
Cryptochrome 2 from Lilium × formolongi Regulates Photoperiodic Flowering in Transgenic Arabidopsis thaliana

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cloning and Sequence Analysis of LfCRY2
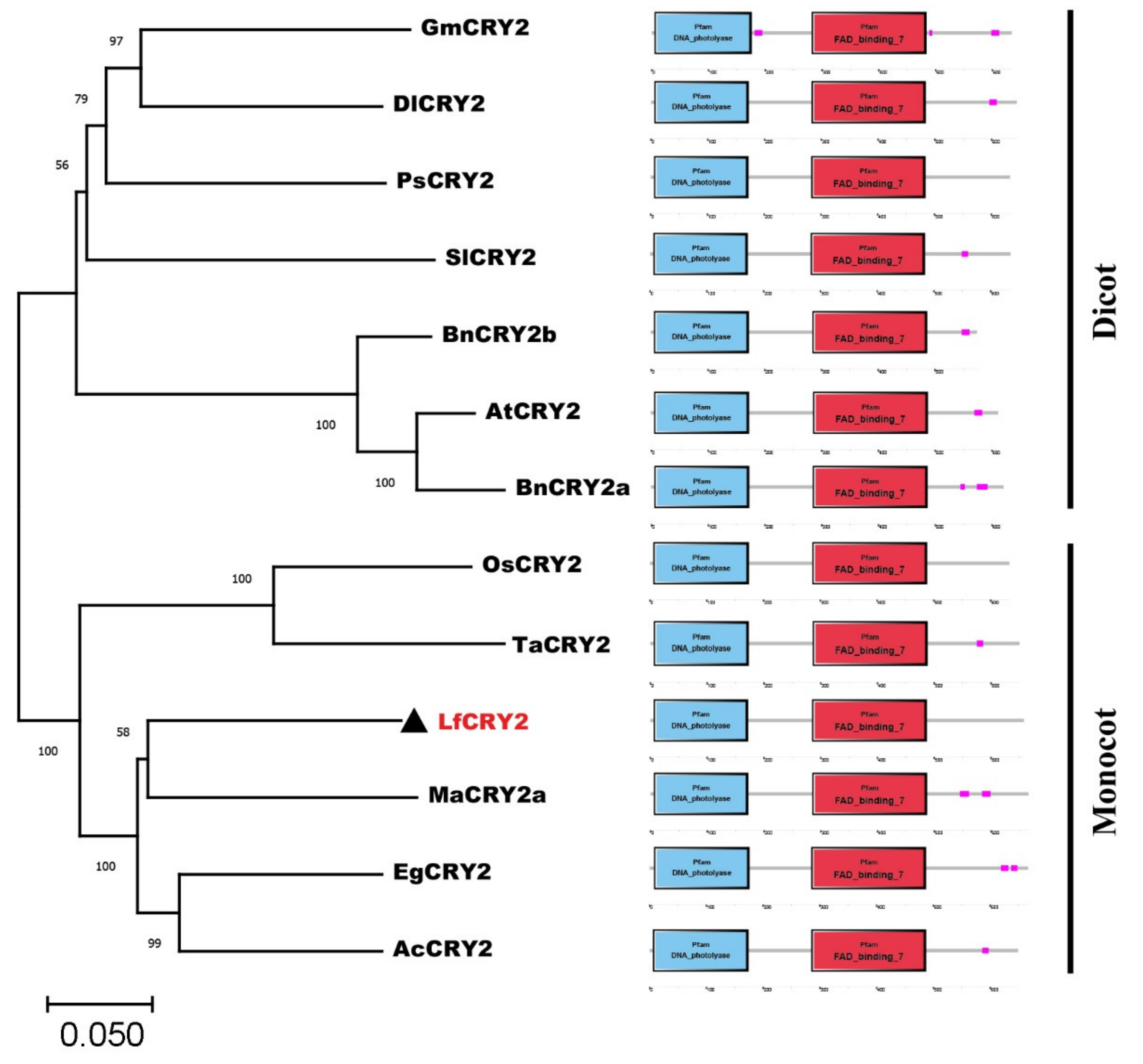
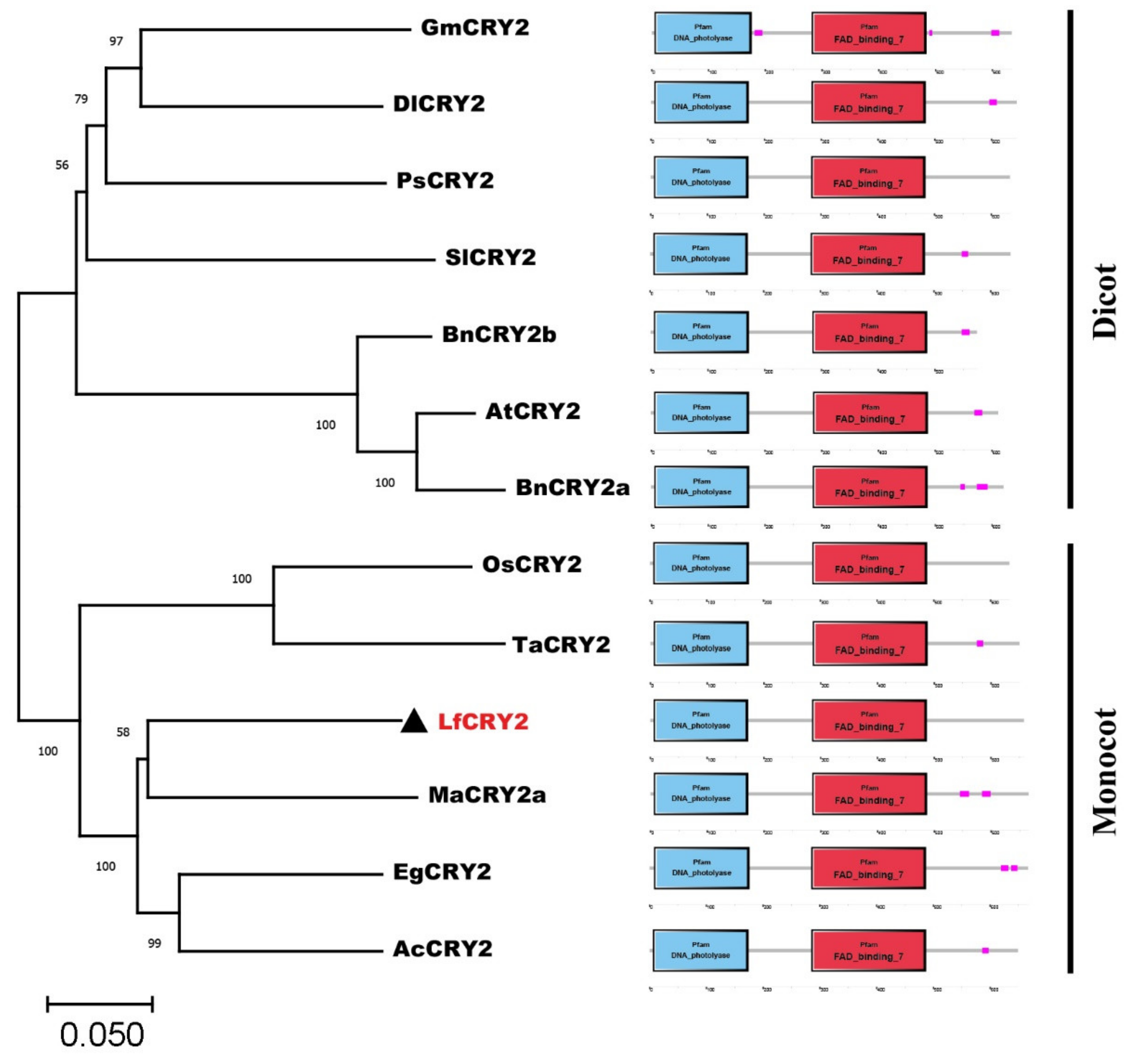
2.2. Phylogenetic and Amino Acid Similarity Analysis of LfCRY2
2.3. Tissue-Specific Expression and Photoperiodic Regulation of CRY2 Gene in L. × formolongi
2.4. Circadian and Light Regulation of CRY2 Gene in L. × formolongi
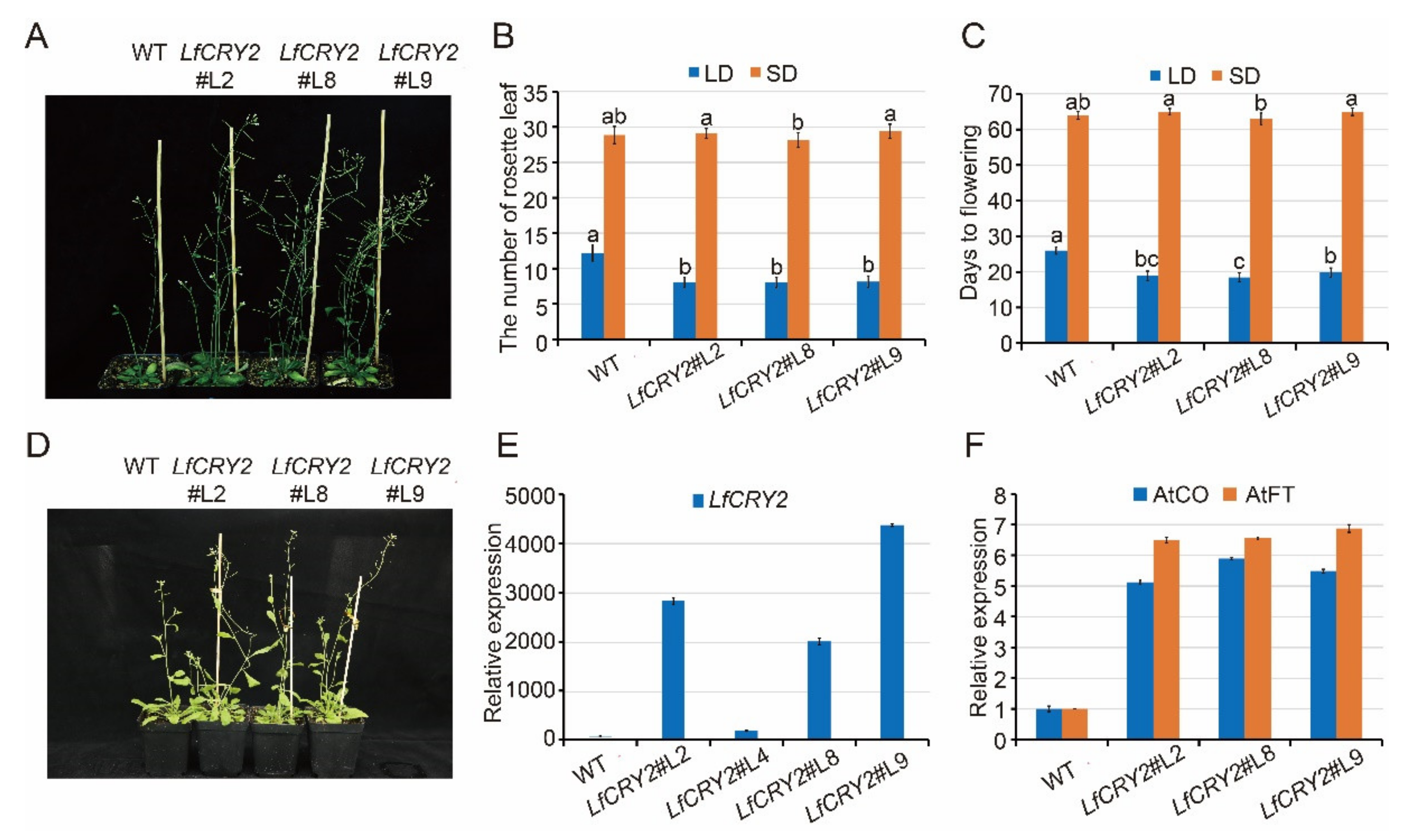
2.5. Overexpression of LfCRY2 Promoted the Flowering of Arabidopsis under LDs
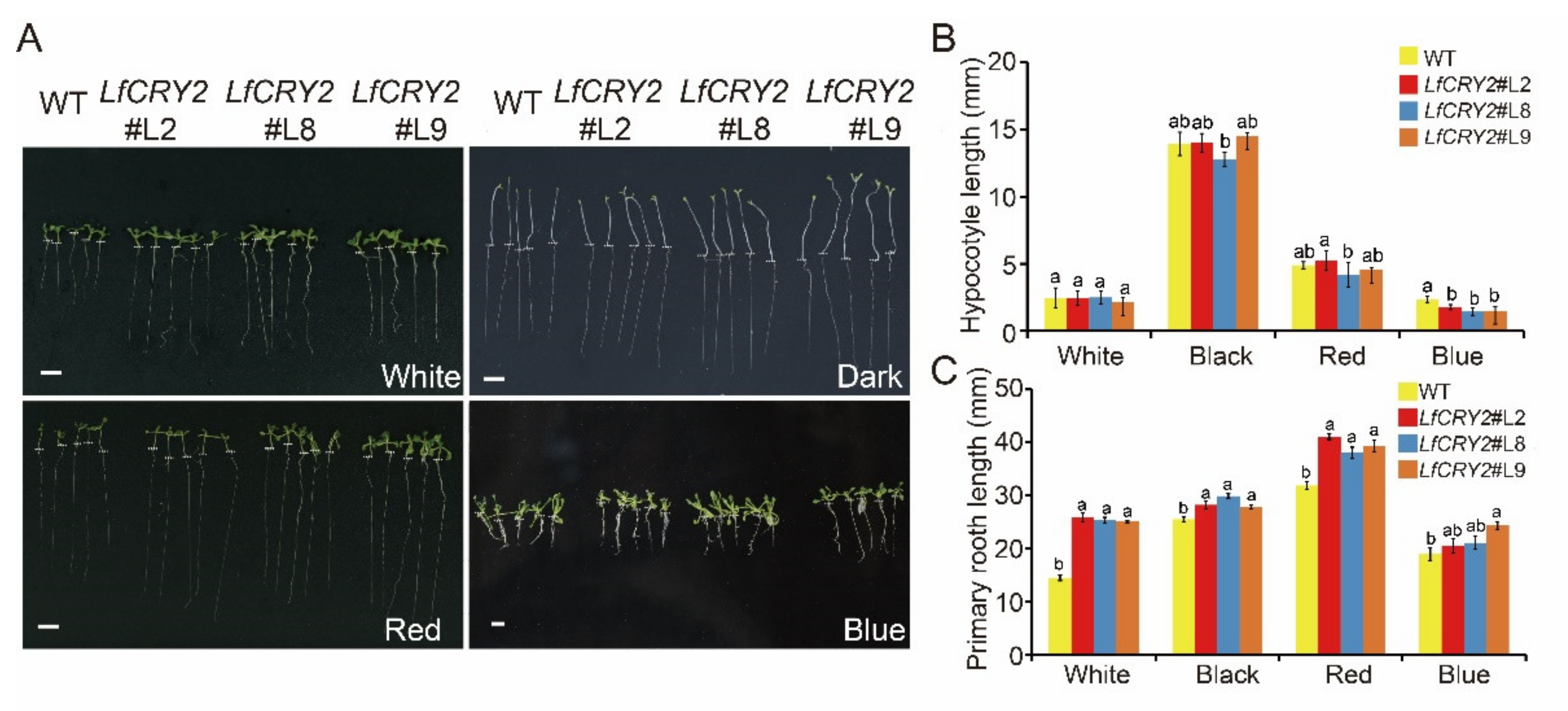
2.6. Overexpression of LfCRY2 Results in Longer Roots and Blue Light-Specific Hypocotyl in the Transgenic Arabidopsis
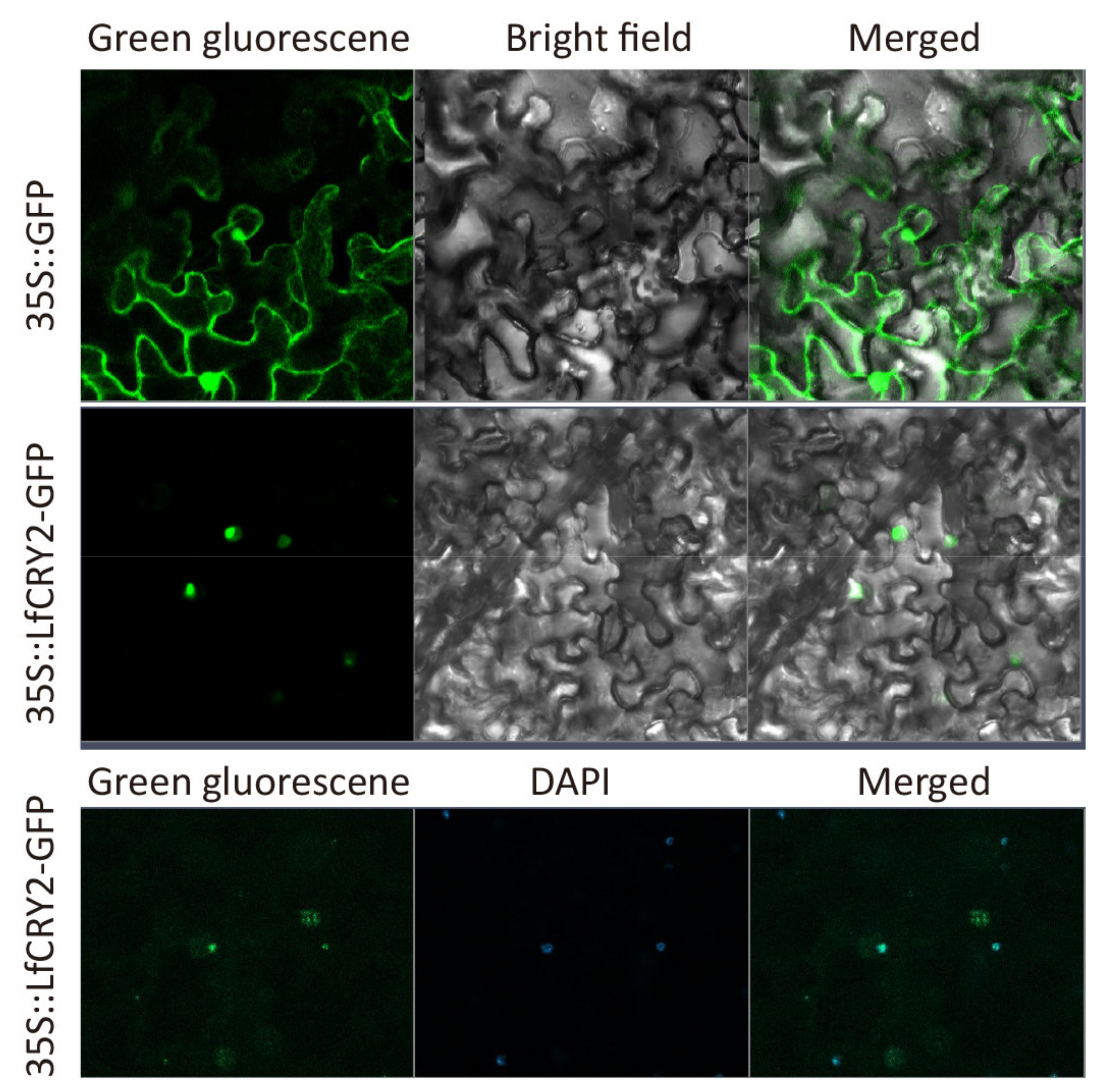
2.7. Subcelluar Localization
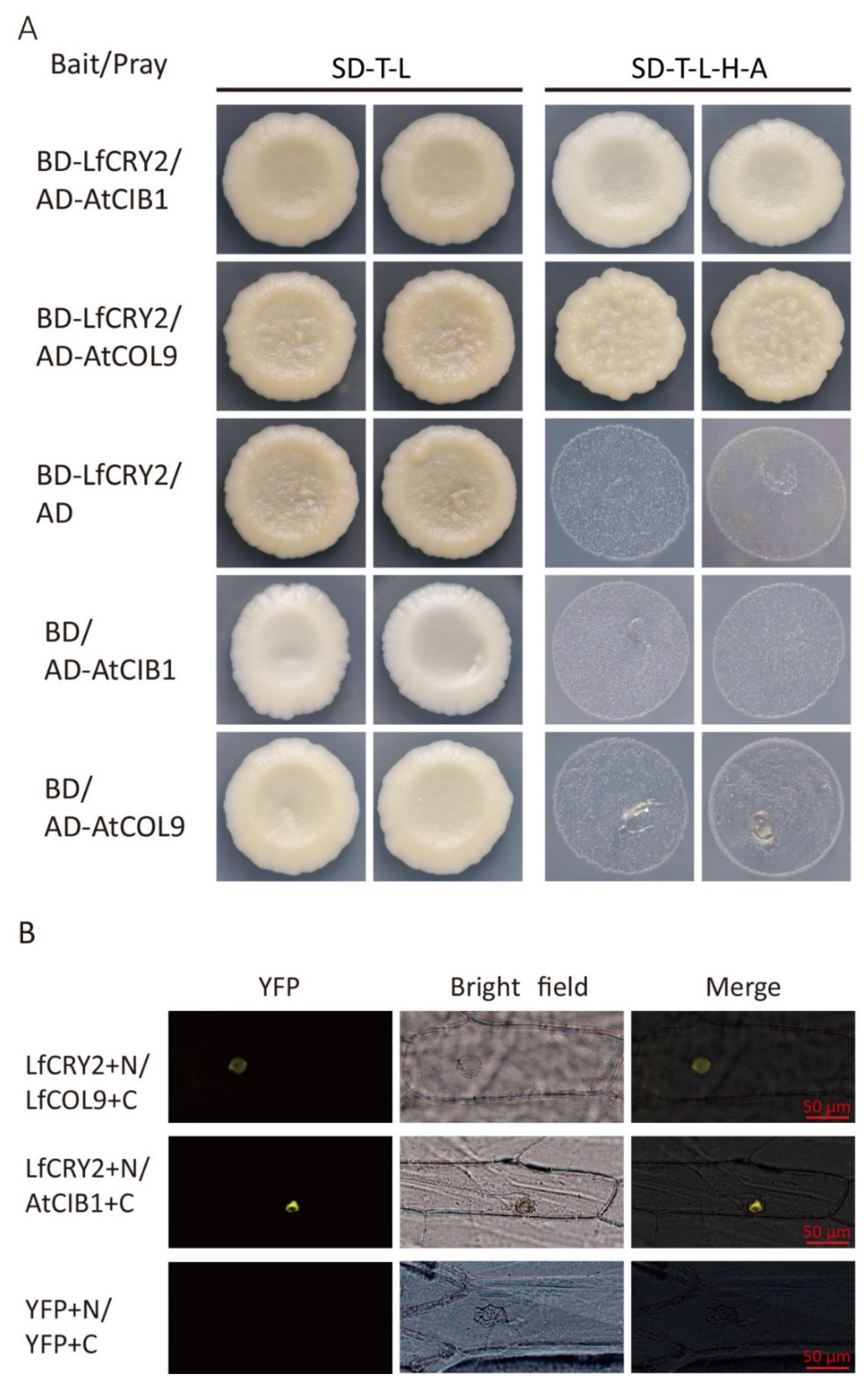
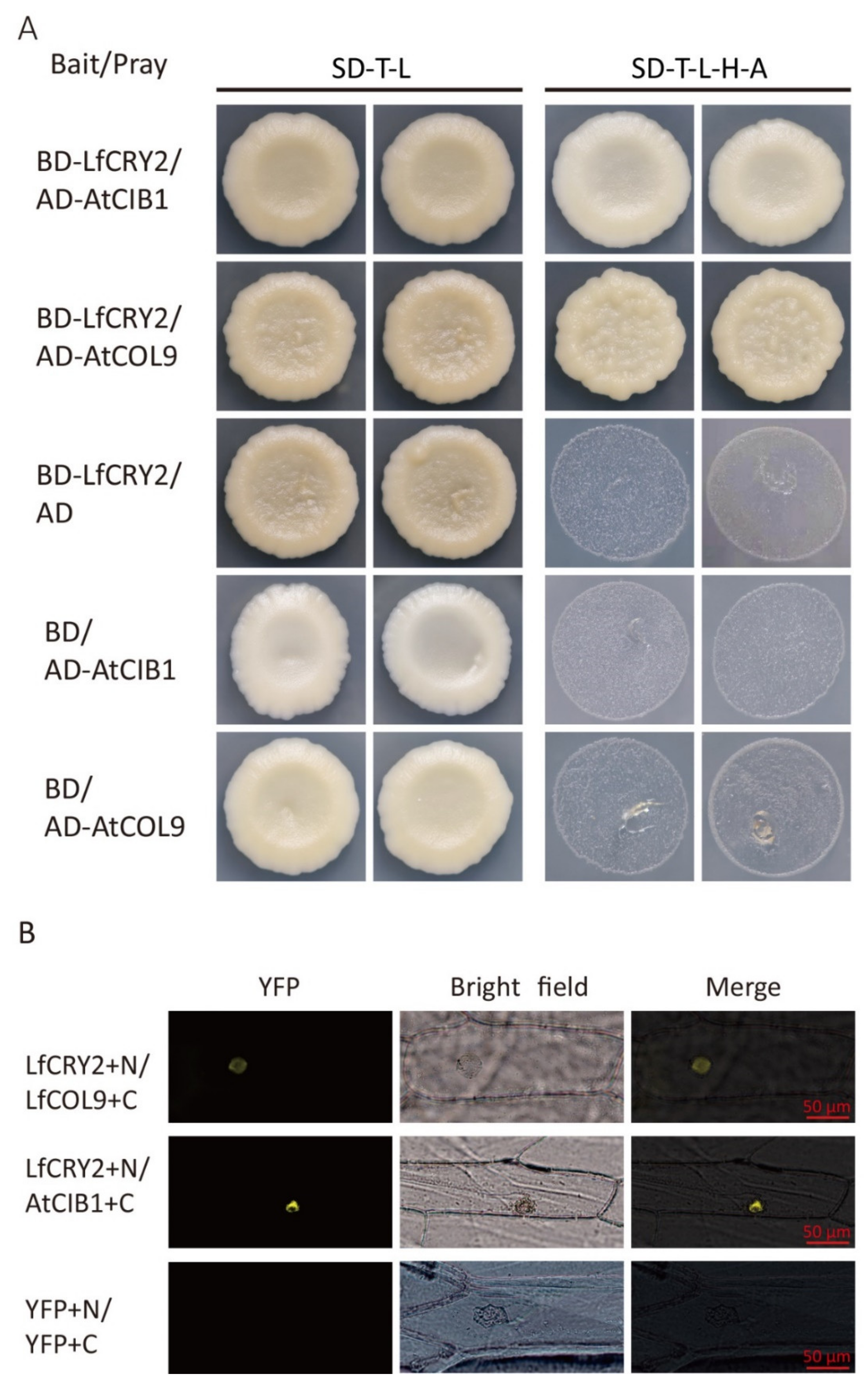
2.8. LfCRY2 Interacts with LfCOL9 and AtCOB1 in Yeast and Plant Cells
3. Discussion
4. Materials and Methods
4.1. Phylogenetic and Bioinformatic Analyses
4.2. Plant Materials and Growth Conditions
4.3. Expression of LfCRY2 Gene
4.4. Arabidopsis Transformation and Functional Analysis
4.4.1. Arabidopsis Transformation
4.4.2. Flowering Studies
4.5. Subcellular Localization
4.6. Yeast Two-Hybrid Assay
4.7. Bimolecular Fluorescence Complementation (BiFC) Assay
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lumsden, P.J. Photoperiodism in plants. In Biological Rhythms; Kumar, V., Ed.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Mouradov, A.; Cremer, F.; Coupland, G. Control of flowering time: Interacting pathways as a basis for diversity. Plant Cell 2002, 14, S111–S130. [Google Scholar] [CrossRef] [Green Version]
- Quail, P.H. Phytochrome photosensory signalling networks. Nat. Rev. Mol. Cell Biol. 2002, 3, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Batschauer, A. Plant blue-light receptors. Planta 2005, 220, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Song, Y.H.; Imaizumi, T. LOV domain-containing F-Box proteins: Light-dependent protein degradation modules in Arabidopsis. Mol. Plant 2012, 5, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Rizzini, L.; Favory, J.J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schafer, E.; Nagy, F.; Jenkins, G.I. Perception of UV-B by the Arabidopsis UVR8 protein. Science 2011, 332, 103–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cashmore, A.R. Cryptochromes enabling plants and animals to determine circadian time. Cell 2003, 114, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A. Structure and function of DNA photolyase and cryptochrome blue-light photoreceptors. Chem. Rev. 2003, 103, 2203–2238. [Google Scholar] [CrossRef] [PubMed]
- Partch, C.L.; Sancar, A. Photochemistry and photobiology of cryptochrome blue-light photopigments: The search for a photocycle. Photochem. Photobiol. 2010, 81, 1291–1304. [Google Scholar] [CrossRef]
- Briggs, W.R.; Huala, E. Blue-light photoreceptors in higher plants. Annu. Rev. Cell Dev. Biol. 1999, 15, 33–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.; Zhang, Y.C.; Sang, Y.; Li, Q.H.; Yang, H.Q. A role for Arabidopsis cryptochromes and COP1 in the regulation of stomatal opening. Proc. Natl. Acad. Sci. USA 2005, 102, 12270–12275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.; Cashmore, A.R. HY4 gene of A. thaliana encodes a protein with characteristics of a blue-light photoreceptor. Nature 1993, 366, 162–166. [Google Scholar] [CrossRef]
- Guo, H.; Yang, H.; Mockler, T.C.; Lin, C. Regulation of flowering time by Arabidopsis photoreceptors. Science 1998, 279, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.H.; Liu, H.T.; Klejnot, J.; Lin, C.T. The cryptochrome blue light receptors. Arabidopsis Book 2010, 8, e0135. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Ma, L.G.; Li, J.M.; Zhao, H.Y.; Deng, X.W. Direct interaction of Arabidopsis cryptochromes with COP1 in light control cevelopment. Science 2001, 294, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.Q.; Wu, Y.J.; Tang, R.H.; Liu, D.; Liu, Y.; Cashmore, A.R. The C termini of Arabidopsis cryptochromes mediate a constitutive light response. Cell 2000, 103, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.Q.; Tang, R.H.; Cashmore, A.R. The signaling mechanism of Arabidopsis CRY1 involves direct interaction with COP1. Plant Cell 2001, 13, 2573–2587. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Jarillo, J.A.; Cashmore, A.R. Chimeric proteins between CRY1 and CRY2 Arabidopsis blue light photoreceptors indicate overlapping functions and varying protein stability. Plant Cell 1998, 10, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Shalitin, D.; Yang, H.Y.; Mockler, T.C.; Maymon, M.; Guo, H.W.; Whitelam, G.C.; Lin, C.T. Regulation of Arabidopsis cryptochrome 2 by blue-light-dependent phosphorylation. Nature 2002, 417, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.J.; Zhang, Y.C.; Li, Q.H.; Sang, Y.; Mao, J.; Lian, H.L.; Wang, L.; Yang, H.Q. COP1-mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering in Arabidopsis. Plant Cell 2008, 20, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Kaya, H.; Goto, K.; Iwabuchi, M.; Araki, T. A pair of related genes with antagonistic roles in mediating flowering signals. Science 1999, 286, 1960–1962. [Google Scholar] [CrossRef] [PubMed]
- Samach, A.; Onouchi, H.; Gold, S.E.; Ditta, G.S.; Schwarz-Sommer, Z.; Yanofsky, M.F.; Coupland, G. Distinct roles of CONSTANS target genes in reproductive development of Arabidopsis. Science 2000, 288, 1613–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onouchi, H.; Igeño, M.I.; Périlleux, C.; Graves, K.; Coupland, G. Mutagenesis of plants overexpressing CONSTANS demonstrates novel interactions among Arabidopsis flowering-time genes. Plant Cell 2000, 12, 885–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbesier, L.; Vincent, C.; Jang, S.; Fornara, F.; Fan, Q.; Searle, L.; Giakountis, A.; Farrona, S.; Gissot, L.; Turnbull, C. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 2007, 316, 1030–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.; Marchal, V.; Panigrahi, K.; Wenkel, S.; Soppe, W.; Deng, X.W.; Valverde, F.; Coupland, G. Arabidopsis COP1 shapes the temporal pattern of CO accumulation conferring a photoperiodic flowering response. EMBO J. 2008, 27, 1277–1288. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Z.C.; Liu, H.T.; Liu, B.; Liu, X.M.; Lin, C.T. Blue light-dependent interaction of CRY2 with SPA1 regulates COP1 activity and floral initiation in Arabidopsis. Curr. Biol. 2011, 21, 841–847. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.W.; Matsui, M.; Ning, W.; Wagner, D.; Chu, A.M.; Feldmann, K.A.; Quail, P.H. COP1, an Arabidopsis regulatory gene, encodes a protein with both a zinc-binding motif and a G beta homologous domain. Cell 1992, 71, 791–801. [Google Scholar] [CrossRef]
- Liu, H.T.; Yu, X.H.; Li, K.W.; Klejnot, J.; Yang, H.Y.; Lisiero, D.; Lin, C.T. Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis. Science 2008, 322, 1535–1539. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.W.; Li, X.; Ma, D.B.; Chen, Z.; Wang, J.W.; Liu, H.T. CIB1 and CO interact to mediate CRY2-dependent regulation of flowering. EMBO Rep. 2018, 19, e45762. [Google Scholar] [CrossRef]
- Du, S.S.; Li, L.; Li, L.; Wei, X.X.; Xu, F.; Xu, P.B.; Wang, W.X.; Xu, P.; Cao, X.L.; Miao, L.X. Photoexcited cryptochrome 2 interacts directly with TOE1 and TOE2 in flowering regulation. Plant Physiol. 2020, 184, 485–505. [Google Scholar] [CrossRef]
- Hirose, F.; Shinomura, T.; Tanabata, T.; Shimada, H.; Takano, M. Involvement of rice cryptochromes in de-etiolation responses and flowering. Plant Cell Physiol. 2006, 47, 915–925. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Li, H.Y.; Li, R.B.; Hu, R.; Fan, C.M.; Chen, F.L.; Wang, Z.H.; Liu, X.; Fu, Y.F.; Lin, C.T. Association of the circadian rhythmic expression of GmCRY1a with a latitudinal cline in photoperiodic flowering of soybean. Proc. Natl. Acad. Sci. USA 2008, 105, 21028–21033. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.T.; Zhou, L.X.; Ma, Y.; Gao, J.; Li, W.L.; Piao, M.X.; Zeng, B.Z.; Yang, Z.M.; Bian, M.D. Cryptochrome 1b from sweet sorghum regulates photoperiodic flowering, photomorphogenesis, and ABA response in transgenic Arabidopsis thaliana. Plant Mol. Biol. Rep. 2017, 36, 13–22. [Google Scholar] [CrossRef]
- Yang, L.W.; Fu, J.X.; Qi, S.; Hong, Y.; Huang, H.; Dai, S. Molecular cloning and function analysis of ClCRY1a and ClCRY1b, two genes in Chrysanthemum lavandulifolium that play vital roles in promoting floral transition. Gene 2017, 617, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.T.; Yang, H.Y.; Guo, H.W.; Mockler, T.; Chen, J.; Cashmore, A.R. Enhancement of blue-light sensitivity of Arabidopsis seedlings by a blue light receptor cryptochrome 2. Proc. Natl. Acad. Sci. USA 1998, 95, 2686–2690. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.H.; Klejnot, J.; Zhao, X.Y.; Shalitin, D.; Maymon, M.; Yang, H.Y.; Lee, J.; Liu, X.M.; Lopez, J.; Lin, C.T. Arabidopsis cryptochrome 2 completes its posttranslational life cycle in the nucleus. Plant Cell 2007, 19, 3146–3156. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.F.; Zhang, M.F.; Jia, G.X. Analysis of global gene expression profiles during the flowering initiation process of Lilium × formolongi. Plant Mol. Biol. 2017, 94, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Zhao, Y.Q.; Zhang, M.; Jia, G.X.; Zaccai, M. Functional and evolutionary characterization of the CONSTANS-like family in Lilium × formolongi. Plant Cell Physiol. 2018, 9, 1874–1888. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Robertson, D.E.; Ahmad, M.; Raibekas, A.A.; Jorns, M.S.; Dutton, P.L.; Cashmore, A.R. Association of flavin adenine dinucleotide with the Arabidopsis blue light receptor CRY1. Science 1995, 269, 968–970. [Google Scholar] [CrossRef]
- Malhotra, K.; Kim, S.T.; Batschauer, A.; Dawut, L.; Sancar, A. Putative blue-light photoreceptors from Arabidopsis thaliana and Sinapis alba with a high degree of sequence homology to DNA photolyase contain the two photolyase cofactors but lack DNA repair activity. Biochemistry 1995, 34, 6892–6899. [Google Scholar] [CrossRef]
- Canamero, R.C.; Bakrim, N.; Bouly, J.P.; Garay, A.; Dudkin, E.E.; Habricot, Y.; Ahmad, M. Cryptochrome photoreceptors CRY1 and CRY2 antagonistically regulate primary root elongation in Arabidopsis thaliana. Planta 2006, 224, 995–1003. [Google Scholar] [CrossRef]
- Platten, J.D.; Foo, E.; Elliott, R.C.; Hecht, V.; Reid, J.B.; Weller, J.L. Cryptochrome 1 contributes to blue-light sensing in pea. Plant Physiol. 2005, 139, 1472–1482. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zuo, Z.C.; Wang, X.; Gu, L.F.; Yoshizumi, T.; Yang, Z.H.; Yang, L.; Liu, Q.; Liu, W.; Han, Y.J.; et al. Photoactivation and inactivation of Arabidopsis cryptochrome 2. Science 2016, 354, 343–347. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.T. Blue light receptors and signal transduction. Plant Cell 2002, 14, S207–S225. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.T.; Shalitin, D. Cryptochrome structure and signal transduction. Annu. Rev. Plant Biol. 2003, 54, 469–496. [Google Scholar] [CrossRef] [Green Version]
- Tóth, R.; Kevei, E.; Hall, A.; Millar, A.J.; Kozma-Bognár, L. Circadian clock-regulated expression of phytochrome and cryptochrome genes in Arabidopsis. Plant Physiol. 2001, 127, 1607–1616. [Google Scholar] [CrossRef]
- Chatterjee, M.; Sharma, P.; Khurana, J.P. Cryptochrome 1 from Brassica napus is up-regulated by blue light and controls hypocotyl/stem growth and anthocyanin accumulation. Plant Physiol. 2006, 141, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y.; Mao, K.; Zhao, C.; Zhang, R.F.; Zhao, X.Y.; Zhang, H.L.; Shu, H.R.; Hao, Y.J. Molecular cloning of cryptochrome 1 from apple and its functional characterization in Arabidopsis. Plant Physiol. Biochem. 2013, 67, 169–177. [Google Scholar] [CrossRef]
- Platten, J.D.; Foo, E.; Foucher, F.; Hecht, V.; Reid, J.B.; Weller, J.L. The cryptochrome gene family in pea includes two differentially expressed CRY2 genes. Plant Mol. Biol. 2005, 59, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Devlin, P.F.; Kay, S.A. Cryptochromes are required for phytochrome signaling to the circadian clock but not for rhythmicity. Plant Cell 2000, 12, 2499–2510. [Google Scholar] [CrossRef] [Green Version]
- Valverde, F.; Mouradov, A.; Soppe, W.; Ravenscroft, D.; Coupland, G. Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science 2004, 303, 1003–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, H.; Roussot, C.; Suarez-Lopez, P.; Corbesier, L.; Vincent, C.; Pineiro, M.; Hepworth, S.; Mouradov, A.; Justin, S.; Turnbull, C.; et al. CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis. Development 2004, 131, 3615–3626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-S.; Liau, Y.-E.; Huang, J.-C.; Wu, T.-D.; Su, C.-C.; Lin, C.H. Characterization of a desiccation-related protein in lily pollen during development and stress. Plant Cell Physiol. 1998, 39, 1307–1314. [Google Scholar] [CrossRef]
- Koornneef, M.; Hanhart, C.J.; Van, J.H.V.D. A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana. Mol. Gen. Genet. MGG 1991, 229, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Mao, K.; Zhao, C.; Zhao, X.Y.; Zhang, R.F.; Zhang, H.L.; Shu, H.R.; Hao, Y.J. Molecular cloning and functional analysis of a blue light receptor gene MdCRY2 from apple (Malus domestica). Plant Cell Rep. 2013, 32, 555–566. [Google Scholar] [CrossRef]
- Liu, Y.W.; Li, X.; Li, K.W.; Liu, H.T.; Lin, C.T. Multiple bHLH Proteins form heterodimers to mediate CRY2-dependent regulation of flowering-time in Arabidopsis. PLoS Genet. 2013, 9, e1003816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.F.; Liu, Q.; Jia, G.X. Reference gene selection for gene expression studies in lily using quantitative real-time PCR. Genet. Mol. Res. 2016, 15, gmr.15027982. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]
- Burow, M.D.; Chlan, C.A.; Sen, P.; Lisca, A.; Murai, N. High-frequency generation of transgenic tobacco plants after modified leaf disk cocultivation with Agrobacterium tumefaciens. Plant Mol. Biol. Rep. 1990, 8, 124–139. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.-M.; Yang, Z.-M.; Yang, L.-H.; Chen, J.-R.; Chen, H.-X.; Zheng, S.-X.; Zeng, J.-G.; Jia, G.-X.; Li, Y.-F. Cryptochrome 2 from Lilium × formolongi Regulates Photoperiodic Flowering in Transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 12929. https://doi.org/10.3390/ijms222312929
Wu X-M, Yang Z-M, Yang L-H, Chen J-R, Chen H-X, Zheng S-X, Zeng J-G, Jia G-X, Li Y-F. Cryptochrome 2 from Lilium × formolongi Regulates Photoperiodic Flowering in Transgenic Arabidopsis thaliana. International Journal of Molecular Sciences. 2021; 22(23):12929. https://doi.org/10.3390/ijms222312929
Chicago/Turabian StyleWu, Xiao-Mei, Zheng-Min Yang, Lin-Hao Yang, Ji-Ren Chen, Hai-Xia Chen, Si-Xiang Zheng, Jian-Guo Zeng, Gui-Xia Jia, and Yu-Fan Li. 2021. "Cryptochrome 2 from Lilium × formolongi Regulates Photoperiodic Flowering in Transgenic Arabidopsis thaliana" International Journal of Molecular Sciences 22, no. 23: 12929. https://doi.org/10.3390/ijms222312929
APA StyleWu, X.-M., Yang, Z.-M., Yang, L.-H., Chen, J.-R., Chen, H.-X., Zheng, S.-X., Zeng, J.-G., Jia, G.-X., & Li, Y.-F. (2021). Cryptochrome 2 from Lilium × formolongi Regulates Photoperiodic Flowering in Transgenic Arabidopsis thaliana. International Journal of Molecular Sciences, 22(23), 12929. https://doi.org/10.3390/ijms222312929