Systematic Identification of Protein Targets of Sub5 Using Saccharomyces cerevisiae Proteome Microarrays


Abstract
1. Introduction
2. Results
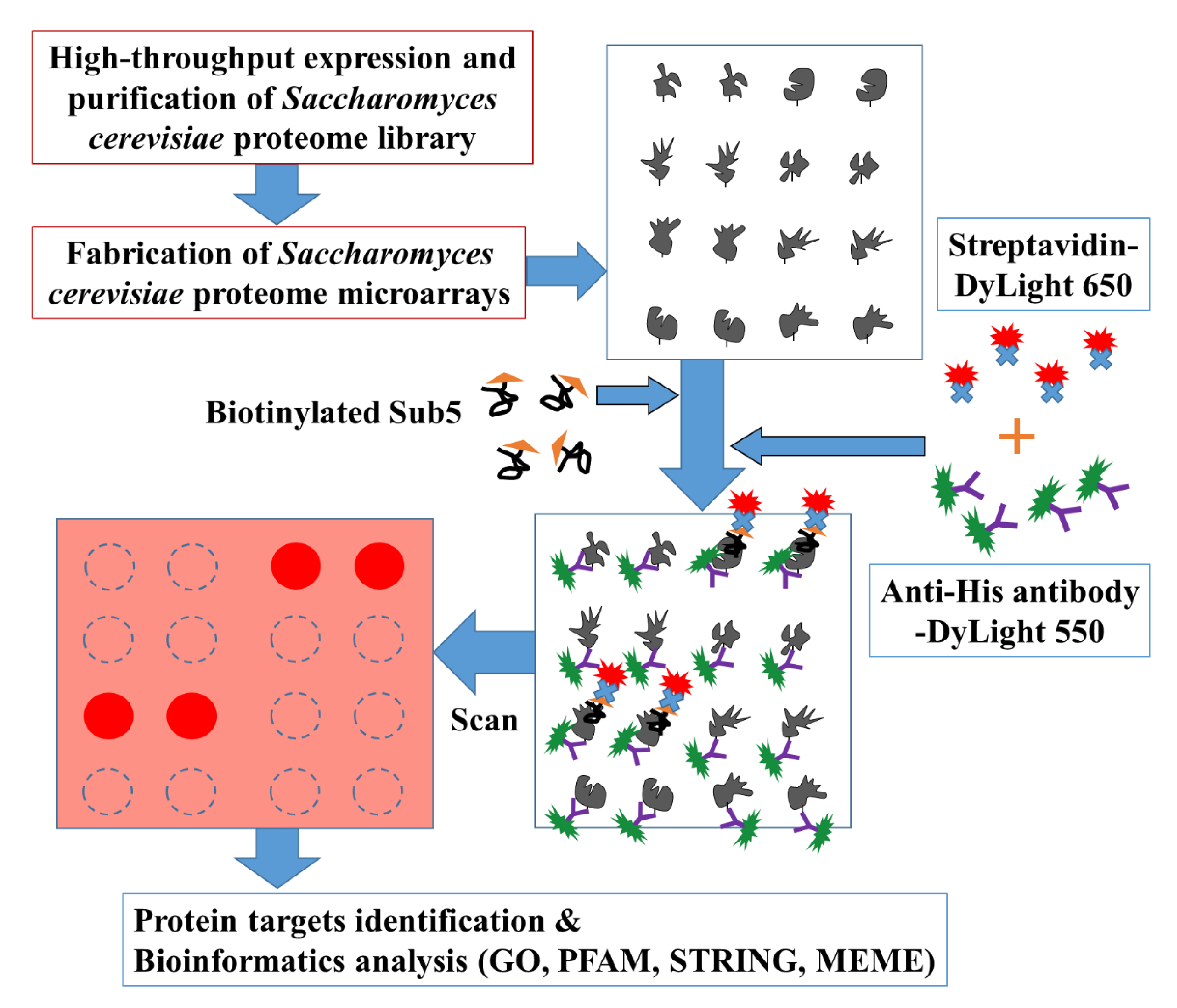
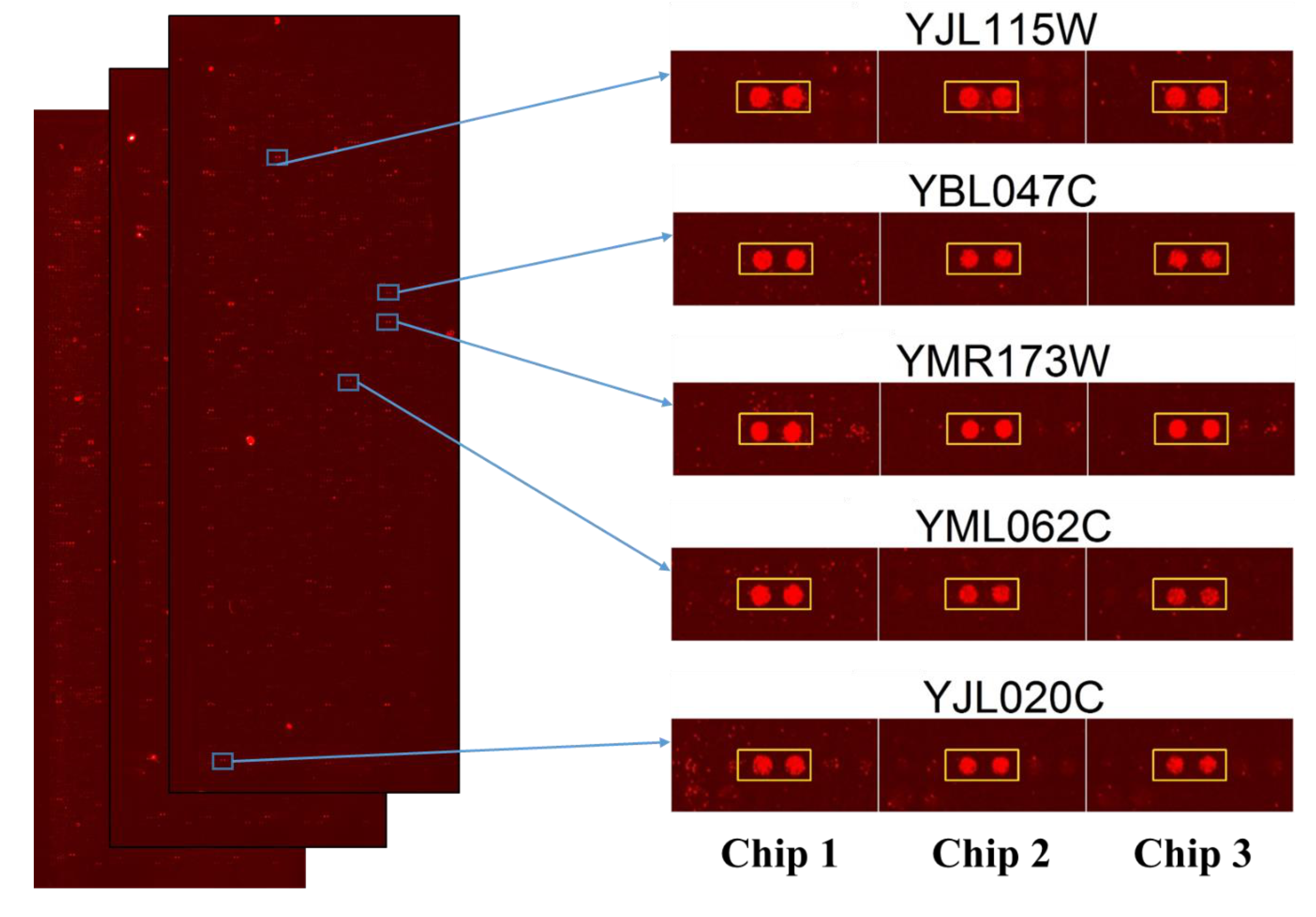
2.1. Saccharomyces Cerevisiae Proteome Microarrays Assays of Sub5
2.2. Statistical Analysis of Protein Targets of Sub5
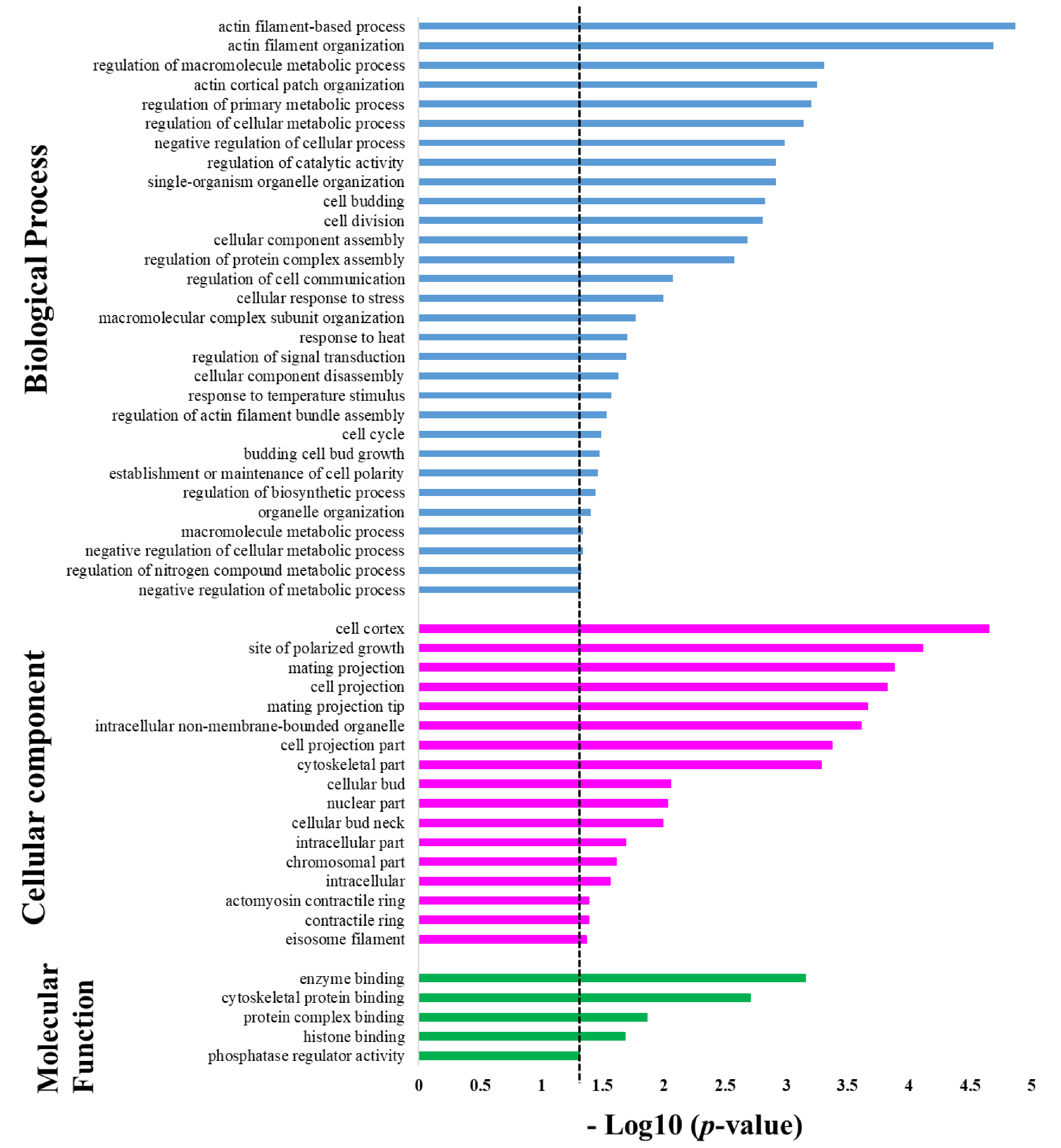
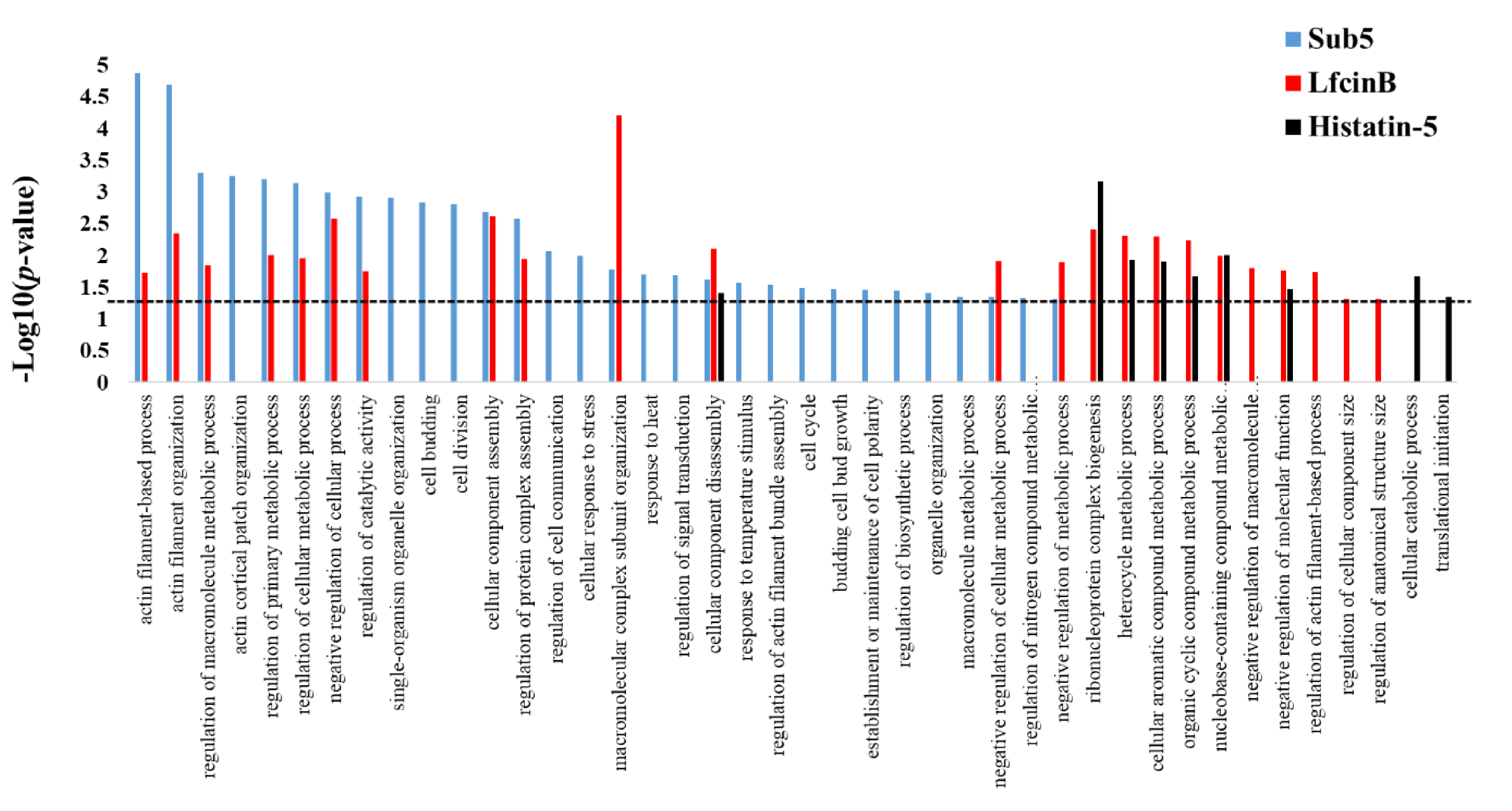
2.3. Bioinformatics Enrichment Analysis in Gene Ontology of the Protein Targets of Sub5
2.4. Protein Domain Enrichment among the Protein Targets of Sub5
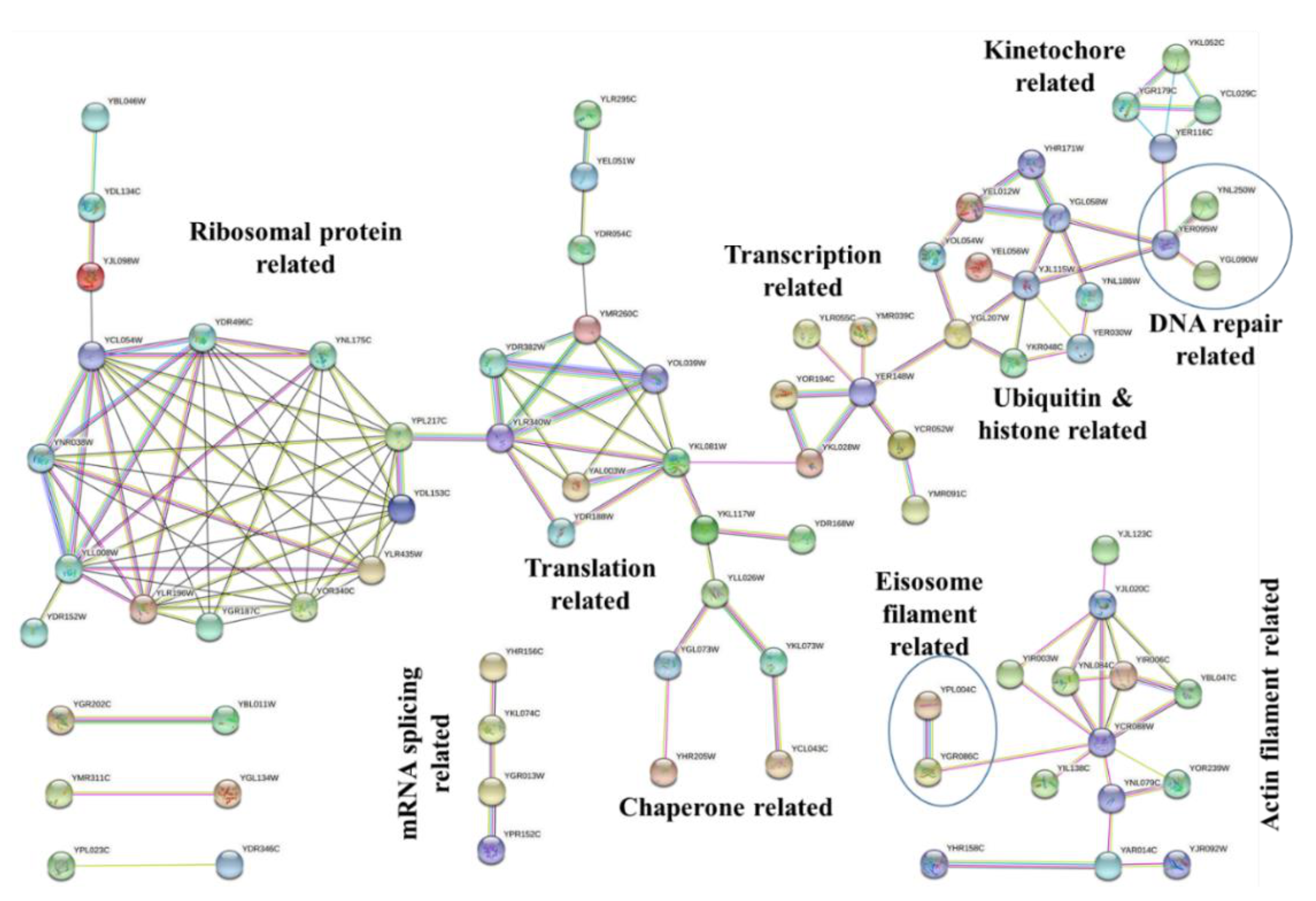
2.5. STRING Analysis for Interaction between of Protein Targets of Sub5
2.6. Consensus Motif Search among the Protein Targets of Sub5
3. Discussion
4. Materials and Methods
4.1. Expression and Purification of Entire Saccharomyces Cerevisiae Proteome
4.2. Fabrication of Saccharomyces Cerevisiae Proteome Microarrays
4.3. Saccharomyces Cerevisiae Proteome Microarray Assays with Sub5
4.4. Identification of Protein Targets of Sub5 from Saccharomyces Cerevisiae Proteome Microarrays
4.5. Bioinformatics Analysis of Gene Ontology
4.6. Bioinformatics Analysis for Protein–Protein Interaction
4.7. Motif Search by MEME
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shah:, P.; Hsiao, F.S.; Ho, Y.H.; Chen, C.S. The proteome targets of intracellular targeting antimicrobial peptides. Proteomics 2016, 16, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- McCormick, T.S.; Weinberg, A. Epithelial cell-derived antimicrobial peptides are multifunctional agents that bridge innate and adaptive immunity. Periodontol 2000 2010, 54, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Nissen-Meyer, J.; Nes, I.F. Ribosomally synthesized antimicrobial peptides: Their function, structure, biogenesis, and mechanism of action. Arch Microbiol. 1997, 167, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E. Peptide antibiotics. Lancet 1997, 349, 418–422. [Google Scholar] [CrossRef]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Biopolymers 2002, 66, 236–248. [Google Scholar] [CrossRef]
- Hilpert, K.; Volkmer-Engert, R.; Walter, T.; Hancock, R.E. High-throughput generation of small antibacterial peptides with improved activity. Nat. Biotechnol. 2005, 23, 1008–1012. [Google Scholar] [CrossRef]
- Wu, M.; Hancock, R.E. Improved derivatives of bactenecin, a cyclic dodecameric antimicrobial cationic peptide. Antimicrob. Agents. Chemother. 1999, 43, 1274–1276. [Google Scholar] [CrossRef]
- Mania, D.; Hilpert, K.; Ruden, S.; Fischer, R.; Takeshita, N. Screening for antifungal peptides and their modes of action in Aspergillus nidulans. Appl. Environ. Microbiol. 2010, 76, 7102–7108. [Google Scholar] [CrossRef]
- Chen, C.S.; Zhu, H. Protein microarrays. Biotechniques 2006, 40, 423–425. [Google Scholar] [CrossRef]
- Sutandy, F.X.; Qian, J.; Chen, C.S.; Zhu, H. Overview of protein microarrays. Curr. Protoc. Protein Sci. 2013, Chapter 27. 27.1.1–27.1.16. [Google Scholar] [CrossRef]
- Shah, P.; Wu, W.S.; Chen, C.S. Systematical Analysis of the Protein Targets of Lactoferricin B and Histatin-5 Using Yeast Proteome Microarrays. Int. J. Mol. Sci. 2019, 20, 4218. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.H.; Ho, Y.H.; Chuang, Y.C.; Chen, P.C.; Chen, C.S. Identification of lactoferricin B intracellular targets using an Escherichia coli proteome chip. PLoS ONE 2011, 6, e28197. [Google Scholar] [CrossRef]
- Ho, Y.H.; Sung, T.C.; Chen, C.S. Lactoferricin B inhibits the phosphorylation of the two-component system response regulators BasR and CreB. Mol. Cell. Proteomics. 2012, 11, M111.014720. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.H.; Shah, P.; Chen, Y.W.; Chen, C.S. Systematic Analysis of Intracellular-targeting Antimicrobial Peptides, Bactenecin 7, Hybrid of Pleurocidin and Dermaseptin, Proline-Arginine-rich Peptide, and Lactoferricin B, by Using Escherichia coli Proteome Microarrays. Mol. Cell. Proteomics. 2016, 15, 1837–1847. [Google Scholar] [CrossRef] [PubMed]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar]
- Low, C.Y.; Rotstein, C. Emerging fungal infections in immunocompromised patients. F1000 Med. Rep. 2011, 3, 14. [Google Scholar] [CrossRef]
- Scorzoni, L.; de Paula, E.S.A.C.; Marcos, C.M.; Assato, P.A.; de Melo, W.C.; de Oliveira, H.C.; Costa-Orlandi, C.B.; Mendes-Giannini, M.J.; Fusco-Almeida, A.M. Antifungal Therapy: New Advances in the Understanding and Treatment of Mycosis. Front. Microbiol. 2017, 8, 36. [Google Scholar] [CrossRef]
- McCarthy, M.W.; Kontoyiannis, D.P.; Cornely, O.A.; Perfect, J.R.; Walsh, T.J. Novel Agents and Drug Targets to Meet the Challenges of Resistant Fungi. J. Infect. Dis. 2017, 216, S474–S483. [Google Scholar] [CrossRef]
- Ebbensgaard, A.; Mordhorst, H.; Overgaard, M.T.; Nielsen, C.G.; Aarestrup, F.M.; Hansen, E.B. Comparative Evaluation of the Antimicrobial Activity of Different Antimicrobial Peptides against a Range of Pathogenic Bacteria. PLoS ONE 2015, 10, e0144611. [Google Scholar] [CrossRef] [PubMed]
- Hilpert, K.; McLeod, B.; Yu, J.; Elliott, M.R.; Rautenbach, M.; Ruden, S.; Burck, J.; Muhle-Goll, C.; Ulrich, A.S.; Keller, S.; et al. Short cationic antimicrobial peptides interact with ATP. Antimicrob. Agents. Chemother. 2010, 54, 4480–4483. [Google Scholar] [CrossRef]
- Dominguez, R.; Holmes, K.C. Actin structure and function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef] [PubMed]
- Gourlay, C.W.; Carpp, L.N.; Timpson, P.; Winder, S.J.; Ayscough, K.R. A role for the actin cytoskeleton in cell death and aging in yeast. J. Cell. Biol. 2004, 164, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, A.; Geng, M.; Hull, K.G.; Li, J.; Romo, D.; Lu, S.E.; Albee, A.; Nutter, C.; Gordon, D.M.; Ghannoum, M.A.; et al. A Novel Actin Binding Drug with In Vivo Efficacy. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Zhu, H.; Bilgin, M.; Bangham, R.; Hall, D.; Casamayor, A.; Bertone, P.; Lan, N.; Jansen, R.; Bidlingmaier, S.; Houfek, T.; et al. Global analysis of protein activities using proteome chips. Science 2001, 293, 2101–2105. [Google Scholar] [CrossRef]
- Lu, K.Y.; Tao, S.C.; Yang, T.C.; Ho, Y.H.; Lee, C.H.; Lin, C.C.; Juan, H.F.; Huang, H.C.; Yang, C.Y.; Chen, M.S.; et al. Profiling lipid-protein interactions using nonquenched fluorescent liposomal nanovesicles and proteome microarrays. Mol. Cell. Proteomics 2012, 11, 1177–1190. [Google Scholar] [CrossRef]
- Gelperin, D.M.; White, M.A.; Wilkinson, M.L.; Kon, Y.; Kung, L.A.; Wise, K.J.; Lopez-Hoyo, N.; Jiang, L.; Piccirillo, S.; Yu, H.; et al. Biochemical and genetic analysis of the yeast proteome with a movable ORF collection. Genes Dev. 2005, 19, 2816–2826. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Enrichment in Molecular Function | p-Value | Hit in This Category | Total Gene in This Category |
|---|---|---|---|---|
| GO:0030029 | actin filament-based process | 0.0000136 | 13 | 129 |
| GO:0007015 | actin filament organization | 0.0000203 | 10 | 74 |
| GO:0060255 | regulation of macromolecule metabolic process | 0.000494 | 40 | 1124 |
| GO:0044396 | actin cortical patch organization | 0.000558 | 5 | 19 |
| GO:0080090 | regulation of primary metabolic process | 0.000628 | 40 | 1137 |
| GO:0031323 | regulation of cellular metabolic process | 0.000726 | 40 | 1145 |
| GO:0048523 | negative regulation of cellular process | 0.001034 | 25 | 592 |
| GO:0050790 | regulation of catalytic activity | 0.001206 | 19 | 392 |
| GO:1902589 | single-organism organelle organization | 0.00121 | 23 | 528 |
| GO:0007114 | cell budding | 0.00149 | 7 | 60 |
| GO:0051301 | cell division | 0.00157 | 16 | 304 |
| GO:0022607 | cellular component assembly | 0.00207 | 31 | 848 |
| GO:0043254 | regulation of protein complex assembly | 0.00265 | 8 | 90 |
| GO:0010646 | regulation of cell communication | 0.00844 | 9 | 139 |
| GO:0033554 | cellular response to stress | 0.01015 | 25 | 705 |
| GO:0043933 | macromolecular complex subunit organization | 0.01683 | 30 | 938 |
| GO:0009408 | response to heat | 0.02003 | 6 | 75 |
| GO:0009966 | regulation of signal transduction | 0.02036 | 8 | 132 |
| GO:0022411 | cellular component disassembly | 0.02356 | 8 | 136 |
| GO:0009266 | response to temperature stimulus | 0.02696 | 6 | 81 |
| GO:0032231 | regulation of actin filament bundle assembly | 0.02925 | 3 | 13 |
| GO:0007049 | cell cycle | 0.03250 | 25 | 782 |
| GO:0007117 | budding cell bud growth | 0.03356 | 4 | 34 |
| GO:0007163 | establishment or maintenance of cell polarity | 0.03434 | 7 | 116 |
| GO:0009889 | regulation of biosynthetic process | 0.03619 | 29 | 956 |
| GO:0006996 | organelle organization | 0.03937 | 46 | 1704 |
| GO:0043170 | macromolecule metabolic process | 0.04550 | 77 | 3178 |
| GO:0031324 | negative regulation of cellular metabolic process | 0.04558 | 16 | 447 |
| GO:0051171 | regulation of nitrogen compound metabolic process | 0.04726 | 29 | 979 |
| GO:0009892 | negative regulation of metabolic process | 0.04787 | 16 | 450 |
| Term | Enrichment in Molecular Function | p-Value | Hit in This Category | Total Gene in This Category |
|---|---|---|---|---|
| GO:0005938 | cell cortex | 0.000022 | 15 | 175 |
| GO:0030427 | site of polarized growth | 0.000077 | 18 | 274 |
| GO:0005937 | mating projection | 0.000131 | 12 | 133 |
| GO:0042995 | cell projection | 0.00015 | 12 | 135 |
| GO:0043332 | mating projection tip | 0.000219 | 11 | 118 |
| GO:0043232 | intracellular non-membrane-bounded organelle | 0.000245 | 50 | 1429 |
| GO:0044463 | cell projection part | 0.000425 | 11 | 128 |
| GO:0044430 | cytoskeletal part | 0.000524 | 15 | 235 |
| GO:0005933 | cellular bud | 0.008774 | 13 | 254 |
| GO:0044428 | nuclear part | 0.00938 | 40 | 1260 |
| GO:0005935 | cellular bud neck | 0.01021 | 11 | 197 |
| GO:0044424 | intracellular part | 0.02049 | 127 | 5588 |
| GO:0044427 | chromosomal part | 0.02443 | 17 | 433 |
| GO:0005622 | intracellular | 0.02741 | 127 | 5603 |
| GO:0070938 | contractile ring | 0.04085 | 3 | 15 |
| GO:0005826 | actomyosin contractile ring | 0.04085 | 3 | 15 |
| GO:0036286 | eisosome filament | 0.04297 | 2 | 2 |
| Term | Enrichment in Molecular Function | p-Value | Hit in This Category | Total Gene in This Category |
|---|---|---|---|---|
| GO:0019899 | enzyme binding | 0.000705 | 12 | 153 |
| GO:0008092 | cytoskeletal protein binding | 0.0019716 | 9 | 100 |
| GO:0032403 | protein complex binding | 0.0138161 | 7 | 86 |
| GO:0042393 | histone binding | 0.0207646 | 5 | 46 |
| GO:0019208 | phosphatase regulator activity | 0.049133 | 4 | 36 |
| p-Value | Hit in This Category | Total Gene in This Category | Genes | |
|---|---|---|---|---|
| Cytoskeletal-regulatory complex EF hand (EF hand represent helix E and F in parvalbumin) | 0.001542 | 3 | 3 | YBL047C, YNL084C, YIR006C |
| SRC homology 3 domain (SH3 domain) | 0.011565 | 4 | 21 | YER118C, YAR014C, YJL020C, YCR088W |
| Ubiquitin-conjugating enzyme | 0.039661 | 3 | 14 | YDR054C, YEL012W, YGL058W |
| Eisosome component PIL1 protein | 0.045367 | 2 | 2 | YGR086C, YPL004C |
| Tropomyosin like | 0.045367 | 2 | 2 | YNL079C, YIL138C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, P.; Chen, C.-S. Systematic Identification of Protein Targets of Sub5 Using Saccharomyces cerevisiae Proteome Microarrays. Int. J. Mol. Sci. 2021, 22, 760. https://doi.org/10.3390/ijms22020760
Shah P, Chen C-S. Systematic Identification of Protein Targets of Sub5 Using Saccharomyces cerevisiae Proteome Microarrays. International Journal of Molecular Sciences. 2021; 22(2):760. https://doi.org/10.3390/ijms22020760
Chicago/Turabian StyleShah, Pramod, and Chien-Sheng Chen. 2021. "Systematic Identification of Protein Targets of Sub5 Using Saccharomyces cerevisiae Proteome Microarrays" International Journal of Molecular Sciences 22, no. 2: 760. https://doi.org/10.3390/ijms22020760
APA StyleShah, P., & Chen, C.-S. (2021). Systematic Identification of Protein Targets of Sub5 Using Saccharomyces cerevisiae Proteome Microarrays. International Journal of Molecular Sciences, 22(2), 760. https://doi.org/10.3390/ijms22020760