Platelet-Rich Plasma Promotes the Proliferation of Human Keratinocytes via a Progression of the Cell Cycle. A Role of Prolidase

 , , , and
, , , and 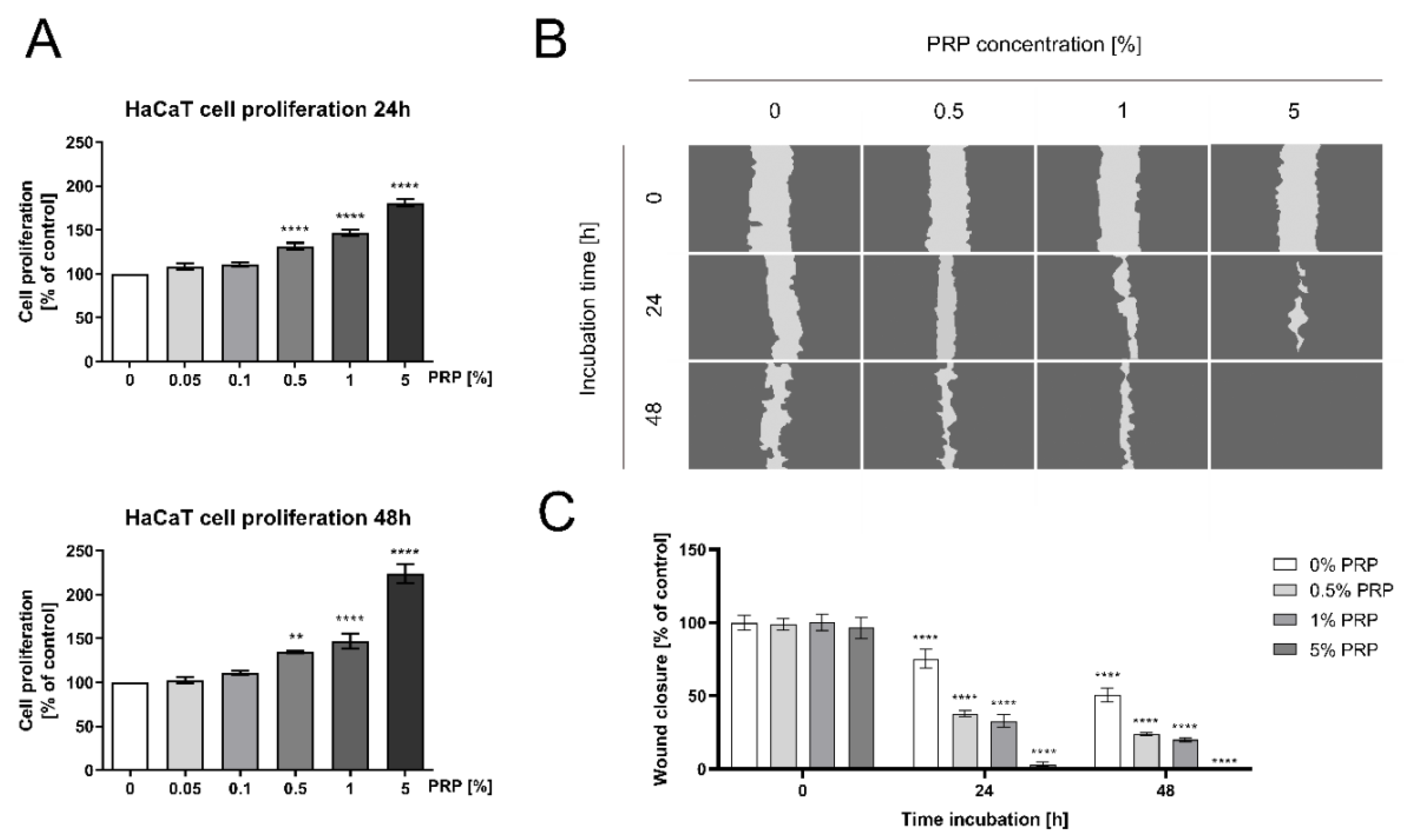{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. PRP Promotes DNA Biosynthesis and Cell Proliferation/Migration of HaCaT Keratinocytes
2.2. PRP Stimulates Cell Cycle Progression of HaCaT Keratinocytes
2.3. PRP Induces EGFR and EGFR-Downstream Signaling Proteins Expression in HaCaT Keratinocytes
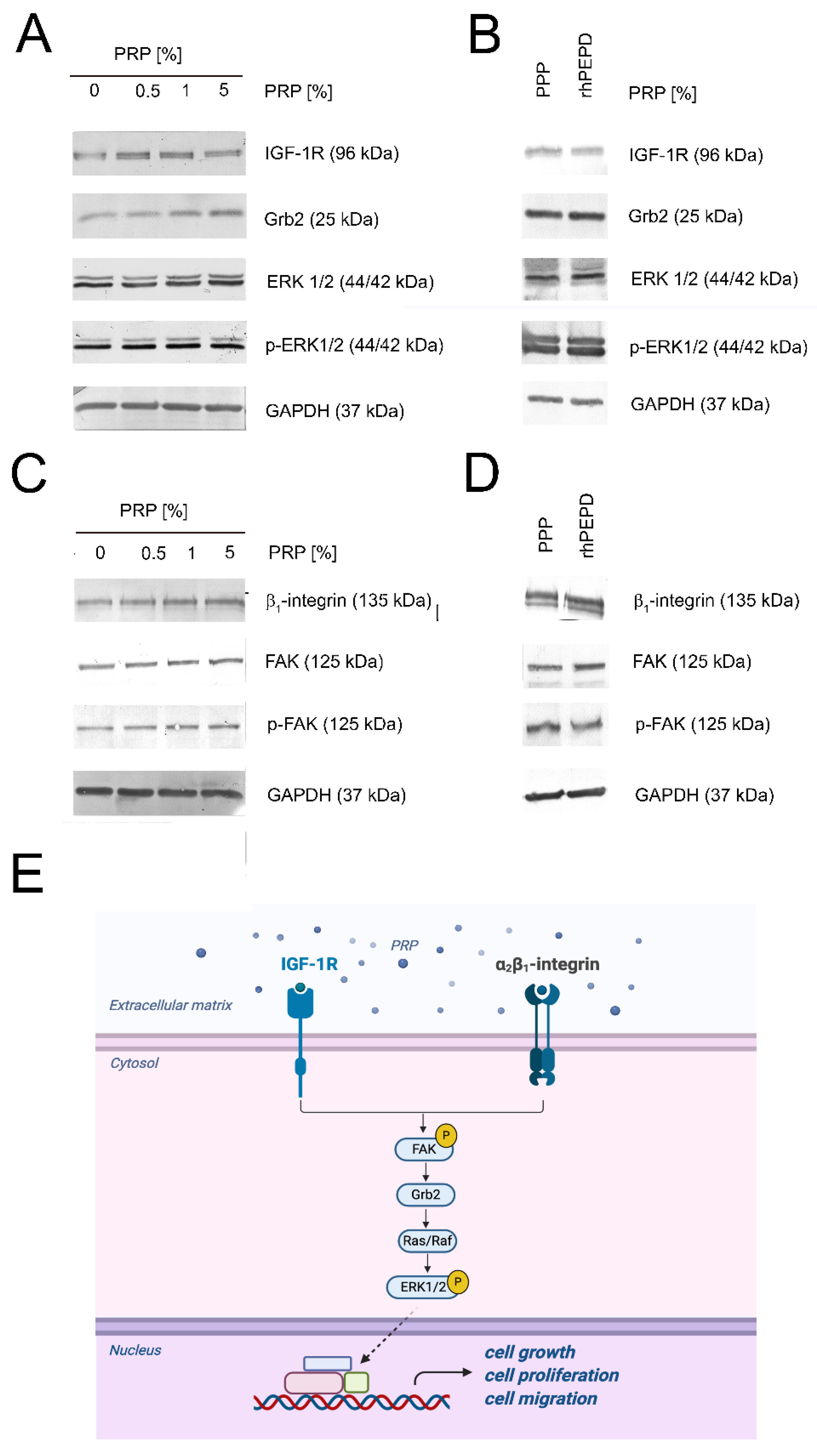
2.4. Activation of the β1-Integrin and IGF-1R Signaling in PRP-Treated HaCaT Cells
2.5. PRP Induces NF-κβ Expression in PRP-Treated HaCaT Cells
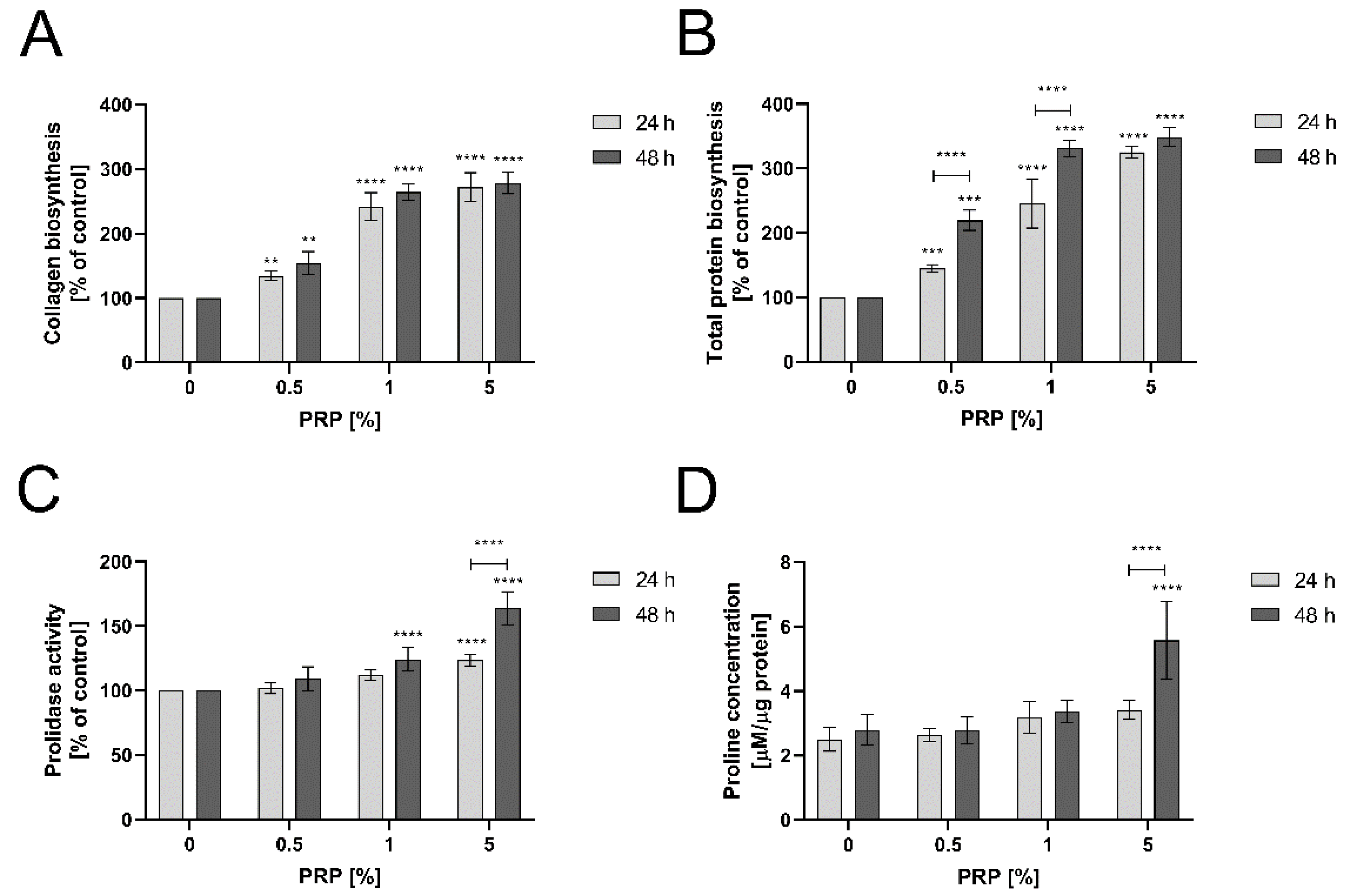
2.6. PRP Stimulates Collagen Biosynthesis through Enhancing Prolidase Activity and Proline Availability in An Experimental Model of Wound Healing in HaCaT Cells
3. Discussion
4. Materials and Methods
4.1. Platelet-Rich Plasma Production
4.2. HaCaT Cell Culture and Treatment
4.3. DNA Biosynthesis Assay
4.4. Wound Healing Assay
4.5. Cell Cycle Analysis
4.6. Preparation of Cell Lysates
4.7. Western Immunoblotting
4.8. Confocal Microscopy (Immunocytochemistry)
4.9. Prolidase Activity Measurement
4.10. Determination of Collagen Biosynthesis
4.11. Prolidase Concentration Measurement by ELISA
4.12. LC-MS Analysis of Proline Concentration
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Akt | protein kinase B |
| EGFR | epidermal growth factor receptor |
| ERK1/2 | extracellular signal-regulated kinase 1/2 |
| FAK | focal adhesion kinase pp125FAK |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| Glut-1 | glucose transporter-1 |
| Grb2 | growth factor receptor-bound protein 2 |
| HIF-1α | hypoxia-inducible factor 1 alpha |
| IGF-1R | insulin-like growth factor 1 receptor |
| mTOR | mammalian target of rapamycin |
| NF-κβ | nuclear factor kappa beta |
| IκBα | inhibitor of nuclear factor-kappa beta |
| IκKα | IκB kinase alpha |
| IκKβ | IκB kinase beta |
| PEPD | prolidase |
| PI3K | phosphoinositide 3 kinase |
| PRP | platelet-rich plasma |
| PPP | platelet-poor plasma |
| TGF-β1 | transforming growth factor-beta 1 |
| VEGF | vascular endothelial growth factor |
References
- Guszczyn, T.; Surażyński, A.; Zaręba, I.; Rysiak, E.; Popko, J.; Pałka, J. Differential effect of platelet-rich plasma fractions on β1-integrin signaling, collagen biosynthesis, and prolidase activity in human skin fibroblasts. Drug Des. Dev. Ther. 2017, 11, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Gou, L.; Zhang, P.; Li, H.; Qiu, S. Platelet-rich plasma and regenerative dentistry. Aust. Dent. J. 2020, 65, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Nurden, A.T. Platelets, inflammation and tissue regeneration. Thromb. Haemost. 2011, 105, S13–S33. [Google Scholar] [CrossRef] [PubMed]
- Nurden, A.T.; Nurden, P.; Sanchez, M.; Andia, I.; Anitua, E. Platelets and wound healing. Front. Biosci. 2008, 13, 3532–3548. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wu, Y.; Zhou, L.; Yang, Z.; Zhang, X.; Hu, X.; Yang, J.; Wang, M.; Wang, B.; Luo, G.; et al. Platelet-rich plasma accelerates skin wound healing by promoting re-epithelialization. Burn. Trauma 2020, 8, tkaa028. [Google Scholar] [CrossRef] [PubMed]
- Marx, R.E. Platelet-rich plasma: Evidence to support its use. J. Oral Maxillofac. Surg. 2004, 62, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Blair, P.; Flaumenhaft, R. Platelet α-granules: Basic biology and clinical correlates. Blood Rev. 2009, 23, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Garcovich, S. Systematic Review—The Potential Implications of Different Platelet-Rich Plasma (PRP) Concentrations in Regenerative Medicine for Tissue Repair. Int. J. Mol. Sci. 2020, 21, 5702. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cao, Y.; Zhao, R.; Xu, F.; Wu, D.; Wang, Y. The Role of Autologous PRP on Deep Partial-Thickness Burn Wound Healing in Bama Pigs. J. Burn. Care Res. 2020, 41, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Elsaid, A.; El-Said, M.; Emile, S.H.; Youssef, M.; Khafagy, W.; Elshobaky, A. Randomized Controlled Trial on Autologous Platelet-Rich Plasma Versus Saline Dressing in Treatment of Non-healing Diabetic Foot Ulcers. World J. Surg. 2019, 44, 1294–1301. [Google Scholar] [CrossRef]
- Evans, A.G.; Ibrahim, M.; Pope, R.; Mwangi, J.; Botros, M.; Johnson, S.P.; Al Kassis, S. Treating hand and foot osteoarthritis using a patient’s own blood: A systematic review and meta-analysis of platelet-rich plasma. J. Orthop. 2020, 18, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Volakakis, E.; Papadakis, M.; Manios, A.; Ioannou, C.V.; Zoras, O.; De Bree, E. Platelet-rich Plasma Improves Healing of Pressure Ulcers as Objectively Assessed by Digital Planimetry. Wounds 2019, 31, 252–256. [Google Scholar] [PubMed]
- Misiura, M.; Baszanowska, W.; Ościłowska, I.; Pałka, J.; Miltyk, W. Prolidase Stimulates Proliferation and Migration Through Activation of the PI3K/Akt/mTOR Signaling Pathway in Human Keratinocytes. Int. J. Mol. Sci. 2020, 21, 9243. [Google Scholar] [CrossRef] [PubMed]
- Karna, E.; Surazynski, A.; Orłowski, K.; Łaszkiewicz, J.; Puchalski, Z.; Nawrat, P.; Pałka, J. Serum and tissue level of insulin-like growth factor-I (IGF-I) and IGF-I binding proteins as an index of pancreatitis and pancreatic cancer. Int. J. Exp. Pathol. 2002, 83, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Surazynski, A.; Miltyk, W.; Prokop, I.; Palka, J. Prolidase-dependent regulation of TGF c and TGF β receptor expressions in human skin fibroblasts. Eur. J. Pharmacol. 2010, 649, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Misiura, M.; Miltyk, W. Current Understanding of the Emerging Role of Prolidase in Cellular Metabolism. Int. J. Mol. Sci. 2020, 21, 5906. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, Y.; Bhattacharya, A.; Zhang, Y. PEPD is a pivotal regulator of p53 tumor suppressor. Nat. Commun. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the Human Tissue-specific Expression by Genome-wide Integration of Transcriptomics and Antibody-based Proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef]
- Lupi, A.; Tenni, R.; Rossi, A.; Cetta, G.; Forlino, A. Human prolidase and prolidase deficiency: An overview on the characterization of the enzyme involved in proline recycling and on the effects of its mutations. Amino Acids 2008, 35, 739–752. [Google Scholar] [CrossRef]
- Gonzalez, A.C.D.O.; Andrade, Z.D.A.; Costa, T.F.; Medrado, A.R.A.P. Wound Healing—A Literature Review. An. Bras. Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef]
- Munch-Petersen, B. Enzymatic Regulation of Cytosolic Thymidine Kinase 1 and Mitochondrial Thymidine Kinase 2: A Mini Review. Nucl. Nucleotides Nucleic Acids 2010, 29, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Caillat, C.; Perrakis, A. Cdt1 and Geminin in DNA Replication Initiation. Alzheimer’s Dis. 2012, 62, 71–87. [Google Scholar] [CrossRef]
- Atherton-Fessler, S.; Liu, F.; Gabrielli, B.; Lee, M.S.; Peng, C.Y.; Piwnica-Worms, H. Cell cycle regulation of the p34cdc2 inhibitory kinases. Mol. Biol. Cell 1994, 5, 989–1001. [Google Scholar] [CrossRef] [PubMed]
- Castilho, R.M.; Squarize, C.H.; Gutkind, J.S. Exploiting PI3K/mTOR signaling to accelerate epithelial wound healing. Oral Dis. 2013, 19, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, Y.; Ding, Y.; Choi, K.-S.; Kazim, A.L.; Zhang, Y. Prolidase Directly Binds and Activates Epidermal Growth Factor Receptor and Stimulates Downstream Signaling. J. Biol. Chem. 2013, 288, 2365–2375. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J. Ubiquitin signalling in the NF-κB pathway. Nat. Cell Biol. 2005, 7, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Krane, S.M. The importance of proline residues in the structure, stability and susceptibility to proteolytic degradation of collagens. Amino Acids 2008, 35, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Namiduru, E.S. Prolidase. Bratisl. Lek. Listy 2016, 117, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, V.; Ciric, M.; Jovanovic, V.; Stojanovic, P. Platelet Rich Plasma: A short overview of certain bioactive components. Open Med. 2016, 11, 242–247. [Google Scholar] [CrossRef]
- Surazynski, A.; Miltyk, W.; Pałka, J.; Phang, J.M. Prolidase-dependent regulation of collagen biosynthesis. Amino Acids 2008, 35, 731–738. [Google Scholar] [CrossRef]
- Palka, J.A.; Phang, J.M. Prolidase activity in fibroblasts is regulated by interaction of extracellular matrix with cell surface integrin receptors. J. Cell. Biochem. 1997, 67, 166–175. [Google Scholar] [CrossRef]
- Juliano, R.L.; Haskill, S. Signal transduction from the extracellular matrix. J. Cell Biol. 1993, 120, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Seger, R.; Krebs, E.G. The MAPK signaling cascade. FASEB J. 1995, 9, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Labat-Robert, J.; Robert, L. Interaction Between Cells and Extracellular Matrix: Signaling by Integrins and the Elastin-Laminin Receptor. Prog. Mol. Subcell. Biol. 2000, 25, 57–70. [Google Scholar] [CrossRef]
- Kim, S.-A.; Ryu, H.-W.; Lee, K.-S.; Cho, J.-W. Application of platelet-rich plasma accelerates the wound healing process in acute and chronic ulcers through rapid migration and upregulation of cyclin A and CDK4 in HaCaT cells. Mol. Med. Rep. 2012, 7, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-L.; Tsai, Y.-C.; Korivi, M.; Chang, C.-T.; Hseu, Y.-C. Lucidone Promotes the Cutaneous Wound Healing Process via Activation of the PI 3 K/AKT, Wnt/β-catenin and NF-κB Signaling Pathways. Biochim. Biophys. Acta (BBA) Bioenerg. 2017, 1864, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, M.; Giladi, A.; Leibovich, S.J. Regulation of Vascular Endothelial Growth Factor Gene Expression in Murine Macrophages by Nitric Oxide and Hypoxia. Exp. Biol. Med. 2003, 228, 697–705. [Google Scholar] [CrossRef]
- Kiriakidis, S.; Andreakos, E.; Monaco, C.; Foxwell, B.; Feldmann, M.; Paleolog, E. VEGF expression in human macrophages is NF-kappaB-dependent: Studies using adenoviruses expressing the endogenous NF-kappaB inhibitor IkappaBalpha and a kinase-defective form of the IkappaB kinase. J. Cell Sci. 2003, 116, 665–674. [Google Scholar] [CrossRef]
- Yaron, A.; Naider, F.; Scharpe, S. Proline-Dependent Structural and Biological Properties of Peptides and Proteins. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 31–81. [Google Scholar] [CrossRef]
- Amable, P.R.; Carias, R.B.V.; Teixeira, M.V.T.; da Cruz Pacheco, Í.; Amaral, R.J.D.; Granjeiro, J.M.; Borojevic, R. Platelet-rich plasma preparation for regenerative medicine: Optimization and quantification of cytokines and growth factors. Stem Cell Res. Ther. 2013, 4, 67. [Google Scholar] [CrossRef]
- Marx, R.E. Platelet-Rich Plasma (PRP): What Is PRP and What Is Not PRP? Implant. Dent. 2001, 10, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Etulain, J. Platelets in wound healing and regenerative medicine. Platelets 2018, 29, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Emer, J. Platelet-Rich Plasma (PRP): Current Applications in Dermatology. Skin Ther. Lett. 2019, 24, 1–6. [Google Scholar]
- Besio, R.; Monzani, E.; Gioia, R.; Nicolis, S.; Rossi, A.; Casella, L.; Forlino, A. Improved prolidase activity assay allowed enzyme kinetic characterization and faster prolidase deficiency diagnosis. Clin. Chim. Acta 2011, 412, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Peterkofsky, B.; Diegelmann, R. Use of a mixture of proteinase-free collagenases for the specific assay of radioactive colla-gen in the presence of other proteins. Biochemistry 1971, 10, 988–994. [Google Scholar] [CrossRef]
- Klupczynska, A.; Misiura, M.; Miltyk, W.; Ościłowska, I.; Pałka, J.; Kokot, Z.J.; Matysiak, J. Development of an LC-MS Targeted Metabolomics Methodology to Study Proline Metabolism in Mammalian Cell Cultures. Molecules 2020, 25, 4639. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Misiura, M.; Guszczyn, T.; Oscilowska, I.; Baszanowska, W.; Palka, J.; Miltyk, W. Platelet-Rich Plasma Promotes the Proliferation of Human Keratinocytes via a Progression of the Cell Cycle. A Role of Prolidase. Int. J. Mol. Sci. 2021, 22, 936. https://doi.org/10.3390/ijms22020936
Misiura M, Guszczyn T, Oscilowska I, Baszanowska W, Palka J, Miltyk W. Platelet-Rich Plasma Promotes the Proliferation of Human Keratinocytes via a Progression of the Cell Cycle. A Role of Prolidase. International Journal of Molecular Sciences. 2021; 22(2):936. https://doi.org/10.3390/ijms22020936
Chicago/Turabian StyleMisiura, Magdalena, Tomasz Guszczyn, Ilona Oscilowska, Weronika Baszanowska, Jerzy Palka, and Wojciech Miltyk. 2021. "Platelet-Rich Plasma Promotes the Proliferation of Human Keratinocytes via a Progression of the Cell Cycle. A Role of Prolidase" International Journal of Molecular Sciences 22, no. 2: 936. https://doi.org/10.3390/ijms22020936
APA StyleMisiura, M., Guszczyn, T., Oscilowska, I., Baszanowska, W., Palka, J., & Miltyk, W. (2021). Platelet-Rich Plasma Promotes the Proliferation of Human Keratinocytes via a Progression of the Cell Cycle. A Role of Prolidase. International Journal of Molecular Sciences, 22(2), 936. https://doi.org/10.3390/ijms22020936