Experimental Model Systems for Understanding Human Axonal Injury Responses


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
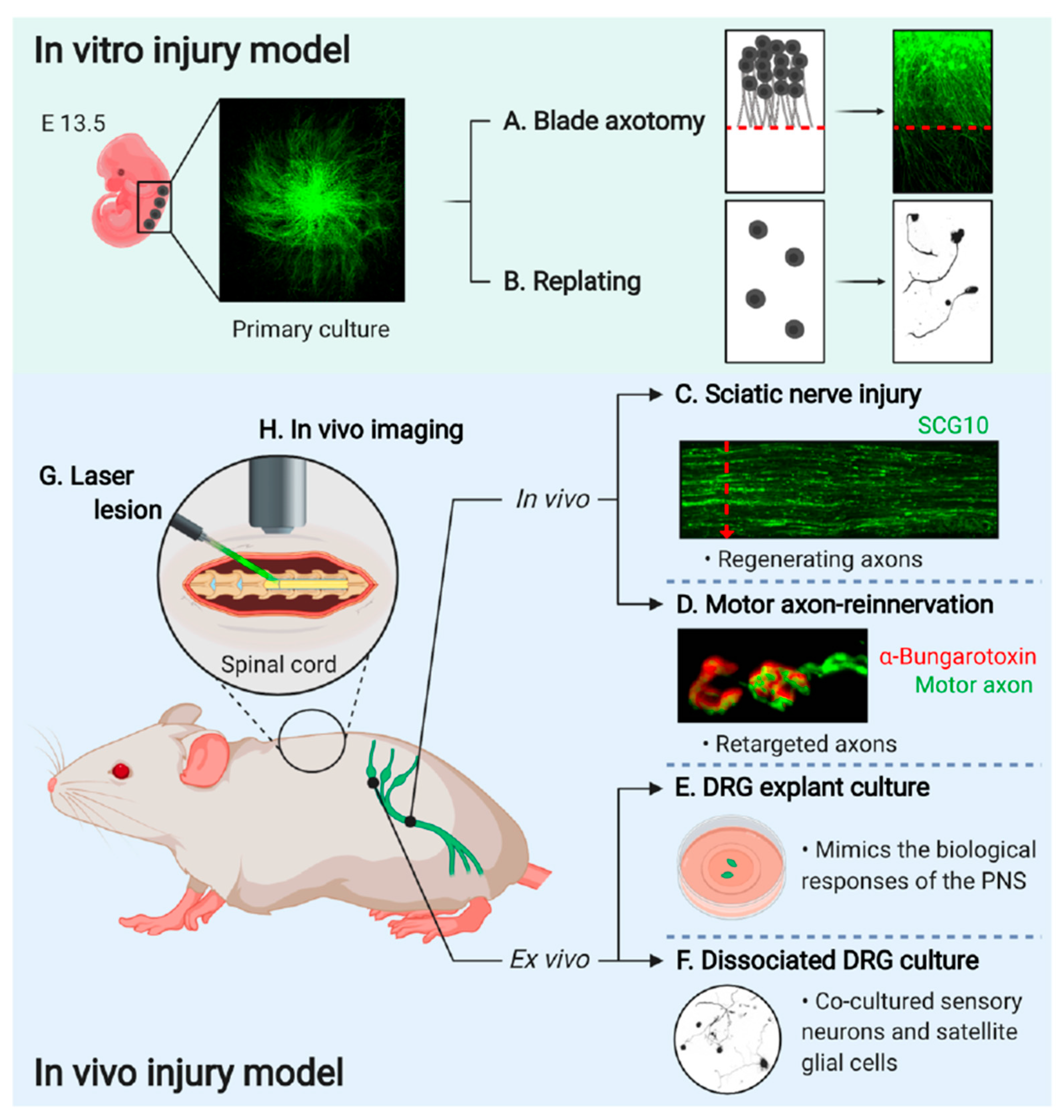
2. In Vitro Axon Injury Model
2.1. Blade Axotomy
2.2. Replating Assay
3. In Vivo Axon Injury Model
3.1. Sciatic Nerve Injury Model
3.2. Motor Axon-Reinnervation Model
3.3. Spinal Cord Injury Model
3.4. In Vivo Imaging
4. Ex Vivo Injury Model
4.1. DRG Explant Culture
4.2. Dissociated DRG Neuron Culture
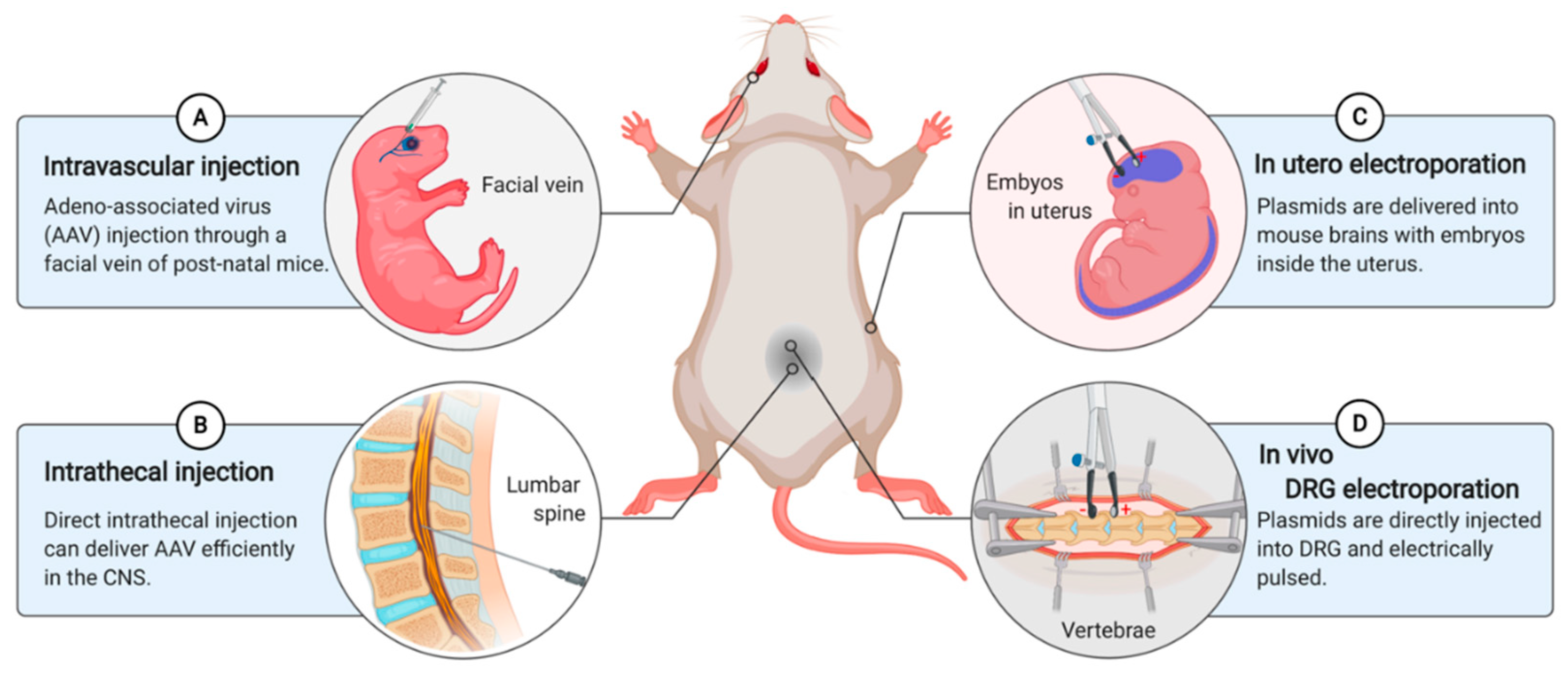
5. In Vivo Gene Delivery
5.1. AAV-Mediated In Vivo Gene Delivery
5.2. In Utero Electroporation
5.3. In Vivo DRG Electroporation
6. Recent Innovations Using Experimental Systems
6.1. SARM1 and NMNAT2
6.2. DLK
7. Conclusions and Perspectives
7.1. The Advantages of Experimenting with Rodent Models
7.2. Limitations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Meaning |
| PNS | peripheral nervous system |
| CNS | central nervous system |
| RNP | ribonucleotide protein |
| RBP | RNA binding protein |
| CSPG | chondroitin sulfate proteoglycan |
| OMpg | oligodendrocyte-myelin glycoprotein |
| MAG | myelin-associated glycoprotein |
| DRG | dorsal root ganglion |
| DEG | differentially expressed gene |
| TMEM184b | Transmembrane protein 184b |
| MORN4 | MORN repeat containing 4 |
| GSK3 | Glycogen synthase kinase 3 |
| SCG10 | superior cervical ganglion 10 |
| EHL | extensor hallucis longus |
| AAV | Adeno-associated virus |
| IUE | in utero electroporation |
| Wlds | Wallerian degeneration slow |
| NMNAT | nicotinamide mononucleotide adenylyltransferase |
| SARM1 | Sterile Alpha and Toll Interleukin Receptor Motif-containing protein 1 |
| DLK | dual leucine zipper kinase |
| JNK | c-Jun N-terminal kinase |
| JIP3 | JNK-interacting protein 3 |
| ALS | Amyotrophic lateral sclerosis |
| iPSC | induced pluripotent stem cell |
References
- Santiago, P. The Croonian lecture—La fine structure des centres nerveux. Proc. R. Soc. Lond. 1894, 55, 444–468. [Google Scholar]
- Campbell, D.S.; Holt, C.E. Chemotropic responses of retinal growth cones mediated by rapid local protein synthesis and degradation. Neuron 2001, 32, 1013–1026. [Google Scholar] [CrossRef]
- Holt, C.E.; Schuman, E.M. The central dogma decentralized: New perspectives on RNA function and local translation in neurons. Neuron 2013, 80, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Rishal, I.; Fainzilber, M. Axon-soma communication in neuronal injury. Nat. Rev. Neurosci. 2014, 15, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Ertürk, A.; Hellal, F.; Enes, J.; Bradke, F. Disorganized microtubules underlie the formation of retraction bulbs and the failure of axonal regeneration. J. Neurosci. 2007, 27, 9169–9180. [Google Scholar] [CrossRef] [PubMed]
- Ziv, N.E.; Spira, M.E. Localized and transient elevations of intracellular Ca2+ induce the dedifferentiation of axonal segments into growth cones. J. Neurosci. 1997, 17, 3568–3579. [Google Scholar] [CrossRef] [PubMed]
- Bradke, F.; Fawcett, J.W.; Spira, M.E. Assembly of a new growth cone after axotomy: The precursor to axon regeneration. Nat. Rev. Neurosci. 2012, 13, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Tom, V.J.; Steinmetz, M.P.; Miller, J.H.; Doller, C.M.; Silver, J. Studies on the development and behavior of the dystrophic growth cone, the hallmark of regeneration failure, in an in vitro model of the glial scar and after spinal cord injury. J. Neurosci. 2004, 24, 6531–6539. [Google Scholar] [CrossRef]
- Ming, G.L.; Wong, S.T.; Henley, J.; Yuan, X.B.; Song, H.J.; Spitzer, N.C.; Poo, M.M. Adaptation in the chemotactic guidance of nerve growth cones. Nature 2002, 417, 411–418. [Google Scholar] [CrossRef]
- Ziv, N.E.; Spira, M.E. Spatiotemporal Distribution of Ca2+ Following Axotomy and Throughout the Recovery Process of Cultured Aplysia Neurons. Eur. J. Neurosci. 1993, 5, 657–668. [Google Scholar] [CrossRef]
- Ji, S.J.; Jaffrey, S.R. Axonal transcription factors: Novel regulators of growth cone- to-nucleus signaling. Dev Neurobiol. 2014, 74, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Baleriola, J.; Walker, C.A.; Jean, Y.Y.; Crary, J.F.; Troy, C.M.; Nagy, P.L.; Hengst, U. Axonally synthesized ATF4 transmits a neurodegenerative signal across brain regions Jimena. Cell 2014, 158, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Cavalli, V. HDAC5 is a novel injury-regulated tubulin deacetylase controlling axon regeneration. EMBO J. 2012, 31, 3063–3078. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.S.; Skene, J.H. A transcription-dependent switch controls competence of adult neurons for distinct modes of axon growth. J. Neurosci. 1997, 17, 646–658. [Google Scholar] [CrossRef] [PubMed]
- Shigeoka, T.; Jung, H.; Jung, J.; Turner-Bridger, B.; Ohk, J.; Lin, J.Q.; Amieux, P.S.; Holt, C.E. Dynamic Axonal Translation in Developing and Mature Visual Circuits. Cell 2016, 166, 181–192. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Smith, D.S.; Perrone-Bizzozero, N.; Twiss, J.L. Axonal mRNA transport and translation at a glance. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef]
- Magill, C.K.; Tong, A.; Kawamura, D.; Hayashi, A.; Hunter, D.A.; Parsadanian, A.; Mackinnon, S.E.; Myckatyn, T.M. Reinnervation of the tibialis anterior following sciatic nerve crush injury: A confocal microscopic study in transgenic mice. Exp. Neurol. 2007, 207, 64–74. [Google Scholar] [CrossRef]
- De Lima, S.; Koriyama, Y.; Kurimoto, T.; Oliveira, J.T.; Yin, Y.; Li, Y.; Gilbert, H.Y.; Fagiolini, M.; Martinez, A.M.B.; Benowitz, L. Full-length axon regeneration in the adult mouse optic nerve and partial recovery of simple visual behaviors. Proc. Natl. Acad. Sci. USA 2012, 109, 9149–9154. [Google Scholar] [CrossRef]
- Gerdts, J.; Brace, E.J.; Sasaki, Y.; DiAntonio, A.; Milbrandt, J. SARM1 activation triggers axon degeneration locally via NAD+ destruction. Science (80-) 2015, 348, 453–457. [Google Scholar] [CrossRef]
- Osterloh, J.M.; Yang, J.; Rooney, T.M.; Fox, A.N.; Adalbert, R.; Powell, E.H.; Sheehan, A.E.; Avery, M.A.; Hackett, R.; Logan, M.A.; et al. dSarm/Sarm1 is required for activation of an injury-induced axon death pathway. Science (80-) 2012, 337, 481–484. [Google Scholar] [CrossRef]
- Waller, A. Experiments on the section of the glossopharyngeal and hypoglossal nerves of the frog, and observations of the alterations produced thereby in the structure of their primitive fibres. Abstr. Pap. Commun. R. Soc. Lond. 1851, 5, 924–925. [Google Scholar] [CrossRef]
- Coleman, M.P.; Höke, A. Programmed axon degeneration: From mouse to mechanism to medicine. Nat. Rev. Neurosci. 2020, 21, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.T.; Medress, Z.A.; Barres, B.A. Axon degeneration: Molecular mechanisms of a self-destruction pathway. J. Cell Biol. 2012, 196, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Gerdts, J.; Summers, D.W.; Milbrandt, J.; DiAntonio, A. Axon Self-Destruction: New Links among SARM1, MAPKs, and NAD+ Metabolism. Neuron 2016, 89, 449–460. [Google Scholar] [CrossRef]
- Yiu, G.; He, Z. Glial inhibition of CNS axon regeneration. Nat. Rev. Neurosci. 2006, 7, 617–627. [Google Scholar] [CrossRef]
- David, S.; Aguayo, A.J. Axonal elongation into peripheral nervous system “bridges” after central nervous system injury in adult rats. Science (80-) 1981, 214, 931–933. [Google Scholar] [CrossRef]
- Mahar, M.; Cavalli, V. Intrinsic mechanisms of neuronal axon regeneration. Nat. Rev. Neurosci. 2018, 19, 323–337. [Google Scholar] [CrossRef]
- Tedeschi, A.; Bradke, F. Spatial and temporal arrangement of neuronal intrinsic and extrinsic mechanisms controlling axon regeneration. Curr. Opin. Neurobiol. 2017, 42, 118–127. [Google Scholar] [CrossRef]
- Lindner, R.; Puttagunta, R.; Di Giovanni, S. Epigenetic Regulation of Axon Outgrowth and Regeneration in CNS Injury: The First Steps Forward. Neurotherapeutics 2013, 10, 771–781. [Google Scholar] [CrossRef]
- Shin, J.E.; Cho, Y. Epigenetic Regulation of Axon Regeneration after Neural Injury. Mol. Cells 2017, 40, 10–16. [Google Scholar] [CrossRef]
- Shin, J.E.; Ha, H.; Cho, E.H.; Kim, Y.K.; Cho, Y. Comparative analysis of the transcriptome of injured nerve segments reveals spatiotemporal responses to neural damage in mice. J. Comp. Neurol. 2018, 526, 1195–1208. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.P.; Sahoo, P.K.; Kar, A.N.; Twiss, J.L. Intra-axonal mechanisms driving axon regeneration. Brain Res. 2020, 1740. [Google Scholar] [CrossRef] [PubMed]
- Figley, M.D.; DiAntonio, A. The SARM1 axon degeneration pathway: Control of the NAD+ metabolome regulates axon survival in health and disease. Curr. Opin. Neurobiol. 2020, 63, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.R.C.; Geisler, S.; Pittman, S.K.; Doan, R.A.; Weihl, C.C.; Milbrandt, J.; DiAntonio, A. TMEM184b Promotes Axon Degeneration and Neuromuscular Junction Maintenance. J. Neurosci. 2016, 36, 4681–4689. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.R.C.; Gerdts, J.; Naylor, S.A.; Royse, E.X.; Ebstein, S.Y.; Sasaki, Y.; Milbrandt, J.; DiAntonio, A. A model of toxic neuropathy in Drosophila reveals a role for MORN4 in promoting axonal degeneration. J. Neurosci. 2012, 32, 5054–5061. [Google Scholar] [CrossRef] [PubMed]
- Gerdts, J.; Sasaki, Y.; Vohra, B.; Marasa, J.; Milbrandt, J. Image-based screening identifies novel roles for IkappaB kinase and glycogen synthase kinase 3 in axonal degeneration. J. Biol. Chem. 2011, 286, 28011–28018. [Google Scholar] [CrossRef]
- Frey, E.; Valakh, V.; Karney-Grobe, S.; Shi, Y.; Milbrandt, J.; DiAntonio, A. An in vitro assay to study induction of the regenerative state in sensory neurons. Exp. Neurol. 2015, 263, 350–363. [Google Scholar] [CrossRef]
- Shin, J.E.; Geisler, S.; DiAntonio, A. Dynamic regulation of SCG10 in regenerating axons after injury. Exp. Neurol. 2014, 252, 1–11. [Google Scholar] [CrossRef]
- Sharthiya, H.; Seng, C.; Van Kuppevelt, T.H.; Tiwari, V.; Fornaro, M. HSV-1 interaction to 3-O-sulfated heparan sulfate in mouse-derived DRG explant and profiles of inflammatory markers during virus infection. J. Neurovirol. 2017, 23, 483–491. [Google Scholar] [CrossRef]
- Ylera, B.; Ertürk, A.; Hellal, F.; Nadrigny, F.; Hurtado, A.; Tahirovic, S.; Oudega, M.; Kirchhoff, F.; Bradke, F. Chronically CNS-Injured Adult Sensory Neurons Gain Regenerative Competence upon a Lesion of Their Peripheral Axon. Curr. Biol. 2009, 19, 930–936. [Google Scholar] [CrossRef]
- Schaffran, B.; Hilton, B.J.; Bradke, F. Imaging in vivo dynamics of sensory axon responses to CNS injury. Exp. Neurol. 2019, 317, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Mason, M.R.J.; Lieberman, A.R.; Grenningloh, G.; Anderson, P.N. Transcriptional upregulation of SCG10 and CAP-23 is correlated with regeneration of the axons of peripheral and central neurons in vivo. Mol. Cell. Neurosci. 2002, 20, 595–615. [Google Scholar] [CrossRef] [PubMed]
- Voria, I.; Hauser, J.; Axis, A.; Schenker, M.; Bichet, S.; Kuntzer, T.; Grenningloh, G.; Barakat-Walter, I. Improved sciatic nerve regeneration by local thyroid hormone treatment in adult rat is accompanied by increased expression of SCG10. Exp. Neurol. 2006, 197, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Saijilafu; Hur, E.M.; Liu, C.M.; Jiao, Z.; Xu, W.L.; Zhou, F.Q. PI3K-GSK3 signalling regulates mammalian axon regeneration by inducing the expression of Smad1. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Ho, C.; Wong, K.; Tessier-Lavigne, M. Axotomy-induced smad1 activation promotes axonal growth in adult sensory neurons. J. Neurosci. 2009, 29, 7116–7123. [Google Scholar] [CrossRef]
- Valakh, V.; Frey, E.; Babetto, E.; Walker, L.J.; DiAntonio, A. Cytoskeletal disruption activates the DLK/JNK pathway, which promotes axonal regeneration and mimics a preconditioning injury. Neurobiol. Dis. 2015, 77, 13–25. [Google Scholar] [CrossRef]
- Lee, J.K.; Zheng, B. Axon regeneration after spinal cord injury: Insight from genetically modified mouse models. Restor. Neurol. Neurosci. 2008, 26, 175–182. [Google Scholar]
- Zurborg, S.; Piszczek, A.; Martínez, C.; Hublitz, P.; Al Banchaabouchi, M.; Moreira, P.; Perlas, E.; Heppenstall, P.A. Generation and characterization of an Advillin-Cre driver mouse line. Mol. Pain 2011, 7, 1–10. [Google Scholar] [CrossRef]
- Wilson, J.M.; Hartley, R.; Maxwell, D.J.; Todd, A.J.; Lieberam, I.; Kaltschmidt, J.A.; Yoshida, Y.; Jessell, T.M.; Brownstone, R.M. Conditional rhythmicity of ventral spinal interneurons defined by expression of the Hb9 homeodomain protein. J. Neurosci. 2005, 25, 5710–5719. [Google Scholar] [CrossRef]
- Arber, S.; Han, B.; Mendelsohn, M.; Smith, M.; Jessell, T.M.; Sockanathan, S. Requirement for the Homeobox Gene Hb9 in the Consolidation of Motor Neuron Identity. Neuron 1999, 23, 659–674. [Google Scholar] [CrossRef]
- Caldeira, V.; Dougherty, K.J.; Borgius, L.; Kiehn, O. Spinal Hb9::Cre-derived excitatory interneurons contribute to rhythm generation in the mouse. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Shin, J.E.; Lee, B.; Kim, H.; Jeon, Y.; Ahn, S.H.; Chi, S.W.; Cho, Y. The stem cell marker Prom1 promotes axon regeneration by down-regulating cholesterol synthesis via Smad signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 15955–15966. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.M.; Ebendal, T. Nerve growth activities in rat peripheral nerve. Brain Res. 1982, 246, 57–64. [Google Scholar] [CrossRef]
- Neumann, S.; Woolf, C.J. Regeneration of dorsal column fibers into and beyond the lesion site following adult spinal cord injury. Neuron 1999, 23, 83–91. [Google Scholar] [CrossRef]
- Abe, N.; Cavalli, V. Nerve injury signaling. Curr. Opin. Neurobiol. 2008, 18, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.E.; Cho, Y.; Beirowski, B.; Milbrandt, J.; Cavalli, V.; DiAntonio, A. Dual Leucine Zipper Kinase Is Required for Retrograde Injury Signaling and Axonal Regeneration. Neuron 2012, 74, 1015–1022. [Google Scholar] [CrossRef]
- Hanz, S.; Fainzilber, M. Retrograde signaling in injured nerve-The axon reaction revisited. J. Neurochem. 2006, 99, 13–19. [Google Scholar] [CrossRef]
- Cavalli, V.; Kujala, P.; Klumperman, J.; Goldstein, L.S.B. Sunday driver links axonal transport to damage signaling. J. Cell Biol. 2005, 168, 775–787. [Google Scholar] [CrossRef]
- Vannucci, B.; Santosa, K.B.; Keane, A.M.; Jablonka-Shariff, A.; Lu, C.Y.; Yan, Y.; MacEwan, M.; Snyder-Warwick, A.K. What is Normal? Neuromuscular junction reinnervation after nerve injury. Muscle Nerve 2019, 60, 604–612. [Google Scholar] [CrossRef]
- Hilton, B.J.; Assinck, P.; Duncan, G.J.; Lu, D.; Lo, S.; Tetzlaff, W. Dorsolateral funiculus lesioning of the mouse cervical spinal cord at C4 but not at C6 results in sustained forelimb motor deficits. J. Neurotrauma 2013, 30, 1070–1083. [Google Scholar] [CrossRef]
- Sun, F.; Park, K.K.; Belin, S.; Wang, D.; Lu, T.; Chen, G.; Zhang, K.; Yeung, C.; Feng, G.; Yankner, B.A.; et al. Sustained axon regeneration induced by co-deletion of PTEN and SOCS3. Nature 2011, 480, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Tuszynski, M.H.; Steward, O. Concepts and Methods for the Study of Axonal Regeneration in the CNS. Neuron 2012, 74, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Cregg, J.M.; DePaul, M.A.; Filous, A.R.; Lang, B.T.; Tran, A.; Silver, J. Functional regeneration beyond the glial scar. Exp. Neurol. 2014, 253, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Canty, A.J.; Teles-Grilo ruivo, L.M.; Nesarajah, C.; Song, S.; Jackson, J.S.; Little, G.E.; De Paola, V. Synaptic elimination and protection after minimal injury depend on cell type and their prelesion structural dynamics in the adult cerebral cortex. J. Neurosci. 2013, 33, 10374–10383. [Google Scholar] [CrossRef] [PubMed]
- Kerschensteiner, M.; Schwab, M.E.; Lichtman, J.W.; Misgeld, T. In vivo imaging of axonal degeneration and regeneration in the injured spinal cord. Nat. Med. 2005, 11, 572–577. [Google Scholar] [CrossRef]
- Lorenzana, A.O.; Lee, J.K.; Mui, M.; Chang, A.; Zheng, B. A Surviving Intact Branch Stabilizes Remaining Axon Architecture after Injury as Revealed by InVivo Imaging in the Mouse Spinal Cord. Neuron 2015, 86, 947–954. [Google Scholar] [CrossRef]
- Farrar, M.J.; Bernstein, I.M.; Schlafer, D.H.; Cleland, T.A.; Fetcho, J.R.; Schaffer, C.B. Chronic in vivo imaging in the mouse spinal cord using an implanted chamber. Nat. Methods 2012, 9, 297–302. [Google Scholar] [CrossRef]
- Fenrich, K.K.; Weber, P.; Hocine, M.; Zalc, M.; Rougon, G.; Debarbieux, F. Long-term in vivo imaging of normal and pathological mouse spinal cord with subcellular resolution using implanted glass windows. J. Physiol. 2012, 590, 3665–3675. [Google Scholar] [CrossRef]
- Figley, S.A.; Chen, Y.; Maeda, A.; Conroy, L.; McMullen, J.D.; Silver, J.I.; Stapleton, S.; Vitkin, A.; Lindsay, P.; Burrell, K.; et al. A Spinal Cord Window Chamber Model for In Vivo Longitudinal Multimodal Optical and Acoustic Imaging in a Murine Model. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Tedeschi, A.; Dupraz, S.; Laskowski, C.J.; Xue, J.; Ulas, T.; Beyer, M.; Schultze, J.L.; Bradke, F. The Calcium Channel Subunit Alpha2delta2 Suppresses Axon Regeneration in the Adult CNS. Neuron 2016, 92, 419–434. [Google Scholar] [CrossRef]
- Fornaro, M.; Sharthiya, H.; Tiwari, V. Adult mouse DRG explant and dissociated cell models to investigate neuroplasticity and responses to environmental insults including viral infection. J. Vis. Exp. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Borson, S.H.; Gambello, M.J.; Wang, F.; Cavalli, V. Mammalian Target of Rapamycin (mTOR) activation increases axonal growth capacity of injured peripheral nerves. J. Biol. Chem. 2010, 285, 28034–28043. [Google Scholar] [CrossRef] [PubMed]
- Gombash Lampe, S.E.; Kaspar, B.K.; Foust, K.D. Intravenous Injections in Neonatal Mice. J. Vis. Exp. 2014, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Gessler, D.J.; Tai, P.W.L.; Li, J.; Gao, G. Intravenous Infusion of AAV for Widespread Gene Delivery to the Nervous System. In Adeno-Associated Virus Vectors: Design and Delivery; Castle, M.J., Ed.; Springer: New York, NY, USA, 2019; pp. 143–163. ISBN 9781493991396. [Google Scholar]
- Wang, D.; Tai, P.W.L.; Gao, G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat. Rev. Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef]
- Vickers, D.A.L.; Hur, S.C. Microscale vortex-assisted electroporator for sequential molecular delivery. J. Vis. Exp. 2014, 1–7. [Google Scholar] [CrossRef]
- Tabata, H.; Nakajima, K. Efficient in utero gene transfer system to the developing mouse brain using electroporation: Visualization of neuronal migration in the developing cortex. Neuroscience 2001, 103, 865–872. [Google Scholar] [CrossRef]
- Dal Maschio, M.; Ghezzi, D.; Bony, G.; Alabastri, A.; Deidda, G.; Brondi, M.; Sato, S.S.; Zaccaria, R.P.; Di Fabrizio, E.; Ratto, G.M.; et al. High-performance and site-directed in utero electroporation by a triple-electrode probe. Nat. Commun. 2012, 3. [Google Scholar] [CrossRef]
- Saijilafu; Zhang, B.-Y.; Zhou, F.-Q. In vivo electroporation of adult mouse sensory neurons for studying peripheral axon regeneration. Methods Mol. Biol. 2014, 1162, 167–175. [Google Scholar] [CrossRef]
- Li, Q.; Qian, C.; Zhou, F.Q. Investigating mammalian axon regeneration: In vivo electroporation of adult mouse dorsal root ganglion. J. Vis. Exp. 2018, 2018, 2–7. [Google Scholar] [CrossRef]
- Perry, V.H.; Lunn, E.R.; Brown, M.C.; Cahusac, S.; Gordon, S. Evidence that the Rate of Wallerian Degeneration is Controlled by a Single Autosomal Dominant Gene. Eur. J. Neurosci. 1990, 2, 408–413. [Google Scholar] [CrossRef]
- Beuche, W.; Friede, R.L. Naked axon bundles enclosed by single segments of myelin sheaths in the nerves of non-dystrophic C57BL-ob/+mice. Neuropathol. Appl. Neurobiol. 1984, 10, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Lunn, E.R.; Perry, V.H.; Brown, M.C.; Rosen, H.; Gordon, S. Absence of Wallerian Degeneration does not Hinder Regeneration in Peripheral Nerve. Eur. J. Neurosci. 1989, 1, 27–33. [Google Scholar] [CrossRef]
- Beuche, W.; Friede, R.L. The role of non-resident cells in Wallerian degeneration. J. Neurocytol. 1984, 13, 767–796. [Google Scholar] [CrossRef]
- Martin, S.M.; O’Brien, G.S.; Portera-Cailliau, C.; Sagasti, A. Wallerian degeneration of zebrafish trigeminal axons in the skin is required for regeneration and developmental pruning. Development 2010, 137, 3985–3994. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, J.M.; Beach, M.G.; Porpiglia, E.; Sheehan, A.E.; Watts, R.J.; Freeman, M.R. The Drosophila Cell Corpse Engulfment Receptor Draper Mediates Glial Clearance of Severed Axons. Neuron 2006, 50, 869–881. [Google Scholar] [CrossRef]
- Laser, H.; Conforti, L.; Morreale, G.; Mack, T.G.M.; Heyer, M.; Haley, J.E.; Wishart, T.M.; Beirowski, B.; Walker, S.A.; Haase, G.; et al. The slow Wallerian degeneration protein, WldS, binds directly to VCP/p97 and partially redistributes it within the nucleus. Mol. Biol. Cell 2006, 17, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Kitay, B.M.; McCormack, R.; Wang, Y.; Tsoulfas, P.; Zhai, R.G. Mislocalization of neuronal mitochondria reveals regulation of Wallerian degeneration and NMNAT/WLD(S)-mediated axon protection independent of axonal mitochondria. Hum. Mol. Genet. 2013, 22, 1601–1614. [Google Scholar] [CrossRef] [PubMed]
- Araki, T.; Sasaki, Y.; Milbrandt, J. Increased nuclear NAD biosynthesis and SIRT1 activation prevent axonal degeneration. Science (80-) 2004, 305, 1010–1013. [Google Scholar] [CrossRef] [PubMed]
- Conforti, L.; Tarlton, A.; Mack, T.G.A.; Mi, W.; Buckmaster, E.A.; Wagner, D.; Perry, V.H.; Coleman, M.P. A Ufd2/D4Cole1e chimeric protein and overexpression of Rbp7 in the slow Wallerian degeneration (Wld(s)) mouse. Proc. Natl. Acad. Sci. USA 2000, 97, 11377–11382. [Google Scholar] [CrossRef]
- Gilley, J.; Orsomando, G.; Nascimento-Ferreira, I.; Coleman, M.P. Absence of SARM1 rescues development and survival of NMNAT2-Deficient axons. Cell Rep. 2015, 10, 1975–1982. [Google Scholar] [CrossRef]
- Gilley, J.; Coleman, M.P. Endogenous Nmnat2 Is an Essential Survival Factor for Maintenance of Healthy Axons. PLoS Biol. 2010, 8. [Google Scholar] [CrossRef]
- Loreto, A.; Hill, C.S.; Hewitt, V.L.; Orsomando, G.; Angeletti, C.; Gilley, J.; Lucci, C.; Sanchez-Martinez, A.; Whitworth, A.J.; Conforti, L.; et al. Mitochondrial impairment activates the Wallerian pathway through depletion of NMNAT2 leading to SARM1-dependent axon degeneration. Neurobiol. Dis. 2020, 134, 104678. [Google Scholar] [CrossRef] [PubMed]
- Mink, M.; Fogelgren, B.; Olszewski, K.; Maroy, P.; Csiszar, K. A novel human gene (SARM) at chromosome 17q11 encodes a protein with a SAM motif and structural similarity to Armadillo/beta-catenin that is conserved in mouse, Drosophila, and Caenorhabditis elegans. Genomics 2001, 74, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.J.; Summers, D.W.; Sasaki, Y.; Brace, E.; Milbrandt, J.; Diantonio, A. MAPK signaling promotes axonal degeneration by speeding the turnover of the axonal maintenance factor NMNAT2. eLife 2017, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Geisler, S.; Huang, S.X.; Strickland, A.; Doan, R.A.; Summers, D.W.; Mao, X.; Park, J.; DiAntonio, A.; Milbrandt, J. Gene therapy targeting SARM1 blocks pathological axon degeneration in mice. J. Exp. Med. 2019, 216, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Hirai, S.I.; De, F.C.; Miyata, T.; Ogawa, M.; Kiyonari, H.; Suda, Y.; Aizawa, S.; Banba, Y.; Ohno, S. The c-Jun N-terminal kinase activator dual leucine zipper kinase regulates axon growth and neuronal migration in the developing cerebral cortex. J. Neurosci. 2006, 26, 11992–12002. [Google Scholar] [CrossRef]
- Fan, G.; Merritt, S.E.; Kortenjann, M.; Shaw, P.E.; Holzman, L.B. Dual leucine zipper-bearing kinase (DLK) activates p46(SAPK) and p38(mapk) but not ERK2. J. Biol. Chem. 1996, 271, 24788–24793. [Google Scholar] [CrossRef]
- Shin, J.E.; DiAntonio, A. Highwire Regulates Guidance of Sister Axons in the Drosophila Mushroom Body. J. Neurosci. 2011, 31, 17689–17700. [Google Scholar] [CrossRef]
- Nix, P.; Hisamoto, N.; Matsumoto, K.; Bastiani, M. Axon regeneration requires coordinate activation of p38 and JNK MAPK pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 10738–10743. [Google Scholar] [CrossRef]
- Ghosh, A.S.; Wang, B.; Pozniak, C.D.; Chen, M.; Watts, R.J.; Lewcock, J.W. DLK induces developmental neuronal degeneration via selective regulation of proapoptotic JNK activity. J. Cell Biol. 2011, 194, 751–764. [Google Scholar] [CrossRef]
- Xiong, X.; Collins, C.A. A conditioning lesion protects axons from degeneration via the Wallenda/DLK MAP kinase signaling cascade. J. Neurosci. 2012, 32, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.R.; Press, C.; Daniels, R.W.; Sasaki, Y.; Milbrandt, J.; Diantonio, A. A dual leucine kinase-dependent axon self-destruction program promotes Wallerian degeneration. Nat. Neurosci. 2009, 12, 387–389. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Wu, Z.; Chisholm, A.D.; Jin, Y. The DLK-1 Kinase Promotes mRNA Stability and Local Translation in C. elegans Synapses and Axon Regeneration. Cell 2009, 138, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, A.; Bradke, F. The DLK signalling pathway—A double-edged sword in neural development and regeneration. Embo Rep. 2013, 14, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Watkins, T.A.; Wang, B.; Huntwork-Rodriguez, S.; Yang, J.; Jiang, Z.; Eastham-Anderson, J.; Modrusan, Z.; Kaminker, J.S.; Tessier-Lavigne, M.; Lewcock, J.W. DLK initiates a transcriptional program that couples apoptotic and regenerative responses to axonal injury. Proc. Natl. Acad. Sci. USA 2013, 110, 4039–4044. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.E.; Ha, H.; Kim, Y.K.; Cho, Y.; DiAntonio, A. DLK regulates a distinctive transcriptional regeneration program after peripheral nerve injury. Neurobiol. Dis. 2019, 127, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Le Pichon, C.E.; Meilandt, W.J.; Dominguez, S.; Solanoy, H.; Lin, H.; Ngu, H.; Gogineni, A.; Ghosh, A.S.; Jiang, Z.; Lee, S.H.; et al. Loss of dual leucine zipper kinase signaling is protective in animal models of neurodegenerative disease. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Bloom, A.J.; Miller, B.R.; Sanes, J.R.; DiAntonio, A. The requirement for Phr1 in CNS axon tract formation reveals the corticostriatal boundary as a choice point for cortical axons. Genes Dev. 2007, 21, 2593–2606. [Google Scholar] [CrossRef]
- Chen, X.; Rzhetskaya, M.; Kareva, T.; Bland, R.; During, M.J.; Tank, A.W.; Kholodilov, N.; Burke, R.E. Antiapoptotic and trophic effects of dominant-negative forms of dual leucine zipper kinase in dopamine neurons of the substantia nigra in vivo. J. Neurosci. 2008, 28, 672–680. [Google Scholar] [CrossRef]
- Xiong, X.; Wang, X.; Ewanek, R.; Bhat, P.; DiAntonio, A.; Collins, C.A. Protein turnover of the Wallenda/DLK kinase regulates a retrograde response to axonal injury. J. Cell Biol. 2010, 191, 211–223. [Google Scholar] [CrossRef]
- Shin, J.E.; Miller, B.R.; Babetto, E.; Cho, Y.; Sasaki, Y.; Qayum, S.; Russler, E.V.; Cavalli, V.; Milbrandt, J.; DiAntonio, A. SCG10 is a JNK target in the axonal degeneration pathway. Proc. Natl. Acad. Sci. USA 2012, 109, E3696–E3705. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Raber, J.; Yang, D.; Su, B.; Mucke, L. Dynamic regulation of c-Jun N-terminal kinase activity in mouse brain by environmental stimuli. Proc. Natl. Acad. Sci. USA 1997, 94, 12655–12660. [Google Scholar] [CrossRef] [PubMed]
- Kuan, C.Y.; Whitmarsh, A.J.; Yang, D.D.; Liao, G.; Schloemer, A.J.; Dong, C.; Bao, J.; Banasiak, K.J.; Haddad, G.G.; Flavell, R.A.; et al. A critical role of neural-specific JNK3 for ischemic apoptosis. Proc. Natl. Acad. Sci. USA 2003, 100, 15184–15189. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.A.; Harder, J.M.; Fornarola, L.B.; Freeman, R.S.; Clark, A.F.; Pang, I.H.; John, S.W.M.; Libby, R.T. JNK2 and JNK3 are major regulators of axonal injury-induced retinal ganglion cell death. Neurobiol. Dis. 2012, 46, 393–401. [Google Scholar] [CrossRef]
- Siu, M.; Sengupta Ghosh, A.; Lewcock, J.W. Dual Leucine Zipper Kinase Inhibitors for the Treatment of Neurodegeneration. J. Med. Chem. 2018, 61, 8078–8087. [Google Scholar] [CrossRef]
- von Scheidt, M.; Zhao, Y.; Kurt, Z.; Pan, C.; Zeng, L.; Yang, X.; Schunkert, H.; Lusis, A.J. Applications and Limitations of Mouse Models for Understanding Human Atherosclerosis. Cell Metab. 2017, 25, 248–261. [Google Scholar] [CrossRef]
- Ellenbroek, B.; Youn, J. Rodent models in neuroscience research: Is it a rat race? Dmm Dis. Model. Mech. 2016, 9, 1079–1087. [Google Scholar] [CrossRef]
- Perlman, R.L. Mouse Models of Human Disease: An Evolutionary Perspective. Evol. Med. Public Health 2016, eow014. [Google Scholar] [CrossRef]
- Aldiri, I.; Xu, B.; Wang, L.; Chen, X.; Hiler, D.; Griffiths, L.; Valentine, M.; Shirinifard, A.; Thiagarajan, S.; Sablauer, A.; et al. The Dynamic Epigenetic Landscape of the Retina During Development, Reprogramming, and Tumorigenesis. Neuron 2017, 94, 550–568.e10. [Google Scholar] [CrossRef]
- Gunn, T.M.; Canine, B. Application of Mouse Genetics to Human Disease: Generation and Analysis of Mouse Models, 5th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; ISBN 9780124105294. [Google Scholar]
- Justice, M.J.; Dhillon, P. Using the mouse to model human disease: Increasing validity and reproducibility. Dmm Dis. Model. Mech. 2016, 9, 101–103. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.; Cho, Y. Experimental Model Systems for Understanding Human Axonal Injury Responses. Int. J. Mol. Sci. 2021, 22, 474. https://doi.org/10.3390/ijms22020474
Lee B, Cho Y. Experimental Model Systems for Understanding Human Axonal Injury Responses. International Journal of Molecular Sciences. 2021; 22(2):474. https://doi.org/10.3390/ijms22020474
Chicago/Turabian StyleLee, Bohm, and Yongcheol Cho. 2021. "Experimental Model Systems for Understanding Human Axonal Injury Responses" International Journal of Molecular Sciences 22, no. 2: 474. https://doi.org/10.3390/ijms22020474
APA StyleLee, B., & Cho, Y. (2021). Experimental Model Systems for Understanding Human Axonal Injury Responses. International Journal of Molecular Sciences, 22(2), 474. https://doi.org/10.3390/ijms22020474