
An ATP-Binding Cassette Transporter, LaABCB11, Contributes to Alkaloid Transport in Lycoris aurea

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
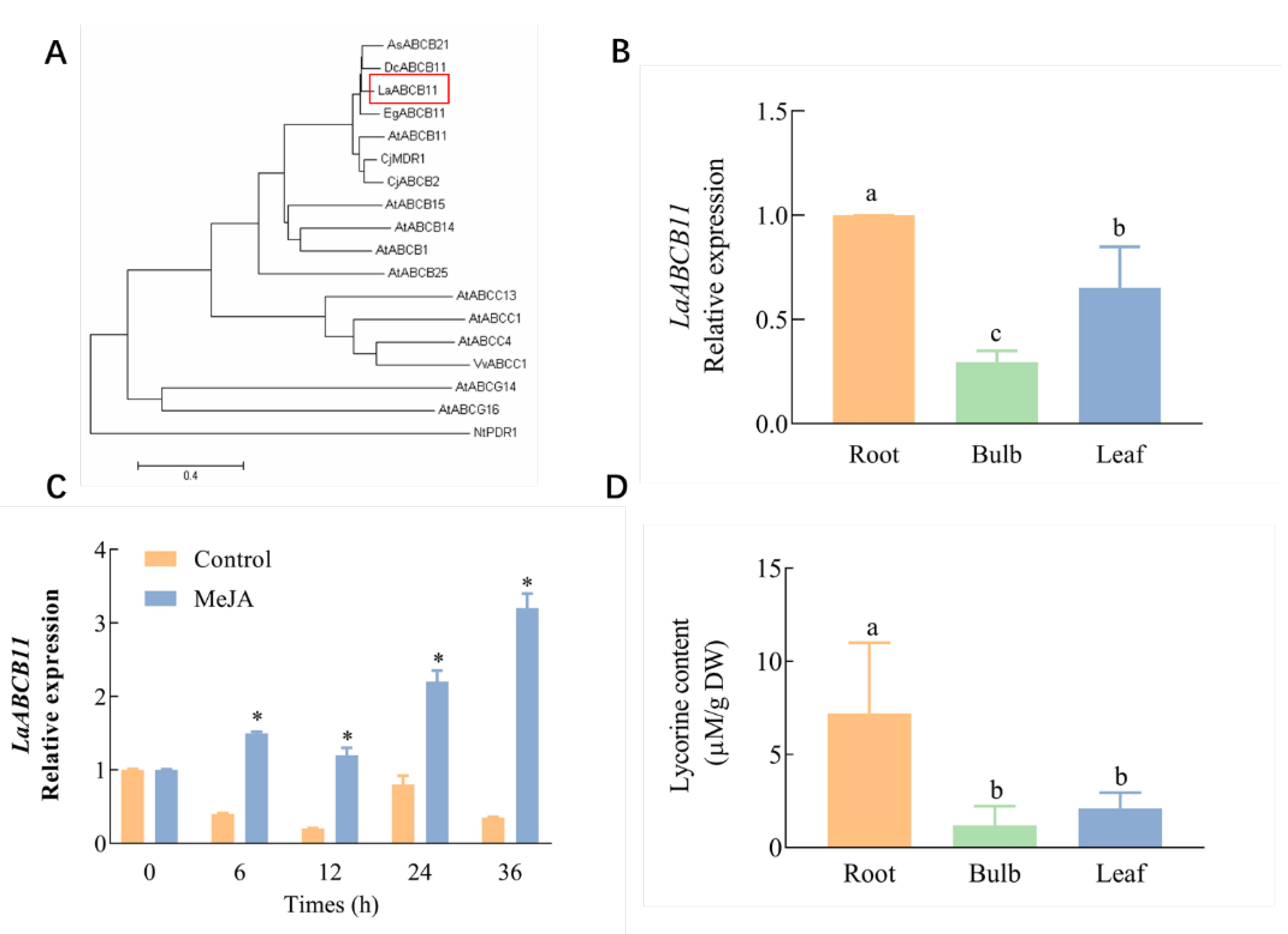
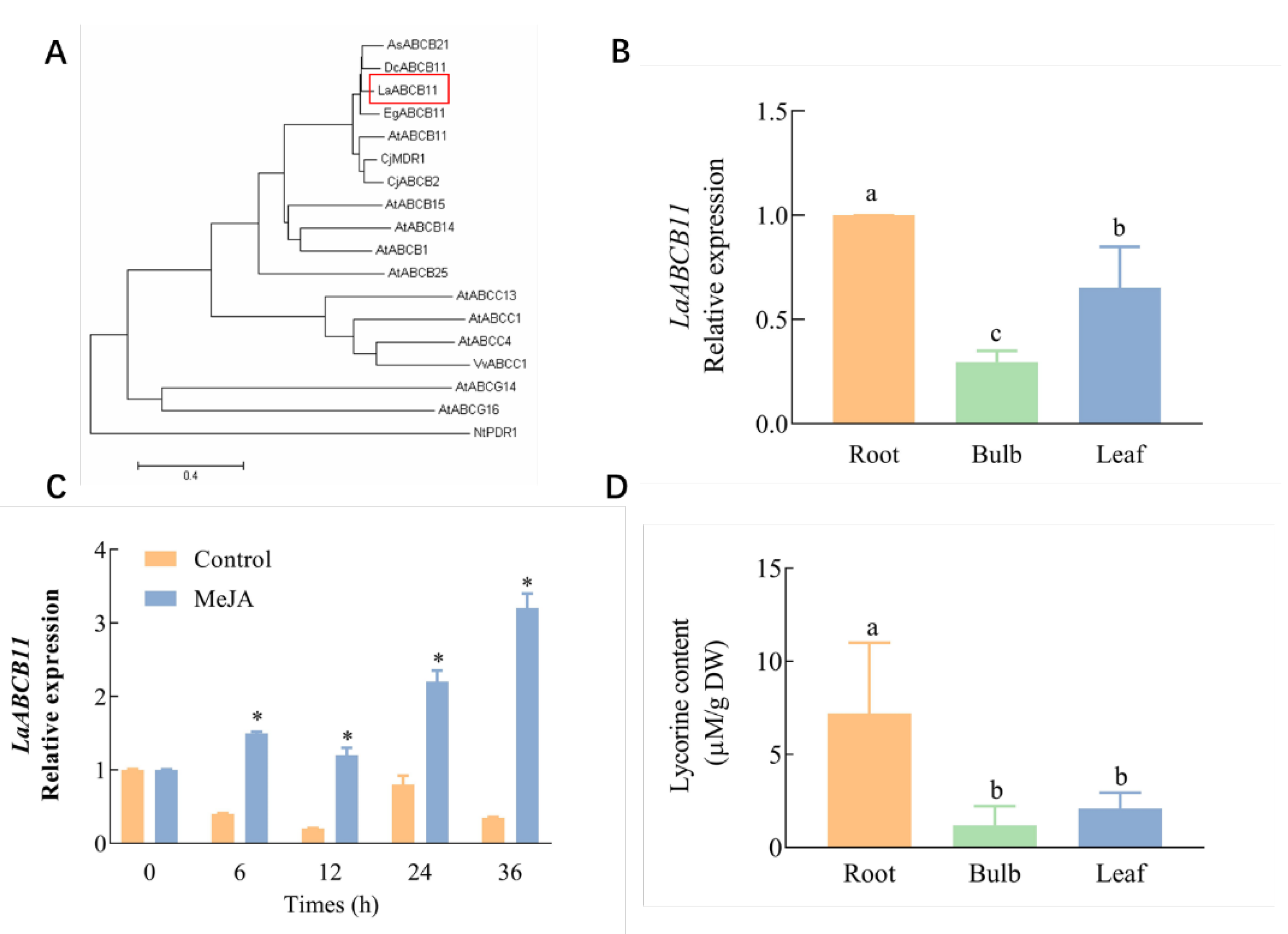
2.1. Isolation of Lycorine Transporter from L. aurea and Lycorine Content in Different Tissues
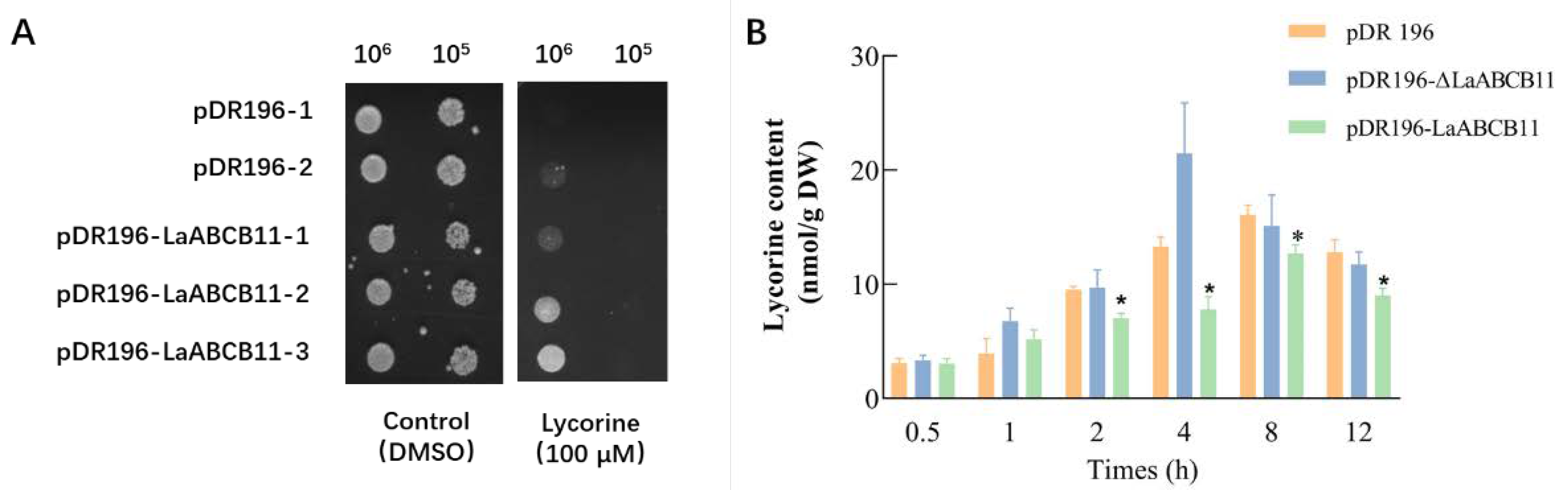
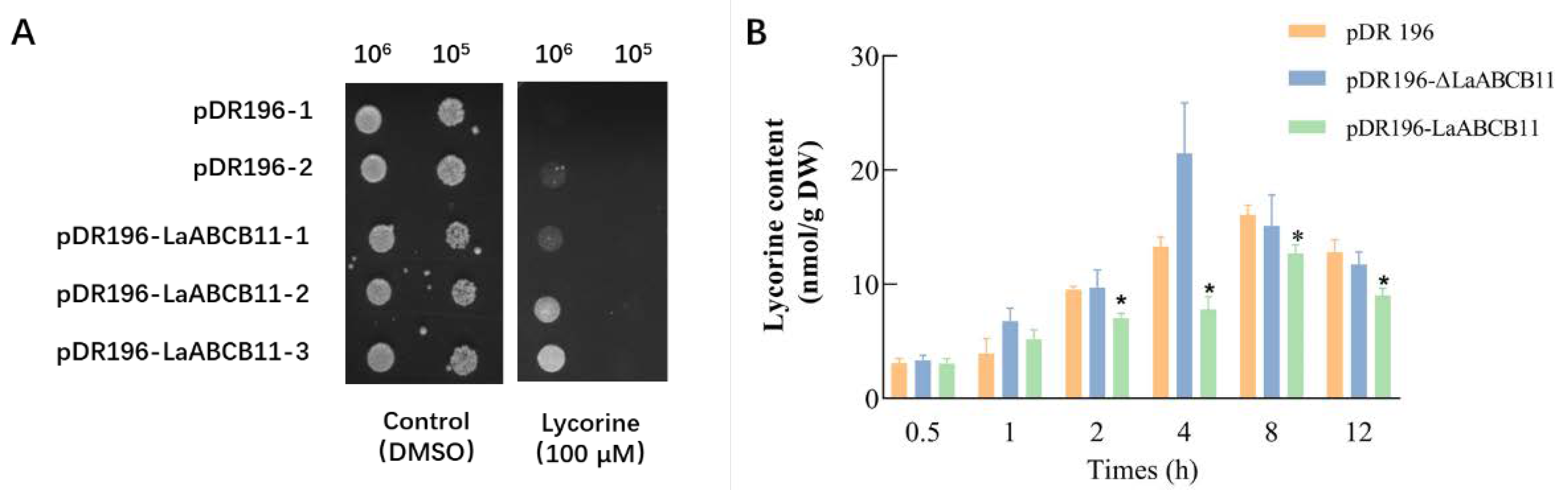
2.2. LaABCB11 Functions as a Lycorine Efflux in Yeast
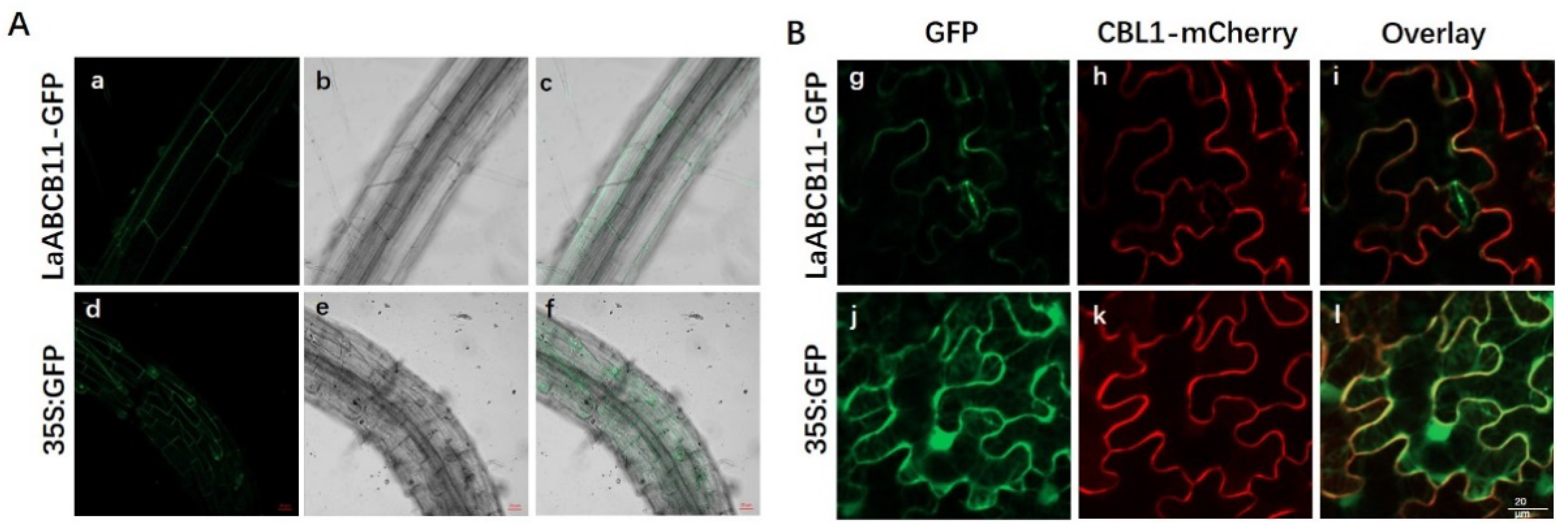
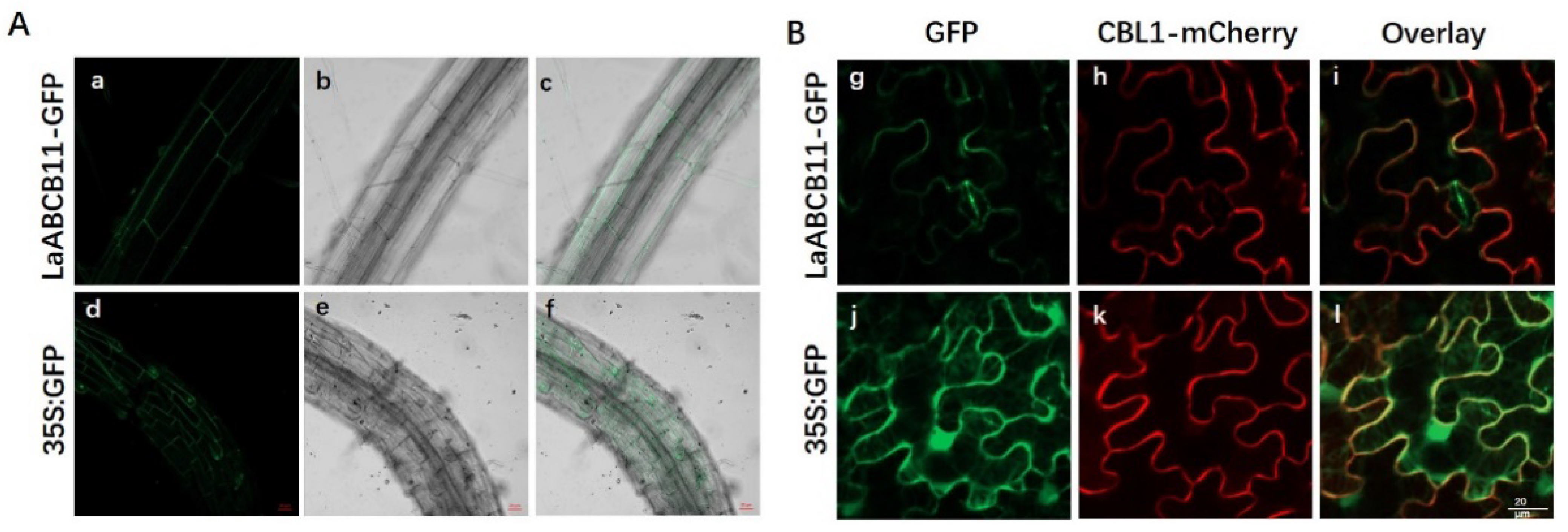
2.3. Sub-Cellular Localization of LaABCB11
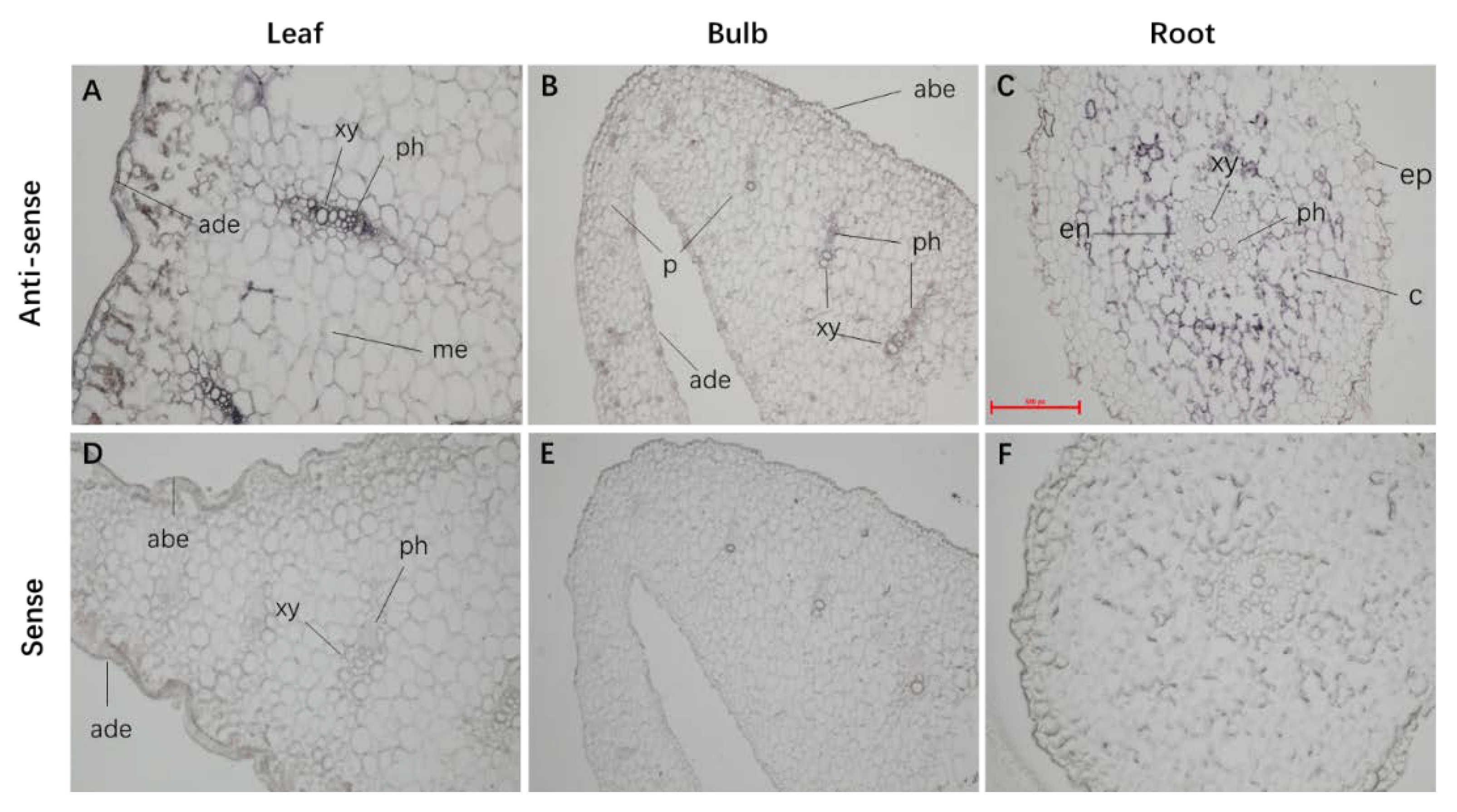
2.4. LaABCB11 Expression Pattern in Accordance with Its Function
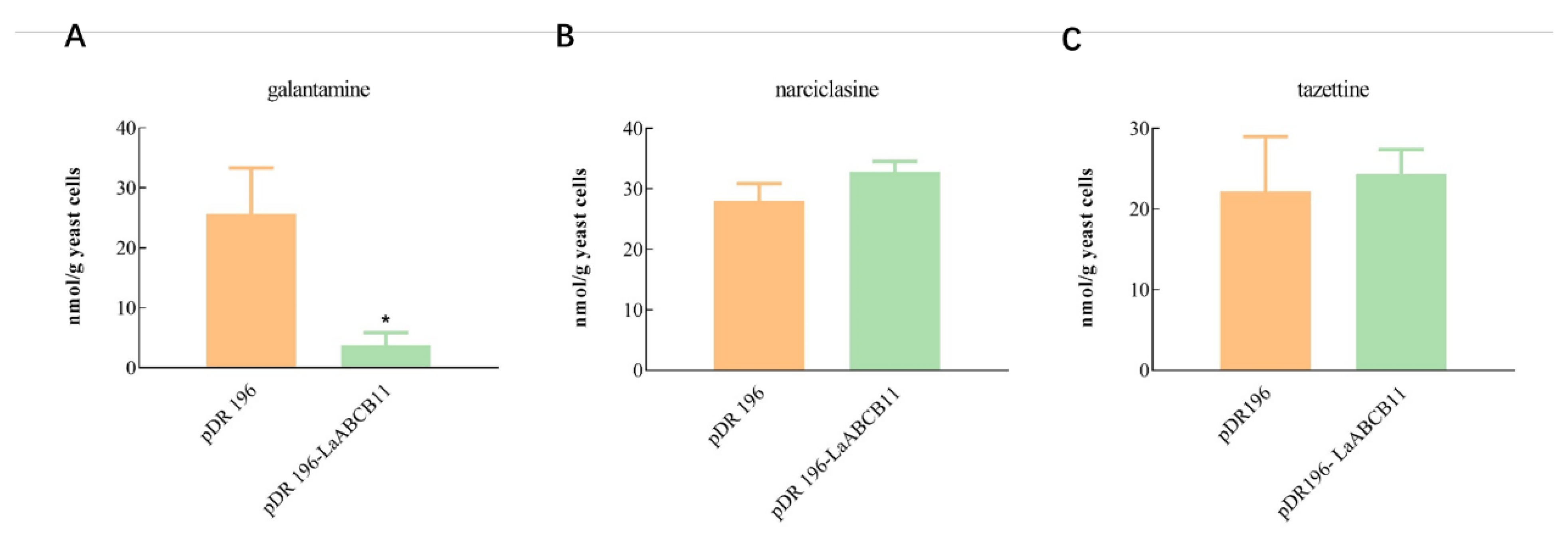
2.5. Substrate Specificity of LaABCB11
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Isolation of LaABCGB11 cDNA
4.3. RNA Isolation, cDNA Synthesis and qRT-PCR
4.4. Functional Analysis of LaABCB11 in Yeast
4.5. Transient Expression in N. benthamiana Leaves
4.6. Transformation of A. thaliana
4.7. In Situ Hybridization
4.8. HPLC Analysis
4.9. Phylogenetic Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lamoral-Theys, D.; Decaestecker, C.; Mathieu, V.; Dubois, J.; Kornienko, A.; Kiss, R.; Evidente, A.; Pottier, L. Lycorine and its derivatives for anticancer drug design. Mini Rev. Med. Chem. 2010, 10, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Boukhatem, M.N.; Setzer, W.N. Aromatic herbs, medicinal plant-derived essential oils, and phytochemical extracts as potential therapies for coronaviruses: Future perspectives. Plants 2020, 9, 800. [Google Scholar] [CrossRef]
- Heinrich, M.; Lee Teoh, H. Galanthamine from snowdrop--the development of a modern drug against Alzheimer’s disease from local Caucasian knowledge. J. Ethnopharmacol. 2004, 92, 147–162. [Google Scholar] [CrossRef]
- Kilgore, M.B.; Augustin, M.M.; Starks, C.M.; O’Neil-Johnson, M.; May, G.D.; Crow, J.A.; Kutchan, T.M. Cloning and characterization of a norbelladine 4′-O-methyltransferase involved in the biosynthesis of the Alzheimer’s drug galanthamine in Narcissus sp. aff. pseudonarcissus. PLoS ONE 2014, 9, e103223. [Google Scholar] [CrossRef] [Green Version]
- Kilgore, M.B.; Kutchan, T.M. The Amaryllidaceae alkaloids: Biosynthesis and methods for enzyme discovery. Phytochem. Rev. Proc. Phytochem. Soc. Eur. 2016, 15, 317–337. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Xu, S.; Wang, N.; Xia, B.; Jiang, Y.; Wang, R. Transcriptome analysis of secondary metabolism pathway, transcription factors, and transporters in response to methyl jasmonate in Lycoris aurea. Front. Plant Sci. 2016, 7, 1971. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Massicotte, M.A.; Garand, A.; Tousignant, L.; Ouellette, V.; Bérubé, G.; Desgagné-Penix, I. Cloning and characterization of norbelladine synthase catalyzing the first committed reaction in Amaryllidaceae alkaloid biosynthesis. BMC Plant Biol. 2018, 18, 338. [Google Scholar] [CrossRef]
- Barton, D.H.; Bhakuni, D.S.; Chapman, G.M.; Kirby, G.W.; Haynes, L.J.; Stuart, K.L. Phenol oxidation and biosynthesis. XIV. (Alkaloids from Croton species. VII.) The biosynthesis of crotonosine. J. Chem. Soc. Perkin Trans. 1 1967, 14, 1295–1298. [Google Scholar] [CrossRef]
- Kilgore, M.B.; Augustin, M.M.; May, G.D.; Crow, J.A.; Kutchan, T.M. CYP96T1 of Narcissus sp. aff. pseudonarcissus Catalyzes Formation of the Para-Para’ C-C Phenol Couple in the Amaryllidaceae Alkaloids. Front. Plant Sci. 2016, 7, 225. [Google Scholar] [CrossRef] [Green Version]
- Desgagné-Penix, I. Biosynthesis of alkaloids in Amaryllidaceae plants: A review. Phytochem. Rev. 2021, 20, 409–431. [Google Scholar] [CrossRef]
- Yu, F.; De Luca, V. ATP-binding cassette transporter controls leaf surface secretion of anticancer drug components in Catharanthus roseus. Proc. Natl. Acad. Sci. USA 2013, 110, 15830–15835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shitan, N. Secondary metabolites in plants: Transport and self-tolerance mechanisms. Biosci. Biotechnol. Biochem. 2016, 80, 1283–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, M.; Shitan, N.; Sawada, K.; Van Montagu, M.C.; Inzé, D.; Rischer, H.; Goossens, A.; Oksman-Caldentey, K.M.; Moriyama, Y.; Yazaki, K. Vacuolar transport of nicotine is mediated by a multidrug and toxic compound extrusion (MATE) transporter in Nicotiana tabacum. Proc. Natl. Acad. Sci. USA 2009, 106, 2447–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shitan, N.; Minami, S.; Morita, M.; Hayashida, M.; Ito, S.; Takanashi, K.; Omote, H.; Moriyama, Y.; Sugiyama, A.; Goossens, A.; et al. Involvement of the leaf-specific multidrug and toxic compound extrusion (MATE) transporter Nt-JAT2 in vacuolar sequestration of nicotine in Nicotiana tabacum. PLoS ONE 2014, 9, e108789. [Google Scholar] [CrossRef] [Green Version]
- Jelesko, J.G. An expanding role for purine uptake permease-like transporters in plant secondary metabolism. Front. Plant. Sci. 2012, 3, 78. [Google Scholar] [CrossRef] [Green Version]
- Shitan, N.; Bazin, I.; Dan, K.; Obata, K.; Kigawa, K.; Ueda, K.; Sato, F.; Forestier, C.; Yazaki, K. Involvement of CjMDR1, a plant multidrug-resistance-type ATP-binding cassette protein, in alkaloid transport in Coptis japonica. Proc. Natl. Acad. Sci. USA 2003, 100, 751–756. [Google Scholar] [CrossRef] [Green Version]
- Takanashi, K.; Yamada, Y.; Sasaki, T.; Yamamoto, Y.; Sato, F.; Yazaki, K. A multidrug and toxic compound extrusion transporter mediates berberine accumulation into vacuoles in Coptis japonica. Phytochemistry 2017, 138, 76–82. [Google Scholar] [CrossRef]
- Srinivasan, P.; Smolke, C.D. Engineering cellular metabolite transport for biosynthesis of computationally predicted tropane alkaloid derivatives in yeast. Proc. Natl. Acad. Sci. USA 2021, 118, e2104460118. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, F.; Boutry, M. Towards Identification of the Substrates of ATP-Binding Cassette Transporters. Plant Physiol. 2018, 178, 18–39. [Google Scholar] [CrossRef] [Green Version]
- Decottignies, A.; Grant, A.M.; Nichols, J.W.; de Wet, H.; McIntosh, D.B.; Goffeau, A. ATPase and multidrug transport activities of the overexpressed yeast ABC protein Yor1p. J. Biol. Chem. 1998, 273, 12612–12622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, R.J.; Locher, K.P. Structure of a bacterial multidrug ABC transporter. Nature 2006, 443, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Held, K.; Pascaud, F.; Eckert, C.; Gajdanowicz, P.; Hashimoto, K.; Corratgé-Faillie, C.; Offenborn, J.N.; Lacombe, B.; Dreyer, I.; Thibaud, J.B.; et al. Calcium-dependent modulation and plasma membrane targeting of the AKT2 potassium channel by the CBL4/CIPK6 calcium sensor/protein kinase complex. Cell Res. 2011, 21, 1116–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, T.; Brodsky, D.; Costa, A.; Kaneko, T.; Lo Schiavo, F.; Katsuhara, M.; Schroeder, J.J.P. K+ transport by the OsHKT2; 4 transporter from rice with atypical Na+ transport properties and competition in permeation of K+ over Mg2+ and Ca2+ ions. 2011, 156, 1493–1507. Plant Physiol. 2011, 156, 1493–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caputi, L.; Franke, J.; Farrow, S.C.; Chung, K.; Payne, R.M.E.; Nguyen, T.D.; Dang, T.T.; Soares Teto Carqueijeiro, I.; Koudounas, K.; Dugé de Bernonville, T.; et al. Missing enzymes in the biosynthesis of the anticancer drug vinblastine in Madagascar periwinkle. Science 2018, 360, 1235–1239. [Google Scholar] [CrossRef] [Green Version]
- Miao, G.P.; Han, J.; Zhang, J.F.; Zhu, C.S.; Zhang, X. A MDR transporter contributes to the different extracellular production of sesquiterpene pyridine alkaloids between adventitious root and hairy root liquid cultures of Tripterygium wilfordii Hook.f. Plant Mol. Biol. 2017, 95, 51–62. [Google Scholar] [CrossRef]
- Sun, B.; Wang, P.; Wang, R.; Li, Y.; Xu, S. Molecular Cloning and Characterization of a meta/para-O-Methyltransferase from Lycoris aurea. Int. J. Mol. Sci. 2018, 1911. [Google Scholar] [CrossRef] [Green Version]
- Demurtas, O.C.; de Brito Francisco, R.; Diretto, G.; Ferrante, P.; Frusciante, S.; Pietrella, M.; Aprea, G.; Borghi, L.; Feeney, M.; Frigerio, L.; et al. ABCC Transporters Mediate the Vacuolar Accumulation of Crocins in Saffron Stigmas. Plant cell 2019, 31, 2789–2804. [Google Scholar] [CrossRef] [Green Version]
- Mu, H.M.; Wang, R.; Li, X.D.; Jiang, Y.M.; Wang, C.Y.; Quan, J.P.; Peng, F.; Xia, B. Effect of abiotic and biotic elicitors on growth and alkaloid accumulation of Lycoris chinensis seedlings. Z. Naturforsch. C J. Biosci. 2009, 64, 541–550. [Google Scholar] [CrossRef]
- Kaneda, M.; Schuetz, M.; Lin, B.S.; Chanis, C.; Hamberger, B.; Western, T.L.; Ehlting, J.; Samuels, A.L. ABC transporters coordinately expressed during lignification of Arabidopsis stems include a set of ABCBs associated with auxin transport. J. Exp. Bot. 2011, 62, 2063–2077. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Wang, P.K.; Huang, M.R.; Zhou, J.; Han, Z.M. The localization of galanthamine in vegetative organ of Lycoris aurea Herb. J. Mol. Cell Biol. 2007, 40, 339–345. [Google Scholar] [CrossRef]
- Xu, S.; Jiang, Y.; Wang, N.; Xia, B.; Jiang, Y.; Li, X.; Zhang, Z.; Li, Y.; Wang, R. Identification and differential regulation of microRNAs in response to methyl jasmonate treatment in Lycoris aurea by deep sequencing. BMC Genom. 2016, 17, 789. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.; Xu, S.; Zhao, Y.; Xia, B.; Wang, R. Selection and validation of appropriate reference genes for quantitative Real-Time PCR analysis of gene expression in Lycoris aurea. Front. Plant Sci. 2016, 7, 536. [Google Scholar] [CrossRef] [Green Version]
- Martinoia, E.; Klein, M.; Geisler, M.; Bovet, L.; Forestier, C.; Kolukisaoglu, U.; Müller-Röber, B.; Schulz, B. Multifunctionality of plant ABC transporters--more than just detoxifiers. Planta 2002, 214, 345–355. [Google Scholar] [CrossRef]
- Kang, J.; Hwang, J.U.; Lee, M.; Kim, Y.Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Li, R.; Qi, M. In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. Plant J. Cell Mol. Biol. 2000, 22, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Bechtold, N.; Pelletier, G. In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol. Biol. 1998, 82, 259–266. [Google Scholar] [CrossRef]
- Jabnoune, M.; Espeout, S.; Mieulet, D.; Fizames, C.; Verdeil, J.L.; Conéjéro, G.; Rodríguez-Navarro, A.; Sentenac, H.; Guiderdoni, E.; Abdelly, C.; et al. Diversity in expression patterns and functional properties in the rice HKT transporter family. Plant Physiol. 2009, 150, 1955–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Liu, Z.; Wang, S.; Li, J.; Li, Y.; Chen, W.K.; Wang, R. Fungal endophytes promote the accumulation of Amaryllidaceae alkaloids in Lycoris radiata. Environ. Microbiol. 2020, 22, 1421–1434. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Liu, Y.; Xu, S.; Li, J.; Zhou, J.; Wang, R. An ATP-Binding Cassette Transporter, LaABCB11, Contributes to Alkaloid Transport in Lycoris aurea. Int. J. Mol. Sci. 2021, 22, 11458. https://doi.org/10.3390/ijms222111458
Wang R, Liu Y, Xu S, Li J, Zhou J, Wang R. An ATP-Binding Cassette Transporter, LaABCB11, Contributes to Alkaloid Transport in Lycoris aurea. International Journal of Molecular Sciences. 2021; 22(21):11458. https://doi.org/10.3390/ijms222111458
Chicago/Turabian StyleWang, Rong, Yantong Liu, Sheng Xu, Jie Li, Jiayu Zhou, and Ren Wang. 2021. "An ATP-Binding Cassette Transporter, LaABCB11, Contributes to Alkaloid Transport in Lycoris aurea" International Journal of Molecular Sciences 22, no. 21: 11458. https://doi.org/10.3390/ijms222111458
APA StyleWang, R., Liu, Y., Xu, S., Li, J., Zhou, J., & Wang, R. (2021). An ATP-Binding Cassette Transporter, LaABCB11, Contributes to Alkaloid Transport in Lycoris aurea. International Journal of Molecular Sciences, 22(21), 11458. https://doi.org/10.3390/ijms222111458