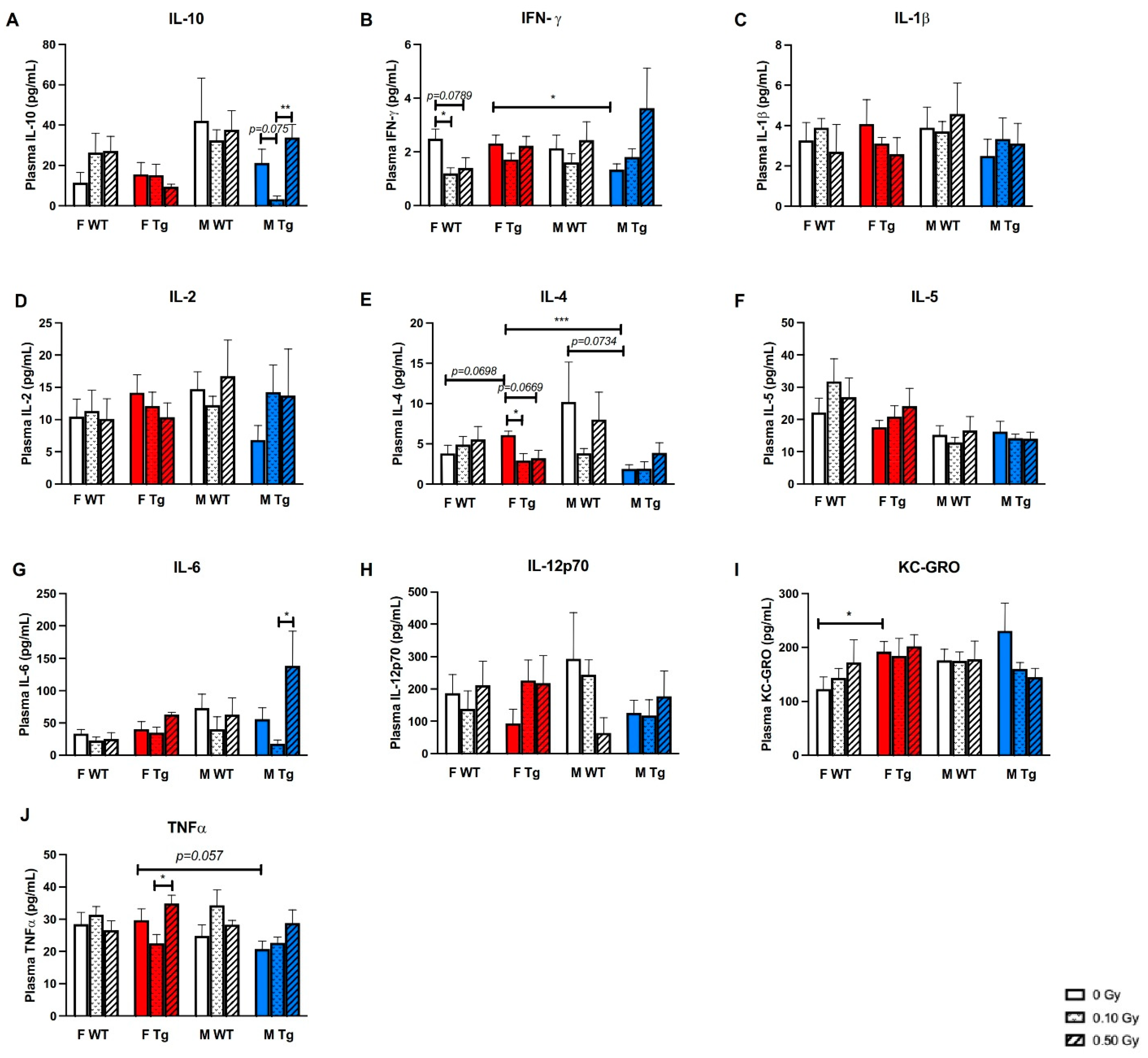
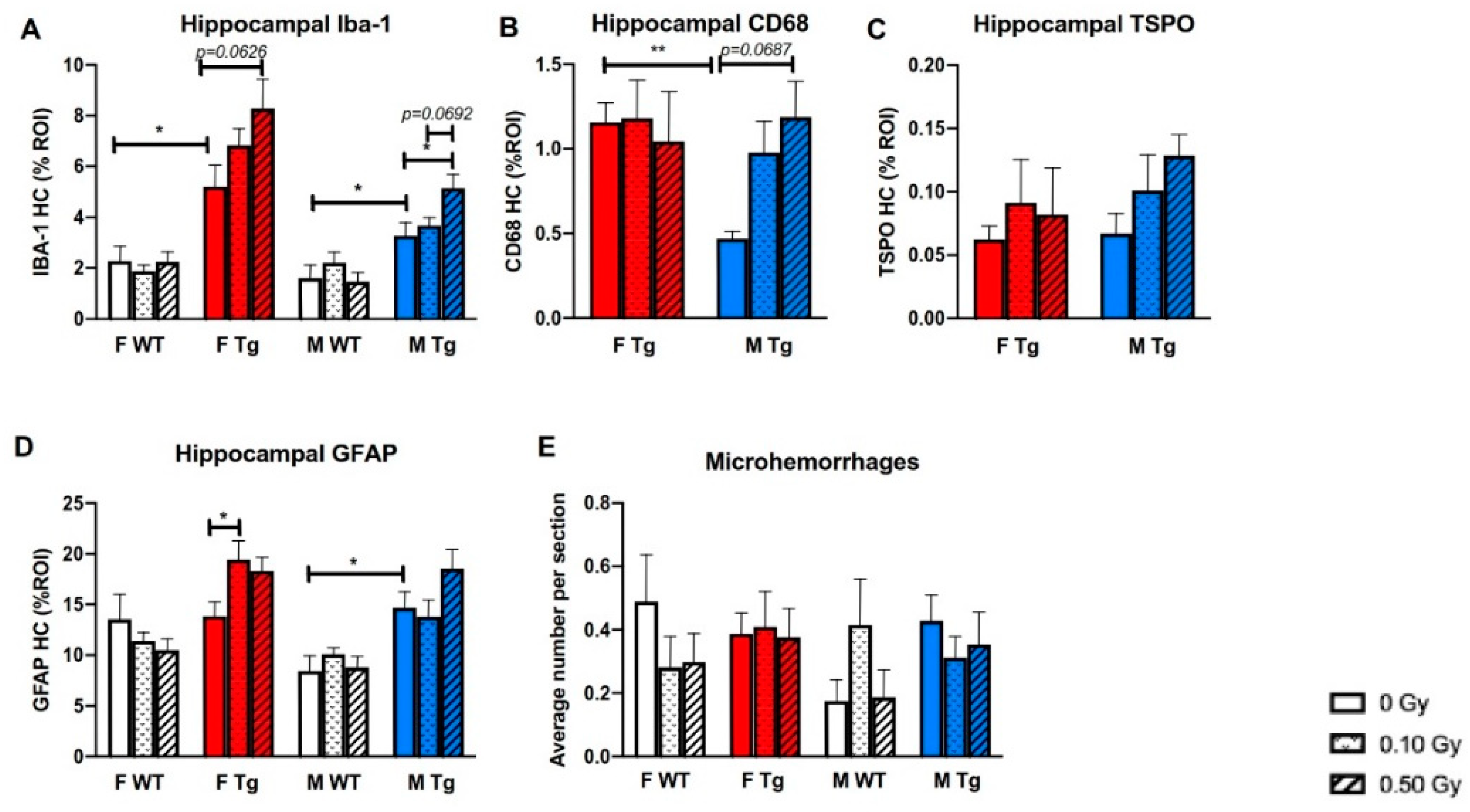
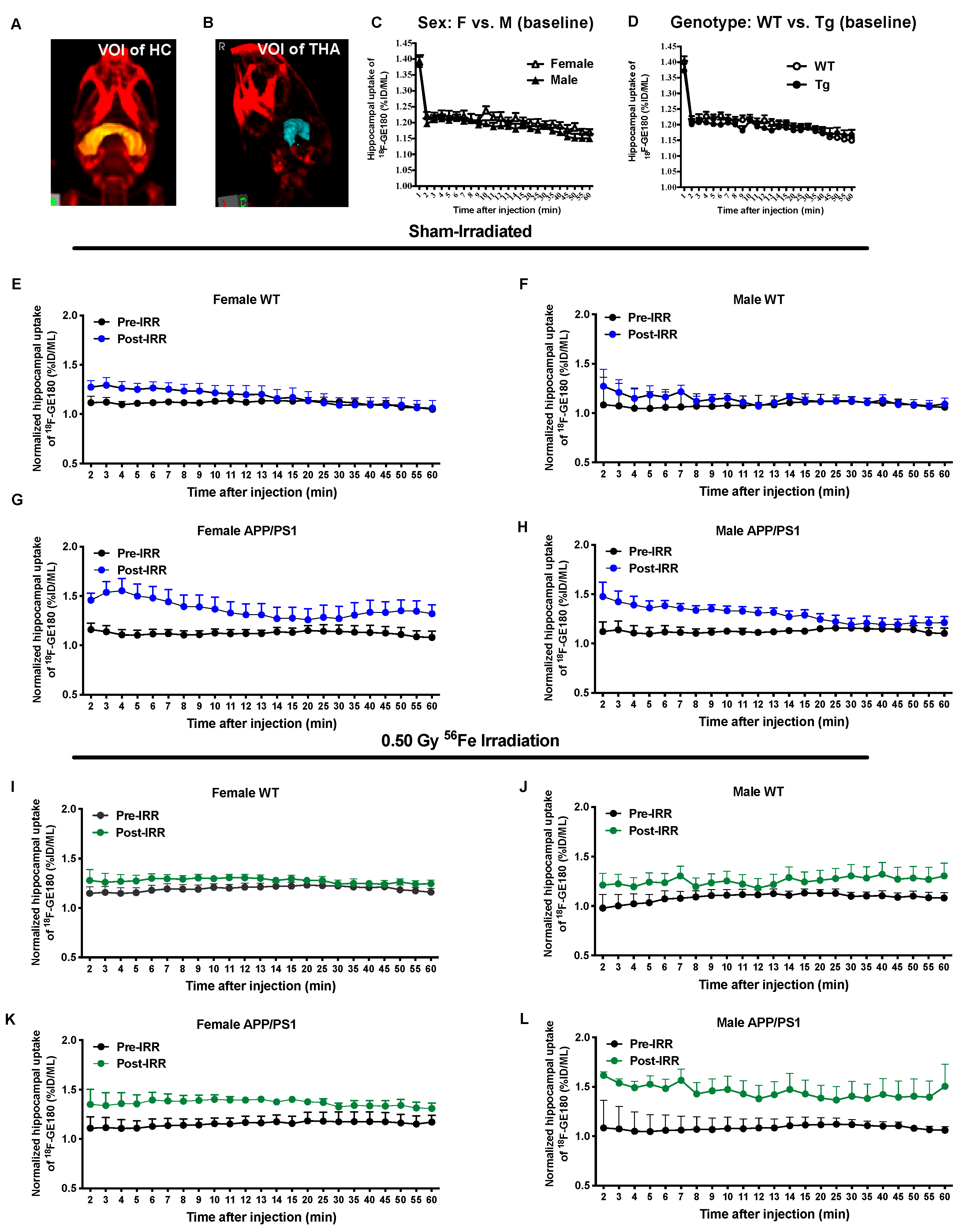
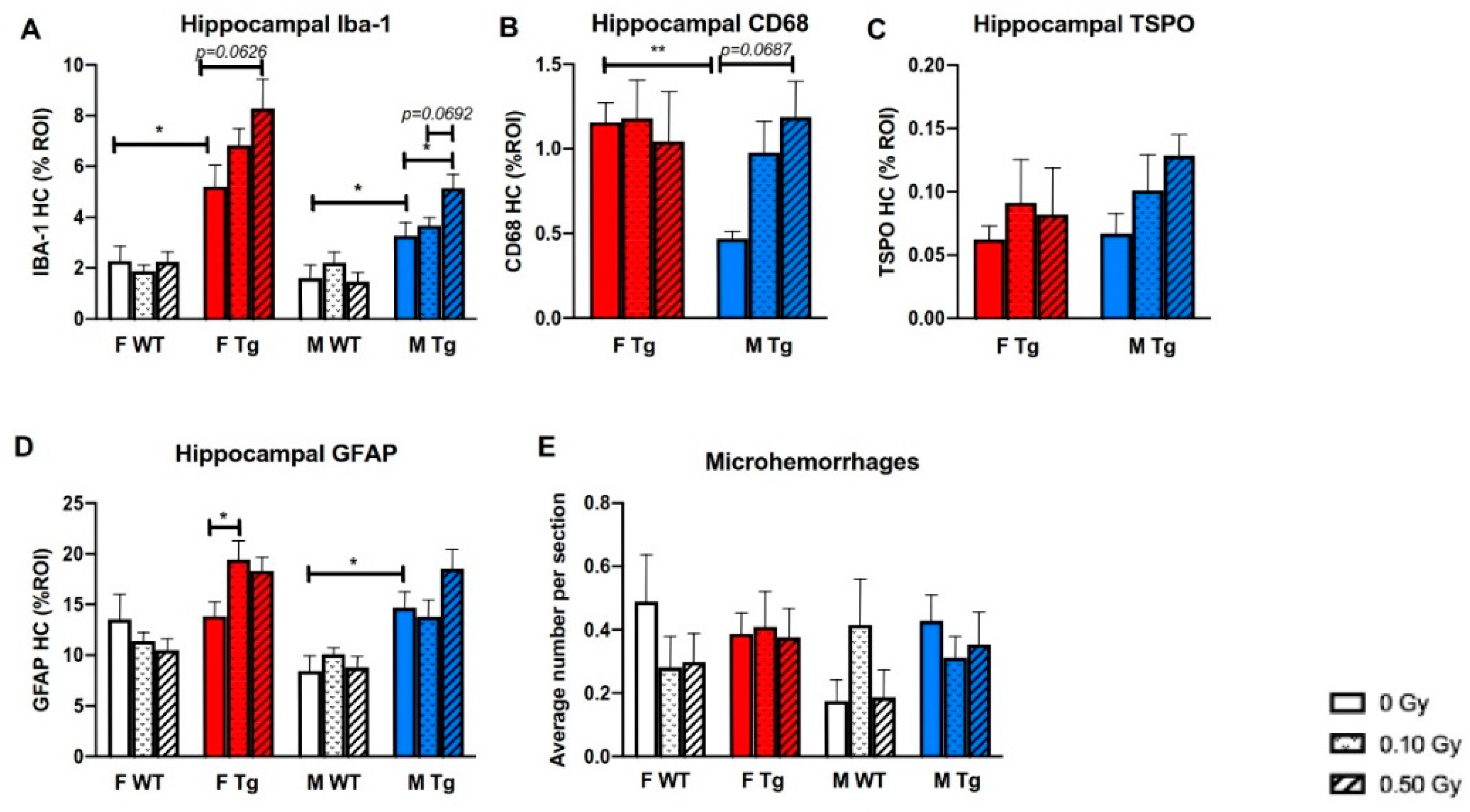
2.3. 56Fe Irradiation Increased Neuroinflammation and Altered Plasma Cytokine Levels
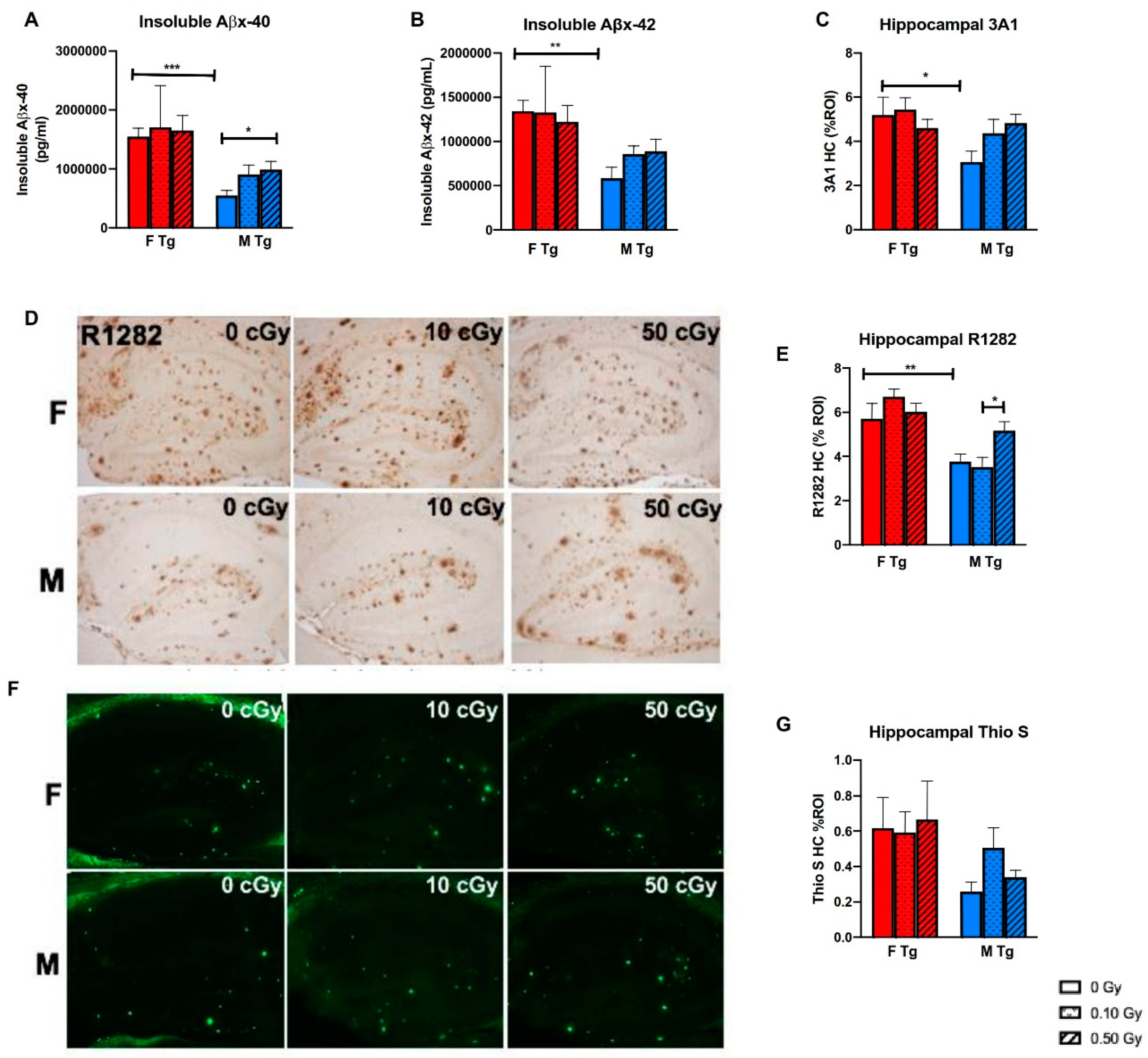
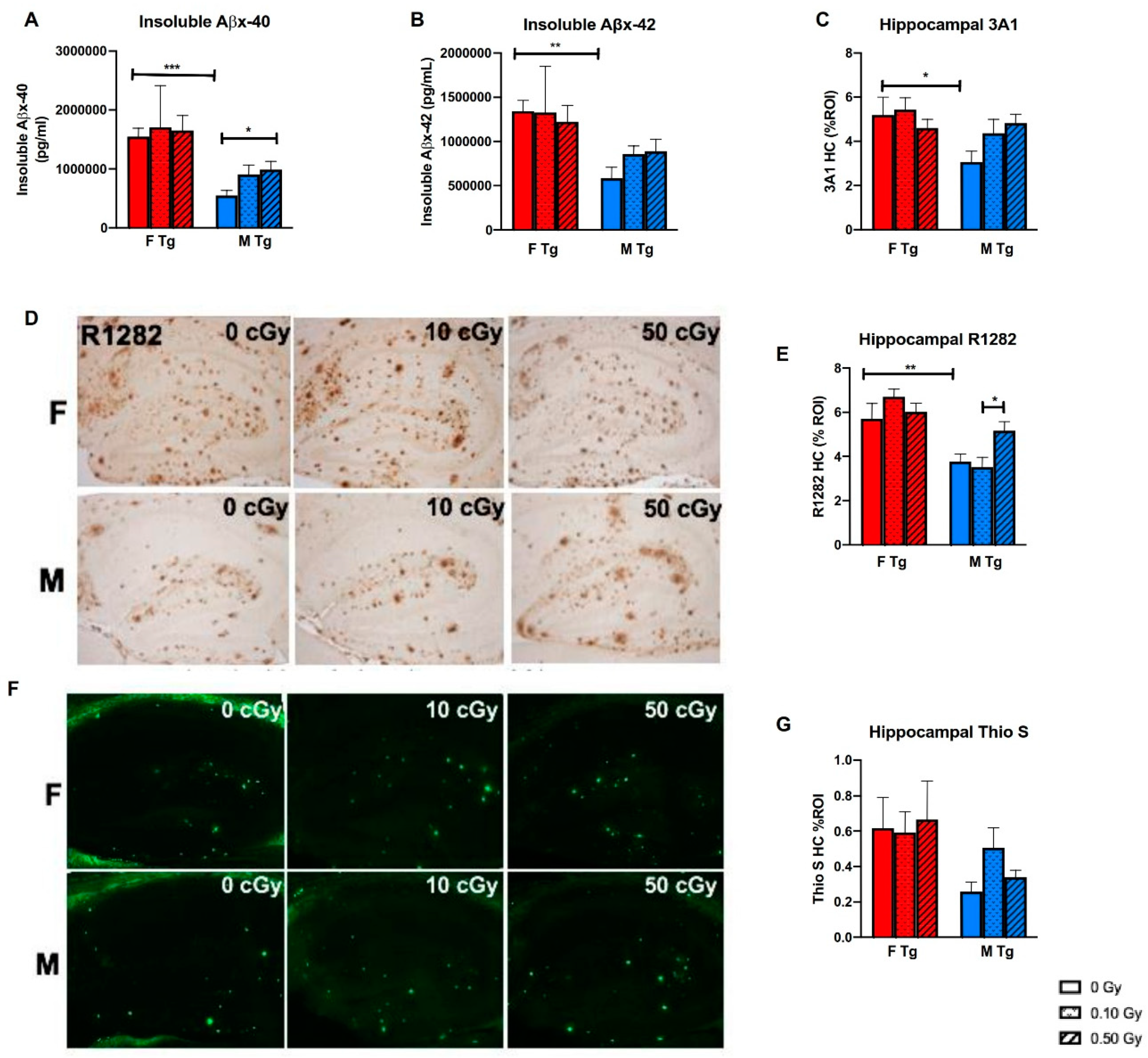
2.4. 56Fe Irradiation Increased Insoluble Aβ Load and Microgliosis in APP/PS1 Male Mice but Did Not Affect Synaptic Markers
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. 56Fe Irradiation of Mice
4.3. Behavioral Tests
4.3.1. Open Field (OF)
4.3.2. Y Maze Spontaneous Alternation Test
4.3.3. Rotarod Test
4.3.4. Elevated Plus Maze (EPM)
4.3.5. Tail Suspension Test (TST)
4.3.6. Contextual Fear Conditioning Test (CFC)
4.3.7. Startle Test
4.3.8. Prepulse Inhibition (PPI)
4.3.9. Grip Strength (GS) Test
4.3.10. Wire Hanging (WH) Test
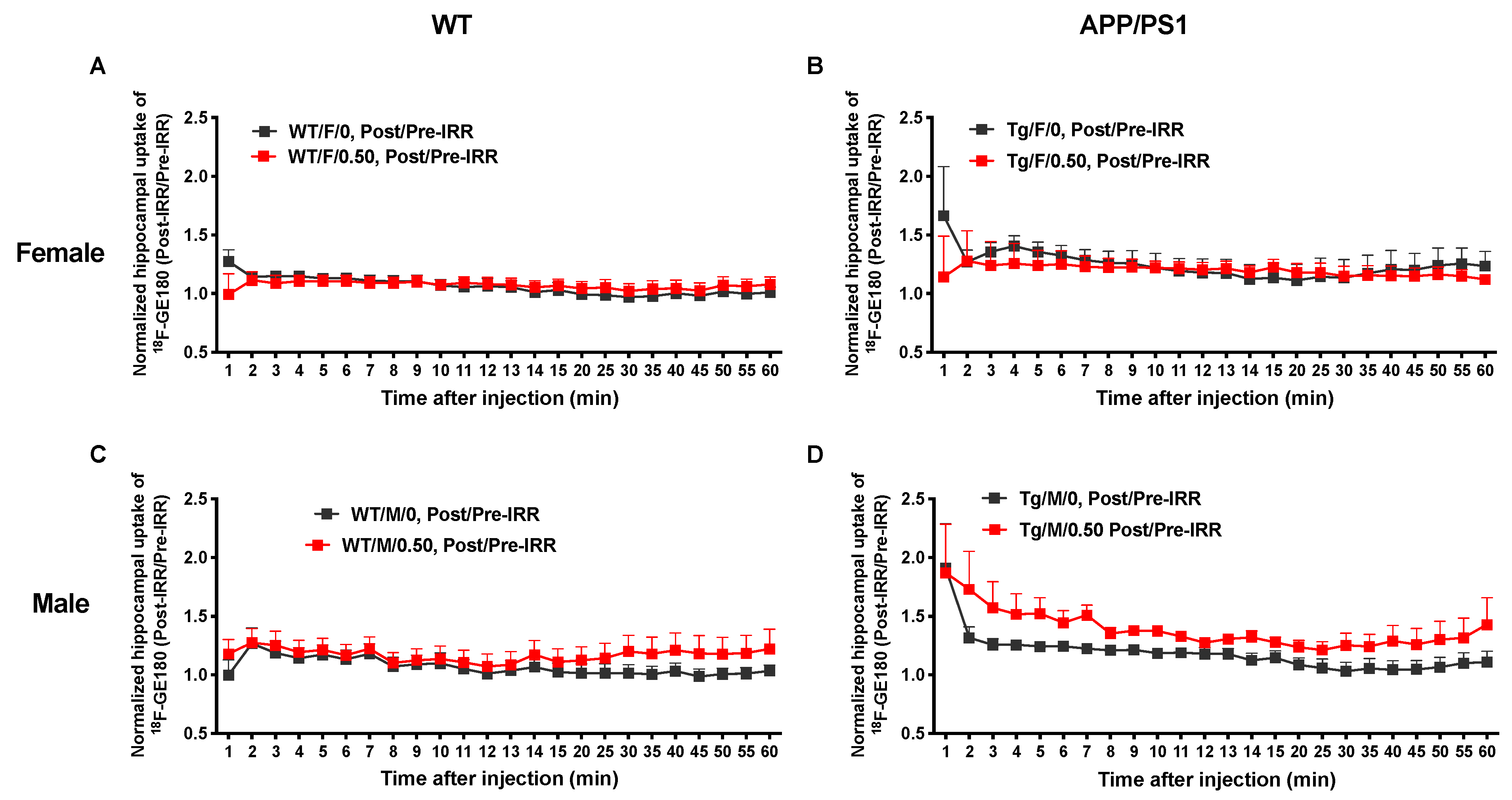
4.4. MicroPET Imaging
4.5. Euthanasia and Tissue Preparation
4.6. MSD Multiplex AB Triplex-38/40/42 Protein ELISA
4.7. MSD Cytokine ELISA
4.8. Immunohistochemistry, Histology, and Quantification
4.9. Western Blot Analysis for Synaptic Markers
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cucinotta, F.; Nikjoo, H.; Goodhead, D. The effects of delta rays on the number of particle-track traversals per cell in laboratory and space exposures. Radiat. Res. 1998, 150, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Hassler, D.M.; Zeitlin, C.; Wimmer-Schweingruber, R.F.; Ehresmann, B.; Rafkin, S.; Eigenbrode, J.L.; Brinza, D.E.; Weigle, G.; Böttcher, S.; Böhm, E.; et al. Mars’ surface radiation environment measured with the Mars Science Laboratory’s Curiosity Rover. Science 2014, 343, 1244797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nzabarushimana, E.; Miousse, I.R.; Shao, L.; Chang, J.; Allen, A.R.; Turner, J.; Stewart, B.; Raber, J.; Koturbash, I. Long-term epigenetic effects of exposure to low doses of 56Fe in the mouse lung. J. Radiat. Res. 2014, 55, 823–828. [Google Scholar] [CrossRef] [Green Version]
- Gridley, D.S.; Obenaus, A.; Bateman, T.A.; Pecaut, M.J. Long-term changes in rat hematopoietic and other physiological systems after high-energy iron ion irradiation. Int. J. Radiat. Biol. 2008, 84, 549–559. [Google Scholar] [CrossRef]
- Durante, M.; Cucinotta, F.A. Heavy ion carcinogenesis and human space exploration. Nat. Rev. Cancer 2008, 8, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Rabin, B.M.; Bielinski, D.F.; Kelly, M.E.; Miller, M.G.; Thanthaeng, N.; Shukitt-Hale, B. Neurochemical differences in learning and memory paradigms among rats supplemented with anthocyanin-rich blueberry diets and exposed to acute doses of 56Fe particles. Life Sci. Space Res. 2017, 12, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Rivera, P.D.; Shih, H.Y.; Leblanc, J.A.; Cole, M.G.; Amaral, W.Z.; Mukherjee, S.; Zhang, S.; Lucero, M.J.; Decarolis, N.A.; Chen, B.P.; et al. Acute and fractionated exposure to high-LET 56Fe HZE-particle radiation both result in similar long-term deficits in adult hippocampal neurogenesis. Radiat. Res 2013, 180, 658–667. [Google Scholar] [CrossRef] [Green Version]
- Sweet, T.B.; Hurley, S.D.; Wu, M.D.; Olschowka, J.A.; Williams, J.P.; O’Banion, M.K. Neurogenic Effects of Low-Dose Whole-Body HZE (Fe) Ion and Gamma Irradiation. Radiat. Res. 2016, 186, 614–623. [Google Scholar] [CrossRef] [Green Version]
- Shukitt-Hale, B.; Lau, F.C.; Cheng, V.; Luskin, K.; Carey, A.N.; Carrihill-Knoll, K.; Rabin, B.M.; Joseph, J.A. Changes in gene expression in the rat hippocampus following exposure to 56Fe particles and protection by berry diets. Cent. Nerv. Syst. Agents Med. Chem. 2013, 13, 36–42. [Google Scholar] [CrossRef] [PubMed]
- DeCarolis, N.A.; Rivera, P.D.; Ahn, F.; Amaral, W.Z.; LeBlanc, J.A.; Malhotra, S.; Shih, H.Y.; Petrik, D.; Melvin, N.; Chen, B.P.; et al. 56Fe Particle Exposure Results in a Long-Lasting Increase in a Cellular Index of Genomic Instability and Transiently Suppresses Adult Hippocampal Neurogenesis In Vivo. Life Sci. Space Res. 2014, 2, 70–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rola, R.; Fishman, K.; Baure, J.; Rosi, S.; Lamborn, K.R.; Obenaus, A.; Nelson, G.A.; Fike, J.R. Hippocampal neurogenesis and neuroinflammation after cranial irradiation with 56Fe particles. Radiat. Res. 2008, 169, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Liu, Y.; Zhao, Q.; Li, J.; Mao, A.; Li, H.; Di, C.; Zhang, H. 56Fe irradiation-induced cognitive deficits through oxidative stress in mice. Toxicol. Res. 2016, 5, 1672–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denisova, N.A.; Shukitt-Hale, B.; Rabin, B.M.; Joseph, J.A. Brain signaling and behavioral responses induced by exposure to 56Fe-particle radiation. Radiat. Res. 2002, 158, 725–734. [Google Scholar] [CrossRef]
- Machida, M.; Lonart, G.; Britten, R.A. Low (60 cGy) doses of 56Fe HZE-particle radiation lead to a persistent reduction in the glutamatergic readily releasable pool in rat hippocampal synaptosomes. Radiat. Res. 2010, 174, 618–623. [Google Scholar] [CrossRef]
- Britten, R.A.; Jewell, J.S.; Miller, V.D.; Davis, L.K.; Hadley, M.M.; Wyrobek, A.J. Impaired Spatial Memory Performance in Adult Wistar Rats Exposed to Low (5–20 cGy) Doses of 1 GeV/n 56Fe Particles. Radiat. Res. 2016, 185, 332–337. [Google Scholar] [CrossRef]
- Britten, R.A.; Davis, L.K.; Jewell, J.S.; Miller, V.D.; Hadley, M.M.; Sanford, L.D.; Machida, M.; Lonart, G. Exposure to mission relevant doses of 1 GeV/Nucleon 56Fe particles leads to impairment of attentional set-shifting performance in socially mature rats. Radiat. Res. 2014, 182, 292–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonart, G.; Parris, B.; Johnson, A.M.; Miles, S.; Sanford, L.D.; Singletary, S.J.; Britten, R.A. Executive function in rats is impaired by low (20 cGy) doses of 1 GeV/u 56Fe particles. Radiat. Res. 2012, 178, 289–294. [Google Scholar] [CrossRef]
- Villasana, L.; Rosenberg, J.; Raber, J. Sex-dependent effects of 56Fe irradiation on contextual fear conditioning in C57BL/6J mice. Hippocampus 2010, 20, 19–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raber, J.; Allen, A.R.; Rosi, S.; Sharma, S.; Dayger, C.; Davis, M.J.; Fike, J.R. Effects of whole body 56Fe radiation on contextual freezing and Arc-positive cells in the dentate gyrus. Behav. Brain Res. 2013, 246, 162–167. [Google Scholar] [CrossRef]
- Cherry, J.D.; Liu, B.; Frost, J.L.; Lemere, C.A.; Williams, J.P.; Olschowka, J.A.; O’Banion, M.K. Galactic cosmic radiation leads to cognitive impairment and increased abeta plaque accumulation in a mouse model of Alzheimer’s disease. PLoS ONE 2012, 7, e53275. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Joseph, J.A.; Shukitt-Hale, B. Effects of age and diet on the heavy particle-induced disruption of operant responding produced by a ground-based model for exposure to cosmic rays. Brain Res. 2005, 1036, 122–129. [Google Scholar] [CrossRef]
- Parihar, V.K.; Maroso, M.; Syage, A.; Allen, B.D.; Angulo, M.C.; Soltesz, I.; Limoli, C.L. Persistent nature of alterations in cognition and neuronal circuit excitability after exposure to simulated cosmic radiation in mice. Exp. Neurol. 2018, 305, 44–55. [Google Scholar] [CrossRef]
- Villasana, L.E.; Benice, T.S.; Raber, J. Long-term effects of 56Fe irradiation on spatial memory of mice: Role of sex and apolipoprotein E isoform. Int. J. Radiat. Oncol. Biol. Phys. 2011, 80, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.J.; Zhang, X.; Chen, W.W. Role of oxidative stress in Alzheimer’s disease. Biomed. Rep. 2016, 4, 519–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenkorang, M.A.; Snyder, B.; Cunningham, R.L. Sex-related differences in oxidative stress and neurodegeneration. Steroids 2018, 133, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Hinshaw, R.G.; Le, K.X.; Park, M.A.; Wang, S.; Belanger, A.P.; Dubey, S.; Frost, J.L.; Shi, Q.; Holton, P.; et al. Space-like 56Fe irradiation manifests mild, early sex-specific behavioral and neuropathological changes in wildtype and Alzheimer’s-like transgenic mice. Sci. Rep. 2019, 9, 12118. [Google Scholar] [CrossRef]
- Villasana, L.; Acevedo, S.; Poage, C.; Raber, J. Sex- and APOE isoform-dependent effects of radiation on cognitive function. Radiat. Res. 2006, 166, 883–891. [Google Scholar] [CrossRef]
- Yeiser, L.A.; Villasana, L.E.; Raber, J. ApoE isoform modulates effects of cranial 56Fe irradiation on spatial learning and memory in the water maze. Behav. Brain Res. 2013, 237, 207–214. [Google Scholar] [CrossRef]
- Villasana, L.E.; Weber, S.; Akinyeke, T.; Raber, J. Genotype differences in anxiety and fear learning and memory of WT and ApoE4 mice associated with enhanced generation of hippocampal reactive oxygen species. J. Neurochem. 2016, 138, 896–908. [Google Scholar] [CrossRef] [Green Version]
- Haley, G.E.; Villasana, L.; Dayger, C.; Davis, M.J.; Raber, J. Apolipoprotein e genotype-dependent paradoxical short-term effects of 56Fe irradiation on the brain. Int. J. Radiat. Oncol. Biol. Phys. 2012, 84, 793–799. [Google Scholar] [CrossRef] [Green Version]
- Gatz, M.; Reynolds, C.A.; Fratiglioni, L.; Johansson, B.; Mortimer, J.A.; Berg, S.; Fiske, A.; Pedersen, N.L. Role of genes and environments for explaining Alzheimer disease. Arch. Gen. Psychiatry 2006, 63, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Vlkolinsky, R.; Titova, E.; Krucker, T.; Chi, B.B.; Staufenbiel, M.; Nelson, G.A.; Obenaus, A. Exposure to 56Fe-particle radiation accelerates electrophysiological alterations in the hippocampus of APP23 transgenic mice. Radiat. Res. 2010, 173, 342–352. [Google Scholar] [CrossRef]
- Townsend, L.W.; Fry, R.J. Radiation protection guidance for activities in low-Earth orbit. Adv. Space Res. 2002, 30, 957–963. [Google Scholar] [CrossRef]
- Whoolery, C.W.; Walker, A.K.; Richardson, D.R.; Lucero, M.J.; Reynolds, R.P.; Beddow, D.H.; Clark, K.L.; Shih, H.Y.; LeBlanc, J.A.; Cole, M.G.; et al. Whole-Body Exposure to 28Si-Radiation Dose-Dependently Disrupts Dentate Gyrus Neurogenesis and Proliferation in the Short Term and New Neuron Survival and Contextual Fear Conditioning in the Long Term. Radiat. Res. 2017, 188, 532–551. [Google Scholar] [CrossRef]
- Silasi, G.; Diaz-Heijtz, R.; Besplug, J.; Rodriguez-Juarez, R.; Titov, V.; Kolb, B.; Kovalchuk, O. Selective brain responses to acute and chronic low-dose X-ray irradiation in males and females. Biochem. Biophys. Res. Commun. 2004, 325, 1223–1235. [Google Scholar] [CrossRef]
- Krukowski, K.; Grue, K.; Frias, E.S.; Pietrykowski, J.; Jones, T.; Nelson, G.; Rosi, S. Female mice are protected from space radiation-induced maladaptive responses. Brain Behav. Immun. 2018, 74, 106–120. [Google Scholar] [CrossRef]
- Patel, R.; Arakawa, H.; Radivoyevitch, T.; Gerson, S.L.; Welford, S.M. Long-Term Deficits in Behavior Performances Caused by Low- and High-Linear Energy Transfer Radiation. Radiat. Res. 2017, 188, 672–680. [Google Scholar] [CrossRef]
- Casadesus, G.; Shukitt-Hale, B.; Cantuti-Castelvetri, I.; Rabin, B.M.; Joseph, J.A. The effects of heavy particle irradiation on exploration and response to environmental change. Adv. Space Res. 2004, 33, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Pecaut, M.J.; Haerich, P.; Miller, C.N.; Smith, A.L.; Zendejas, E.D.; Nelson, G.A. The effects of low-dose, high-LET radiation exposure on three models of behavior in C57BL/6 mice. Radiat. Res. 2004, 162, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Miry, O.; Zhang, X.L.; Vose, L.R.; Gopaul, K.R.; Subah, G.; Moncaster, J.A.; Wojnarowicz, M.W.; Fisher, A.M.; Tagge, C.A.; Goldstein, L.E.; et al. Life-long brain compensatory responses to galactic cosmic radiation exposure. Sci. Rep. 2021, 11, 4292. [Google Scholar] [CrossRef]
- Soler, I.; Yun, S.; Reynolds, R.P.; Whoolery, C.W.; Tran, F.H.; Kumar, P.L.; Rong, Y.; DeSalle, M.J.; Gibson, A.D.; Stowe, A.M.; et al. Multi-Domain Touchscreen-Based Cognitive Assessment of C57BL/6J Female Mice Shows Whole-Body Exposure to 56Fe Particle Space Radiation in Maturity Improves Discrimination Learning Yet Impairs Stimulus-Response Rule-Based Habit Learning. Front. Behav. Neurosci. 2021, 15, 722780. [Google Scholar] [CrossRef]
- Parihar, V.K.; Angulo, M.C.; Allen, B.D.; Syage, A.; Usmani, M.T.; Passerat de la Chapelle, E.; Amin, A.N.; Flores, L.; Lin, X.; Giedzinski, E.; et al. Sex-Specific Cognitive Deficits Following Space Radiation Exposure. Front. Behav. Neurosci. 2020, 14, 535885. [Google Scholar] [CrossRef]
- Klein, P.M.; Parihar, V.K.; Szabo, G.G.; Zoldi, M.; Angulo, M.C.; Allen, B.D.; Amin, A.N.; Nguyen, Q.A.; Katona, I.; Baulch, J.E.; et al. Detrimental impacts of mixed-ion radiation on nervous system function. Neurobiol. Dis. 2021, 151, 105252. [Google Scholar] [CrossRef] [PubMed]
- Keiser, A.A.; Kramar, E.A.; Dong, T.; Shanur, S.; Pirodan, M.; Ru, N.; Acharya, M.M.; Baulch, J.E.; Limoli, C.L.; Wood, M.A. Systemic HDAC3 inhibition ameliorates impairments in synaptic plasticity caused by simulated galactic cosmic radiation exposure in male mice. Neurobiol. Learn. Mem. 2021, 178, 107367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Forkstam, C.; Engel, J.A.; Svensson, L. Role of dopamine in prepulse inhibition of acoustic startle. Psychopharmacology 2000, 149, 181–188. [Google Scholar] [CrossRef]
- Rabin, B.M.; Joseph, J.A.; Shukitt-Hale, B. Long-term changes in amphetamine-induced reinforcement and aversion in rats following exposure to 56Fe particle. Adv. Space Res. 2003, 31, 127–133. [Google Scholar] [CrossRef]
- Rabin, B.M.; Joseph, J.A.; Shukitt-Hale, B.; McEwen, J. Effects of exposure to heavy particles on a behavior mediated by the dopaminergic system. Adv. Space Res. 2000, 25, 2065–2074. [Google Scholar] [CrossRef]
- Rabin, B.M.; Joseph, J.A.; Erat, S. Effects of exposure to different types of radiation on behaviors mediated by peripheral or central systems. Adv. Space Res. 1998, 22, 217–225. [Google Scholar] [CrossRef]
- Haerich, P.; Eggers, C.; Pecaut, M.J. Investigation of the effects of head irradiation with gamma rays and protons on startle and pre-pulse inhibition behavior in mice. Radiat. Res. 2012, 177, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Impey, S.; Jopson, T.; Pelz, C.; Tafessu, A.; Fareh, F.; Zuloaga, D.; Marzulla, T.; Riparip, L.K.; Stewart, B.; Rosi, S.; et al. Short- and long-term effects of 56Fe irradiation on cognition and hippocampal DNA methylation and gene expression. BMC Genom. 2016, 17, 825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marty, V.N.; Vlkolinsky, R.; Minassian, N.; Cohen, T.; Nelson, G.A.; Spigelman, I. Radiation-induced alterations in synaptic neurotransmission of dentate granule cells depend on the dose and species of charged particles. Radiat. Res. 2014, 182, 653–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philpott, D.E.; Sapp, W.; Miquel, J.; Kato, K.; Corbett, R.; Stevenson, J.; Black, S.; Lindseth, K.A.; Benton, E.V. The effect of high energy (HZE) particle radiation (40Ar) on aging parameters of mouse hippocampus and retina. Scanning Electron. Microsc. 1985, 1177–1182. [Google Scholar]
- Britten, R.A.; Miller, V.D.; Hadley, M.M.; Jewell, J.S.; Macadat, E. Performance in hippocampus- and PFC-dependent cognitive domains are not concomitantly impaired in rats exposed to 20cGy of 1GeV/n 56Fe particles. Life Sci. Space Res. 2016, 10, 17–22. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, H.; Sun, G.; Zhang, J.; Edwards, N.J.; Aronowski, J. Neuronal Interleukin-4 as a Modulator of Microglial Pathways and Ischemic Brain Damage. J. Neurosci. 2015, 35, 11281–11291. [Google Scholar] [CrossRef] [Green Version]
- Fiorentino, D.F.; Zlotnik, A.; Mosmann, T.R.; Howard, M.; O’Garra, A. IL-10 inhibits cytokine production by activated macrophages. J. Immunol. 1991, 147, 3815–3822. [Google Scholar]
- Cianciulli, A.; Dragone, T.; Calvello, R.; Porro, C.; Trotta, T.; Lofrumento, D.D.; Panaro, M.A. IL-10 plays a pivotal role in anti-inflammatory effects of resveratrol in activated microglia cells. Int. Immunopharmacol. 2015, 24, 369–376. [Google Scholar] [CrossRef]
- Chiang, C.S.; Hong, J.H.; Stalder, A.; Sun, J.R.; Withers, H.R.; McBride, W.H. Delayed molecular responses to brain irradiation. Int. J. Radiat. Biol. 1997, 72, 45–53. [Google Scholar] [CrossRef]
- Moravan, M.J.; Olschowka, J.A.; Williams, J.P.; O’Banion, M.K. Cranial irradiation leads to acute and persistent neuroinflammation with delayed increases in T-cell infiltration and CD11c expression in C57BL/6 mouse brain. Radiat. Res. 2011, 176, 459–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowsky, J.L.; Fadale, D.J.; Anderson, J.; Xu, G.M.; Gonzales, V.; Jenkins, N.A.; Copeland, N.G.; Lee, M.K.; Younkin, L.H.; Wagner, S.L.; et al. Mutant presenilins specifically elevate the levels of the 42 residue beta-amyloid peptide in vivo: Evidence for augmentation of a 42-specific gamma secretase. Hum. Mol. Genet. 2004, 13, 159–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimbel, D.A.; Nygaard, H.B.; Coffey, E.E.; Gunther, E.C.; Lauren, J.; Gimbel, Z.A.; Strittmatter, S.M. Memory impairment in transgenic Alzheimer mice requires cellular prion protein. J. Neurosci. 2010, 30, 6367–6374. [Google Scholar] [CrossRef]
- Jankowsky, J.L.; Melnikova, T.; Fadale, D.J.; Xu, G.M.; Slunt, H.H.; Gonzales, V.; Younkin, L.H.; Younkin, S.G.; Borchelt, D.R.; Savonenko, A.V. Environmental enrichment mitigates cognitive deficits in a mouse model of Alzheimer’s disease. J. Neurosci. 2005, 25, 5217–5224. [Google Scholar] [CrossRef]
- Savonenko, A.; Xu, G.M.; Melnikova, T.; Morton, J.L.; Gonzales, V.; Wong, M.P.; Price, D.L.; Tang, F.; Markowska, A.L.; Borchelt, D.R. Episodic-like memory deficits in the APPswe/PS1dE9 mouse model of Alzheimer’s disease: Relationships to beta-amyloid deposition and neurotransmitter abnormalities. Neurobiol. Dis. 2005, 18, 602–617. [Google Scholar] [CrossRef]
- Wickstrom, T.; Clarke, A.; Gausemel, I.; Horn, E.; Jorgensen, K.; Khan, I.; Mantzilas, D.; Rajanayagam, T.; In’t Veld, D.J.; Trigg, W. The development of an automated and GMP compliant FASTlab Synthesis of [18F]GE-180; a radiotracer for imaging translocator protein (TSPO). J. Label. Compd. Radiopharm. 2014, 57, 42–48. [Google Scholar] [CrossRef]
- Shi, Q.; Chowdhury, S.; Ma, R.; Le, K.X.; Hong, S.; Caldarone, B.J.; Stevens, B.; Lemere, C.A. Complement C3 deficiency protects against neurodegeneration in aged plaque-rich APP/PS1 mice. Sci. Transl. Med. 2017, 9, 392. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Measures | Tests |
|---|---|
| General Health | SHIRPA |
| Grip and Muscle Strength Endurance | Grip Strength (GS) Wire Hanging (WH) |
| Depression | Tail Suspension Test (TST) |
| Anxiety | Open Field (OF) Elevated Plus Maze (EPM) |
| Locomotor Activity | Open Field (OF) Y Maze (YM) Elevated Plus Maze (EPM) |
| Motor Coordination | Rotarod |
| Motor Learning | Rotarod |
| Short-Term Memory | Y Maze (YM) |
| Fear Memory | Contextual Fear Conditioning (CFC) |
| Sensory Reactivity | Startle |
| Sensory Gating | Pre-Pulse Inhibition (PPI) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schroeder, M.K.; Liu, B.; Hinshaw, R.G.; Park, M.-A.; Wang, S.; Dubey, S.; Liu, G.G.; Shi, Q.; Holton, P.; Reiser, V.; et al. Long-Term Sex- and Genotype-Specific Effects of 56Fe Irradiation on Wild-Type and APPswe/PS1dE9 Transgenic Mice. Int. J. Mol. Sci. 2021, 22, 13305. https://doi.org/10.3390/ijms222413305
Schroeder MK, Liu B, Hinshaw RG, Park M-A, Wang S, Dubey S, Liu GG, Shi Q, Holton P, Reiser V, et al. Long-Term Sex- and Genotype-Specific Effects of 56Fe Irradiation on Wild-Type and APPswe/PS1dE9 Transgenic Mice. International Journal of Molecular Sciences. 2021; 22(24):13305. https://doi.org/10.3390/ijms222413305
Chicago/Turabian StyleSchroeder, Maren K., Bin Liu, Robert G. Hinshaw, Mi-Ae Park, Shuyan Wang, Shipra Dubey, Grace Geyu Liu, Qiaoqiao Shi, Peter Holton, Vladimir Reiser, and et al. 2021. "Long-Term Sex- and Genotype-Specific Effects of 56Fe Irradiation on Wild-Type and APPswe/PS1dE9 Transgenic Mice" International Journal of Molecular Sciences 22, no. 24: 13305. https://doi.org/10.3390/ijms222413305
APA StyleSchroeder, M. K., Liu, B., Hinshaw, R. G., Park, M.-A., Wang, S., Dubey, S., Liu, G. G., Shi, Q., Holton, P., Reiser, V., Jones, P. A., Trigg, W., Di Carli, M. F., Caldarone, B. J., Williams, J. P., O’Banion, M. K., & Lemere, C. A. (2021). Long-Term Sex- and Genotype-Specific Effects of 56Fe Irradiation on Wild-Type and APPswe/PS1dE9 Transgenic Mice. International Journal of Molecular Sciences, 22(24), 13305. https://doi.org/10.3390/ijms222413305