
Challenges and Perspectives in the Study of Self-Incompatibility in Orchids


Abstract
:1. Introduction
2. Morphological Types of SI in Orchidaceae
2.1. Subtribe Dendrobiinae
2.2. Subtribe Pleurothallidinae
2.3. Subtribe Oncidiinae
2.4. Subtribe Aeridinae
3. Physiology of SI in Orchidaceae
4. Molecular Mechanisms of SI in Orchidaceae
5. Challenges and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- De Nettancourt, D. Incompatibility and Incongruity in Wild and Cultivated Plants; Springer: New York, NY, USA, 2001. [Google Scholar]
- Dressler, R.L. Phylogeny and Classification of the Orchid Family; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Fujii, S.; Kubo, K.; Takayama, S. Non-self- and self-recognition models in plant self-incompatibility. Nat. Plants 2016, 2, 16130. [Google Scholar] [CrossRef]
- Wu, Q.; Han, T.-S.; Chen, X.; Chen, J.-F.; Zou, Y.-P.; Li, Z.-W.; Xu, Y.-C.; Guo, Y.-L. Long-term balancing selection contributes to adaptation in Arabidopsis and its relatives. Genome Biol. 2017, 18, 217. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.-L.; Zhao, X.; Lanz, C.; Weigel, D. Evolution of the S-Locus Region in Arabidopsis Relatives. Plant Physiol. 2011, 157, 937–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, G. Recent progress in plant reproduction research: The story of the male gametophyte through to successful fertilization. Plant Cell Physiol. 2009, 50, 1857–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopfer, C.R.; Nasrallah, M.E.; Nasrallah, J.B. The male determinant of self-incompatibility in Brassica. Science 1999, 286, 1697–1700. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Shiba, H.; Iwano, M.; Shimosato, H.; Che, F.S.; Kai, N.; Watanabe, M.; Suzuki, G.; Hinata, K.; Isogai, A. The pollen determinant of self-incompatibility in Brassica campestris. Proc. Natl. Acad. Sci. USA 2000, 97, 1920–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akagi, T.; Henry, I.M.; Morimoto, T.; Tao, R. Insights into the Prunus-specific S-RNase-based self-incompatibility system from a genome-wide analysis of the evolutionary radiation of S locus-related F-box genes. Plant Cell Physiol. 2016, 57, 1281–1294. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, M.J.; de Graaf, B.H.; Hadjiosif, N.; Perry, R.M.; Poulter, N.S.; Osman, K.; Vatovec, S.; Harper, A.; Franklin, F.C.; Franklin-Tong, V.E. Identification of the pollen self-incompatibility determinant in Papaver rhoeas. Nature 2009, 459, 992–995. [Google Scholar] [CrossRef]
- Yang, B.; Thorogood, D.; Armstead, I.; Barth, S. How far are we from unravelling self-incompatibility in grasses? New Phytol. 2008, 178, 740–753. [Google Scholar] [CrossRef]
- Igic, B.; Lande, R.; Kohn, J.R. Loss of self-incompatibility and its evolutionary consequences. Int. J. Plant Sci. 2008, 169, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Brewbaker, J.L.; Gorrez, D.D. Genetics of self-incompatibility in the monocot genera, Ananas (pineapple) and Gasteria. Am. J. Bot. 1967, 54, 611. [Google Scholar]
- East, E.M. The distribution of self sterility in the flowering plants. Proc. Am. Philos. Soc. 1940, 82, 449–518. [Google Scholar]
- Franklin-Tong, V.E. Self-Incompatibility in Flowering Plants—Evolution, Diversity, and Mechanisms; Springer: New York, NY, USA, 2008. [Google Scholar]
- Millner, H.J.; McCrea, A.R.; Baldwin, T.C. An investigation of self-incompatibility within the genus Restrepia. Am. J. Bot. 2015, 102, 487–494. [Google Scholar] [CrossRef] [Green Version]
- Freudenstein, J.V. Fundamentals of orchid biology. Nord. J. Bot. 1994, 14, 204. [Google Scholar] [CrossRef]
- Johansen, B. Incompatibility in Dendrobium (Orchidaceae). Bot. J. Linn. Soc. 1990, 103, 165–196. [Google Scholar] [CrossRef]
- Borba, E.L.; Barbosa, A.R.; Melo, M.; Gontijo, S.L.; Oliveira, H. Mating systems in the Pleurothallidinae (Orchidaceae): Evolutionary and systematic implications. Lankesteriana Int. J. Orchid. 2011, 11, 207–221. [Google Scholar]
- Castro, J.B.; Singer, R.B. A literature review of the pollination strategies and breeding systems in Oncidiinae orchids. Acta Bot. Bras. 2019, 33, 618–643. [Google Scholar] [CrossRef]
- Oh, G.S.; Chung, M.Y.; Chung, S.G.; Chung, M.G. Contrasting breeding systems: Liparis Kumokiri and L. Makinoana (Orchidaceae). Ann. Bot. Fenn. 2001, 38, 281–284. [Google Scholar]
- Whigham, D.F.; O’Neill, J.P. Dynamics of flowering and fruit production in two eastern North American terrestrial orchids, Tipularia discolor and Liparis lilifolia. In Population Ecology of Terrestrial Orchids; Wells, T.C.E., Willems, J.H., Eds.; SPB Academic Publishers: The Hague, The Netherlands, 1991; pp. 89–101. [Google Scholar]
- Aragón, S.; Ackerman, J.D. Density effects on the reproductive success and herbivory of Malaxis massonii. Lindleyana 2001, 16, 3–12. [Google Scholar]
- Ackerman, J.D. Limitations to Sexual Reproduction in Encyclia krugii (Orchidaceae). Syst. Bot. 1989, 14, 101–109. [Google Scholar] [CrossRef]
- Agnew, J.D. Self-Compatibility/Incompatibility in Some Orchids of the Subfamily Vandoideae. Plant Breed. 1986, 97, 183–186. [Google Scholar] [CrossRef]
- Li, Z.-J.; Wang, Y.; Yu, Y.; Zhang, Y.; Miao, K. Studies on floral and pollination biology in endangered Dendrobium orchid. Guangdong Agric. Sci. 2009, 231, 43–49. [Google Scholar]
- Pridgeon, A.M.; Cribb, P.J.; Chase, M.W.; Rasmussen, F.N. Genera Orchidacearum Volume 6: Epidendroideae; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Niu, S.C. Morphology and Molecular Mechanism of Dendrobium Self-Incompatibility. Ph.D. Thesis, The Chinese Academy of Sciences, Beijing, China, 2018. [Google Scholar]
- Niu, S.C.; Huang, J.; Xu, Q.; Li, P.X.; Yang, H.J.; Zhang, Y.Q.; Zhang, G.Q.; Chen, L.J.; Niu, Y.X.; Luo, Y.B.; et al. Morphological type identification of self-incompatibility in Dendrobium and its phylogenetic evolution pattern. Int. J. Mol. Sci. 2018, 19, 2595. [Google Scholar] [CrossRef] [Green Version]
- Huang, J. Study on Cross-Compatibility of The Genus Dendrobium. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2016. [Google Scholar]
- Singer, R.B.; Koehler, S. Notes on the Pollination Biology of Notylia nemorosa (Orchidaceae): Do pollinators necessarily promote cross pollination? J. Plant Res. 2003, 116, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Gontijo, S.L.; Barbosa, A.R.; de Melo, M.C. Occurrence of different sites of self-incompatibility reaction infour Anathallis (Orchidaceae, Pleurothallidinae) species. Plant Spec. Biol. 2010, 25, 129–135. [Google Scholar] [CrossRef]
- Duarte, M.O.; Oliveira, D.; Borba, E.L. Two self-incompatibility sites occur simultaneously in the same Acianthera species (Orchidaceae, Pleurothallidinae). Plants 2020, 9, 1758. [Google Scholar] [CrossRef]
- Borba, E.L.; Semir, J.; Shepherd, G.J. Self-incompatibility inbreeding depression and crossing potential in five Brazilian Pleurothallis (Orchidaceae) Species. Ann. Bot. 2001, 88, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, H. Dry stigmas, water and self-incompatibility in Brassica. Sex. Plant Reprod. 1995, 8, 1–10. [Google Scholar] [CrossRef]
- Elleman, C.J.; Franklin-Tong, V.; Dickinson, H.G. Pollination in species with dry stigmas—The nature of the early stigmatic response and the pathway taken by pollen tubes. New Phytol. 1992, 121, 413–424. [Google Scholar] [CrossRef]
- Duarte, M.O. Sementes Abortadas ou Óvulos não Fecundados? Investigação da Possível Ocorrência de Dois Sítios de Autoincompatibilidade em Acianthera (Orchidaceae). Ph.D. Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil, 2020. [Google Scholar]
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef] [Green Version]
- Neubig, K.M.; Whitten, W.M.; Williams, N.H.; Blanco, M.A.; Endara, L.; Burleigh, J.G.; Silvera, K.; Cushman, J.C.; Chase, M.W. Generic recircumscriptions of Oncidiinae (Orchidaceae: Cymbidieae) based on maximum likelihood analysis of combined DNA datasets. Bot. J. Linn. Soc. 2012, 168, 117–146. [Google Scholar] [CrossRef]
- Tremblay, R.L.; Ackerman, J.D.; Zimmerman, J.K.; Calvo, R.N. Variation in sexual reproduction in orchids and its evolutionary consequences: A spasmodic journey to diversification. Biol. J. Linn. Soc. 2005, 84, 1–54. [Google Scholar] [CrossRef]
- Zhang, Z. Conservation Biology of Three Phalaenopsis (Orchidaceae) Species in Hainan Island. Ph.D. Thesis, Hainan University, Haikou, China, 2018. [Google Scholar]
- Chai, L.; Ge, X.; Biswas, M.K.; Qiang, X.; Deng, X. Self-sterility in the Mutant ‘zigui Shatian’ Pummelo (Citrus grandis Osbeck) Is Due to Abnormal Post-zygotic Embryo Development and Not Self-incompatibility. Plant Cell Tissue Organ Cult. 2011, 104, 1–11. [Google Scholar] [CrossRef]
- Müller, R. Zur quantitativen Bestimmung von Indolylessigsaure mittels Papirchromatographie und Papirelektrophorese. Beitr. Biol. Pflanz. 1953, 30, 1–32. [Google Scholar]
- Arditti, J.; Flick, B.H. Post-pollination Phenomena in Orchid Flowers. Vi. Excised Floral Segments of Cymbidium. Am. J. Bot. 1976, 63, 201–211. [Google Scholar] [CrossRef]
- Heslop-Harrison, J. The physiology of reproduction in Dactylorchis. I. Auxin and the control of meiosis, ovule formation and ovary growth. Rotaniska Not. 1957, 110, 28–48. [Google Scholar]
- Gregory, L.E.; Gaskins, M.H.; Colberg, C. Parthenocarpic Pod Development By Vanilla planifolia Andrews Induced with Growth-regulating Chemicals. Econ. Bot. 1967, 21, 351–357. [Google Scholar] [CrossRef]
- Zhang, X.S.; O’Neill, S.D. Ovary and gametophyte development are coordinately regulated by auxin and ethylene following pollination. Plant Cell 1993, 5, 403–418. [Google Scholar] [CrossRef]
- Burg, S.P.; Dijkman, M.J. Ethylene and Auxin Participation in Pollen Induced Fading of Vanda Orchid Blossoms. Plant Physiol. 1967, 42, 1648–1650. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.C.; Huang, J.; Zhang, Y.Q.; Li, P.X.; Zhang, G.Q.; Xu, Q.; Chen, L.J.; Wang, J.Y.; Luo, Y.B.; Liu, Z.J. Lack of S-RNase-based gametophytic self-incompatibility in orchids suggests that this system evolved after the monocot-eudicot split. Front. Plant Sci. 2017, 8, 1106. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Eaves, D.J.; Sanchez-Moran, E.; Franklin, F.C.; Franklin-Tong, V.E. The Papaver rhoeas S determinants confer self-incompatibility to Arabidopsis thaliana in planta. Science 2015, 350, 684–687. [Google Scholar] [CrossRef]
- De Graaf, B.H.; Rudd, J.J.; Wheeler, M.J.; Perry, R.M.; Bell, E.M.; Osman, K.; Franklin, F.C.; Franklin-Tong, V.E. Self-incompatibility in Papaver Targets Soluble Inorganic Pyrophosphatases in Pollen. Nature 2006, 444, 490–493. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Peng, Z.; Tang, D.; Yang, Z.; Li, D.; Xu, Y.; Zhang, C.; Huang, S. Generation of self-compatible diploid potato by knockout of S-RNase. Nat. Plants 2018, 4, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.B.; Jia, J.S.; Wang, C.L. A general review of the conservation status of Chinese orchids. Biodivers. Sci. 2003, 11, 70–77. [Google Scholar]


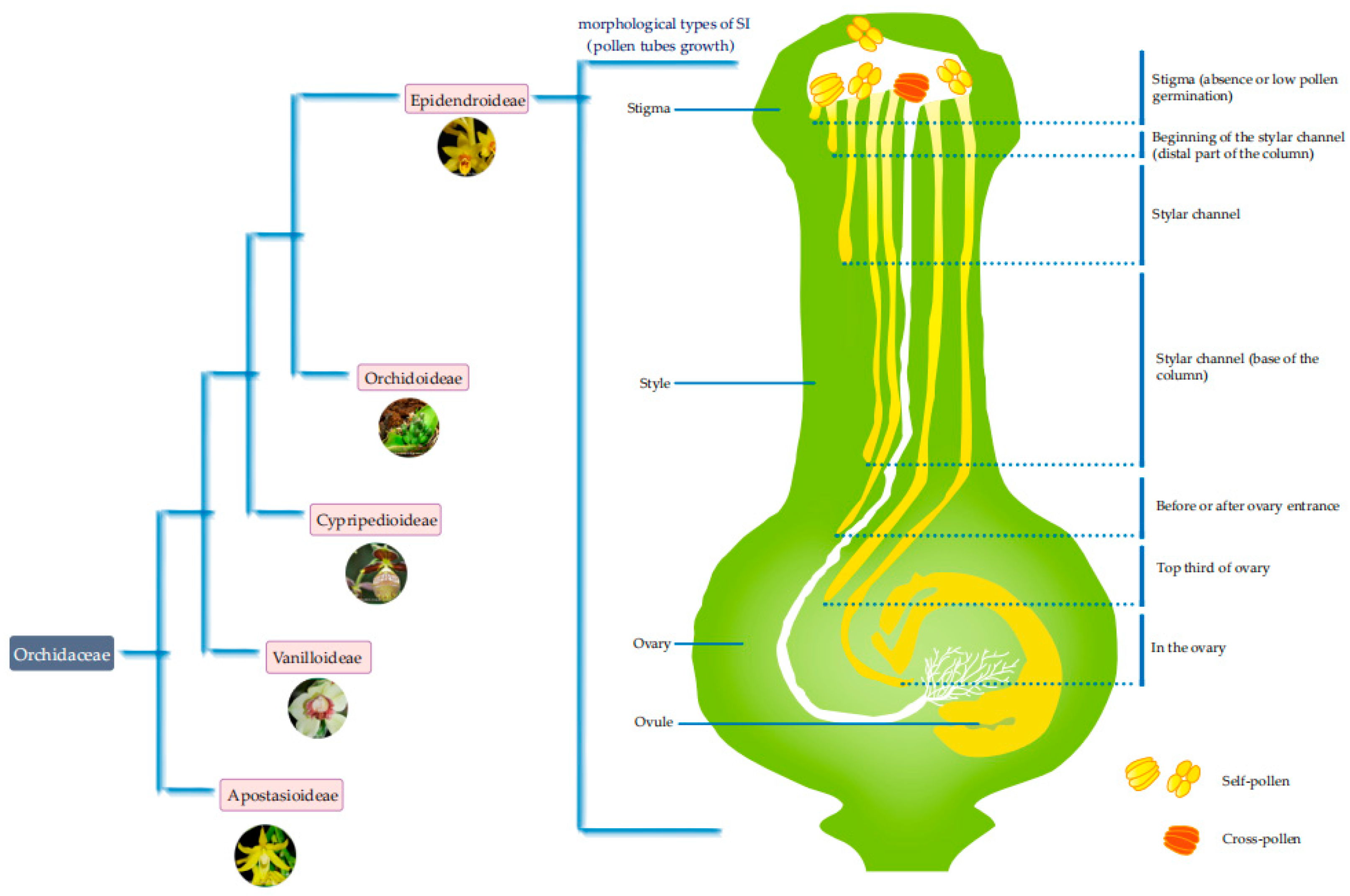{kind=link}
| Species | Site Where Pollen Tubes Stop Growing | Time When Pollen Tubes Stop Growing | Reference(s) |
|---|---|---|---|
| D. longicornu in Shenzhen | Stigma (absence or low pollen germination) | [28] | |
| D. cucullatum | [30] | ||
| Notylia nemorosa | [31] | ||
| Anathallis rubens1 | [19,32] | ||
| A. sclerophylla | [19,32] | ||
| A. heterophylla | [19,32] | ||
| Masdevallia infracta2 | [19] | ||
| Octomeria crassifolia3 | [19] | ||
| Octomeria grandiflora | [19] | ||
| Octomeria praestans | [19] | ||
| Stelis aff. Hypnicola4 | [19] | ||
| Stelis aff.peliochyla | [19] | ||
| Stelis sp. | [19] | ||
| Specklinia pristeoglossa5 | [19] | ||
| Specklinia sp. | [19] | ||
| D. densiflorum | Beginning of the stylar channel (distal part of the column) | 1 d | [28] |
| D. thyrsiflorum | 1 d | [28] | |
| D. farmeri | 4 d | [18] | |
| D. moniliforme | 3 d | [28] | |
| D. catenatum | 2–3 d | [28] | |
| D. longicornu in Yunnan | 4 d | [28] | |
| D. chrysanthum | 3 d | [28] | |
| Acianthera saurocephala6 | - | [19] | |
| D. unicum | Stylar channel | 4–5 d | [28] |
| D. devonianum | 4–5 d | [28] | |
| D. denneanum | 4–5 d | [28] | |
| A. johannensis | Stylar channel (base of the column) | 7 d | [33] |
| Acianthera adamantinensis | - | [19] | |
| A. fabiobarrosii | - | [19] | |
| Acianthera hamosa | - | [19] | |
| Acianthera limae | - | [19] | |
| Acianthera modestissima | - | [19] | |
| Acianthera ochreata | - | [19] | |
| Acianthera prolifera | - | [19] | |
| Acianthera teres | - | [19] | |
| Anathallis microphyta | 15 d | [32] | |
| Pleurothallis teres4 | - | [34] | |
| D. lindleyi | Before or after ovary entrance | 2 d | [28] |
| D. hancockii | 3–5 d | [28] | |
| D. jenkinsii | 3–5 d | [28] | |
| Restrepia brachypus7 | Top third of ovary | 21 d | [16] |
| Pleurothallis johannensis | In the ovary | - | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Jia, Y.; Liu, Y.; Chen, D.; Luo, Y.; Niu, S. Challenges and Perspectives in the Study of Self-Incompatibility in Orchids. Int. J. Mol. Sci. 2021, 22, 12901. https://doi.org/10.3390/ijms222312901
Zhang X, Jia Y, Liu Y, Chen D, Luo Y, Niu S. Challenges and Perspectives in the Study of Self-Incompatibility in Orchids. International Journal of Molecular Sciences. 2021; 22(23):12901. https://doi.org/10.3390/ijms222312901
Chicago/Turabian StyleZhang, Xiaojing, Yin Jia, Yang Liu, Duanfen Chen, Yibo Luo, and Shance Niu. 2021. "Challenges and Perspectives in the Study of Self-Incompatibility in Orchids" International Journal of Molecular Sciences 22, no. 23: 12901. https://doi.org/10.3390/ijms222312901
APA StyleZhang, X., Jia, Y., Liu, Y., Chen, D., Luo, Y., & Niu, S. (2021). Challenges and Perspectives in the Study of Self-Incompatibility in Orchids. International Journal of Molecular Sciences, 22(23), 12901. https://doi.org/10.3390/ijms222312901