
Potential Effects of Sweet Potato (Ipomoea batatas) in Hyperglycemia and Dyslipidemia—A Systematic Review in Diabetic Retinopathy Context

 ,
,  , , ,
, , ,
Abstract
1. Introduction
1.1. Hyperglycemia
1.2. Dyslipidemia
2. Methods
2.1. Search Strategy
2.2. Inclusion Criteria
2.3. Exclusion Criteria
2.4. Data Extraction and Management
2.5. Strategy for Data Extraction
3. Results
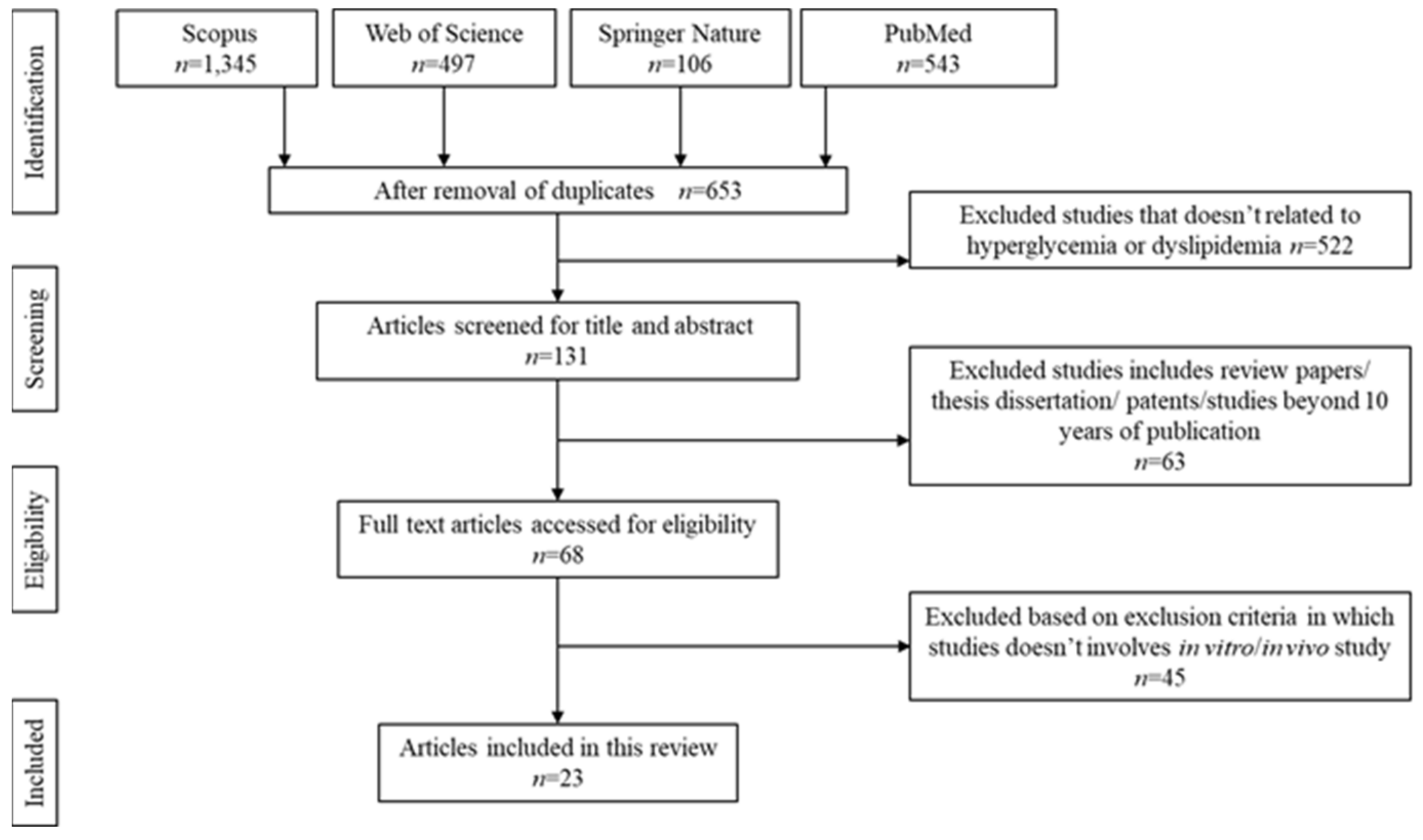
3.1. Literature Search
3.2. The Diabetic Retinopathy as a Consequence of Hyperglycemia
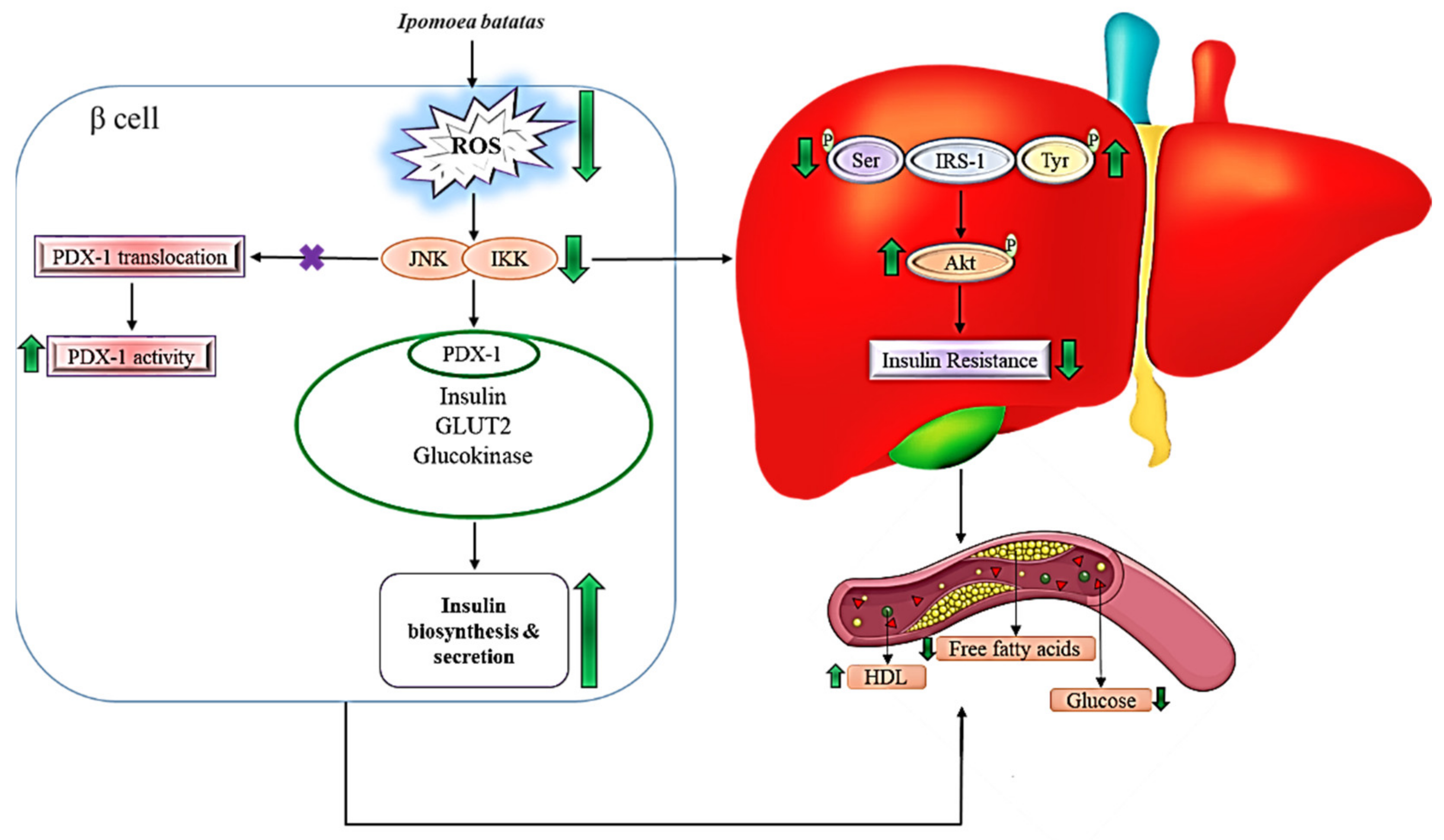
3.3. Mechanism of Ipomoea batatas in Hyperglycemia and Dyslipidemia
3.4. Signaling Mechanism of Ipomoea batatas in Reducing Hyperglycemic Condition
3.5. Signaling Mechanism of Ipomoea batatas in Regulating Dyslipidemia
4. Discussion
5. Safety Concern and Dosage Recommendation
6. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, C.; Liu, D.; Wu, L.; Zhang, J.; Li, X.; Wu, W. Chemical Characterization and Antioxidant Properties of Ethanolic Extract and Its Fractions from Sweet Potato (Ipomoea batatas L.) Leaves. Foods 2019, 9, 15. [Google Scholar] [CrossRef]
- Drapal, M.; Rossel, G.; Heider, B.; Fraser, P.D. Metabolic diversity in sweet potato (Ipomoea batatas, Lam.) leaves and storage roots. Hortic. Res. 2019, 6, 1–9. [Google Scholar] [CrossRef]
- Gupta, S.; Rosenthal, D.M.; Stinchcombe, J.R.; Baucom, R.S. The remarkable morphological diversity of leaf shape in sweet potato (Ipomoea batatas): The influence of genetics, environment, and G×E. New Phytol. 2020, 225, 2183–2195. [Google Scholar] [CrossRef] [PubMed]
- Food & Agriculture Organizations of the United Nations Crops. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 3 April 2021).
- Fu, Z.F.; Tu, Z.C.; Zhang, L.; Wang, H.; Wen, Q.H.; Huang, T. Antioxidant activities and polyphenols of sweet potato (Ipomoea batatas L.) leaves extracted with solvents of various polarities. Food Biosci. 2016, 15, 11–18. [Google Scholar] [CrossRef]
- Meents, A.K.; Chen, S.P.; Reichelt, M.; Lu, H.H.; Bartram, S.; Yeh, K.W.; Mithöfer, A. Volatile DMNT systemically induces jasmonate-independent direct anti-herbivore defense in leaves of sweet potato (Ipomoea batatas) plants. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Qin, L. Growth and photosynthetic characteristics of sweet potato (Ipomoea batatas) leaves grown under natural sunlight with supplemental LED lighting in a tropical greenhouse. J. Plant Physiol. 2020, 252, 153239. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Koh, E. Antioxidant content and activity in leaves and petioles of six sweet potato (Ipomoea batatas L.) and antioxidant properties of blanched leaves. Food Sci. Biotechnol. 2019, 28, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-L.; Lee, S.-L.; Chen, C.-J.; Chen, H.-C.; Kao, M.-C.; Liu, C.-H.; Chen, J.-Y.; Lai, Y.-T.; Wu, Y.-C. Characterization of Secondary Metabolites from Purple Ipomoea batatas Leaves and Their Effects on Glucose Uptake. Molecules 2016, 21, 745. [Google Scholar] [CrossRef]
- Nyirjesy, S.C.; Sheikh, S.; Hadjiliadis, D.; De Leon, D.D.; Peleckis, A.J.; Eiel, J.N.; Kubrak, C.; Stefanovski, D.; Rubenstein, R.C.; Rickels, M.R.; et al. β-Cell secretory defects are present in pancreatic insufficient cystic fibrosis with 1-hour oral glucose tolerance test glucose ≥155 mg/dL. Pediatr. Diabetes 2018, 19, 1173–1182. [Google Scholar] [CrossRef]
- Marcovecchio, M.L. Complications of acute and chronic hyperglycemia. US Endocrinol. 2017, 13, 17–21. [Google Scholar] [CrossRef]
- Duan, W.; Shen, X.; Lei, J.; Xu, Q.; Yu, Y.; Li, R.; Wu, E.; Ma, Q. Hyperglycemia, a neglected factor during cancer progression. Biomed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.-J. Pathogenesis of chronic hyperglycemia: From reductive stress to oxidative stress. J. Diabetes Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Bonora, M.; Patergnani, S.; Rimessi, A.; de Marchi, E.; Suski, J.M.; Bononi, A.; Giorgi, C.; Marchi, S.; Missiroli, S.; Poletti, F.; et al. ATP synthesis and storage. Purinergic Signal. 2012, 8, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.J.; Xiong, S.Q.; Ding, L.X.; Peng, J.; Xia, X.B. Diabetic retinopathy: Focus on NADPH oxidase and its potential as therapeutic target. Eur. J. Pharmacol. 2019, 853, 381–387. [Google Scholar] [CrossRef]
- Kopin, L.; Lowenstein, C.J. Dyslipidemia. Ann. Intern. Med. 2017, 167, ICT82–ICT95. [Google Scholar] [CrossRef] [PubMed]
- Hammer, S.S.; Busik, J.V. The role of dyslipidemia in diabetic retinopathy. Vision Res. 2017, 139, 228–236. [Google Scholar] [CrossRef]
- Patni, N.; Ahmad, Z.; Wilson, D.P. Genetics and Dyslipidemia. In Endocrinology; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Grossman, A., Hershman, J.M., Hofland, J., et al., Eds.; National Center for Biotechnology Information, U.S. National Library of Medicine: Bethesda, MD, USA, 2020; pp. 1–12. [Google Scholar]
- Mancini, G.B.J.; Hegele, R.A.; Leiter, L.A. Dyslipidemia. Can. J. Diabetes 2018, 42, S178–S185. [Google Scholar] [CrossRef]
- Jamal, R.; Zakaria, S.Z.S.; Kamaruddin, M.A.; Jalal, N.A.; Ismail, N.; Kamil, N.M.; Abdullah, N.; Baharudin, N.; Hussin, N.H.; Othman, H.; et al. Cohort Profile: The Malaysian Cohort (TMC) project: A prospective study of non-communicable diseases in a multi-ethnic population. Int. J. Epidemiol. 2014, 44, 423–431. [Google Scholar] [CrossRef]
- Chang, Y.C.; Wu, W.C. Dyslipidemia and diabetic retinopathy. Rev. Diabet. Stud. 2013, 10, 121–132. [Google Scholar] [CrossRef]
- Hutton, B.; Salanti, G.; Caldwell, D.M.; Chaimani, A.; Schmid, C.H.; Cameron, C.; Ioannidis, J.P.A.; Straus, S.; Thorlund, K.; Jansen, J.P.; et al. The PRISMA Extension Statement for Reporting of Systematic Reviews Incorporating Network Meta-analyses of Health Care Interventions: Checklist and Explanations. Ann. Intern. Med. 2015, 162, 777–784. [Google Scholar] [CrossRef]
- Grewal, A.; Kataria, H.; Dhawan, I. Literature search for research planning and identification of research problem. Indian J. Anaesth. 2016, 60, 635–639. [Google Scholar] [CrossRef]
- Niwa, A.; Tajiri, T.; Higashino, H. Ipomoea batatas and Agarics blazei ameliorate diabetic disorders with therapeutic antioxidant potential in streptozotocin-induced diabetic rats. J. Clin. Biochem. Nutr. 2011, 48, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Gautam, S.; Mishra, A.; Maurya, R.; Srivastava, A.K. Antihyperglycemic and Antidyslipidemic Potential of Ipomoea Batatas Leaves in Validated Diabetic Animal Models. Int. J. Pharm. Pharm. Sci. 2015, 7, 176–186. Available online: https://innovareacademics.in/journals/index.php/ijpps/article/view/5603 (accessed on 7 August 2021).
- Lin, K.-H.; Low, P.-Y.; Chao, P.-Y.; Shih, M.-C.; Chiang, M.-C.; Lai, Y.-C.; Wu, S.-B. Antioxidant Properties and Glucose Uptake Effect of Ethanol Extracts from Different Sweet Potato Leaves Prepared by Lyophilization and Oven- Drying at 40 °C. Curr. Nutr. Food Sci. 2017, 13, 227–236. [Google Scholar] [CrossRef]
- Tahir, I.M.; Akhter, N.; Parveen, D.A.; Mehboob, H.; Saleem, S.; Munir, N.; Shah, S.M.-A.; Zaheer, J.; Khan, F.S.; Sultana, S.; et al. Effects of methanolic and aqueous extracts of Ipomoea batatas L on mineral contents level (calcium and magnesium) in alloxan-induced diabetic rats. Pak. J. Pharm. Sci. 2018, 31, 2077–2083. [Google Scholar]
- Akhtar, N.; Akram, M.; Daniyal, M.; Ahmad, S. Evaluation of antidiabetic activity of Ipomoea batatas L. extract in alloxan-induced diabetic rats. Int. J. Immunopathol. Pharmacol. 2018, 32, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kamal, S.; Akhter, N.; Khan, S.G.; Kiran, S.; Farooq, T.; Akram, M.; Shah, S.M.A.; Tahir, I.M.; Akhlaq, M.; Said, F.; et al. Anti-diabetic activity of aqueous extract of Ipomoea batatas L. in alloxan induced diabetic Wistar rats and its effects on biochemical parameters in diabetic rats. Pakistan J. Pharm. Sci. 2018, 31, 1539–1548. [Google Scholar]
- Omodamiro, O.; Omodamiro, R. Evaluation of Hypoglycemic and Hypolipidemic Potentials of Sweet Potato on a Wistar Albino Rat. Am. J. Adv. Drug Deliv. 2018, 6. [Google Scholar] [CrossRef]
- Rafiu, A.; Luka, C. Effects of Aqueous Extract of Ipomoea batatas Leaf on Blood Glucose, Kidney Functions and Hematological Parameters of Streptozotocin-Induced Diabetic Rats. J. Res. Diabetes Metab. 2018, 4, 004–009. [Google Scholar]
- Rafiu, A.A.; Luka, C. Evaluation of the Antidiabetic Property of Aqueous Extract of Ipomoea batatas Leaf on Hyperglycemia, Hyperlipidemia, Blood Electrolytes, and Enzymatic Antioxidants of Streptozotocin-Induced Diabetic Rats. J. Res. Diabetes Metab. 2018, 4, 022–026. [Google Scholar]
- Almoraie, N.M. The Role of Ipomoea Batatas Leaves Extract on the Treatment of Diabetes Induced by Streptozotocin. Pharmacophore 2019, 10, 14–20. [Google Scholar]
- Novrial, D.; Soebowo, S.; Widjojo, P. Protective effect of Ipomoea batatas L leaves extract on histology of pancreatic langerhans islet and beta cell insulin expression of rats induced by streptozotocin. Molekul 2020, 15, 48–55. [Google Scholar] [CrossRef]
- Refaat, W.A.; El-Kholie, E.M.; Abo El-Nasr, A.F. Possible Effects of Sweet Potato Leaves, Roots and their Mixture Feeding on Alloxan-Induced Diabetic Ratsm. J. Home Econ. 2020, 30, 1–18. [Google Scholar]
- Shih, C.K.; Chen, C.M.; Varga, V.; Shih, L.C.; Chen, P.R.; Lo, S.F.; Shyur, L.F.; Li, S.C. White sweet potato ameliorates hyperglycemia and regenerates pancreatic islets in diabetic mice. Food Nutr. Res. 2020, 64, 1–11. [Google Scholar] [CrossRef]
- Jiang, T.; Shuai, X.; Li, J.; Yang, N.; Deng, L.; Li, S.; He, Y.; Guo, H.; Li, Y.; He, J. Protein-Bound Anthocyanin Compounds of Purple Sweet Potato Ameliorate Hyperglycemia by Regulating Hepatic Glucose Metabolism in High-Fat Diet/Streptozotocin-Induced Diabetic Mice. J. Agric. Food Chem. 2020, 68, 1596–1608. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Mu, T.; Sun, H. Sweet potato (Ipomoea batatas L.) leaf polyphenols ameliorate hyperglycemia in type 2 diabetes mellitus mice. Food Funct. 2021. [Google Scholar] [CrossRef]
- Park, J.H.; Choi, S.Y.; Lee, K.W.; Kim, S.S.; Cho, K.D.; Han, C.K. Effect of diets with red yeast sweet potato powder supplement on fecal amount and lipid metabolism in rats fed a high-fat diet. J. Korean Soc. Food Sci. Nutr. 2012, 41, 487–493. [Google Scholar] [CrossRef]
- Jawi, I.M.; Indrayani, A.W.; Sutirta-Yasa, I.W.P. Aqueous Extract of Balinese Purple Sweet Potato (Ipomoea Batatas L.) Prevents Oxidative Stress and Decreases Blood Interleukin-1 in Hypercholesterolemic Rabbits. Bali Med. J. 2015, 4, 37–40. [Google Scholar]
- Kurata, R.; Kobayashi, T.; Ishii, T.; Niimi, H.; Niisaka, S.; Kubo, M.; Kishimoto, M. Influence of sweet potato (Ipomoea batatas L.) leaf consumption on rat lipid metabolism. Food Sci. Technol. Res. 2017, 23, 57–62. [Google Scholar] [CrossRef]
- Nasoetion, M.H.; Atmomarsono, U.; Sunarti, D.; Suthama, N. Growth performance and lipid profile of broilers fed different levels of purple sweet potato extract and raised under different stocking densities. Livest. Res. Rural Dev. 2019, 31, 97. [Google Scholar]
- Nur, A.; Karina Riskawati, Y.; Widodo, E.; Ratnawati, R.; Firdausy, A.; Kusuma Dewi, L.; Karina, M. Effect of Purple Sweet Potato Extract (Ipomoea batatas L) on Zebrafish (Danio rerio) By Diet Induced Obesity. Res. J. Life Sci. 2019, 6, 83–93. [Google Scholar] [CrossRef][Green Version]
- Heriwijaya, I.P.P.D.; Jawi, I.M.; Satriyasa, B.K. Uji efektivitas ekstrak air daun ubi jalar ungu (Ipomoea batatas) terhadap profil lipid tikus putih jantan galur wistar yang diinduksi pakan dislipidemia. Intisari Sains Medis 2020, 11, 452. [Google Scholar] [CrossRef]
- Khairani, A.F.; Islami, U.; Anggun Syamsunarno, M.R.; Lantika, U.A. Synbiotic purple sweet potato yogurt ameliorate lipid metabolism in high fat diet mice model. Biomed. Pharmacol. J. 2020, 13, 175–184. [Google Scholar] [CrossRef]
- Ntchapda, F.; Tchatchouang, F.C.; Miaffo, D.; Maidadi, B.; Vecchio, L.; Talla, R.E.; Bonabe, C.; Seke Etet, P.F.; Dimo, T. Hypolipidemic and anti-atherosclerogenic effects of aqueous extract of Ipomoea batatas leaves in diet-induced hypercholesterolemic rats. J. Integr. Med. 2021, 19, 243–250. [Google Scholar] [CrossRef]
- Safi, S.Z.; Qvist, R.; Kumar, S.; Batumalaie, K.; Ismail, I.S. Bin Molecular mechanisms of diabetic retinopathy, general preventive strategies, and novel therapeutic targets. Biomed Res. Int. 2014, 2014, 801269. [Google Scholar] [CrossRef] [PubMed]
- Bek, T. Diameter Changes of Retinal Vessels in Diabetic Retinopathy. Curr. Diab. Rep. 2017, 17, 82. [Google Scholar] [CrossRef] [PubMed]
- Hammes, H.P.; Lin, J.; Renner, O.; Shani, M.; Lundqvist, A.; Betsholtz, C.; Brownlee, M.; Deutsch, U. Pericytes and the pathogenesis of diabetic retinopathy. Diabetes 2002, 51, 3107–3112. [Google Scholar] [CrossRef]
- Wiley, H.E.; Ferris, F.L. Nonproliferative Diabetic Retinopathy and Diabetic Macular Edema. In Retina; Stephen, J.R., SriniVas, R.S., Wiedemann, P., Wilkinson, C.P., Schachat, A.P., Eds.; Elsevier Inc.: Los Angeles, CA, USA, 2012; Volume 2, pp. 940–968. ISBN 9781455707379. [Google Scholar]
- Beltramo, E.; Porta, M. Pericyte Loss in Diabetic Retinopathy: Mechanisms and Consequences. Curr. Med. Chem. 2013, 20, 3218–3225. [Google Scholar] [CrossRef]
- Penn, J.S.; Madan, A.; Caldwell, R.B.; Bartoli, M.; Caldwell, R.W.; Hartnett, M.E. Vascular endothelial growth factor in eye disease. Prog. Retin. Eye Res. 2008, 27, 331–371. [Google Scholar] [CrossRef]
- Aiello, L.P. The potential role of PKC β in diabetic retinopathy and macular edema. Surv. Ophthalmol. 2002, 47, S263–S269. [Google Scholar] [CrossRef]
- Neve, A.; Cantatore, F.P.; Maruotti, N.; Corrado, A.; Ribatti, D. Extracellular matrix modulates angiogenesis in physiological and pathological conditions. Biomed Res. Int. 2014, 2014, 756078. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef]
- Augustine, J.; Troendle, E.P.; Barabas, P.; McAleese, C.A.; Friedel, T.; Stitt, A.W.; Curtis, T.M. The Role of Lipoxidation in the Pathogenesis of Diabetic Retinopathy. Front. Endocrinol. 2021, 11, 1146. [Google Scholar] [CrossRef]
- Shin, E.S.; Sorenson, C.M.; Sheibani, N. Diabetes and Retinal Vascular Dysfunction. J. Ophthalmic Vis. Res. 2014, 9, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Omar, B.; Ahrén, B. Pleiotropic mechanisms for the glucose-lowering action of DPP-4 inhibitors. Diabetes 2014, 63, 2196–2202. [Google Scholar] [CrossRef] [PubMed]
- Hue, S.-M.; Boyce, A.N.; Somasundram, C. Antioxidant activity, phenolic and flavonoid contents in the leaves of different varieties of sweet potato (Ipomoea batatas). Aust. J. Crop Sci. 2012, 6, 375–380. [Google Scholar]
- Bondeva, T.; Wolf, G. Reactive oxygen species in diabetic nephropathy: Friend or foe? Nephrol. Dial. Transplant. 2014, 29, 1998–2003. [Google Scholar] [CrossRef]
- Soares, J.M.D.; Leal, A.E.B.P.; Silva, J.C.; Almeida, J.R.G.S.; Oliveira, H.P. De Influence of flavonoids on mechanism of modulation of insulin secretion. Pharmacogn. Mag. 2017, 13, 639–646. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef]
- Zeka, K.; Ruparelia, K.; Arroo, R.; Budriesi, R.; Micucci, M. Flavonoids and Their Metabolites: Prevention in Cardiovascular Diseases and Diabetes. Diseases 2017, 5, 19. [Google Scholar] [CrossRef]
- Makino, R.; Takano, K.; Kita, K.; Nishimukai, M. Influence of long-term feeding of high-fat diet on quercetin and fat absorption from the small intestine in lymph duct-cannulated rats. Biosci. Biotechnol. Biochem. 2018, 82, 2007–2011. [Google Scholar] [CrossRef]
- Wei, X.; Gu, N.; Feng, N.; Guo, X.; Ma, X. Inhibition of p38 mitogen-activated protein kinase exerts a hypoglycemic effect by improving β cell function via inhibition of β cell apoptosis in db/db mice. J. Enzyme Inhib. Med. Chem. 2018, 33, 1494–1500. [Google Scholar] [CrossRef]
- Luciani, D.S.; White, S.A.; Widenmaier, S.B.; Saran, V.V.; Taghizadeh, F.; Hu, X.; Allard, M.F.; Johnson, J.D. Bcl-2 and Bcl-xL suppress glucose signaling in pancreatic β-cells. Diabetes 2013, 62, 170–182. [Google Scholar] [CrossRef]
- Stöckli, J.; Fazakerley, D.J.; James, D.E. GLUT4 exocytosis. J. Cell Sci. 2011, 124, 4147–4159. [Google Scholar] [CrossRef]
- Kaneto, H.; Katakami, N.; Matsuhisa, M.; Matsuoka, T.A. Role of reactive oxygen species in the progression of type 2 diabetes and atherosclerosis. Mediators Inflamm. 2010, 2010, 1–11. [Google Scholar] [CrossRef]
- Yang, J.; Wang, C.; Guan, Y. Cytokines in the progression of pancreatic -cell dysfunction. Int. J. Endocrinol. 2010, 2010, 515136–515146. [Google Scholar] [CrossRef]
- Alejandro, E.U.; Kalynyak, T.B.; Taghizadeh, F.; Gwiazda, K.S.; Rawstron, E.K.; Jacob, K.J.; Johnson, J.D. Acute insulin signaling in pancreatic beta-cells is mediated by multiple Raf-1 dependent pathways. Endocrinology 2010, 151, 502–512. [Google Scholar] [CrossRef] [PubMed]
- Ranta, F.; Düfer, M.; Stork, B.; Wesselborg, S.; Drews, G.; Häring, H.U.; Lang, F.; Ullrich, S. Regulation of calcineurin activity in insulin-secreting cells: Stimulation by Hsp90 during glucocorticoid-induced apoptosis. Cell. Signal. 2008, 20, 1780–1786. [Google Scholar] [CrossRef] [PubMed]
- Musilová, J.; Bystrick, J.; Árvay, J.; Harangózo, L. Polyphenols and phenolic acids in sweet potato (Ipomoea batatas L.) roots. Potravin. Slovak J. Food Sci. 2017, 11, 82–87. [Google Scholar] [CrossRef]
- Longo, N.; Frigeni, M.; Pasquali, M. Carnitine transport and fatty acid oxidation. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 2422–2435. [Google Scholar] [CrossRef] [PubMed]
- Dessì, M.; Noce, A.; Bertucci, P.; di Villahermosa, S.M.; Zenobi, R.; Castagnola, V.; Addessi, E.; Di Daniele, N. Atherosclerosis, Dyslipidemia, and Inflammation: The Significant Role of Polyunsaturated Fatty Acids. Int. Sch. Res. Not. 2013, 2013, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Gou, L.; Yan, S.; Huang, T. Inhibition of acetyl-CoA carboxylase by PP-7a exerts beneficial effects on metabolic dysregulation in a mouse model of diet-induced obesity. Exp. Ther. Med. 2020, 20, 521–529. [Google Scholar] [CrossRef]
- Ravaut, G.; Légiot, A.; Bergeron, K.F.; Mounier, C. Monounsaturated fatty acids in obesity-related inflammation. Int. J. Mol. Sci. 2021, 22, 330. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Sasahara, T.; Kiritoshi, S.; Sonoda, K.; Senokuchi, T.; Matsuo, T.; Kukidome, D.; Wake, N.; Matsumura, T.; Miyamura, N.; et al. Evaluation of urinary 8-hydroxydeoxy-guanosine as a novel biomarker of macrovascular complications in type 2 diabetes. Diabetes Care 2003, 26, 1507–1512. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A.; Quagliaro, L.; Catone, B.; Pascon, R.; Piazzola, M.; Bais, B.; Marra, G.; Tonutti, L.; Taboga, C.; Motz, E. Role of hyperglycemia in nitrotyrosine postprandial generation. Diabetes Care 2002, 25, 1439–1443. [Google Scholar] [CrossRef]
- Kumar, S.; Narwal, S.; Kumar, V.; Prakash, O. α-glucosidase inhibitors from plants: A natural approach to treat diabetes. Pharmacogn. Rev. 2011, 5, 19–29. [Google Scholar] [CrossRef]
- Swaminathan, S.; Fonseca, V.A.; Alam, M.G.; Shah, S.V. The role of iron in diabetes and its complications. Diabetes Care 2007, 30, 1926–1933. [Google Scholar] [CrossRef]
- Shaaban, M.; Dawod, A.E.; Nasr, M. Role of iron in diabetes mellitus and its complications. Menoufia Med. J. 2016, 29, 11. [Google Scholar] [CrossRef]
- Bystrom, L.M.; Guzman, M.L.; Rivella, S. Iron and reactive oxygen species: Friends or foes of cancer cells? Antioxidants Redox Signal. 2014, 20, 1917–1924. [Google Scholar] [CrossRef]
- Zhu, J.; Xun, P.; Bae, J.C.; Kim, J.H.; Kim, D.J.; Yang, K.; He, K. Circulating calcium levels and the risk of type 2 diabetes: A systematic review and meta-analysis. Br. J. Nutr. 2019, 122, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Mohanraj, R.; Sivasankar, S. Sweet potato (Ipomoea batatas [L.] Lam)—A valuable medicinal food: A review. J. Med. Food 2014, 17, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Imafidon, K.; Durojaiye, M.; Daniel, B. Safety Assessment of Ipomoea Batatas (L.). Lam Leaf Extract, A Traditional Haematinic; In Male Wistar Rats. J. Ghana Sci. Assoc. 2015, 16, 45–51. [Google Scholar]
- Dutta, S. Sweet Potatoes for Diabetes Mellitus: A Systematic Review. Pharmacophore 2015, 6, 72. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Author | Type of Subject | Dosage of Treatment | Extraction Method | Follow Up Duration | Findings | Conclusion |
|---|---|---|---|---|---|---|
| Niwa et al., 2011 [24] | Male Wistar rats injected with Streptozotocin | 5 g/kg/day for 8 weeks | Not stated | 1st, 3rd, 5th, and 7th week |
| I. batatas exhibits anti-hyperglycemic effects through the p38 MAP kinase signaling pathway. |
| Pal et al., 2015 [25] | Male albino Sprague Dawley rats injected with Streptozotocin | 100 mg/kg/day for 30 days | Ethanol and aqueous | 7th, 14th, 21st, and 28th day |
| Aqueous extraction of I. batatas is an effective anti- hyperglycemic agent. |
| Lin et al., 2017 [26] | CN1927, CYY98 of sweet potato leaf extract (SPLE), and 40 °C oven-dried TN64 | Not relevant | Ethanol | - |
| SPLE improves TNF-α -induced insulin resistance by activating insulin signaling. |
| Tahir et al., 2018 [27] | Male Wistar rats induced with Alloxan monohydrate | 4 g/kg/day for 15 days | Methanol and aqueous | 3rd, 6th, 9th, 12th, and 15th day |
| I. batatas decreases diabetic complications by decreasing calcium and magnesium level in various organs. |
| Akhtar et al., 2018 [28] | Male Wistar rats induced with Alloxan monohydrate | 4 g/kg/day for 14 days | Methanol | 3rd, 6th, 9th, 12th, and 15th day |
| Methanol extraction of I. batatas is an effective anti-hyperglycemic agent. |
| Kamal et al., 2018 [29] | Male Wistar rats induced with Alloxan monohydrate | 4 g/kg/day for 14 days | Aqueous | 3rd, 6th, 9th, 12th, and 15th day |
| Aqueous extraction of I. batatas is an effective anti-hyperglycemic agent in older aged subjects. |
| Omodamiro et al., 2018 [30] | Adult female albino rats | 1000 mg/kg/day or 750 mg/kg/day or 500 mg/kg/day or 250 mg/kg/day for 7 days | Methanol | Every 2 days |
| Methanol extraction of 500 mg/kg is an optimum dose for anti-hyperglycemic. |
| Rafiu et al., 2018 [31] | Adult male Wister strains albino rats induced with Streptozotocin | 400 mg/kg/day for 28 days. | Aqueous | Every day |
| Aqueous extraction of 400 mg/kg is an optimum dose for anti-hyperglycemic. |
| Rafiu et al., 2018 [32] | Adult male Wister strain albino rats induced with Streptozotocin | 400 mg/kg/day for 28 days. | Aqueous | Every 2 days |
| I. batatas is able to reverse hyperglycemic condition. |
| Almoraie, 2019 [33] | Adult male Wister albino rats induced with Streptozotocin | 200 mg/kg/day for 4 weeks | Aqueous | Not stated |
| I. batatas is able to reverse hyperglycemic condition, hyperinsulinemia, oxidative stress, inflammatory and histopathological changes in pancreas. |
| Novrial et al., 2020 [34] | Male Sprague Dawley rats induced with Streptozotocin | 0.25 g/kg/day or 0.8 g/kg/day or 2.5 g/kg/day for 14 days | Ethanol | 6th and 14th day |
| I. batatas is an effective hyperglycemic agent. |
| Refaat et al., 2020 [35] | Adult male albino Sprague Dawley rats induced with Alloxan monohydrate | 2.5% or 5% of diet for 28 days | Powdered form (freeze dried) | 7th, 14th, 21st, and 28th day |
| 5% of grinded I. batatas leaves in diet is an effective anti-hyperglycemic agent. |
| Shih et al., 2020 [36] | Male mice induced with Streptozotocin | 0.5% or 5% of diet for 8 weeks | Powdered form (freeze dried) | Once a week |
| I. batatas exhibits anti- hyperglycemic effects by stimulating the regeneration of pancreatic islet and insulin resistance. |
| Jiang et al., 2020 [37] | Male mice induced with Streptozotocin | 500 mg/kg/day for 8 weeks | Ethanol | Once a week |
| Sweet potato extract stimulates glycolysis and reduces gluconeogenesis. |
| Luo et al., 2021 [38] | Mice | Not specified | Not specified | Not specified |
| I. batatas exhibits anti-hyperglycemic effects through PI3K and glycogen synthase kinase-3β signaling pathway. |
| Author | Type of Subject | Dosage of Treatment | Extraction Method | Follow up Duration | Findings | Conclusion |
|---|---|---|---|---|---|---|
| Park et al., 2012 [39] | Male Sprague Dawley rats fed with high-fat diet (HFD) | 5% of total diet for 4 weeks | Fleshed | 4th week |
| Sweet potato extraction is an effective agent to enhance fecal lipid excretion. |
| Jawi et al., 2015 [40] | Local Balinese male rabbits fed with high cholesterol diet | 4 mL/kg/day for 60 days | Aqueous | 60th day |
| Sweet potato extract is able to reduce cholesterol content. |
| Kurata et al., 2017 [41] | Male Sprague Dawley rats fed with HFD | 0%, 1%, 3%, and 5% of total diet for 35 days | freeze-dried powder | 35th day |
| I. batatas is able to stimulate lipid metabolism. |
| Nasoetion et al., 2019 [42] | Broiler chicks fed with crude protein diet | 25 mL/kg/day and 50 mL/kg/day for 35 days | Aqueous | Weekly for 35 days |
| Sweet potato extract is able to reduce fat deposition. |
| Nur et al., 2019 [43] | Zebrafish (Danio rerio) fed with HFD | 80 ppm, 120 ppm, and 160 ppm for 40 days | Ethanol | Weekly for 40 days |
| Sweet potato extract is able to reduce fat deposition. |
| Heriwijaya et al., 2020 [44] | White male Wister rats fed with high cholesterol diet | 3 cc, 6 cc, and 9cc for 12 weeks | Aqueous | 12th week |
| Purple sweet potato leaf extract is able to reverse hyperlipidemia condition |
| Khairani et al., 2020 [45] | Male Mus Musculus mice fed with HFD | 0.3 mL/day, 0.5 mL/day, and 1.0 mL/day for 9 weeks | Aqueous | 9th week |
| 0.3 mL/20 g/bw is the optimum dosage of the extract to exhibit maximum effect. |
| Ntchapda et al., 2021 [46] | Male Wistar rats fed with HFD | 400 mg/kg, 500 mg/kg, and 600 mg/kg for 4 weeks | Aqueous | 4th week |
| I. batatas is able to reverse hyperlipidemia condition. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naomi, R.; Bahari, H.; Yazid, M.D.; Othman, F.; Zakaria, Z.A.; Hussain, M.K. Potential Effects of Sweet Potato (Ipomoea batatas) in Hyperglycemia and Dyslipidemia—A Systematic Review in Diabetic Retinopathy Context. Int. J. Mol. Sci. 2021, 22, 10816. https://doi.org/10.3390/ijms221910816
Naomi R, Bahari H, Yazid MD, Othman F, Zakaria ZA, Hussain MK. Potential Effects of Sweet Potato (Ipomoea batatas) in Hyperglycemia and Dyslipidemia—A Systematic Review in Diabetic Retinopathy Context. International Journal of Molecular Sciences. 2021; 22(19):10816. https://doi.org/10.3390/ijms221910816
Chicago/Turabian StyleNaomi, Ruth, Hasnah Bahari, Muhammad Dain Yazid, Fezah Othman, Zainul Amiruddin Zakaria, and Mohd Khairi Hussain. 2021. "Potential Effects of Sweet Potato (Ipomoea batatas) in Hyperglycemia and Dyslipidemia—A Systematic Review in Diabetic Retinopathy Context" International Journal of Molecular Sciences 22, no. 19: 10816. https://doi.org/10.3390/ijms221910816
APA StyleNaomi, R., Bahari, H., Yazid, M. D., Othman, F., Zakaria, Z. A., & Hussain, M. K. (2021). Potential Effects of Sweet Potato (Ipomoea batatas) in Hyperglycemia and Dyslipidemia—A Systematic Review in Diabetic Retinopathy Context. International Journal of Molecular Sciences, 22(19), 10816. https://doi.org/10.3390/ijms221910816