
Doxycycline Ameliorates the Severity of Experimental Proliferative Vitreoretinopathy in Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
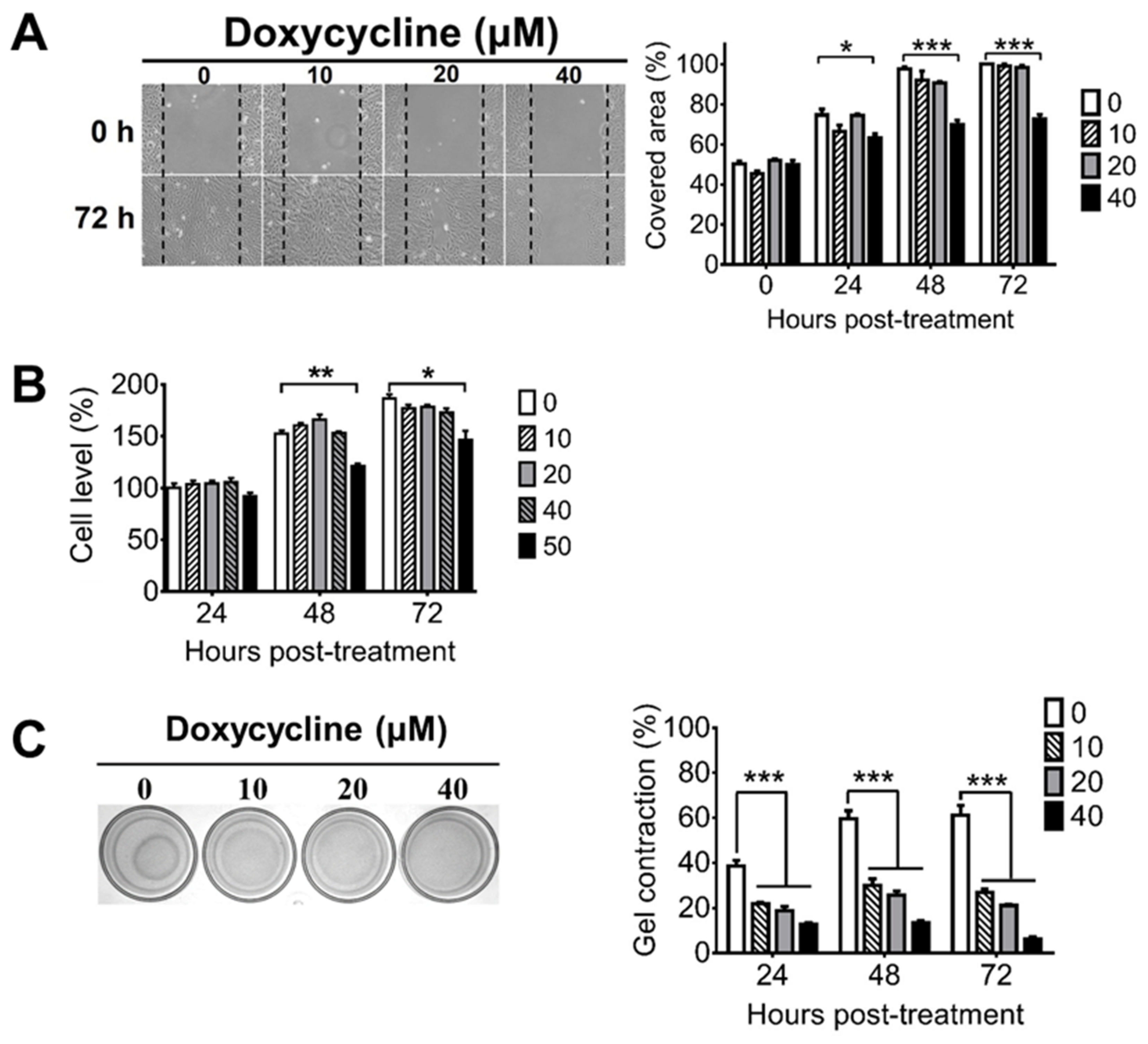
2.1. Doxycycline Decreases the Migration, Proliferation, and Contraction of ARPE-19 Cells
2.2. Doxycycline Diminishes the Total MMP Activity and p38 MAPK Activation of ARPE-19 Cells
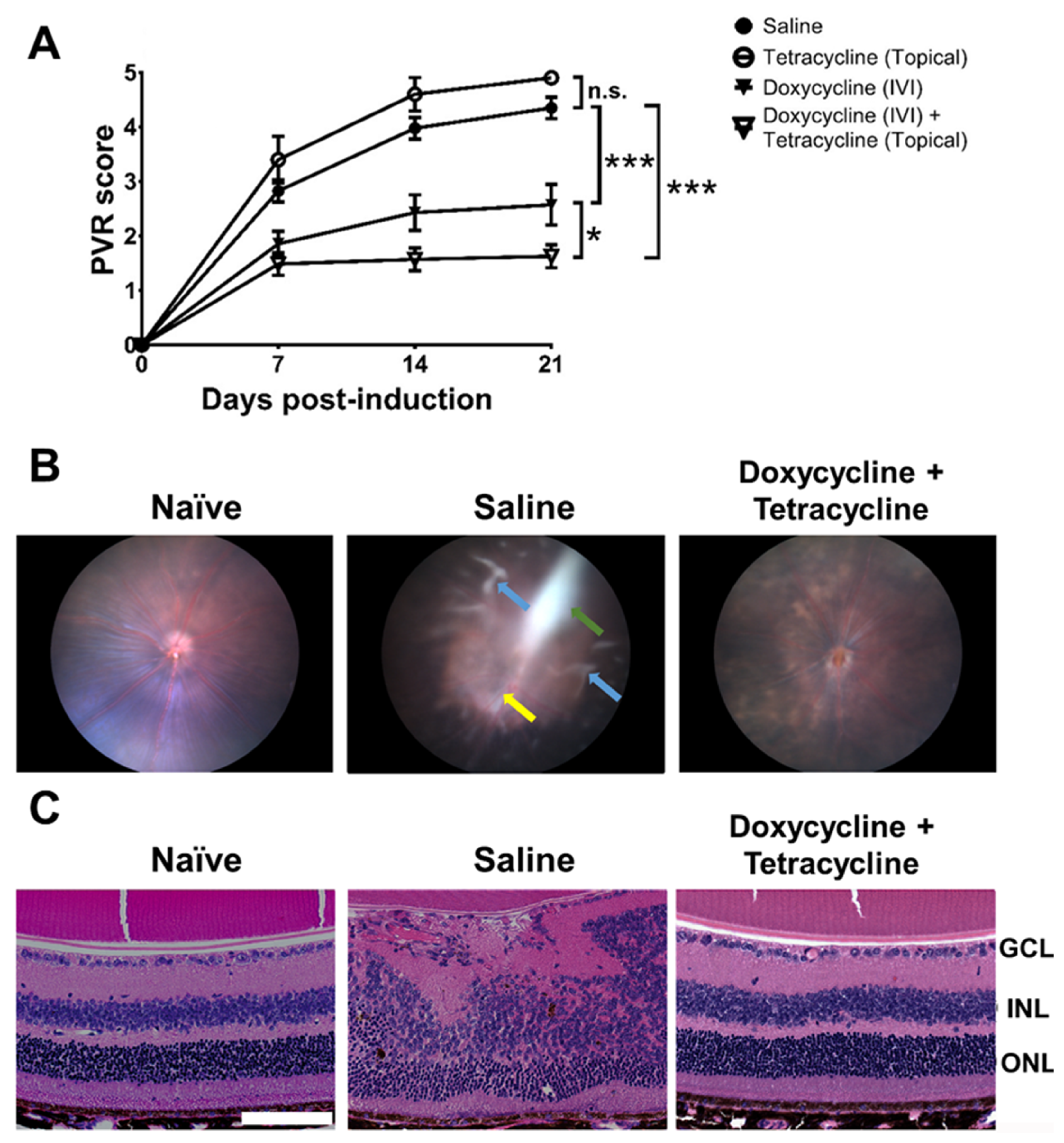
2.3. Doxycycline Reduces PVR Severity in Mice
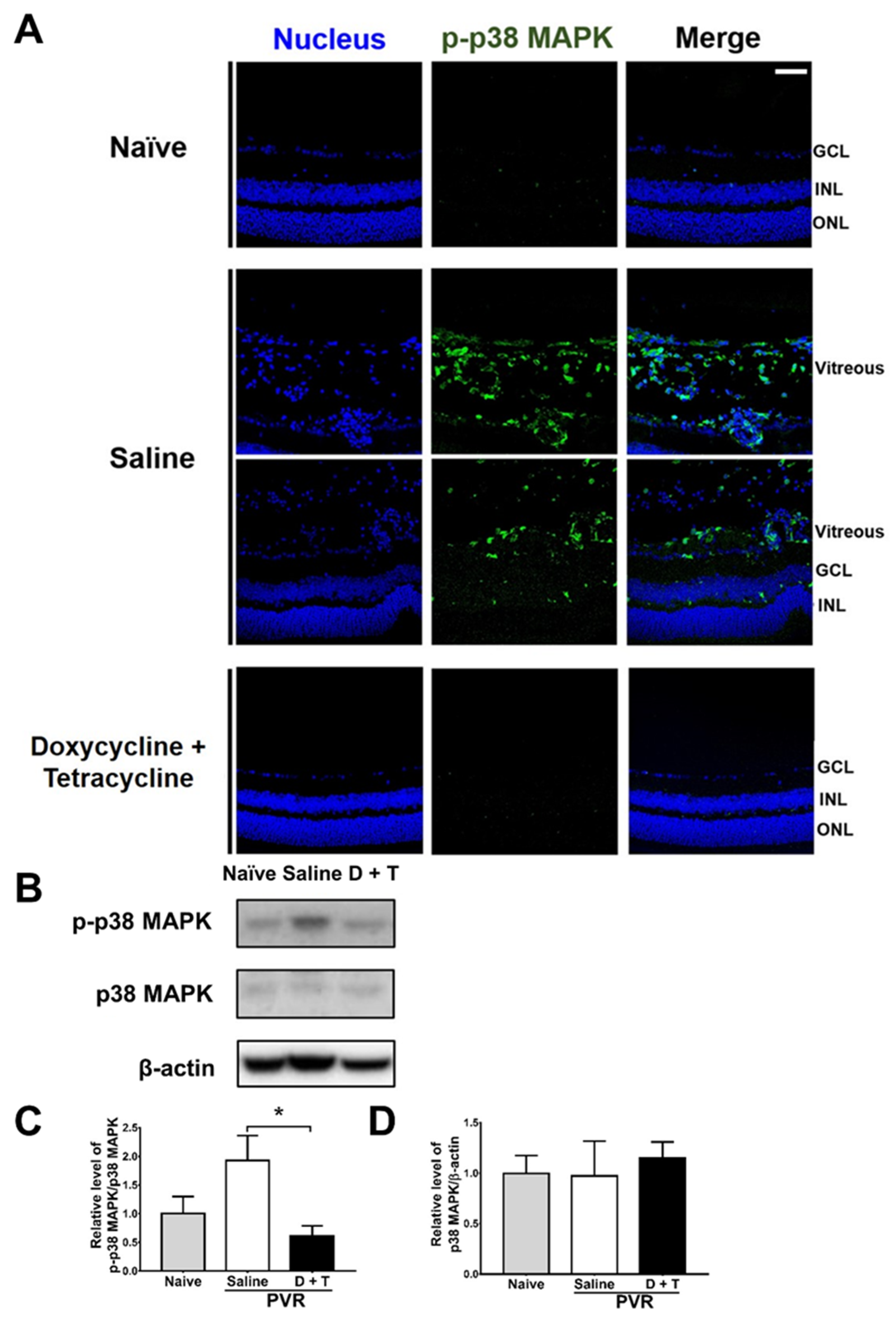
2.4. Doxycycline and Tetracycline Treatment Decreases MMP-9 Expression and p38 MAPK Phosphorylation in the Retina and Vitreous Body of PVR Mice
2.5. Doxycycline and Tetracycline Treatment Decreases IL-4 and CXCL9 but Increases IL-10 in the Retina of PVR Mice
3. Discussion
4. Materials and Methods
4.1. Cell and Mouse
4.2. Assays for Cell Migration, Proliferation, and Contraction
4.3. Assays for the Total MMP Activity
4.4. Western Blotting Analysis
4.5. PVR Induction and Antibiotic Treatment in Mice
4.6. Histological and Immunofluorescence Staining
4.7. Luminex Assay
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Kwon, O.W.; Song, J.H.; Roh, M.I. Retinal Detachment and Proliferative Vitreoretinopathy. Dev. Ophthalmol. 2016, 55, 154–162. [Google Scholar]
- Van de Put, M.A.J.; Hooymans, J.M.M.; Los, L.I. Dutch Rhegmatogenous Retinal Detachment Study Group. The incidence of rhegmatogenous retinal detachment in The Netherlands. Ophthalmology 2013, 120, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Mitry, D.; Charteris, D.G.; Yorston, D.; Siddiqui, M.A.; Campbell, H.; Murphy, A.L.; Fleck, B.W.; Wright, A.F.; Singh, J.; Scottish, R.D.S.G. The epidemiology and socioeconomic associations of retinal detachment in Scotland: A two-year prospective population-based study. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4963–4968. [Google Scholar] [CrossRef]
- Park, S.J.; Choi, N.K.; Park, K.H.; Woo, S.J. Five year nationwide incidence of rhegmatogenous retinal detachment requiring surgery in Korea. PLoS ONE 2013, 8, e80174. [Google Scholar] [CrossRef]
- Kuo, H.K.; Chen, Y.H.; Wu, P.C.; Kuo, Y.H. The Effects of Ozurdex(R) (Dexamethasone Intravitreal Implant) on Experimental Proliferative Vitreoretinopathy. Ophthalmologica 2015, 233, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Heimann, H.; Bartz-Schmidt, K.U.; Bornfeld, N.; Weiss, C.; Hilgers, R.D.; Foerster, M.H. Scleral Buckling versus Primary Vitrectomy in Rhegmatogenous Retinal Detachment Study Group. Scleral buckling versus primary vitrectomy in rhegmatogenous retinal detachment: A prospective randomized multicenter clinical study. Ophthalmology 2007, 114, 2142–2154. [Google Scholar] [CrossRef]
- Falkner-Radler, C.I.; Myung, J.S.; Moussa, S.; Chan, R.V.; Smretschnig, E.; Kiss, S.; Graf, A.; D’Amico, D.J.; Binder, S. Trends in primary retinal detachment surgery: Results of a Bicenter study. Retina 2011, 31, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Brady, C.J.; Kaiser, R.S. Clinical management of proliferative vitreoretinopathy: An update. Retina 2015, 35, 165–175. [Google Scholar] [CrossRef]
- Claes, C.; Lafeta, A.P. Proliferative vitreoretinopathy. Dev. Ophthalmol. 2014, 54, 188–195. [Google Scholar]
- Pastor, J.C.; Rojas, J.; Pastor-Idoate, S.; Di Lauro, S.; Gonzalez-Buendia, L.; Delgado-Tirado, S. Proliferative vitreoretinopathy: A new concept of disease pathogenesis and practical consequences. Prog. Retin. Eye Res. 2016, 51, 125–155. [Google Scholar] [CrossRef]
- Asaria, R.H.; Kon, C.H.; Bunce, C.; Charteris, D.G.; Wong, D.; Khaw, P.T.; Aylward, G.W. Adjuvant 5-fluorouracil and heparin prevents proliferative vitreoretinopathy: Results from a randomized, double-blind, controlled clinical trial. Ophthalmology 2001, 108, 1179–1183. [Google Scholar] [CrossRef]
- Wickham, L.; Bunce, C.; Wong, D.; McGurn, D.; Charteris, D.G. Randomized controlled trial of combined 5-Fluorouracil and low-molecular-weight heparin in the management of unselected rhegmatogenous retinal detachments undergoing primary vitrectomy. Ophthalmology 2007, 114, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.A.; Sanchez, J.G.; Arevalo, J.F. Combined 5-fluorouracil, low-molecular-weight heparin, and silicone oil in the management of complicated retinal detachment with proliferative vitreoretinopathy grade C. Ophthalmic. Surg. Lasers Imaging 2007, 38, 276–282. [Google Scholar] [CrossRef]
- Wiedemann, P.; Hilgers, R.D.; Bauer, P.; Heimann, K. Adjunctive daunorubicin in the treatment of proliferative vitreoretinopathy: Results of a multicenter clinical trial. Daunomycin Study Group. Am. J. Ophthalmol. 1998, 126, 550–559. [Google Scholar] [CrossRef]
- Shinohara, K.; Tanaka, M.; Sakuma, T.; Kobayashi, Y. Efficacy of daunorubicin encapsulated in liposome for the treatment of proliferative vitreoretinopathy. Ophthalmic. Surg. Lasers Imaging 2003, 34, 299–305. [Google Scholar] [CrossRef]
- Chang, Y.C.; Hu, D.N.; Wu, W.C. Effect of oral 13-cis-retinoic acid treatment on postoperative clinical outcome of eyes with proliferative vitreoretinopathy. Am. J. Ophthalmol. 2008, 146, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Berman, D.H.; Gombos, G.M. Proliferative vitreoretinopathy: Does oral low-dose colchicine have an inhibitory effect? A controlled study in humans. Ophthalmic. Surg. 1989, 20, 268–272. [Google Scholar]
- El Baha, S.; Leila, M.; Amr, A.; Lolah, M.M.A. Anatomical and Functional Outcomes of Vitrectomy with/without Intravitreal Methotrexate Infusion for Management of Proliferative Vitreoretinopathy Secondary to Rhegmatogenous Retinal Detachment. J. Ophthalmol. 2021, 2021, 3648134. [Google Scholar] [CrossRef]
- Benner, J.D.; Dao, D.; Butler, J.W.; Hamill, K.I. Intravitreal methotrexate for the treatment of proliferative vitreoretinopathy. BMJ Open Ophthalmol. 2019, 4, e000293. [Google Scholar] [CrossRef]
- Sadaka, A.; Sisk, R.A.; Osher, J.M.; Toygar, O.; Duncan, M.K.; Riemann, C.D. Intravitreal methotrexate infusion for proliferative vitreoretinopathy. Clin. Ophthalmol. 2016, 10, 1811–1817. [Google Scholar] [CrossRef] [PubMed]
- Jahangir, S.; Jahangir, T.; Ali, M.H.; Lateef, Q.; Hamza, U.; Tayyab, H. Use of Intravitreal Methotrexate Infusion in Complicated Retinal Detachment for Prevention of Proliferative Vitreoretinopathy in a Pilot Study. Cureus 2021, 13, e17439. [Google Scholar] [PubMed]
- Liang, C.M.; Tai, M.C.; Chang, Y.H.; Chen, Y.H.; Chen, C.L.; Lu, D.W.; Chen, J.T. Glucosamine inhibits epithelial-to-mesenchymal transition and migration of retinal pigment epithelium cells in culture and morphologic changes in a mouse model of proliferative vitreoretinopathy. Acta Ophthalmol. 2011, 89, e505–e514. [Google Scholar] [CrossRef]
- Wu, W.C.; Hu, D.N.; Mehta, S.; Chang, Y.C. Effects of retinoic acid on retinal pigment epithelium from excised membranes from proliferative vitreoretinopathy. J. Ocul. Pharmcol. Ther. 2005, 21, 44–54. [Google Scholar] [CrossRef]
- Rubsamen, P.E.; Cousins, S.W. Therapeutic effect of periocular corticosteroids in experimental proliferative vitreoretinopathy. Retina 1997, 17, 44–50. [Google Scholar] [CrossRef]
- Limb, G.A.; Little, B.C.; Meager, A.; Ogilvie, J.A.; Wolstencroft, R.A.; Franks, W.A.; Chignell, A.H.; Dumonde, D.C. Cytokines in proliferative vitreoretinopathy. Eye 1991, 5 Pt 6, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Mitamura, Y.; Takeuchi, S.; Yamamoto, S.; Yamamoto, T.; Tsukahara, I.; Matsuda, A.; Tagawa, Y.; Mizue, Y.; Nishihira, J. Monocyte chemotactic protein-1 levels in the vitreous of patients with proliferative vitreoretinopathy. Jpn. J. Ophthalmol. 2002, 46, 218–221. [Google Scholar] [CrossRef]
- Abu El-Asrar, A.M.; Struyf, S.; Kangave, D.; Geboes, K.; Van Damme, J. Chemokines in proliferative diabetic retinopathy and proliferative vitreoretinopathy. Eur. Cytokine Netw. 2006, 17, 155–165. [Google Scholar] [CrossRef]
- Zandi, S.; Pfister, I.B.; Traine, P.G.; Tappeiner, C.; Despont, A.; Rieben, R.; Skowronska, M.; Garweg, J.G. Biomarkers for PVR in rhegmatogenous retinal detachment. PLoS ONE 2019, 14, e0214674. [Google Scholar] [CrossRef] [PubMed]
- Symeonidis, C.; Papakonstantinou, E.; Androudi, S.; Rotsos, T.; Diza, E.; Brazitikos, P.; Karakiulakis, G.; Dimitrakos, S.A. Interleukin-6 and the matrix metalloproteinase response in the vitreous during proliferative vitreoretinopathy. Cytokine 2011, 54, 212–217. [Google Scholar] [CrossRef]
- Symeonidis, C.; Papakonstantinou, E.; Souliou, E.; Karakiulakis, G.; Dimitrakos, S.A.; Diza, E. Correlation of matrix metalloproteinase levels with the grade of proliferative vitreoretinopathy in the subretinal fluid and vitreous during rhegmatogenous retinal detachment. Acta Ophthalmol. 2011, 89, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Townley, W.A.; Cambrey, A.D.; Khaw, P.T.; Grobbelaar, A.O. Matrix metalloproteinase inhibition reduces contraction by dupuytren fibroblasts. J. Hand Surg. Am. 2008, 33, 1608–1616. [Google Scholar] [CrossRef]
- Smith, G.N., Jr.; Mickler, E.A.; Hasty, K.A.; Brandt, K.D. Specificity of inhibition of matrix metalloproteinase activity by doxycycline: Relationship to structure of the enzyme. Arthritis Rheum. 1999, 42, 1140–1146. [Google Scholar] [CrossRef]
- Onoda, T.; Ono, T.; Dhar, D.K.; Yamanoi, A.; Fujii, T.; Nagasue, N. Doxycycline inhibits cell proliferation and invasive potential: Combination therapy with cyclooxygenase-2 inhibitor in human colorectal cancer cells. J. Lab. Clin. Med. 2004, 143, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Son, H.J.; Kim, E.M.; Choi, J.H.; Kim, S.T.; Ji, I.J.; Choi, D.H.; Joh, T.H.; Kim, Y.S.; Hwang, O. Doxycycline is neuroprotective against nigral dopaminergic degeneration by a dual mechanism involving MMP-3. Neurotox. Res. 2009, 16, 361–371. [Google Scholar] [CrossRef]
- Sun, T.; Zhao, N.; Ni, C.S.; Zhao, X.L.; Zhang, W.Z.; Su, X.; Zhang, D.F.; Gu, Q.; Sun, B.C. Doxycycline inhibits the adhesion and migration of melanoma cells by inhibiting the expression and phosphorylation of focal adhesion kinase (FAK). Cancer Lett. 2009, 285, 141–150. [Google Scholar] [CrossRef]
- Castro, M.M.; Tanus-Santos, J.E.; Gerlach, R.F. Matrix metalloproteinases: Targets for doxycycline to prevent the vascular alterations of hypertension. Pharm. Res. 2011, 64, 567–572. [Google Scholar] [CrossRef]
- Li, H.; Ezra, D.G.; Burton, M.J.; Bailly, M. Doxycycline prevents matrix remodeling and contraction by trichiasis-derived conjunctival fibroblasts. Investig. Ophthalmol. Vis. Sci. 2013, 54, 4675–4682. [Google Scholar] [CrossRef]
- Bostanci, N.; Akgul, B.; Tsakanika, V.; Allaker, R.P.; Hughes, F.J.; McKay, I.J. Effects of low-dose doxycycline on cytokine secretion in human monocytes stimulated with Aggregatibacter actinomycetemcomitans. Cytokine 2011, 56, 656–661. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tang, X.; Wang, X.; Zhao, Y.Y.; Curtis, J.M.; Brindley, D.N. Doxycycline attenuates breast cancer related inflammation by decreasing plasma lysophosphatidate concentrations and inhibiting NF-kappaB activation. Mol. Cancer 2017, 16, 36. [Google Scholar] [CrossRef]
- Babaeinejad, S.; Khodaeiani, E.; Fouladi, R.F. Comparison of therapeutic effects of oral doxycycline and azithromycin in patients with moderate acne vulgaris: What is the role of age? J. Dermatol. Treat. 2011, 22, 206–210. [Google Scholar] [CrossRef]
- Scott, I.U.; Jackson, G.R.; Quillen, D.A.; Larsen, M.; Klein, R.; Liao, J.; Holfort, S.; Munch, I.C.; Gardner, T.W. Effect of doxycycline vs placebo on retinal function and diabetic retinopathy progression in patients with severe nonproliferative or non-high-risk proliferative diabetic retinopathy: A randomized clinical trial. JAMA Ophthalmol. 2014, 132, 535–543. [Google Scholar] [CrossRef]
- Massey, J.Y.; Hanna, C.; Goodart, R.; Wallace, T. Effect of drug vehicle on human ocular retention of topically applied tetracycline. Am. J. Ophthalmol. 1976, 81, 151–156. [Google Scholar] [CrossRef]
- Yang, S.; Li, H.; Li, M.; Wang, F. Mechanisms of epithelial-mesenchymal transition in proliferative vitreoretinopathy. Discov. Med. 2015, 20, 207–217. [Google Scholar] [PubMed]
- Kim, H.S.; Luo, L.; Pflugfelder, S.C.; Li, D.Q. Doxycycline inhibits TGF-beta1-induced MMP-9 via Smad and MAPK pathways in human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2005, 46, 840–848. [Google Scholar] [CrossRef]
- Zhang, J.; Yuan, G.; Dong, M.; Zhang, T.; Hua, G.; Zhou, Q.; Shi, W. Notch signaling modulates proliferative vitreoretinopathy via regulating retinal pigment epithelial-to-mesenchymal transition. Histochem. Cell Biol. 2017, 147, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liao, Y.; Sun, Q.; Tang, H.; Wang, G.; Zhao, F.; Jin, Y. Upregulation of Matrix Metalloproteinase-9 in Primary Cultured Rat Astrocytes Induced by 2-Chloroethanol Via MAPK Signal Pathways. Front Cell Neurosci. 2017, 11, 218. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal. Transduct. Target 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Mostafa Mtairag, E.; Chollet-Martin, S.; Oudghiri, M.; Laquay, N.; Jacob, M.P.; Michel, J.B.; Feldman, L.J. Effects of interleukin-10 on monocyte/endothelial cell adhesion and MMP-9/TIMP-1 secretion. Cardiovasc. Res. 2001, 49, 882–890. [Google Scholar] [CrossRef]
- Salminen, L. Penetration of ocular compartments by tetracyclines. I. An experimental study with tetracycline. Albrecht. Von Graefes Arch. Klin. Exp. Ophthalmol. 1977, 204, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Salminen, L. Penetration of ocular compartments by tetracyclines. II. An experimental study with doxycycline. Albrecht. Von Graefes Arch. Klin. Exp. Ophthalmol. 1977, 204, 201–207. [Google Scholar] [CrossRef]
- Gilmour, M.A.; Clarke, C.R.; Macallister, C.G.; Dedeo, J.M.; Caudell, D.L.; Morton, R.J.; Pugh, M. Ocular penetration of oral doxycycline in the horse. Vet. Ophthalmol. 2005, 8, 331–335. [Google Scholar] [CrossRef]
- Radisky, E.S.; Radisky, D.C. Matrix metalloproteinase-induced epithelial-mesenchymal transition in breast cancer. J. Mammary Gland Biol. Neoplasia 2010, 15, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Santa-Cecilia, F.V.; Socias, B.; Ouidja, M.O.; Sepulveda-Diaz, J.E.; Acuna, L.; Silva, R.L.; Michel, P.P.; Del-Bel, E.; Cunha, T.M.; Raisman-Vozari, R. Doxycycline Suppresses Microglial Activation by Inhibiting the p38 MAPK and NF-kB Signaling Pathways. Neurotox. Res. 2016, 29, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Charteris, D.G.; Hiscott, P.; Grierson, I.; Lightman, S.L. Proliferative vitreoretinopathy. Lymphocytes in epiretinal membranes. Ophthalmology 1992, 99, 1364–1367. [Google Scholar] [CrossRef]
- Roybal, C.N.; Velez, G.; Toral, M.A.; Tsang, S.H.; Bassuk, A.G.; Mahajan, V.B. Personalized Proteomics in Proliferative Vitreoretinopathy Implicate Hematopoietic Cell Recruitment and mTOR as a Therapeutic Target. Am. J. Ophthalmol. 2018, 186, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Morishima, Y.; Ishii, Y. Targeting Th2 cytokines in fibrotic diseases. Curr. Opin. Investig. Drugs 2010, 11, 1229–1238. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-H.; Lin, Y.-J.; Wang, L.-C.; Tsai, H.-Y.; Yang, C.-H.; Teng, Y.-T.; Hsu, S.-M. Doxycycline Ameliorates the Severity of Experimental Proliferative Vitreoretinopathy in Mice. Int. J. Mol. Sci. 2021, 22, 11670. https://doi.org/10.3390/ijms222111670
Chen S-H, Lin Y-J, Wang L-C, Tsai H-Y, Yang C-H, Teng Y-T, Hsu S-M. Doxycycline Ameliorates the Severity of Experimental Proliferative Vitreoretinopathy in Mice. International Journal of Molecular Sciences. 2021; 22(21):11670. https://doi.org/10.3390/ijms222111670
Chicago/Turabian StyleChen, Shun-Hua, Yu-Jheng Lin, Li-Chiu Wang, Hsien-Yang Tsai, Chang-Hao Yang, Yu-Ti Teng, and Sheng-Min Hsu. 2021. "Doxycycline Ameliorates the Severity of Experimental Proliferative Vitreoretinopathy in Mice" International Journal of Molecular Sciences 22, no. 21: 11670. https://doi.org/10.3390/ijms222111670
APA StyleChen, S.-H., Lin, Y.-J., Wang, L.-C., Tsai, H.-Y., Yang, C.-H., Teng, Y.-T., & Hsu, S.-M. (2021). Doxycycline Ameliorates the Severity of Experimental Proliferative Vitreoretinopathy in Mice. International Journal of Molecular Sciences, 22(21), 11670. https://doi.org/10.3390/ijms222111670