1. Introduction
Lymphatic vessels transport lymph, consisting of interstitial fluid, immune cells, antigens, and other macromolecules, to the lymph nodes for antigen screening and then into blood circulation [
1]. The flow of lymph is perpetuated by an active process of lymphatic pumping, mediated by smooth muscle cells within the lymphatic vessel wall [
2]. Lymphatic pumping is tightly regulated by intrinsic and extrinsic factors, many of them derived from the endothelial or smooth muscle cells [
3,
4]. These factors include chemical mediators such as nitric oxide (NO) or prostaglandins [
5,
6,
7,
8].
The lymphatic system plays a critical role in osmoregulation, immune cell trafficking, and adaptive immune response. While lymphatic disfunction is frequently observed at sites of inflammation and infection, it is commonly overlooked due to an incomplete understanding of its role in the disease’s pathology. These changes result in a variety of outcomes, including decreased fluid drainage (lymphedema) and reduced nutrient absorption (malnutrition).
Statins are a group of successful cholesterol lowering medications. They are all competitive inhibitors of 3-hydroxy-methylglutarl-CoA (HMG-CoA) reductase, the rate limiting enzyme in the biosynthesis of cholesterol [
9]. Therefore, statin-therapy potently reduces hypertension and the risk of cardiovascular diseases. Unfortunately, statin treatment sometimes results in patients developing myopathies, pain, and weakness in skeletal muscle, which in rare cases can lead to fatal rhabdomyolysis [
10,
11,
12,
13,
14]. While, all commercially available statins have been reported to cause muscle-related side effects, the occurrence of such events is too inconsistent to identify the direct cause in the patient cohort [
15]. Interestingly, the mechanisms underlying this toxicity have been partially identified, although not fully elucidated. Schirris and colleagues recently suggested an off-target interaction of lactone rings, structures found within some statins, were inhibiting mitochondrial respiration, thus inducing apoptosis, a phenomenon that was reduced with the use of the hydrolyzed statin forms [
11]. It was also reported that in isolated muscle fibers, certain statins induced the release of Ca
2+ from the sarcoplasmic reticulum thereby directly affecting the function of the ryanodine receptor RyR1—the predominant receptor involved in skeletal muscle calcium release. The experimental evidence was supported by naturally occurring mutations surrounding RyR1 within the patient cohort, which is associated with many serious musculoskeletal diseases [
11].
Lipophilicity of statins has previously been correlated to increased occurrences of myopathies. Lipophilic drugs increase dissemination throughout the body as they have an increased ability to passively diffuse through thin membrane barriers and single cell barriers such as capillaries, endothelial layers, and the blood brain barrier. Importantly, the lymphatic system participates directly in the increased uptake of lipophilic molecules. This allows for medications, such as lipophilic-statins, to be given at lower dosages but maintain their biological impact [
16].
Simvastatin and Lovastatin are two of the most prescribed statins and are available in a lipophilic lactone close-ringed form or an open-ringed hydroxyacid form. Both medications are regularly prescribed in their closed lactone form, which is converted in vivo, into their active form. With previous groups demonstrating the effect of statin prodrug-forms on skeletal muscle function, we hypothesized that a similar effect would be seen in vascular smooth muscle, specifically those of the lymphatic system. Here, we tested several statins for their acute effect on lymphatic vascular function. We describe novel mechanisms of actions of the closed lactone-ringed statins, Lovastatin and Simvastatin on mesenteric lymphatic vessel function through interference with key components of the lymphatic smooth muscle contractile apparatus. The resulting lymphatic pumping alteration has a biological effect on dietary lipid uptake and dissemination in vivo, suggesting a non-canonical means of reducing cholesterol levels through statin therapy independent of the targeted HMG-CoA-reductase inhibitory function.
3. Discussion
Statin-induced myopathies present a medical phenomenon with pathological implications. While prodrug forms of statins containing a lactone ring have been reported to bind to the Q
o site of mitochondrial complex III, providing an explanation for the association of some statins with skeletal muscle myopathies [
11], it is unknown whether the effect is conserved in other systems containing smooth muscle. We hypothesized that lymphatic smooth muscle cells could be similarity affected, thereby altering their functional activity.
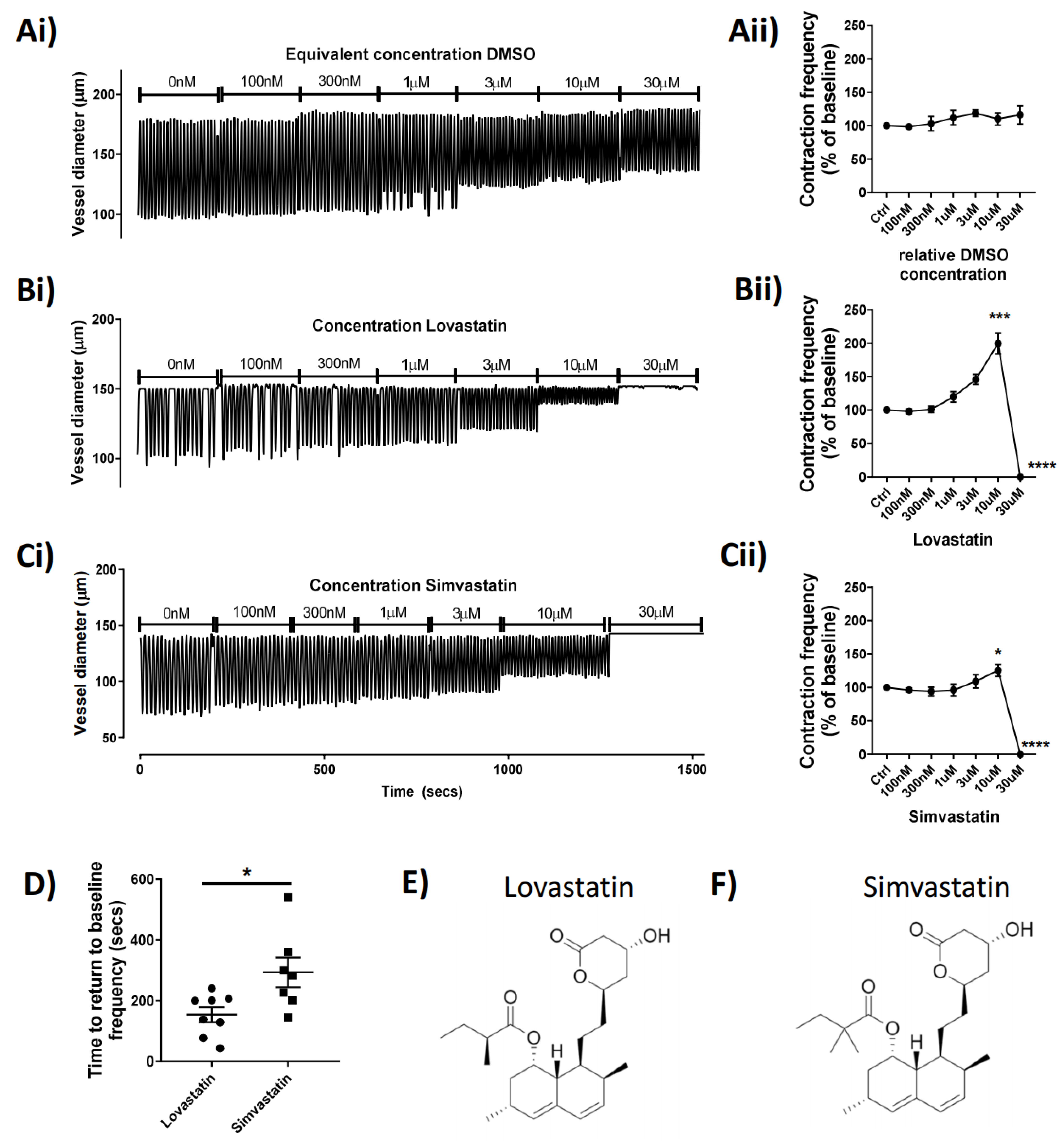
In the present study, we aimed to elucidate the effect of popular commercialized statins on lymphatic vascular function, and we discovered a previously unreported effect of Lovastatin and Simvastatin on the vascular lymphatic system. We demonstrated that these statins in their pro-drug forms impair in a concentration-dependent manner the phasic and rhymical contractions of mesenteric lymphatic vessels, an effect that culminates with a complete loss of tone, suggesting an inability of the vessels to effectively propel lymph. Furthermore, the action of these two statins, while acute, is reversible and seemingly solely dependent on the lactone ring structures found within the pro-drug forms of both Lovastatin and Simvastatin (
Figure 2). Other tested commercial statins (Atorvastatin and Pravastatin) that lack the lactone ring had no impact on lymphatic vessel function, suggesting the importance of this structural moiety (
Supplemental Figure S1).
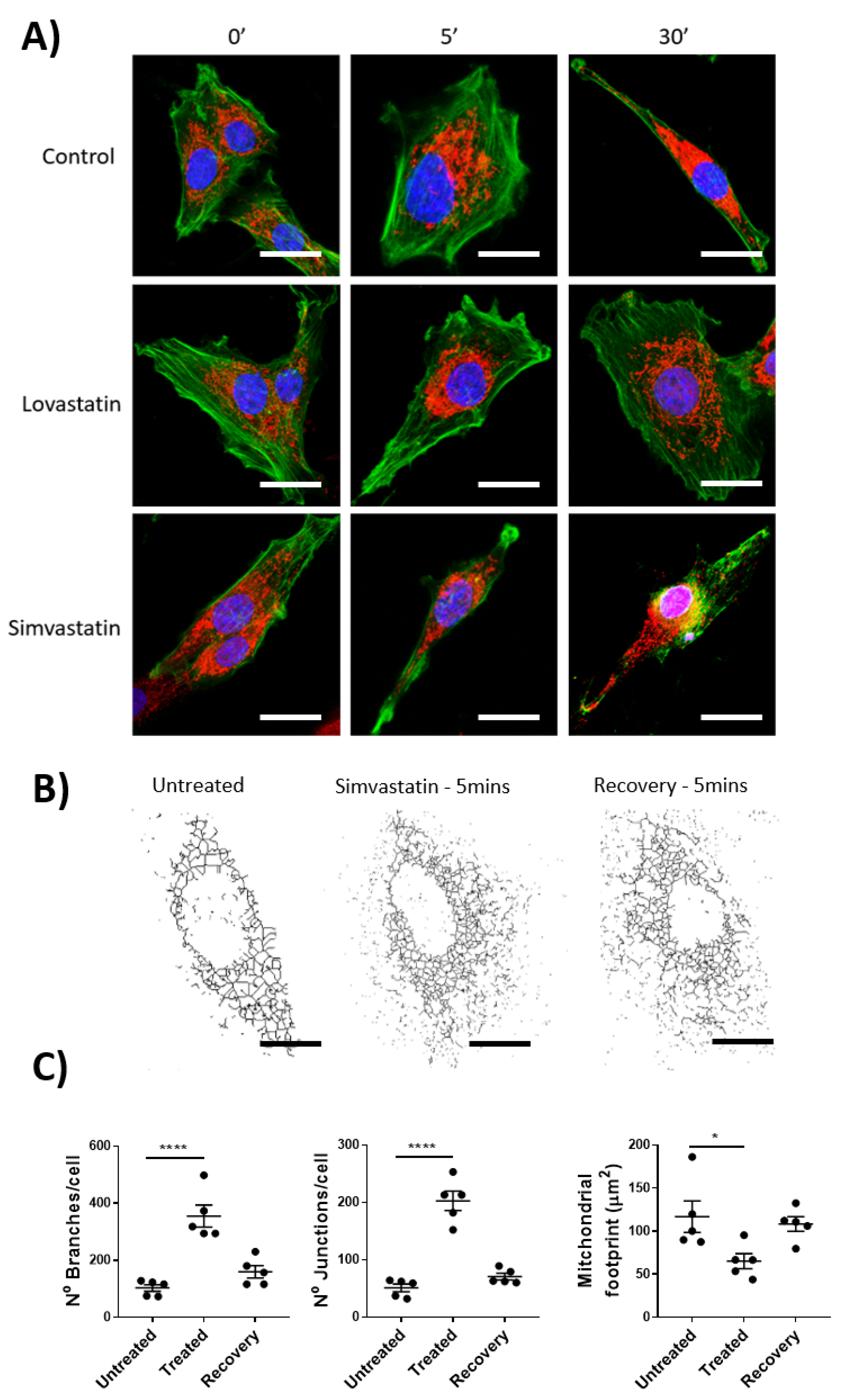
The effects of statins on mitochondrial function is a developing field of research highlighting not only the deleterious effects of statin use, but also novel protective effects. As HMG-CoA is a key enzyme in the mevalonate pathways, its inhibition (by statins) results in the reduced availability of farnesyl pyrophosphate, geranylgeranyl pyrophosphate, heme A, coenzyme Q10, and other metabolites essential for cellular function [
22]. Moreover, as cholesterol is not the end-product of these pathways, there can be a variety of pleiotropic effects with chronic use [
23]. Lactone ring functional groups, contained within several lipophilic statins, have been recently presented as potential inhibitors of the Q
o of the mitochondrial cytochrome complex III subunit [
11]. Using isolated rat lymphatic smooth muscle cells, paired with mitochondrial staining and imaging, we demonstrated that the pro-drug form of Simvastatin, but not Lovastatin, had a significant impact on mitochondrial dynamics. These data, while demonstrating a partially conserved mechanism of action between skeletal and vascular smooth muscle cells, highlight the ambiguity between cells of differing tissue origin, their associated function, and the impact of drug effects. We acknowledge that due to the absence mitochondrial-function assessment through specialized assays such as Seahorse
TM, we cannot definitively prove the absence of a mitochondrial complex III inhibitory effect as has been previously published [
11].
The collecting lymphatic vessel is comprised of an endothelial tube surrounded by specialized lymphatic smooth muscles. While the force and rhythmicity of lymphatic contractions is generated in the lymphatic smooth muscle layer, there is clearly documented regulation of pumping through molecules derived from the endothelium, specifically NO and prostaglandins [
5,
7,
8]. Several reports highlighted an increased production of NO by statins in a way that is independent of its canonical HMG-CoA reductase inhibitory activity by either involving scavenger receptor-B1 [
24] or LOX-1, a receptor for oxidized low-density lipoprotein [
25]. Furthermore, statin treatment was also shown to induce the production of prostaglandins in rat liver [
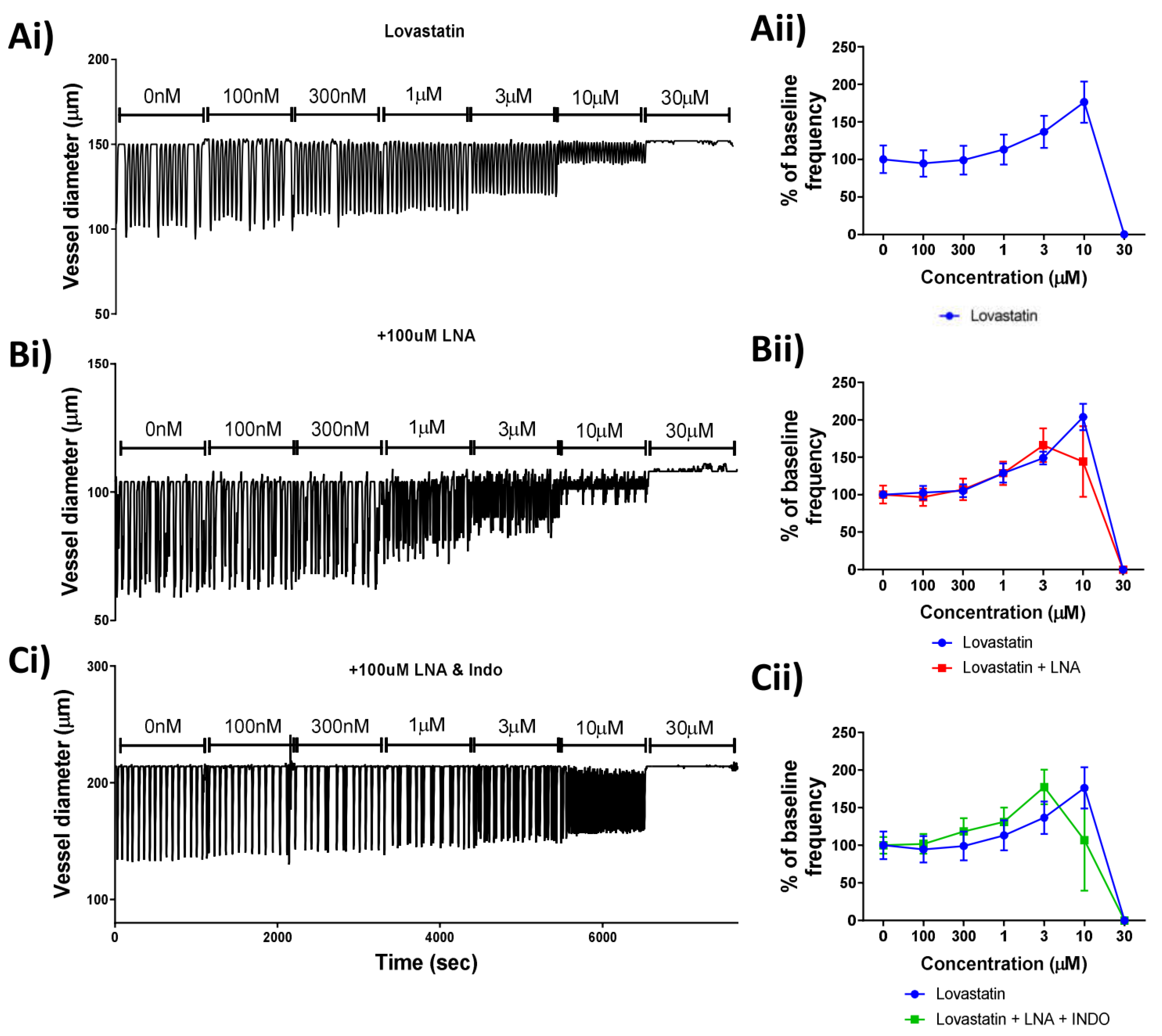
26]. As the production of either product would likely result in the reduction/ablation of vessel contractility that we observed, we performed a combination of ex vivo lymphatic vessel pressure myography and pharmacological interventions to assess whether the endothelium could play a role in the statin-induced lymphatic disfunction. Our findings determined that the action of Lovastatin on vessel contraction was independent of the production of either of these mediators. The highly specific role of FOXC2-positive lymphatic endothelial cells (such as those used here) for their controlled specialization and function within the gastrointestinal tract may provide an explanation for the differing findings when compared to the cardiovascular literature [
27].
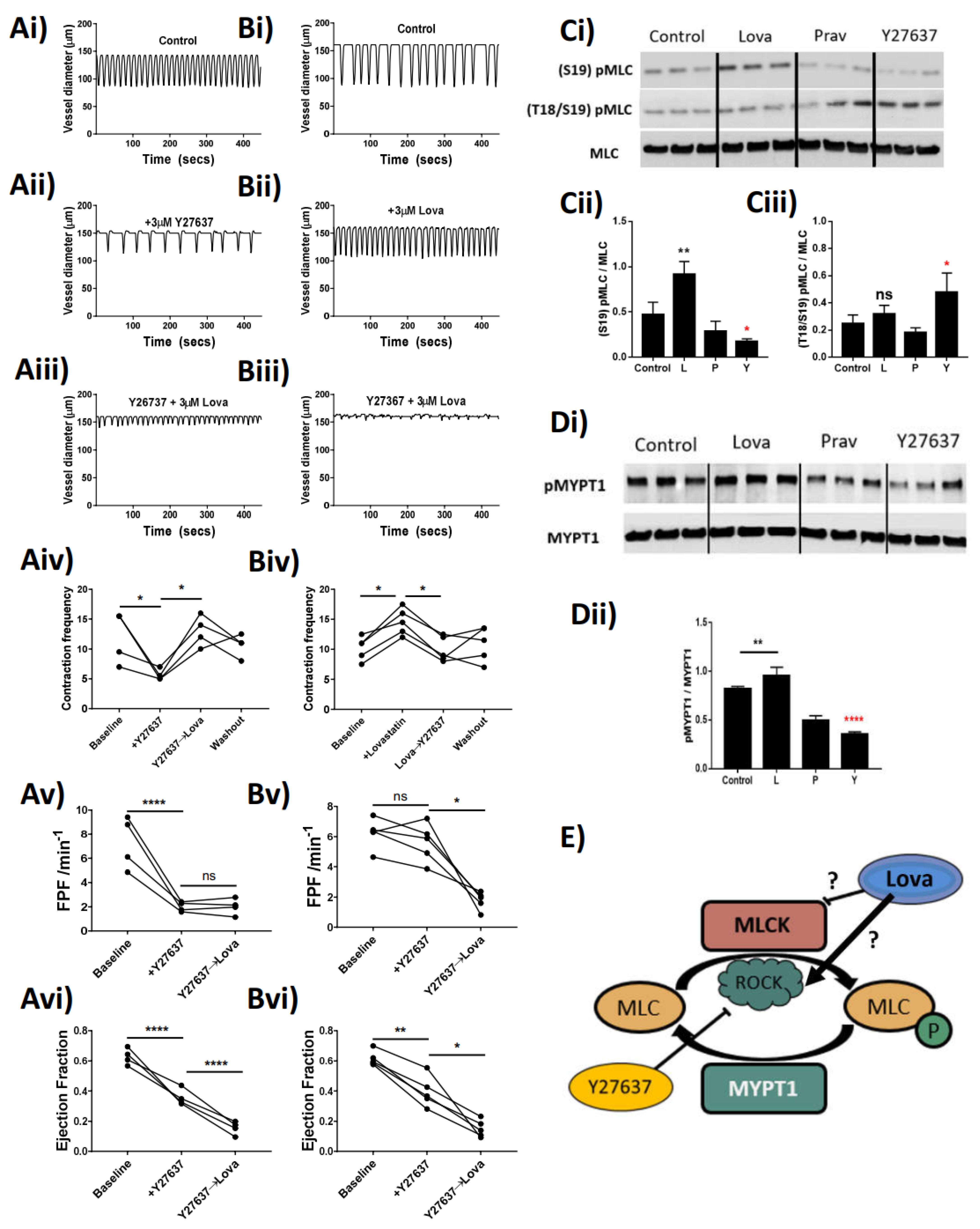
The phasic nature of lymphatic vessel contractility is intrinsically generated in lymphatic smooth muscle cells. As in other smooth muscles, this contraction–relaxation cycle relies on the phosphorylation–dephosphorylation of myosin light chains (MLC) regulating its impact on the actin cytoskeleton [
21] and providing the necessary force for the propulsion of lymph. The regulation of MLC phosphorylation depends on the essential activity of regulatory kinases such as ROCK, myosin light chain kinase (MLCK), and myosin light chain phosphatase (MLCP) via subunits such as MYPT1. Statins act to inhibit the function of HMG-CoA reductase, and as such can have an indirect effect on the activity of ROCK through downregulation of the production of geranylgeranyl pyrophosphate and the associated rhoA synthesis [
28]. There is also evidence within the literature that shows direct in vivo inhibition of ROCK itself by statins [
29]. Our data demonstrate that Lovastatin treatment negates the effects of a ROCK inhibitor (Y27637) on lymphatic contractility, through a proposed action of increasing the function of residual ROCK within the system. We support this claim through the analysis of isolated primary cultured rat mesenteric lymphatic smooth muscle cells, whereby treatment with Lovastatin causes a hyper-phosphorylation of both MLC2 and MYPT1, proteins whose phosphorylation sites are regulated by ROCK. We propose that through this increased functionality of ROCK, the vessel stops contracting as the dephosphorylation of MLC is outweighed by active phosphorylation within the cell (proposed schematic
Figure 5E). Our findings contrast published reports in which other statins (Atorvastatin) have been described to potently reduce ROCK activity in patients with atherosclerosis [
29]. However, whether this is a direct inhibition of the functionality of ROCK or an indirect inhibition through another mechanism is still not fully elucidated and as such remains an area for further investigation. Furthermore, as that study assessed the levels of ROCK activity in circulating leukocytes, it is possible the effect, and therefore function of statins therapy, is unique between stromal and immune cells.
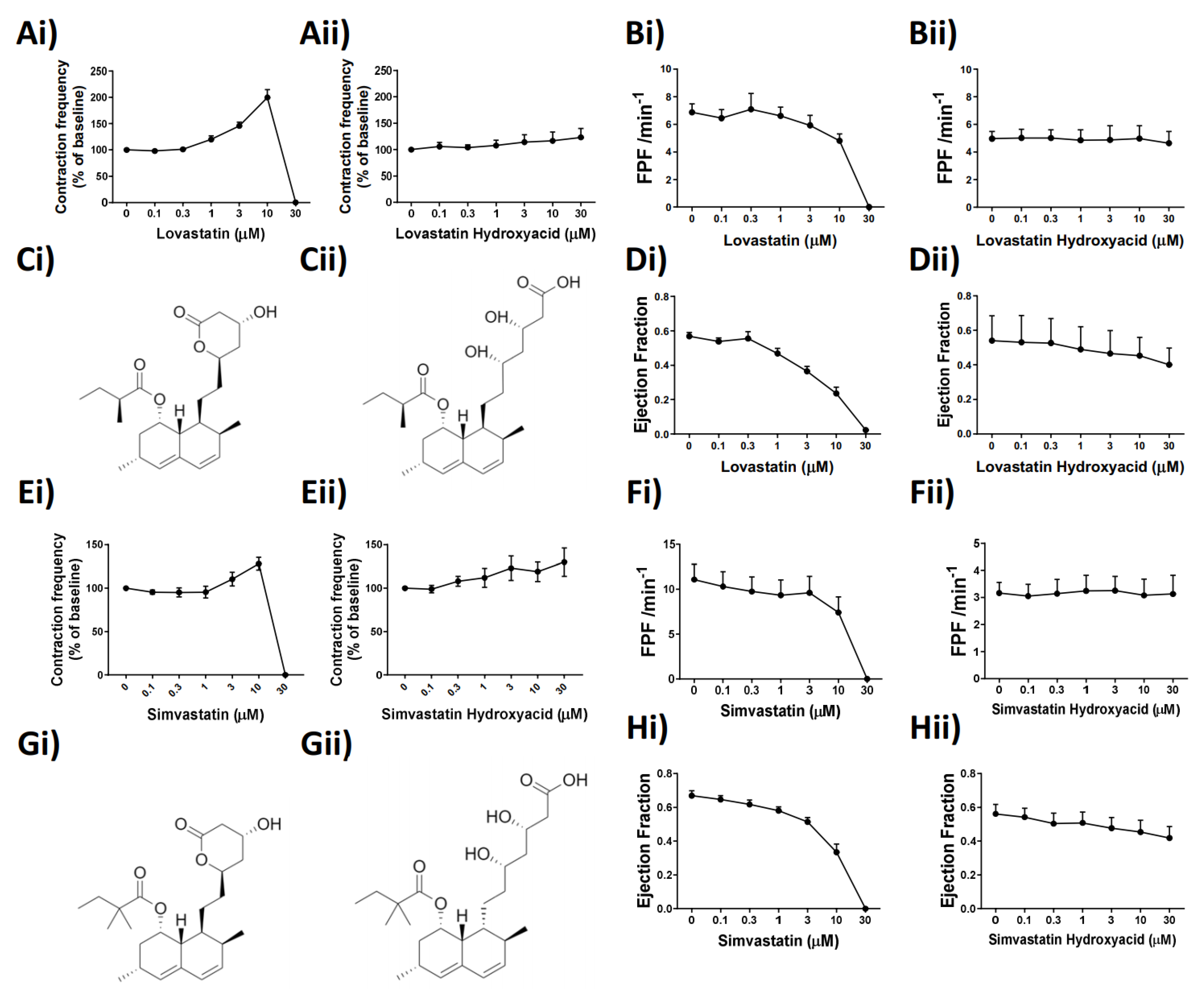
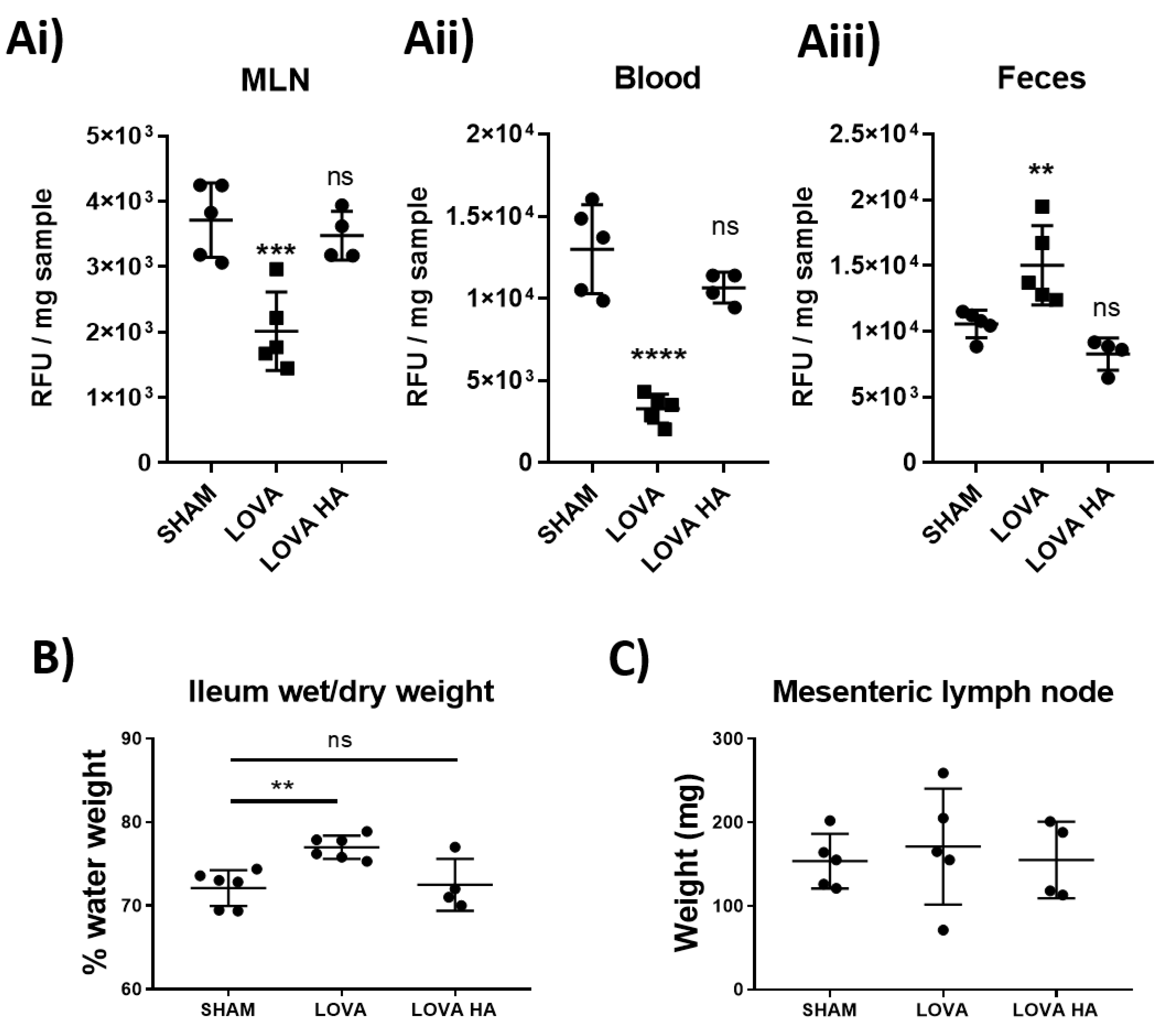
Analysis of lymphatic vessel function using pressure myography clearly demonstrated that Lovastatin treatment caused a significant loss in lymphatic vessel ejection fraction and fractional pump flow (
Figure 5). We posited that this alteration would drastically reduce the ability of the intestinal mesenteric lymphatic system to disseminate dietary absorbed lipids. In line with this hypothesis, we tested and report on a pronounced disruption in dietary cholesterol uptake and dissemination in rats treated acutely with Lovastatin (
Figure 6). This effect was confirmed to be prodrug specific as no impact was observed with treatment of Lovastatin hydroxyacid. These findings reveal a novel mechanism whereby statins actively lower circulating cholesterol by decreasing intestinal lymphatic lipid absorption, transport, and peripheral dissemination. The mechanism is promoted by the action of the lactone ring statin pro-form of Lovastatin, likely transported through the mesenteric lymphatic vessels prior to their enzymatic conversion during first-pass metabolism.
Overall, our study highlights the off-target effect of Lovastatin and Simvastatin pro-form drugs on the lymphatic vasculature. Our novel findings outline a non-canonical mechanism by which Lovastatin regulates the function of the lymphatic smooth muscle cell contractile apparatus, an action that explains the decrease in cholesterol distribution we observed in vivo (
Figure 5). While these data support our overarching hypothesis, we have outstanding questions. Currently we do not fully understand why the treated lymphatic vessels stop pumping in a dilated rather than contracted state. We propose that this dilation is caused by dissociation of pMLC2 from the actin cytoskeleton in a “cutting-of-strings” event that has yet to be witnessed and requires future investigation.
4. Materials and Methods
4.1. Animals
Male Sprague Dawley rats (200–300 g, Charles River, Montreal, QC, Canada) were used to perform all experiments. Animals were housed at a room temperature of 22 °C with 12 h day-and-night cycles and were allowed full access to water and food at all times; both normal circadian rhythms and body temperatures were maintained. All animal protocols were reviewed and approved by the University of Calgary Animal Care and Ethics Committee and were conducted in accordance with the guidelines of the Canadian Council on Animal Care and the National Institutes of Health’s Guide for the Care and Use of Laboratory Animals (Certification No. AC16-187 and AC20-0140).
4.2. Lymphatic Vessels Preparation
Rats were anaesthetized by isofluorane (Pharmaceutical Partners of Canada, Richmond Hill, ON, Canada) inhalation in a sealed box and euthanized by decapitation. The small intestine including jejunum and ileum was exteriorized after laparotomy and kept in albumin physiological saline solution (APSS; in mM: 145.0 NaCl, 4.7 KCl, 2 CaCl2, 1.2 MgSO4, 1.2 NaH2PO4, 5.0 Dextrose, 2.0 Sodium Pyruvate, 0.02 EDTA, 3.0 MOPS, and 5 g/L bovine serum albumin (BSA)) with pH adjusted to 7.4 at room temperature. Pieces of ileal mesentery containing mesenteric arcades (artery, vein, and two collecting lymphatic vessels on each side of the blood vessels) were isolated from the gut and loosely pinned down on a Sylgard-coated Petri dish filled with APSS until ready to use. Individual arcades were transferred to a clean, APSS-filled, Sylgard-coated Petri dish, set on the stage of a dissection microscope and collecting lymphatic vessels were then carefully dissected out of the mesentery and surrounding adipose tissue.
4.3. Pressure Myography
Following dissection, vessels were placed in a 2 mL pressure myograph chamber (Culture Myograph, DMT-USA Inc., Ann Arbor, MI) filled with APSS with pH adjusted to 7.4 at 37 °C, and they were cannulated with two pulled and matched glass micropipettes (original size: OD 1.2 mm, ID 0.69 mm; approximate final ID 80–120 μm). Lymphatic vessels were tied at each end with a fine suture to the glass pipettes, which restricted fluid leakage as well as pressure drops inside the lymphatic vessel. Each pipette was connected to the same adjustable pressure reservoir filled with APSS (pH adjusted to 7.4 at 37 °C). The vessel containing chamber was then placed onto a stage with built-in heating over an integrated DMT inverted microscope. Having both pipettes connected to the same reservoir ensured that no flow was generated by pressure differences and pressure inside the vessel (transmural pressure) was constant and equal to that determined by the height of the single reservoir. The organ bath was superfused at a rate of 0.25 mL/min with APSS. Vessel leak, integrity, and twist were assessed first by applying a positive transmural pressure (3–5 cmH
2O) and then approximate in situ length was established by briefly increasing the transmural pressure to 10 cmH
2O. The vessel was then set to a transmural pressure of 3 cmH
2O for a 30 min equilibration period, and then to 5 cmH
2O for the duration of the experiments. Vessel contractions were recorded via a USB CCD video camera with the MyoView software [
30]. To assess endothelial integrity, acetylcholine was added to the bath at a concentration of 10 μM and a transient (20 to 60 s) relaxation was observed. When used, blockers of eNOS, COX, and guanylate cyclase (N
G-nitro-L-arginine (L-NA) or indomethacin, respectively) were added directly to the superfusion, vessels were then left for 20 min before applying further treatments. Endothelium-dependent responses were abolished in the presence of LNA or indomethacin. Statins were added directly into the bath and mixed by pipetting while superfusion was stopped during those applications to prevent dilutions. The rho-associated protein kinase (ROCK) inhibitor Y-27632 or myosin light-chain kinase (MLCK) inhibitor ML9 was added directly to the bath, and their effect on contractile activity occurred within minutes.
4.4. Western Blot
Isolated lymphatic vessels were lysed in extraction buffer, loaded on SDS polyacrylamide gels, and subjected to electrophoresis (120 V, 90 min). Proteins in the separating gel were then transferred to 0.2 μm nitrocellulose membranes (Bio-Rad Laboratories (Canada) Ltd., Mississauga, ON, Canada) and fixed by complete dehydration. A blocking solution of 5% BSA in 0.1% TBST (a mixture of Tris-buffered saline (TBS, 150 mM NaCl, 25 mM Tris HCl with pH 7.5) and Tween 20 (0.1%) with 5% bovine serum albumin (BSA) for 60 mins was employed to minimize non-specific binding. The membranes were then incubated with primary antibody targeting p-MLC (Ser18/19 and Ser20), MLC, pMYPT1, MYPT1 (all 1:1000 dilution; New England Biolabs Canada, Whitby, ON, Canada), and β-actin (1:10,000 dilution; Biolegend, San Diego, CA, USA) in blocking solution overnight, at 4 °C. After washing, the membranes were then incubated with the appropriate secondary antibodies, anti-Donkey or anti-Mouse IgG-horseradish peroxidase-conjugated secondary antibody (1:5000 dilution; Jackson Immunoresearch, West Grove, PA, USA), for 60 min. After extensive washing in TBST, the chemiluminescence signal was detected with West Dura Substrate (ThermoFisher Scientific, Burlington, ON, Canada). Images were acquired with the Bio-Rad Touch chemiluminescence imaging software and blots quantified using the open-source image processing package Fiji (Previously ImageJ) [
31,
32]. Levels of phosphorylated MLC or MYPT1 were normalized to their total protein expressions, respectively.
4.5. Assessment of Mitochondrial Function via Immunofluorescence Imaging
Lymphatic smooth muscle cells were kindly provided by Dr Sanjukta Chakraborty (Texas A&M University) and were cultured as published [
33]. Cells were plated on 8-well chamber slides 48 h prior to stimulation at 2000 cells/well. Before treatments, cells were pulse-chased with Mitotracker, CFMXrROS (Thermo Fisher Scientific, Burlington, ON, Canada) for 30 min. After treatments, cells were immediately fixed in cold 4% formalin for 20 min without agitation and then transferred to PBS. Cells were stained with Alexa-488 conjugated Phalloidin for 20 min (1 ng/mL; Thermo Fisher Scientific, Burlington, ON, Canada) in PBST before slides were washed and mounted in Vectorshield Fluosave with DAPI (Vector Laboratories, Burlingame, CA, USA). Slides were imaged immediately on a Leica SP8 inverted confocal microscope and images were post-processed using Leica’s LASX software [
34]. Images were taken using sequential scanning on three separate channels to remove all possible overlap near the laser and all images were taken at 63× oil immersion at designated digital zoom. Five cells per treatment were randomly selected, imaged, and the mitochondrial network was assessed as previously published.
4.6. Lymphatic Function Assessment
After a two-hour fasting period, rats were injected intra-peritoneally with 5 mg/kg of the statin of choice. Two hours after statin injection, animals were gavaged with 300 µL of oelic acid containing 100 µg of Bodipy.FL C16 (Thermo Fisher Scientific, Burlington, ON, Canada) One-hour post-gavage, rats were euthanized as described above. The mesenteric lymph node, fecal pellet, and blood were collected for downstream processing. Blood was left to coagulate at room temperature for 15 min before being spun at 3000× g for 10 min to clarify the serum that was then collected and kept on ice. Mesenteric lymph nodes and fecal pellets were weighed and 10× w/v of sterile PBS was added to the sample. Each sample was then homogenized through a 100 µm filter to remove large debris and cells separated by centrifugation at 3000× g for 10 min. The clarified supernatant was collected and kept on ice. Total Bodipy concentration was then analyzed using a Victor X3 fluorescent plate reader (FITC ex 488 nm/em 535 nm) and compared to solution and sample auto fluorescent values from non-gavaged rat controls. All values were then baselined against the reading in wells that only contained PBS.
4.7. Chemicals
Statins were purchased from Cayman Chemical (Ann Arbor, MI, USA) and all other chemicals were from Millipore Sigma (Oakville, ON, Canada) unless otherwise specified.
4.8. Data and Statistical Analysis
Contraction data were expressed relative to each vessel’s own sham control value. Statistical significance was then assessed on the raw data using ANOVA with an appropriate post-hoc test, as indicated in the text. Experimental data were expressed as mean values ± one standard error of the mean (SEM) and p-values less than 0.05 were considered significant.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}